

Известия РАН. Серия биологическая, 2020, № 2, стр. 197-204
Оценка состояния среды Йошкар-Олы по морфометрическим показателям Betula pendula Roth
Н. В. Турмухаметова *
Марийский государственный университет
424000 Йошкар-Ола, пл. Ленина, 1, Россия
* E-mail: bonid@mail.ru
Поступила в редакцию 26.08.2019
После доработки 25.10.2019
Принята к публикации 25.10.2019
Аннотация
Приведены результаты многолетних биоиндикационных исследований оценки состояния березы повислой Betula pendula Roth в различных по антропогенной нагрузке условиях Йошкар-Олы. Показано увеличение морфометрических показателей побеговой системы и площади листовых пластинок березы повислой в условиях антропогенного пресса. Отмечено, что флуктуирующая асимметрия (ФА) листа B. pendula с укороченных побегов не зависит от онтогенетического состояния растения, а у листьев с удлиненных побегов зависит. Установлено, что наиболее информативные морфометрические индикационные параметры состояния среды у особей B. pendula – ФА листьев с укороченных побегов и площадь повреждения листовой пластинки.
Проблема загрязнения природной среды, характера и интенсивности ответной реакции биологических объектов на воздействие поллютантов остается актуальной не одно десятилетие. В качестве объектов биомониторинга загрязнений атмосферы наиболее благоприятны растения, поскольку они подвергаются воздействию всего спектра загрязняющих веществ, присутствующих в воздухе. Использование морфометрических показателей чувствительных растительных биоиндикаторов в сочетании с инструментальными методами измерений позволяет получать необходимую информацию об их ответной реакции на воздействие загрязняющих веществ и давать оценку качества среды обитания (Илькун, 1978; Мэннинг, Федер, 1985; Булохов, 1996; Manual…, 1998; Николаевский, 1999; Бухарина и др., 2007; Khondhodjaeva et al., 2018). Зеленые древесные насаждения – важные объекты средообразования городов, наблюдения и контроля, а также индикаторы состояния окружающей среды (Мозолевская, 1998).
Известно, что древесные растения в условиях воздушного загрязнения, как правило, снижают продуктивность, что выражается, в частности, в уменьшении прироста побегов, поэтому данный показатель часто используется для биоиндикации (Wenzel, 1971; Жеребцова, 1976; Илькун, 1978; Athari, Kramer, 1983; Schweingruber et al., 1983; Mandre, Ots, 1995; Николаевский, 1999; Неверова, Колмогорова, 2003; Бухарина и др., 2007; Lakatos, Mirtchev, 2014). Годичный прирост побегов – определяющий итоговый результат всех физиологических процессов, протекающих в организме древесного растения, комплексный диагностический показатель (Жеребцова, 1976). В ходе индивидуального развития процессы роста у растений протекают с разной скоростью: более интенсивный рост характерен для молодых растений, замедленный – для старых (Заугольнова и др., 1988; Чистякова, 1989).
Листья древесных растений – наиболее активная в метаболическом отношении структура. Ассимиляционные органы растения первыми и в наибольшей степени повреждаются токсическими веществами, содержащимися в воздухе, подвергаются атакам насекомых и патогенных микроорганизмов. Экологическая пластичность и восприимчивость листового аппарата к загрязнению воздушного бассейна предоставляют разнообразные возможности для исследований в области фитомониторинга (Илькун, 1978; Захаров и др., 2000; Кавеленова, Леонтьева, 2003). В подобных работах часто используются морфометрические признаки структуры листа (Захаров и др., 2000; Кавеленова, Леонтьева, 2003; Бухарина и др., 2007; Хузина, 2010; Клевцова, 2014; Беланова и др., 2016), анатомические показатели состояния ассимиляционного аппарата (Неверова, 2009; Рамазанова и др., 2011), физиологические показатели (Бухарина и др., 2007; Василенко, Гончарова, 2017).
С точки зрения проблем индикации различных антропогенных воздействий в городах актуален выбор эффективных биообъектов для постоянного всестороннего контроля, а также признаков-маркеров для оценки состояния окружающей природной среды.
Цель исследования – оценка состояния среды на территории Йошкар-Олы по биоиндикационным морфометрическим показателям побеговой системы и листовой пластинки березы повислой Betula pendula Roth.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились в черте Йошкар-Олы – столицы Республики Марий Эл (РМЭ) в 1999–2017 гг. По сравнению с другими городами Йошкар-Ола характеризуется незначительным загрязнением воздушного бассейна. Доля выбросов автотранспорта составляет 70–85%, промышленных предприятий – 15–30% (Государственный…, 1999, 2017).
В Йошкар-Оле участки исследования были выбраны в градиенте загрязнения среды промышленно-транспортными выбросами. Участок 1 – особо охраняемая природная территория (ООПТ) лесопарк “Сосновая роща” – входит в зону наименьшего загрязнения. Участок 2 – микрорайон “Дубки” – относится к зоне слабого загрязнения. Основные загрязнители воздуха на данной территории – ОАО “Стройкерамика” и автотранспорт. В выбросах предприятия присутствуют пыль, диоксид азота и сажа. Выбросы автотранспорта в этой точке незначительны (Государственный…, 1999, 2017). Участок 3 – Парк культуры и отдыха им. 30-летия ВЛКСМ – относится к зоне умеренного загрязнения. Парк со всех сторон окружен автодорогами. Участок 4 – окрестности фармацевтического завода “Марбиофарм” – также принадлежит к зоне умеренного загрязнения. Завод – одно из предприятий, вносящих наибольший вклад в загрязнение атмосферного воздуха города. В процессе работы предприятия выбрасываются такие вещества, как хлористый водород, ацетон, диоксид серы, диоксид азота (Государственный…, 1999, 2017). В 2016 г. исследование проводили также в условиях естественных лесных массивов двух районов РМЭ.
Материал собирали с деревьев B. pendula трех онтогенетических состояний генеративного периода: молодого (g1), средневозрастного (g2) и старого (g3) (Чистякова, 1989). Для изучения побеговой системы из нижнего яруса кроны 10 модельных деревьев B. pendula разных онтогенетических состояний с южной или юго-западной его сторон брали по 10 ветвей. Главную ось рассматривали как ось (n + 1)-го порядка, ее боковые ветви – как оси (n + 2)-го порядка, идущие затем – как оси (n + 3)-го порядка и т.д. Каждая ветвь была охарактеризована следующими признаками: длиной годичного удлиненного побега (n + 3)-го порядка, числом метамеров, числом укороченных побегов (n + 4)-го порядка, числом листьев на приросте текущего года, числом удлиненных побегов (n + 4)-го порядка. За 1999–2002 гг. были исследованы 4800 годичных побегов B. pendula.
Изучение флуктуирующей асимметрии (ФА) листовой пластинки деревьев проводили по стандартной методике (Захаров и др., 2000). Однако на первоначальном этапе работы в 2000, 2001 гг. анализировали листья не только с укороченных, но и с удлиненных побегов B. pendula. Листья собирали после остановки их роста (начиная с июля); брали по 10 листьев с укороченных побегов прошлого года и с удлиненных побегов текущего года (изучали третий лист) с 10 деревьев B. pendula разных онтогенетических состояний. Выбор третьего листа был обусловлен незначительной модификационной изменчивостью первых трех листьев однолетнего удлиненного побега (Джумабаева, 1978). На гербаризированных листьях изучали по пять признаков с левой и правой половинок листа. Всего было измерено 5400 листьев.
ФА определенного признака листа вычисляли как модуль разности значений признака слева и справа, отнесенный к сумме этих значений. Далее ФА листа оценивали с помощью интегрального показателя стабильности развития – относительного различия между сторонами листа, осредненного по совокупности признаков, ФА всех листьев дерева – как среднее арифметическое значений асимметрии всех измеренных листьев с этого дерева. Степень нарушения стабильности развития дерева определяли с использованием пятибалльной шкалы (Захаров и др., 2000).
На отдельно собранных листьях оценивали площадь листовых пластинок с использованием переводного коэффициента для B. pendula (Федорова, Никольская, 2003). У каждого листа определяли длину, ширину листовой пластинки, площадь листовой пластинки как произведение двух вышеуказанных показателей и переводного коэффициента 0.64. С помощью миллиметровой бумаги определяли площадь повреждений листовых пластинок. Было измерено 300 листьев.
Статистическая обработка данных проводилась в программе Statistica 10 с использованием двух- и трехфакторного дисперсионного анализа, Шеффе-теста, коэффициента корреляции Спирмена (Sokal, Rohlf, 1995).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Ветви B. pendula – симподиальная система, слагающаяся из удлиненных годичных побегов. Изучали малолетнюю разветвленную систему побегов (n + 3)-го порядка B. pendula. В вегетативных почках удлиненного побега (n + 3)-го порядка закладываются удлиненные побеги следующего (n + 4)-го порядка, в смешанных почках – укороченные побеги, несущие, как правило, по два листа и женское соцветие. Укороченные побеги нарастают моноподиально несколько лет, некоторые из них могут стать удлиненными (Серебряков, 1952). Мы попытались выяснить, насколько изменчива побеговая система разновозрастных особей березы повислой генеративного периода в условиях разной степени антропогенной нагрузки.
Двухфакторный дисперсионный анализ выявил различия (р < 0.01–0.05) значений прироста побегов (n + 3)-го порядка в 1999–2002 гг. как у разных онтогенетических групп B. pendula, так и у деревьев в различных условиях произрастания. В относительно чистых условиях произрастания (участок 1) у разновозрастных генеративных особей B. pendula различия длин побегов (n + 3)-го порядка были отмечены только в 1999 г., в условиях среднего загрязнения среды (участок 4) ежегодно у g3-деревьев прирост побегов был выше по сравнению с таковым у других онтогенетических групп (рис. 1), число удлиненных побегов (n + 4)-го порядка также увеличилось. Однако число структурных элементов на побегах (n + 3)-го порядка (метамеров на годичном побеге), а также число укороченных побегов (n + 4)-го порядка не различаются у разновозрастных особей B. pendula (р > 0.05). Следовательно, интенсивный рост побегов g3-особей B. pendula в неблагоприятных условиях определяется более значительным растяжением междоузлий в процессе видимого роста побега. В условиях загрязнения среды промышленно-транспортными выбросами у разновозрастных деревьев B. pendula активизируются ростовые процессы. Так, например, в 2002 г. прирост побегов деревьев в зоне умеренного загрязнения среды был в 1.31–1.47 раза выше такового у контрольных деревьев (p < 0.05).
Рис. 1.
Средняя длина побега (n + 3)-го порядка особей Betula pendula разных онтогенетических групп в 1999 г. (а) и 2002 г. (б). * – р < 0.05, ** – р < 0.01.
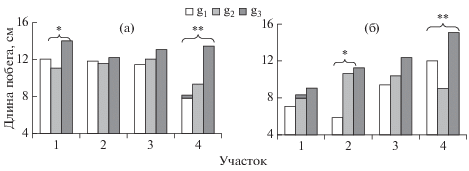
Увеличение прироста побегов у разновозрастных особей B. pendula в условиях воздушного загрязнения среды, по-видимому, обусловлено как повышенными температурами и содержанием углекислого газа в воздухе, так и использованием загрязняющих веществ как дополнительных элементов минерального питания (Илькун, 1978; Бухарина и др., 2007).
Далее была предпринята попытка охарактеризовать состояние окружающей среды в Йошкар-Оле в относительно чистых и антропогенно нарушенных местообитаниях с помощью оценки ФА у листьев B. pendula разных онтогенетических состояний. ФА – незначительные ненаправленные различия между правой и левой сторонами различных морфологических структур, в норме обладающих билатеральной симметрией (Захаров и др., 2000). Выбор листа B. pendula как показателя здоровья среды достаточно эффективен, что подтверждается многочисленными исследованиями в данном направлении (Захаров и др., 2000; Гелашвили и др., 2001; Стрельцов, 2003; Захаров, 2014; Шадрина, Вольперт, 2018). Листья с удлиненных побегов не используются, так как у них выявляется больше нарушений в программе развития (Захаров и др., 2000). Мы полагаем, что укороченные побеги могут подвергаться негативному влиянию факторов окружающей среды в течение нескольких лет, а удлиненные однолетние побеги чувствуют и испытывают краткосрочное воздействие, и поэтому их можно использовать для подобного изучения особенностей развития в текущем вегетационном периоде.
Считается, что уровень “ошибок” в ходе развития организма (онтогенеза или морфогенеза) в нормальных условиях минимален и возрастает при любом стрессирующем воздействии, что и приводит к увеличению асимметрии.
Проведенный двухфакторный дисперсионный анализ выявил зависимость ФА листьев с удлиненных побегов от возрастных особенностей деревьев B. pendula только в зоне умеренного загрязнения (p < 0.01–0.05). Изучаемый показатель у g1-особей двух участков наиболее загрязненной зоны на один балл выше (пятый балл – критическое состояние организма), у g3-особей – на один балл ниже (второй и третий баллы – незначительные и средние нарушения в развитии), чем уровень ФА листьев с укороченных побегов (табл. 1). На остальных участках балльные оценки отклонения состояния листьев B. pendula от нормы по рассматриваемому показателю не различаются при использовании листьев с побегов разного типа.
Таблица 1.
Средние значения показателя флуктуирующей асимметрии листьев с удлиненных побегов у особей Betula pendula в 2000 г.
| Участок | g1-деревья | g2-деревья | g3-деревья |
|---|---|---|---|
| 1 | 0.039 ± 0.002 (I) | 0.039 ± 0.001 (I) | 0.037 ± 0.003 (I) |
| 2 | 0.039 ± 0.003 (I) | 0.037 ± 0.003 (I) | 0.035 ± 0.002 (I) |
| 3 | 0.050 ± 0.0021 (IV)* | 0.049 ± 0.004 (III)* | 0.041 ± 0.002 (II)* |
| 4 | 0.055 ± 0.002 (V)** | 0.048 ± 0.003 (III) | 0.045 ± 0.003 (III)** |
Примечание. * – p < 0.05, ** – p < 0.01. I–V – баллы (Захаров и др., 2000); для табл. 1 и 2.
Следовательно, однолетние побеги у g1-особей B. pendula в условиях среднего загрязнения среды более чувствительны к загрязнению, а у g3-особей менее чувствительны (Турмухаметова, Шивцова, 2007). Это, возможно, свидетельствует о разной устойчивости зачатков листьев в материнской почке, закладывающихся в год, предшествующий сбору материала, к загрязнению на первом и третьем этапах генеративного периода B. pendula.
В результате исследования установлено, что значение интегрального показателя стабильности развития листа B. pendula с укороченных побегов не зависит от онтогенетического состояния растения (р > 0.05). Морфогенез листа с укороченного типа побега у разновозрастных особей B. pendula генеративного периода онтогенеза достаточно стабилен. Следовательно, для проведения подобных работ можно собирать листья с укороченных побегов березы повислой, не учитывая возраст генеративных деревьев. Ранее (Амосова, Феклистов, 2010) была изучена ФА листа с укороченных побегов B. pendula в условиях Архангельска и было выявлено отсутствие различий по изучаемому признаку у прегенеративных и генеративных особей.
В 2016 г. было продолжено мониторинговое исследование с использованием ФА листовой пластинки березы повислой. Анализировали листья, собранные только с укороченных побегов g2-деревьев (участки 1–4). Была выявлена зависимость возрастания ФА листьев с укороченных побегов B. pendula с повышением степени антропогенного пресса (табл. 2, 3). По пятибалльной шкале стабильности развития (Захаров и др., 2000) состояние деревьев B. pendula на участке 1 соответствует условной норме (первый балл), на участке 2 – условной норме (первый балл; 2000 и 2001 гг.) и незначительным нарушениям в развитии (второй балл; 2016 г.), на участке 3 – средним нарушениям в развитии (третий балл), на участке 4 – средним (третий балл; 2001 г.) и существенным нарушениям в развитии (четвертый балл; 2000 и 2016 гг.). Следовательно, неблагоприятные условия в зоне умеренного загрязнения среды отразились на морфоструктуре особей в фрагментах популяции B. pendula, в которых преобладают деревья с высокой ФА листа. Данная тенденция сохранилась в разные годы исследования у замаркированных деревьев в Йошкар-Оле (p < 0.001) (Турмухаметова, Сухорукова, 2017).
Таблица 2.
Средние значения показателя флуктуирующей асимметрии листьев с укороченных побегов у g2-особей Betula pendula в разные годы исследования
| Участок | 2000 г. | 2001 г. | 2016 г. |
|---|---|---|---|
| 1 | 0.036 ± 0.001 (I) | 0.036 ± 0.001 (I) | 0.039 ± 0.004 (I) |
| 2 | 0.039 ± 0.001 (I) | 0.037 ± 0.003 (I) | 0.041 ± 0.002 (II) |
| 3 | 0.046 ± 0.002 (III) | 0.048 ± 0.003 (III) | 0.047 ± 0.002 (III) |
| 4 | 0.050 ± 0.001 (IV) | 0.048 ± 0.002 (III) | 0.053 ± 0.002 (IV) |
| 5 | – | – | 0.034 ± 0.002 (I) |
| 6 | – | – | 0.054 ± 0.003 (IV) |
Таблица 3.
Результаты дисперсионного анализа показателя флуктуирующей асимметрии листа с укороченных побегов Betula pendula в разные годы исследования
| Исследуемый фактор | Число степеней свободы | Средний квадрат эффекта | F-критерий | p |
|---|---|---|---|---|
| Местообитание | 3 | 0.0098 | 19.353 | 10–7 |
| Год | 2 | 0.001 | 2.061 | 0.127 |
| Дерево | 9 | 0.0005 | 1.090 | 0.367 |
| Взаимодействие | 54 | 0.00032 | 0.638 | 0.98 |
| Ошибка | 1080 | 0.00051 |
За 15 лет характеристика качества среды по величине интегрального показателя стабильности развития березы повислой сохранилась на прежнем уровне, что доказывают результаты трехфакторного дисперсионного анализа: отсутствие статистически значимого влияния фактора “год” на показатель ФА (табл. 3) (р > 0.05).
Кроме того, анализировали материал, собранный в 2016 г. в березняках в двух районах РМЭ, в условиях отсутствия явного антропогенного воздействия. На участке 5 (Юринский район, д. Подлесная) ФА = 0.034, что соответствует первому баллу по шкале оценки отклонений (Захаров и др., 2000), на участке 6 (Звениговский р-н, пос. Суслонгер) ФА = 0.054, что соответствует четвертому баллу. Это свидетельствует о нормальном состоянии или о существенном нарушении развития особей. На участке регулярно сливали мазут, что и могло вызвать нарушения стабильности развития листовых пластинок B. pendula.
Значения интегрального показателя стабильности развития листьев B. pendula в разных районах Йошкар-Олы согласуются с зонированием города по уровню загрязнения на основании данных других биоиндикационных исследований (Экология…, 2007), а также с данными по химическому анализу атмосферного воздуха (Государственный…, 1999, 2017).
С помощью морфологического анализа листьев были выявлены как точечные некротические повреждения, хлорозы, так и разнообразные повреждения насекомыми. Было установлено, что площадь повреждений листовой поверхности B. pendula в окрестностях Йошкар-Олы очень небольшая (0–5%) (Красинский, 1950), однако значение этого показателя возрастает с увеличением интенсивности антропогенной нагрузки. Кроме того, в условиях загрязнения у B. pendula уменьшаются некоторые показатели анатомической структуры листа: толщина листовой пластинки, высота верхнего и нижнего эпидермиса, высота столбчатой и губчатой паренхимы (р < 0.05), причем в большей степени поражаются клетки нижнего эпидермиса листьев. Данные негативные процессы компенсируются сохранением относительно большой площади листовой пластинки, что, возможно, определяет адаптацию B. pendula к загрязнению среды промышленно-транспортными выбросами (Турмухаметова, 2005).
Дисперсионный анализ площадей листовых пластинок березы повислой, собранных в 2017 г., и их повреждений выявил увеличение этих показателей в градиенте загрязнения среды (рис. 2) (p < 0.01). Подобная закономерность была отмечена и другими исследователями (Кавеленова, Леонтьева, 2003; Бухарина и др., 2007; Хузина, 2010; Клевцова, 2014).
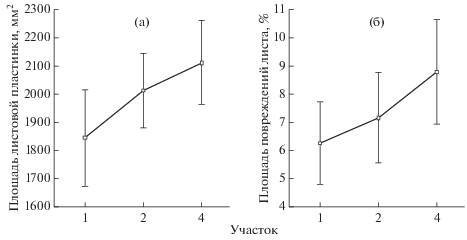
Можно предположить, что отдельные ингредиенты, содержащиеся в воздухе, оказали стимулирующее влияние на рост листовой пластинки. Увеличение площади поврежденной ткани листьев в градиенте загрязнения среды может быть связано с тем, что в условиях антропогенного пресса деревья становятся ослабленными и чаще подвергаются нападениям членистоногих фитофагов. Данная тенденция была отмечена во время исследования структуры энтомокомплексов березы посивлой в условиях Йошкар-Олы (Турмухаметова, 2017): с увеличением степени антропогеннй нагрузки уменьшается видовое разнообразие членистоногих филлофагов березы повислой в 1.5–2.8 раза, однако при этом относительное обилие отдельных наиболее массовых вредителей листьев (тлей, галлиц и клещей) возрастает.
Далее был проведен анализ статистической взаимосвязи показателя стабильности развития B. pendula и числа видов дендробионтов, обитающих в ее кроне. Была выявлена обратная корреляционная связь (rs = –1, p < 0.05). Возрастание интегрального показателя стабильности развития с увеличением антропогенной нагрузки было сопряжено с уменьшением разнообразия насекомых-дендробионтов разных групп от филлофагов и энтомофагов до пантофагов. В условиях среднего загрязнения среды промышленно-транспортными выбросами повышалось относительное обилие членистоногих, ведущих скрытый или полускрытый образ жизни и формирующих защитные приспособления (галлы, внутренние полости и скручивание листьев). Число насекомых-листогрызов, характерных для естественных лесных массивов, в районах с промышленными и транспортными выбросами сокращается, что может быть обусловлено как островной пространственной структурой городских зеленых насаждений, так и прямым воздействием на насекомых поллютантов, а также ухудшением качества кормового ресурса (Турмухаметова, 2017).
ЗАКЛЮЧЕНИЕ
Деревья B. pendula в условиях Йошкар-Олы испытывают комплексное неблагоприятное воздействие специфического микроклимата городской среды, загрязнения атмосферы и почвы промышленно-транспортными выбросами. У старых генеративных деревьев B. pendula в техногенной среде размерная поливариантность побегов проявляется в увеличении прироста побегов и числа побегов следующего порядка без изменения числа метамеров. Специфический микроклимат и загрязнение городской среды способствуют активации ростовых процессов побегов и листовых пластинок особей B. pendula. Применение морфометрических показателей побеговой системы и площади листовых пластинок березы повислой в качестве биоиндикационных может привести к неверным выводам.
В условиях умеренного загрязнения среды у разновозрастных деревьев происходят разнонаправленные изменения в развитии листьев с укороченных и удлиненных побегов, не соответствующие норме. ФА листьев у однолетних удлиненных побегов B. pendula позволяет выявить степень чувствительности разновозрастных особей к загрязнению среды. У генеративных деревьев B. pendula отмечается большее постоянство морфообразовательных процессов листьев именно с укороченных побегов. Так как ФА листьев с укороченных побегов не зависит от онтогенетического состояния особи, то этот показатель более универсален.
Наиболее информативны такие морфометрические индикационные показатели состояния среды у особей B. pendula, как ФА листьев с укороченных побегов и площадь повреждения листовой пластинки. Полученные результаты свидетельствуют о неблагоприятности среды обитания в окрестностях фармацевтического завода в Йошкар-Оле. Использование значений ФА листовой пластинки B. pendula как характеристики интегрального показателя стабильности развития позволило оценить состояние среды в некоторых местообитаниях на территории Йошкар-Олы. Проведенное биоиндикационное исследование согласуется с зонированием городской территории по данным химического и биологического мониторингов.
Список литературы
Амосова И.Б., Феклистов П.А. Асимметрия листовой пластинки березы повислой у особей разного возрастного состояния в пригородных лесах г. Архангельска // Изв. вузов. Лес. журн. 2010. № 2. С. 60–66.
Беланова А.П., Банаев Е.В., Томошевич М.А., Чиндяева Л.Н. Состояние древесных растений в разных экологических зонах сибирского города // Изв. СамарНЦ РАН. 2016. Т. 18. № 2 (2). С. 292–296.
Булохов А.Д. Экологическая оценка среды методами фитоиндикации: монография. Брянск: Изд-во БГПУ, 1996. 104 с.
Бухарина И.Л., Поварницина Т.М., Ведерников К.Е. Эколого-биологические особенности древесных растений в урбанизированной среде: монография. Ижевск: ИжевГСХА, 2007. 216 с.
Василенко М.И., Гончарова Е.Н. Биологические методы в оценке качества городской среды // Управление городом: теория и практика. 2017. № 3(26). С. 26–33.
Гелашвили Д.Б., Краснов А.Н., Логинов В.В., Мокров И.В., Радаев А.А., Силкин А.А., Слепов А.В., Чупрунов Е.В. Методологические и методические аспекты мониторинга здоровья среды. Здоровье среды Керженского заповедника // Тр. Гос. природного заповедника “Керженский”. Н. Новгород: ННГУ, 2001. С. 287–324.
Государственный доклад о состоянии окружающей природной среды Республики Марий Эл в 1998 г. Йошкар-Ола: М-во экологии и природопользования РМЭ, 1999. 190 с.
Государственный доклад о состоянии санитарно-эпидемиологического благополучия населения Республики Марий Эл в 2016 г. Йошкар-Ола: Управление Федеральной службы по надзору в сфере защиты прав потребителей и благополучия человека по РМЭ, 2017. 207 с.
Джумабаева Ш.Б. Ритм сезонного развития, морфологическое и анатомическое строение органов побега некоторых видов древесных растений, используемых для озеленения г. Караганды: Автореф. дис. канд. биол. наук. Л.: ЛГУ, 1978. 17 с.
Жеребцова Г.П. Изменение жизнеспособности древесных растений в условиях городской среды: Автореф. дис. канд. биол. наук. М.: Моск. лесотехн. ин-т, 1976. 31 с.
Захаров В.М. Оценка состояния биоразнообразия и здоровья среды // Поволж. экол. журн. 2014. № 1. С. 50–59.
Захаров В.М., Баранов А.С., Борисов В.И., Валецкий А.В., Кряжева Н.Г., Чистякова Е.К., Чубинишвили А.Т. Здоровье среды: методика оценки. М.: ЦЭПР, 2000. 66 с.
Заугольнова Л.Б., Жукова Л.А., Комаров А.С., Смирнова О.В. Ценопопуляции растений (очерки популяционной биологии). М.: Наука, 1988. 184 с.
Илькун Г.М. Загрязнители атмосферы и растения. Киев: Наук. думка, 1978. 246 с.
Кавеленова Л.М., Леонтьева М.В. К возможности оценки экофизиологических параметров высших растений в условиях ООПТ // Экологические проблемы заповедных территорий России / Под ред. Саксонова С.В. Тольятти: ИЭВБ РАН, 2003. С. 92–96.
Клевцова М.А. Фитоиндикационная оценка состояния заповедных и урбанизированных территорий (на примере Воронежской области) // Вестн. Тамбов. ун-та. Естеств. и техн. науки. 2014. Т. 19. Вып. 5. С. 1301–1303.
Красинский Н.П. Теоретические основы построения ассортиментов газоустойчивых растений // Дымоустойчивость растений и дымоустойчивые ассортименты. Горький: Горьк. гос. ун-т, 1950. С. 111–171.
Мозолевская Е.Г. Концепция мониторинга состояния зеленых насаждений и городских лесов г. Москвы // Вестн. МГУЛ – Лес. вестник. 1998. № 2. С. 5–13.
Мэннинг У.Д., Федер У.А. Биомониторинг загрязнения атмосферы с помощью растений. Л.: Гидрометеоиздат, 1985. 143 с.
Неверова О.А. Применение фитоиндикации в оценке загрязнения окружающей среды // Биосфера. 2009. № 1(1). С. 82–92.
Неверова О.А., Колмогорова Е.Ю. Древесные растения и урбанизированная среда. Экологические и биотехнологические аспекты. Новосибирск: Наука, 2003. 222 с.
Николаевский В.С. Экологическая оценка загрязнения среды и состояния наземных экосистем методами фитоиндикации. М.: МГУЛ, 1999. 193 с.
Рамазанова З.Р., Асадулаев З.М., Магомедова Б.М. Адаптивные особенности анатомической структуры листьев Platanus orientalis L., Quercus robus L., Ulmus pumila L. в условиях Махачкалы // Изв. ДагГПУ. Естеств. и точн. науки. 2011. № 3. С. 54–59.
Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Сов. наука, 1952. 391 с.
Стрельцов А.Б. Региональная система биологического мониторинга. Калуга: Изд-во Калуж. ЦНТИ, 2003. 158 с.
Турмухаметова Н.В. Особенности морфогенеза побегов и феноритмов Betula pendula Roth. и Tilia cordata Mill. в условиях городской среды: Автореф. дис. канд. биол. наук. Новосибирск: ЦСБС СО РАН, 2005. 19 с.
Турмухаметова Н.В. Оценка состояния лиственных деревьев и состава филлофагов в условиях г. Йошкар-Олы // Самар. науч. вестн. 2017. Т. 6. № 4(21). С. 80–84.
Турмухаметова Н.В., Сухорукова М.В. К изучению флуктуирующей асимметрии листа Betula pendula Roth // Проблемы популяционной биологии: Матер. XII Всерос. популяционного семинара памяти Николая Васильевича Глотова (1939–2016), Йошкар-Ола, 11–14 апреля 2017 г. Йошкар-Ола: Стринг, 2017. С. 234–235.
Турмухаметова Н.В., Шивцова И.В. Морфологический подход к оценке состояния среды по асимметрии листа Betula pendula Roth и Fragaria vesca L. // Вестн. МГУЛ – Лес. вестн. 2007. № 5. С. 140–143.
Федорова А.И., Никольская А.Н. Практикум по экологии и охране окружающей среды: Уч. пособие. М.: Владос, 2003. 288 с.
Хузина Г.Р. Влияние урбаносреды на морфометрические показатели листа березы повислой (Betula pendula Roth) // Вестн. Удмурт. ун-та. 2010. Вып. 3. С. 53–57.
Чистякова А.А. Betula pendula Roth – береза повислая // Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники. М.: МГПИ, 1989. С. 42–52.
Шадрина Е.Г., Вольперт Я.Л. Практика оценки здоровья среды: эффективность применения показателя флуктуирующей асимметрии и других биоиндикационных подходов // Жизнь Земли. 2018. № 40(2). С. 183–198.
Экология города Йошкар-Олы: науч. издание / Отв. ред. Воскресенская О.Л. Йошкар-Ола: Мар. гос. ун-т, 2007. 300 с.
Athari S., Kramer H. Zur Problematik der Erfassung von umweltbedingten Zuwachsverbesten in Fichtenbestunden // Forst Holzwirt. 1983. Bd 38. № 8. S. 204–206.
Khondhodjaeva N.B., Ismillaeva K.B., Ruzimbayeva N.T. Bioindication and its importance in the conducting of ecological monitoring // Europ. Sci. 2018. № 4(36). P. 68–70.
Lakatos F., Mirtchev S. Manual for visual assessment of forest crown condition. Pristina: Food and Agriculture Organization of the United Nations, 2014. 23 p.
Mandre M., Ots K. The height growth of trees // Dust Pollution and Forest Ecosystems. Tallin: Inst. Ecol., 1995. № 3. P. 117–118.
Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests. Hamburg; Prague: United Nations Environment Programme and Economic Commission for Europe, 1998. 177 p.
Schweingruber F.H., Kontic R., Winkler-Seifern A. Eine jahrringanalytische Studie zum Nadelbaumster in der Schweiz // Ber. Eidgenoss. Anst. Forstl. Versuch. 1983. № 253. 29 s.
Sokal R.R., Rohlf F.J. Biometry. N.Y.: Freeman, 1995. 887 p.
Wenzel K.F. Habitus-Anderung des Waldbaume durch Luftverunreinigung // Forstarchiv. 1971. Bd 42. № 8–9. S. 165–172.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая