

Известия РАН. Серия биологическая, 2020, № 2, стр. 124-136
Экогеографические единицы и охрана внутривидового разнообразия
Л. А. Животовский 1, *, Г. О. Османова 2
1 Институт общей генетики им. Н.И. Вавилова РАН
ул. Губкина, 3, Москва, 119991 Россия
2 Марийский государственный университет, пл. Ленина, 1, Йошкар-Ола, 424000 Республика Марий Эл, Россия
* E-mail: levazh@gmail.com
Поступила в редакцию 26.08.2019
После доработки 25.10.2019
Принята к публикации 25.10.2019
Аннотация
Обсуждается процедура разбиения вида на экогеографические единицы (ЭГЕ) и экогеографические агрегации (ЭГА), выделяемые по данным о градиентах среды обитания и миграционных особенностях (генных потоках). Отмечено, что каждую ЭГЕ и ЭГА можно рассматривать как единицу запаса, воспроизводства, промысла, охраны.
Согласно Эрнсту Майру (Майр, 1968), популяционная структура вида организована в виде географических клин, изолятов, зон интерградации популяций, возможные пути становления которых подробно обсуждались с позиций основных микроэволюционных факторов (Шварц, 1967; Тимофеев-Ресовский и др., 1973; Lewontin, 1974; Алтухов, 1983; Яблоков, 1987; Животовский, 2016, 2017; и др.).
Основные факторы и процессы становления отдельной популяции и популяционной структуры вида в целом – это условия среды обитания, задающие направление адаптивных изменений; преобразования популяционной изменчивости по генам, которые контролируют морфологические, физиологические и поведенческие признаки, обеспечивающие соответствующие адаптации (Moritz, 2002; и др.).
Ранее (Zhivotovsky et al., 2015; Животовский, 2016, 2017; Животовский, Османова, 2018, 2019а–в) было предложено использовать в целях управления природными биологическими ресурсами (промысла, воспроизводства, охраны) экогеографические единицы (ЭГЕ, или EGU – ecogeographic unit), каждая из которых представляет собой природное тело, скомпонованное из эколого-географических характеристик ареала и эколого-биологических признаков этих популяций.
Цель работы – обзор нашего подхода к выделению внутривидовых ЭГЕ и основанных на них экогеографических агрегаций (ЭГА) совокупности видов.
РАЙОНИРОВАНИЕ ПОПУЛЯЦИЙ – ОСНОВА УПРАВЛЕНИЯ ПРИРОДНЫМИ БИОРЕСУРСАМИ
Управлять природными биологическими ресурсами – значит создать дорожную карту воспроизводства, охраны и промысла популяций тех видов, которые определяют облик экосистемы в данном месте (Животовский, Османова, 2019а). Для этого надо в первую очередь очертить ареал этих популяций, указать их естественные географические границы, т.е. районировать популяции.
Границы популяций определяются двумя основными факторами: условиями среды и пределами генных потоков. Природная популяция (многие поколения, обитающие в одном месте), адаптируется к его условиям и благодаря внутренним генным потокам формирует свои уникальные особенности, общие для особей этой популяции.
Условия среды – главный, определяющий фактор в жизни популяции. Ее фенотипический облик и генетический профиль – эволюционные производные этих условий. Изменение среды обитания – важнейшая причина исчезновения популяций и видов. Сегодня губительные для большинства видов изменения среды во многом обусловлены хозяйственной деятельностью человека (распашка земель, выпас скота, осушение болот, ведение строительных работ, интродукция видов и др.), а также браконьерством и бесконтрольным использованием ресурсных видов. Некоторые популяции, находящиеся на краю ареала и имеющие пониженные адаптационные возможности, гибнут, будучи не в состоянии приспособиться к меняющимся условиям среды и не выдержав конкуренции со стороны других видов. Поэтому важно найти подходы к районированию популяций.
Как сохранять популяции? Были предложены две концепции сохранения природного популяционного разнообразия (Waples, 1991; Moritz, 2002; Allendorf et al., 2012; Funk et al., 2012; и др.): сохранять существующее сейчас генетическое разнообразие, которое обеспечивает адаптацию популяций к условиям их среды обитания; сохранять условия среды и те эволюционные процессы, которые генерируют и поддерживают это генетическое разнообразие в популяциях.
Эти концепции важнейшие, однако их практически невозможно прямо применить к проблеме выделения территориальных границ популяций, так как информация об адаптивных генотипах и обусловливающих их динамику средовых факторах в конкретных популяциях часто отсутствует, а получить ее напрямую пока проблематично. Более того, как показывают данные широкогеномных исследований, такую информацию очень трудно получить в достаточном объеме еще и потому, что локальные адаптации и вызванные ими генетические изменения в популяциях иногда “параллельны” друг другу (Pearse et al., 2014), а иногда и нет (Perrier et al., 2013).
На основе этих концепций были предложены разнообразные подходы к использованию доступных экологических характеристик, которые могли бы быть ассоциированы с адаптивностью популяций. Этого можно достичь, например, выделяя на ареале вида важные экологические градиенты и разбивая вид на соответствующие группировки (Moritz, 2002) или используя подход ландшафтной генетики, чтобы связать генетические характеристики популяций с экологическими параметрами их местообитаний (Dionne et al., 2008; Manel et al., 2010; Sork, Waits, 2010). Можно также выделить “проектируемые единицы” (COSEWIC, 2015): сначала выявить группировки особей путем анализа разных признаков (морфологических, поведенческих, нейтральных генетических маркеров и др.), а затем доказать их филогенетическую и иную значимость и оценить различие популяционных единиц. Помимо этого можно выделить эволюционно значимые единицы в терминах генетических маркеров, жизненных стратегий и условий среды (Waples et al., 2001; Waples, 2006; Waples, Gaggiotti, 2006).
Практическое использование указанных концепций и подходов для выделения популяций затруднено тем, что связи между локальными адаптациями и межпопуляционными различиями скрыты от глаз исследователя из-за взаимодействий генотип–среда и других нелинейных взаимосвязей между генетической изменчивостью, фенотипическими вариациями и экологическими параметрами (Waples, 1991; Gagnaire et al., 2012). К тому же молекулярная природа популяционных адаптаций, за редкими исключениями, неизвестна (Fraser et al., 2011). Более того, геномные данные свидетельствуют о том, что действие множества минорных генов, которые вместе вносят основной вклад в популяционную изменчивость адаптивных количественных признаков, уловить крайне сложно (Yang et al., 2011).
Экологические и генетические маркеры. Для районирования популяций ранее было предложено использовать раздельно средовые/экологические и генетические маркеры (Животовский, 2016, 2017).
Маркеры местообитания (средовые и экологические): физико-химические характеристики условий местообитания вида, биота, параметры воспроизводства популяций, морфофизиологические признаки, численность, индуцированная средой экспрессия генов, миграционные отношения между популяциями, т.е. любые характеристики косной среды и сопряженных видов, а также признаки исследуемого вида, проявление которых зависит не только от генотипа особей, но и, главное, от средовых факторов.
ДНК-маркеры популяции (определяются последовательностями ДНК): однонуклеотидные замены, делеции и вставки, микросателлиты, вариабельные нуклеотидные последовательности, аллозимные полиморфизмы и пр.
Преимущество маркеров местообитания – ассоциация с факторами адаптации, а недостаток – отсутствие данных об их генетической детерминации. Плюс ДНК-маркеров – они представляют собой фрагменты генома, а минус – их неоднозначная связь с адаптацией популяций, так что использование их в таком качестве для выделения популяций пока малореально.
Выделять и сравнивать популяции только по экологическим маркерам недостаточно, так как редко можно строго научно доказать, что выявляемые по ним различия между популяциями наследственно обусловлены. Исследовать же только генетические маркеры также недостаточно из-за возможных смещений в оценках популяционно-генетических параметров, обусловленных особенностями популяционно-генетической динамики и нередко низкой представительностью выборок по отношению к популяционной структуре вида. Для их корректировки как раз и нужны экологические маркеры.
ЭКОГЕОГРАФИЧЕСКИЕ ЕДИНИЦЫ
Принцип выделения экогеографических единиц. При изучении популяций лососевых рыб нами был предложен общий, двухэтапный подход к изучению популяционной структуры вида, основанный на совместном использовании экологических/средовых и генетических маркеров (Zhivotovsky et al., 2015; Животовский, 2016, 2017).
Первый этап. Выделяют ЭГЕ по доступным данным о географии, градиентах среды обитания, миграционных особенностях (генных потоках) и других экологических и средовых маркерах.
ЭГЕ – это популяция или группа соседних популяций, которые отграничены на ареале эколого-географическими градиентами; имеют сходные морфологические, физиологические или иные биологические признаки, предположительно ассоциированные с адаптациями к этим градиентам; связаны друг с другом генными потоками в ряду поколений и значительно репродуктивно изолированы от других популяций этого вида. Под “генным потоком в ряду поколений” понимается возможность обмена генами между популяциями, причем не обязательно за одно поколение, а пошагово – за ряд поколений.
Второй этап. ЭГЕ, определенные на первом этапе, генетически тестируют для оценки корректности объединения тех или иных популяций в ЭГЕ путем анализа изменчивости ДНК-маркеров внутри и между ЭГЕ (по показателям генетического сходства или расстояния между популяциями, генетическим кластерам особей или популяций, оценкам межпопуляционных генных обменов и пр.).
В практических приложениях трудно оценить интенсивность генных потоков и степень репродуктивной изоляции, поэтому в качестве критерия связности генными потоками внутри ЭГЕ и изоляции от других ЭГЕ можно использовать, например, такой: ЭГЕ считаются корректно выделенными, если генетические различия между популяциями внутри ЭГЕ меньше, чем их генетические отличия от популяций других ЭГЕ (Zhivotovsky et al., 2015; Животовский, 2019).
Несомненно, помимо генетического тестирования ЭГЕ нужны сравнительные исследования общей популяционной изменчивости внутри и между ЭГЕ по разным характеристикам: морфофизиологическим, поведенческим, численности, миграционной связности и др.
Как выделять экогеографические единицы? Первый, основной этап нашего подхода – это выделение (районирование) ЭГЕ. Этот этап требует сбора имеющихся и получения новых данных о среде обитания и экологии вида, ботанико- и зоогеографического районирования исследуемой части ареала вида, определения физико-химических и других параметров среды, а также наличия соответствующих тематических географических карт.
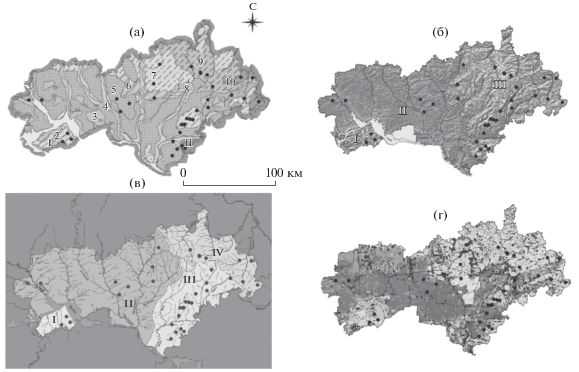
Рассмотрим процедуру выделения ЭГЕ на примере изученных нами ранее речных гидробионтов о. Сахалин (рис. 1а), на которых был впервые опробован принцип выделения ЭГЕ. Исходя из концепции ЭГЕ следует вначале разбить ареал исследуемого вида на эколого-географические районы, используя характеристики местообитания, важные для изучаемого вида, а далее подразделять эти районы согласно данным об экологии вида. Для этого нам следует иметь информацию о градиентах условий среды на исследуемой территории. Например, для пресноводных рыб и других гидробионтов, репродукция которых проходит в реках и озерах, ЭГЕ можно выделять по бассейнам стока рек, ихтиофауне, гидробиологическим особенностям, теплообеспеченности, приуроченности данного вида к тем или иным местообитаниям, миграциям и пр. Источниками такой информации могут быть как результаты специальных исследований, так и данные об изучаемой части ареала вида, полученные, в частности, из тематических географических карт.
Рис. 1.
Этапы выделения экогеографических единиц сахалинского тайменя на Сахалине. а – карта Сахалина с речной сетью; б – основные водоразделы (жирные линии), подразделяющие остров на четыре района; в – зоогеографическое районирование ихтиофауны Сахалина (вместе с водоразделами разбивает территорию Сахалина на шесть районов); г – распределение экологических форм тайменя по характеру весенне-летне-осенних миграций (1 – анадромная, 2 – лагунная, 3 – озерная, 4 – речная); д – подразделение территории Сахалина с одновременным учетом водоразделов, типа ихтиофауны и экоформ тайменя; полученные 15 районов приняты за экогеографические единицы сахалинского тайменя на Сахалине.

Обратимся, например, к разбиению ареала вида по бассейновому принципу. Водосборный бассейн – это целостная единица в организации ландшафта – задает гидрологический режим территории, определяет особенности температурного режима и другие характеристики водоема (Мартыненко, Бочарников, 2008; Корытный, 2017). Поэтому водоразделы между крупными озерно-речными системами являются естественными границами экосистем и составляющих их популяций, а следовательно ЭГЕ. Бассейновый принцип выделения ЭГЕ важен, в частности, для проходных рыб (кеты и других тихоокеанских лососей), так как пресноводный период (период развития от оплодотворенной икры до ската молоди и ее нагула в опресненных водах прибрежья) важнейший в их онтогенезе; на него приходится основная адаптивная нагрузка и закладка механизма хоминга, определяющего возврат рыбы к родным берегам. На рис. 1б показаны основные водоразделы Сахалина.
Однако водоразделы не единственные возможные границы ЭГЕ. Другими границами может служить биота бассейнов – индикатор условий этих бассейнов и среда обитания гидробионтов. В этом качестве для тихоокеанских лососей и других лососевых рыб дальневосточного региона, у которых критический период онтогенеза или вся жизнь проходят в реках и озерах, мы принимаем ихтиологическое районирование, описанное ранее (Черешнев, 1998) (рис. 1в). Можно использовать также и другие подразделения ареала: широтное (теплообеспеченность), ботанико-географическое (характеризующее флору водосборного бассейна), по типу почв и т.д.
Для конкретного вида существуют биогеографические границы популяционных группировок, определяемые их биологическими особенностями, в том числе временем нереста, типом нерестилищ, поведением, питанием, морфофизиологическими признаками, возможными миграционными обменами.
Для конкретизации обратимся к сахалинскому тайменю Parahucho perryi (Brevoort, 1856), краснокнижному виду, основной ареал которого – о. Сахалин. Сахалинские ученые С.Н. Сафронов и В.Д. Никитин выделили у него четыре экологические формы в озерно-речных системах Сахалина (рис. 1г) (Zhivotovsky et al., 2015). Объединение районирования этих экоформ с предыдущим разбиением территории Сахалина по водоразделам и ихтиофауне позволяет выделить 15 районов и соответствующие 15 группировок вида, которые мы и принимаем в качестве ЭГЕ сахалинского тайменя на Сахалине (рис. 1д). Существенно, что анализ микросателлитных ДНК-маркеров позволил тестировать выделенные ЭГЕ и выявить большие генетические различия между ними (Zhivotovsky et al., 2015).
Важно отметить, что географические границы, выделенные для одних видов по части экологических маркеров (например, только по водоразделам или по водоразделам вместе с ихтиофаунистическими границами), могут быть те же самые и для других видов, однако дополненные своими биогеографическими границами. Например, для тихоокеанского лосося кеты Oncorhynchus keta (Walbaum, 1792), заходящей в реки Сахалина, учитывали также наличие сезонных рас (летней и осенней), нерестящихся в разное время в одних и тех же водоемах (хотя и на разных типах нерестилищ), наличие заводских стад и других особенностей вида. Выделенные ЭГЕ этого вида также характеризовали большие генетические отличия друг от друга по сравнению с различиями популяций внутри ЭГЕ (Животовский, 2019).
Интерпретация ЭГЕ. Вне зависимости от того животные это или растения, опираясь на сведения о выделении и особенностях ЭГЕ данного вида, можно сделать вывод, что ЭГЕ – это природное тело, скомпонованное из эколого-географических характеристик района обитания составляющих его популяций и биологических признаков этих популяций. Таким образом, в территориальном аспекте ЭГЕ представляют собой результат эколого-географического районирования популяций (как на рис. 1). Отметим, что ЭГЕ это лишь один из элементов географической организации вида, его популяционной структуры (Животовский, 2016).
ЭГЕ удобно использовать для решения практической задачи – выделения единицы запаса как территориально обозначенной, районированной части вида, в целях разработки единой стратегии управления природными популяциями. При усиливающемся сегодня антропогенном давлении на природу это необходимо делать как для экономически важных, ресурсных видов, так и для находящихся под угрозой исчезновения редких и охраняемых видов животных и растений.
ВЫДЕЛЕНИЕ ЭКОГЕОГРАФИЧЕСКИХ ЕДИНИЦ У РАСТЕНИЙ
Выделение ЭГЕ, которые определяются по параметрам ареала и биологическим признакам популяций, важно не только для животных, но в еще большей степени для растений (Животовский, Османова, 2018, 2019а–в; Османова и др., 2019).
Возможность выделения внутривидовых группировок и их ареала определяется характером адаптации изучаемого вида растений к среде обитания. На внутривидовом уровне адаптация выражается в специфической приспособленности подвидов, популяций, ценопопуляций и отдельных групп растений к своим условиям среды, причем такая приспособленность наследуется в поколениях. Например, у подорожника ланцетолистного Plantago lanceolata L. была обнаружена экотопическая дифференциация, обусловленная разным световым режимом (Teramura, 1983). На хорошо освещенном участке растения P. lanceolata были многорозеточными и свободно растущими, а на периодически затеняемом – преимущественно мелкими, однорозеточными, росли скученно. Растения с каждого из этих участков были расклонированы и высажены на контрастные среды со световыми режимами исходных участков. Результаты показали, что каждая форма была более приспособлена к световому режиму ее родного участка (Teramura, 1983).
Другой пример: были взяты раметы клевера ползучего Amoria repens (L.) C. Presl, произрастающего на участках пастбища с доминированием разных злаков (плевела многолетнего Lolium perenne L., бухарника шерстистого Holcus lanatus L., гребенника обыкновенного Cynosurus cristatus L., полевицы тонкой Agrostis tenuis Sibth.). Перекрестная трансплантация расклонированных рамет выявила, что каждый тип рамет растет лучше всего в исходной, родной для него дернине или в смеси злаков с доминированием того вида, что и на родном участке (Turkington, Harper, 1979).
Таким образом, градиенты условий на ареале вида (разные типы почв и растительности, разная обеспеченность влагой и теплом и другие вариации параметров среды) – источники устойчивой внутривидовой дифференциации. Их можно привлечь для подразделения исследуемой части вида и ее ареала на ЭГЕ.
Для выделения ЭГЕ у растений могут быть выбраны разнообразные характеристики среды обитания (Животовский, Османова, 2018, 2019а–в): почвенные и гидрологические условия, типы растительности, особенности климата, используемые для районирования по ботаническому, эколого-географическому, бассейновому и иным принципам (Абрамов, 2001; Смыков, 2008; Демаков, 2018; Национальный…, 2018; и др.).
Так как часто источник подобной информации – тематические географические карты (типов почв, растительности, климата, районирования, особо охраняемых природных территорий и пр.), то для качественного анализа этой информации важно использовать ГИС-технологии (Животовский, Османова, 2018, 2019а). Кроме того, в целях детальной оценки экологических характеристик местообитаний особую важность приобретают геоботанические описания с дальнейшим анализом флористического состава и оценкой экологических условий по разным шкалам (Раменский и др., 1956; Цыганов, 1983).
Для выделения ЭГЕ важно оценить, как далеко распространяются пыльца и диаспоры (семена, плоды, споры и др.), что в определенной степени характеризует межпопуляционный обмен. Для приблизительной оценки дальности генного потока мы условно выбираем величину, кратную дальности разноса пыльцы и диаспор. Интенсивность генного потока можно оценить также по данным об изменчивости ДНК-маркеров.
ПРИМЕР ВЫДЕЛЕНИЯ ЭКОГЕОГРАФИЧЕСКИХ ЕДИНИЦ У РАСТЕНИЙ
Мы исследовали распространение в районе средней Волги (территория Республики Марий Эл) редкого вида орхидных – башмачка настоящего Cypripedium calceolus L., конкурентно-стресс-толерантного короткокорневищного вида. Обязательное условие для развития проростков башмачка настоящего – присутствие гриба Armillaria mellea (Vahl: Fr.) Kummer. из рода Rhizoctonia (Summerhayes, 1951; Eberle, 1972: цит. по Денисова, Вахрамеева, 1978). Таким образом, территориальное распределение башмачка настоящего во многом определяется наличием в почве гриба-микоризообразователя. О распространении гриба A. mellea известно очень мало. По этой причине мы рассматриваем данный пример с C. calceolus только как иллюстрацию подхода к выделению ЭГЕ.
Уточненные и дополненные Г.А. Богдановым (устное сообщение) данные о распространении этого вида в исследуемом районе взяты из Красной книги Республики Марий Эл (Красная…, 2013). Для анализа эколого-географической информации об условиях произрастания использовали разные карты (Карта лесного…, 2018; Карта рельефа…, 2018; Карта речных…, 2018; Национальный…, 2018).
C. calceolus встречается на исследованной территории практически везде вне зависимости от типа почв (рис. 2а), речных бассейнов (2б), рельефа (2в) и типа растительности (2г). На исследованной территории встречаются дерново-подзолистые, торфяные болотные, серые лесные, пойменные слабокислые и нейтральные почвы; почвообразующими являются песчаные, средне- и легкосуглинистые, глинистые и тяжелые глинистые породы (рис. 2а). Однако распределение этого вида по изученной территории неравномерное: наиболее многочисленные ценопопуляции C. calceolus отмечены в районе Марийско-Вятского увала, который имеет сложное геологическое строение, определившее его высокую почвенную и микроклиматическую гетерогенность (Ступишин и др., 1969), и обусловливает сложный геолого-геоморфологический облик данной части территории (III на рис. 2б). Он представлен двумя физико-географическими районами (Васильева, 1979): территория одного из них – в Камском бассейне (IV на рис. 2в) – сложена песчаниками, глинами, доломитами, мергелями пермского периода, а второго района – в бассейне р. Илеть (III на рис. 2в) – сложена известняково-доломитовыми толщами казанского яруса и глинистыми отложениями татарского яруса. Наличие карбонатных пород определяет разнородный, азональный почвенный покров, из-за чего здесь даже встречаются представители степной флоры (Васильева, 1979).
Рис. 2.
Местообитания Cypripedium calceolus в районе средней Волги (черные точки) в проекциях на эколого-географические карты. a – почвенная карта. Типы почв: 1 – дерново-подзолистые (глинистые и тяжелосуглинистые); 2 – дерново-подзолистые среднесуглинистые; 3 – торфяные болотные низинные; 4 – пойменные слабокислые и нейтральные; 5 – дерново-подзолистые иллювиально-железистые песчаные; 6 – дерново-подзолистые (глинистые и суглинистые), подстилаемые песчаными и супесчаными породами; 7 – дерново-подзолистые суглинистые; 8 – подзолы глеевые торфянистые и торфяные, преимущественно иллювиально-гумусовые; 9 – светло-серые лесные суглинистые; 10 – серые лесные среднесуглинистые; 11 – светло-серые лесные (глинистые и тяжелосуглинистые). б – карта рельефа. I – Приволжская возвышенность, II – Марийская низменность, III – Марийско-Вятский увал с отрогами. в – карта крупных речных бассейнов и природных областей. I – правобережье Волги, лесостепная область; II – южно-таежная область, бассейны рек Марийской низменности; южно-таежная область Марийско-Вятского увала: III – бассейн р. Илеть, IV – Камский бассейн. г – карта растительности исследованного района (белый цвет – обезлесенные территории).
Выделение экогеографических единиц. Принимая радиус потенциальных генных потоков равным 5 или 10 км, можно сопоставить ценопопуляции башмачка обыкновенного по их географической близости друг к другу и по экологическому сходству их местообитаний (рис. 3). Соответственно, применяя изложенные выше принципы, можно выделить ЭГЕ у этого вида. Мы включаем в одну ЭГЕ лишь те ценопопуляции, которые могут быть связаны генными потоками и при этом обитают в сходных условиях. Если таких местообитаний в близком соседстве нет, то ЭГЕ состоит из одной ценопопуляции (рис. 4).
Рис. 3.
Связность исследованных ценопопуляций Cypripedium calceolus потенциальными генными потоками. Черные круги – пятикилометровая дальность потенциальных генных потоков, серые – генные потоки в радиусе 10 км.
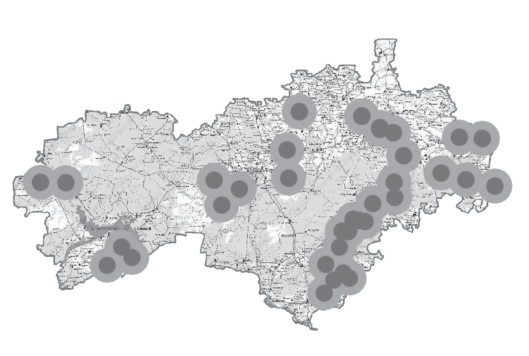
Рис. 4.
Экогеографические единицы Cypripedium calceolus, проецированные на почвенную карту. Каждая ЭГЕ, которая включала в себя две или более популяций, выделена по схожим условиям среды: один тип почв, один речной бассейн. Те ЭГЕ, которые состоят из одной ценопопуляции, лежат вне возможных миграционных связей с другими ценопопуляциями или обитают в иных условиях, чем те ценопопуляции, с которыми такие связи не исключены.
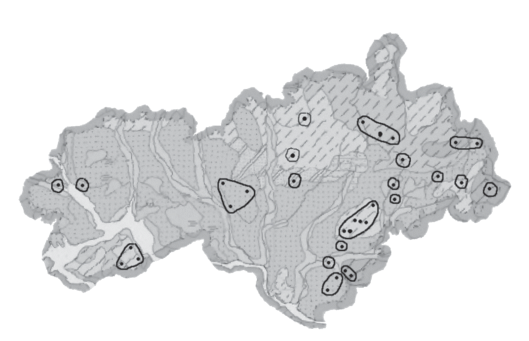
Те ЭГЕ, которые включают в себя две или более ценопопуляций C. calceolus, обитают в близких фитоценозах, растут на сходных почвах, находятся в одинаковых речных бассейнах, и потенциально связаны генными потоками. Многие ценопопуляции C. calceolus образуют ЭГЕ в одиночестве, обитая в своих уникальных условиях и находясь достаточно далеко друг от друга. Скорее всего, это естественное свойство редкого вида, находящегося на краю ареала или в пессимальных условиях обитания. Отметим, что, несмотря на связность цепи популяций C. calceolus вдоль Марийско-Вятского увала (рис. 3), именно здесь мы выделили большое число ЭГЕ (рис. 4). Это связано с высокой почвенной и микроклиматической гетерогенностью Марийско-Вятского увала, обусловленной его сложным геологическим строением (Ступишин и др., 1969).
Интерпретация экогеографических единиц. Как отмечено выше, редкий в среднем Поволжье башмачок настоящий был выбран нами лишь для иллюстрации подхода к выделению ЭГЕ. Тем не менее уже на этом примере видно, что выделенные по экологическим/средовым условиям ЭГЕ позволяют использовать их для разработки стратегии охраны редких или ценных в хозяйственном отношении растений. Например, некоторые местообитания C. calceolus и соответствующие ЭГЕ попадают в особо охраняемые природные территории (Богданов и др., 2015), но большая часть их туда не входит. Поэтому, выделяя ЭГЕ редких видов растений, можно предложить дополненный список отводимых под охрану территорий.
Предложенный нами принцип выделения у растений ЭГЕ с помощью данных об экологии вида, геоботанических описаний и картографического анализа местообитаний позволяет использовать полученные ЭГЕ как единицы эколого-географической структуры вида, его охраны, воспроизводства и эксплуатации. Более того, выделенные большие ЭГЕ могут служить источником сбора образцов растений для разведения в ботанических садах в целях их реинтродукции в естественные местообитания в дальнейшем.
Несомненно, окончательное решение о границах каждой ЭГЕ требует генетического тестирования – использования ДНК-маркеров для сравнения составляющих их ценопопуляций друг с другом, оценки реальных генных потоков между ними, решения о слиянии соседних ЭГЕ друг с другом или, напротив, их разделении в целях дальнейшего уточнения их списка (Животовский, 2016, 2017).
Предложенная методика выделения ЭГЕ у растений применима не только к редким, но и к ценным в хозяйственном отношении растениям, например ресурсным видам. Более того, популяционно-биологический базис методологии выделения ЭГЕ взаимосвязан с проблемами экологии и генетики природных популяций (Миркин, Наумова, 1998; Злобин, 2009; Марков, 2012), а ее картографическая составляющая связывает эти проблемы с практическими запросами землепользования, анализа лесного и почвенного фондов, охраны природы (Заугольнова, 1999; Ханина и др., 2000; Зверев, 2007; Смыков, 2008; Демаков, 2018).
ЭКОГЕОГРАФИЧЕСКИЕ АГРЕГАЦИИ РЕДКИХ ВИДОВ
Проблема выделения многовидовых экогеографических агрегаций. Растение считается редким на данной территории, если оно малочисленное и встречается в небольшом числе местообитаний. Редкие виды – часть общего биоразнообразия, иногда они высоко ценятся из-за своей красоты или полезных свойств, но из-за своей редкости и узких рамок подходящих условий существования находятся на краю выживания (Злобин и др., 2013). Поэтому для них велик риск исчезнуть, если не предпринимать усилий по их сохранению. Основные причины исчезновения редких видов – естественное изменение среды обитания (сильные засухи, лесные пожары неантропогенного происхождения, климатические изменения и пр.), деятельность человека (загрязнение атмосферы и гидросферы, деградация почвенного покрова, сбор ягод и частей растений, выпас скота, осушение болот и др.). Таким образом, изучение редких видов имеет большое прикладное значение, так как они первыми сигнализируют о неблагополучии экосистем.
Редких видов много. Например, на территории Республики Марий Эл >150 видов растений занесены в региональную или национальную Красные книги. Важно выявить территориально близкие скопления ценопопуляций разных видов – экогеографические агрегации (ЭГА) по информации об их распространении и адаптации к условиям их местообитаний в целях разработки стратегии их охраны, в том числе планирования дополнительных охраняемых территорий. Для этой цели можно применить тот же принцип, что и для выделения ЭГЕ.
Объекты исследования. Были выбраны наиболее редкие виды растений в Республике Марий Эл. Все их основные местообитания были известны: они представляли собой разные семейства и жизненные формы (38 видов). В результате такого выбора эти виды характеризовались наличием одной ценопопуляции или небольшим их числом: Apiaceae – берула прямая Berula erecta (Huds.) Hoffm., лазурник трехлопастной Laser trilobum (L.) Borkh., морковь дикая Daucus carota L., мордовник шароголовый Echinops schaerocephalus L.; Araceae – аир обыкновенный Acorus calamus L.; Aspleniaceae – костенец постенный Asplenium ruta-muraria L.; Asteraceae – василек ложнопятнистый Centaurea рseudomaculosa Dobrocz.; Athyriaceae – голокучник Роберта Gymnocarpium robertianum (Hoffm.) Newm., диплазий (орлячок) сибирский Diplazium sibiricum (Turcz. ex Kunze) Kurata; Boraginaceae – пупочник ползучий Omphaloides scorpioides (Hanke) Schrank; Botrychiaceae – гроздовник ромашколистный Botrychium matricariifolium A. Br. ex Koch; Brassicaceae – зубянка пятилистная Dentaria quinquefolia Bieb., лунник оживающий Lunaria rediviva L.; Cyperaceae – осока малоцветковая Carex pauciflora Lightf.; осока метельчатая Carex paniculata L.; Dipsacaceae – короставник татарский Knautia tatarica (L.) Szabo; Dryopteridaceae – многорядник Брауна Polystichum braunii (Spenn) Feé; Fabaceae – астрагал серповидный Astragalus falcatus Lam., вязель пёстрый Coronilla varia L., дрок германский Genista germanica L., люпинник пятилистный или клевер люпиновый узколистный Lupinaster pentaphylloides Moench ssp. angustifolius (Litv.) Soják, остролодочник волосистый Oxytropis pilosa (L.) DC.; Fumariaceae – хохлатка Маршалла Corydalis marschalliana (Pall. ex Willd.) Pers.; Lamiaceae – тимьян блошиный Thymus pulegioides L., шалфей мутовчатый Salvia verticillata L.; Najadaceae – наяда большая Najas major All., наяда гибкая N. flexilis (Willd.) Rostk. Et Schmidt; Oleaceae – ясень обыкновенный Fraxinus excelsior L.); Ophioglossaceae – ужовник обыкновенный Ophioglossum vulgatum L.; Orchidaceae – калипсо луковичная Calypso bulbosa (L.) Oakes; Ranunculaceae – ветреничка алтайская Anemonoides altaica (C. A. Mey.) Holub; Rosaceae кизильник черноплодный Cotoneaster melanocarpus Fisch. ex Blutt, морошка приземистая Rubus chamaemorus L.; Scrophulariaceae – мытник скипетровидный Pedicularis sceptrum-carolinum L.; Trapaceae – водяной орех, чилим плавающий Trapa natans L. s. l.; Zannichelliaceae – занникеллия ползучая Zannichellia repens Boenn.
Данные о распространении вида взяты из Красной книги (Красная…, 2013) и других материалов. В качестве меры связности ценопопуляций одного вида возможными генными потоками мы выбрали пятикилометровый радиус вокруг каждого местообитания исследованных видов. Для описания среды обитания использовали картографические данные по типам почв (Национальный…, 2018), растительности (Карта лесного…, 2018), рельефу (Карта рельефа…, 2018) и охраняемым природным территориям (Богданов и др., 2015) и ГИС-технологию анализа географических карт (Quantum GIS, 2017).
Многовидовые экогеографические агрегации. С помощью рассмотренного выше подходa для каждого из 38 видов мы выделили ЭГЕ, выбрав пятикилометровый радиус потенциальных генных потоков (рис. 5). Те ЭГЕ, которые оказались территориально близки друг к другу и обитали в сходных почвенных и иных условиях, были объединены в ЭГА (всего 11 многовидовых ЭГА) (рис. 6).
Рис. 5.
Местообитания исследованных 38 редких видов (черные точки), спроецированные на почвенную карту Республики Марий Эл. Серые круги – пятикилометровые зоны вокруг каждого местообитания каждого вида.
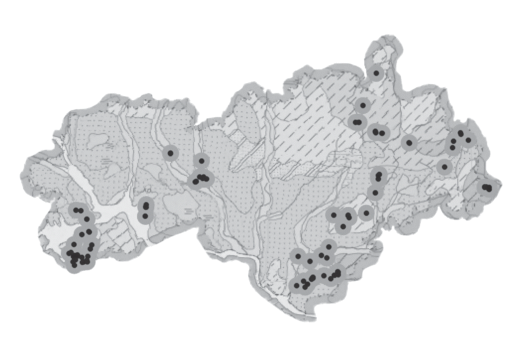
Рис. 6.
Участки территории, занимаемые многовидовыми экогеографическими агрегациями (ЭГА) исследованных редких видов (1–11), спроецированные на карту рельефа Республики Марий Эл. ЭГА выделены на основе группирования территориально близких экогеографических единиц в пределах сходных условий среды: 1 – C. marschalliana, D. carota, D. quinquefolia, F. excelsior, N. major, O. vulgatum; 2 – A. calamus, F. excelsior, T. natans; 3 – C. paniculata, P. sceptrum-carolinum; 4 – B. matricariifolium, C. pauciflora, G. germanica, N. major, R. chamaemorus; 5 – C. varia, L. pentaphylloides, L. trilobum, O. scorpioides, O. vulgatum, Th. pulegioides, Z. repens; 6 – B. erecta, C. pauciflora, K. tatarica, N. flexilis, O. vulgatum; 7 – D. sibiricum, G. robertianum, L. rediviva, O. vulgatum, P. braunii; 8 – A. rutamuraria, C. melanocarpus, D. sibiricum, G. robertianum, O. pilosa; 9 – A. falcatus, E. schaerocephalus, O. pilosa; 10 – A. falcatus, C. рseudomaculosa, O. pilosa; 11 – C. bulbosa, D. sibiricum, S. verticillata.
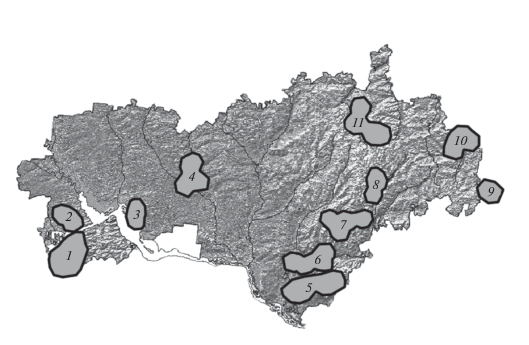
Большинство ЭГА (6–8, 11) сконцентрированы в районе Марийско-Вятского увала (рис. 6). Как указывалось выше, там отмечены карбонатные почвы и близкое залегание мергелей, что является предпочтительным для видов, включенных в эти агрегации, и что привело к группированию их ценопопуляций на одних и тех же участках.
Самая большая ЭГА-5 содержит семь видов, относящихся к разным экологическим группам и разным жизненным формам. Она находится в Волжском районе республики, который является северной границей всех этих видов и примыкает к Марийско-Вятскому увалу.
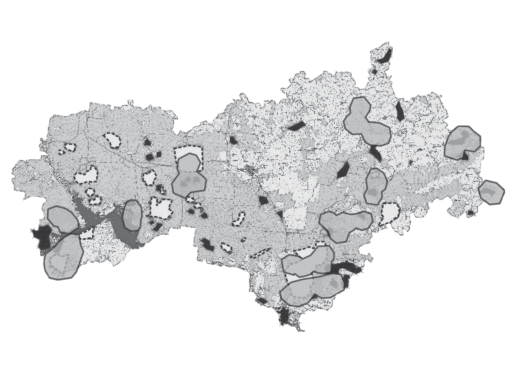
Следует отметить, что, хотя все эти виды редкие и указаны в Красной книге, далеко не все из выделенных ЭГА входят в существующие или планируемые природоохранные зоны, при том что обширная сеть особо охраняемых природных территорий (ООПТ) Республики Марий Эл охватывает основные типы ландшафтов – лесные, луговые, водные и болотные (рис. 7).
Рис. 7.
Взаимное расположение многовидовых экогеографических агрегаций (ЭГА) и охраняемых природных территорий в Республике Марий Эл. Серые области со сплошной границей – ЭГА (те же самые, что на рис. 6). Сквозь них просвечивают границы охраняемых территорий: белые области с пунктирными границами – существующие заказники, памятники природы и особо охраняемые природные территории федерального значения, зачерненные области – проектируемые охраняемые территории.
ВАЖНОСТЬ ВЫДЕЛЕНИЯ ЭКОГЕОГРАФИЧЕСКИХ ЕДИНИЦ И ЭКОГЕОГРАФИЧЕСКИХ АГРЕГАЦИЙ
Выявление ЭГА редких видов позволяет подойти к выделению ООПТ научно обоснованными методами. Выделенные участки территории, на которых расположены многовидовые ЭГА редких видов (рис. 7), можно считать кандидатами на создание дополнительных охраняемых природных территорий (с учетом имеющихся коммуникаций, сельскохозяйственных угодий и других ограничений). Особенно это важно для входящих в ЭГА видов, находящихся на грани исчезновения, например водных растений N. major, N. flexilis, Z. repens и T. natans (загрязнение и эвтрофикация водоемов привели к резкому сокращению ценопопуляций этих видов).
Реализация предложенной методологии выделения ЭГЕ и ЭГА редких видов в большой степени основывается на информации, полученной из тематических географических карт и Красных книг. Создание региональных и национальных карт (растительности, почв, климата и др.) и Красных книг стало значительным шагом в деле защиты природы и объектов животного и растительного мира. Их основное предназначение – накопление и распространение научно обоснованной информации о природном разнообразии, в том числе о редких, исчезающих и находящихся под угрозой исчезновения представителях флоры и фауны.
Не менее важно создание доступных баз данных по региональным тематическим картам, районированию, геоботаническим описаниям и другой эколого-географической информации. Такая информация необходима для разработки и реализации мер по охране, восстановлению и рациональному использованию биоразнообразия, в том числе при подготовке рекомендаций по выделению охраняемых природных территорий.
Работа выполнена в рамках программы гостемы ГЗ 0112-2019-2 (подтема “Эколого-генетическая структура вида”) и Программы фундаментальных исследований президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России”, при финансовой поддержке РФФИ (грант 18-016-00033 в части выделения экогеографических единиц).
Список литературы
Абрамов Н.В. Флора Республики Марий Эл: инвентаризация, анализ, районирование, охрана и проблемы рационального использования ее ресурсов: Автореф. дис. д-ра биол. наук в форме науч. докл. Пермь: ПГУ, 2001. 60 с.
Алтухов Ю.П. Генетические процессы в популяциях. М.: Наука, 1983. 280 с.
Богданов Г.А., Демаков Ю.П., Исаев А.В. Схема развития сети особо охраняемых природных территорий Республики Марий Эл. Научные фонды ГПЗ “Большая Кокшага”. Йошкар-Ола. 2015. 125 с.
Васильева Д.П. Ландшафтная география Марийской АССР. Йошкар-Ола: Мар. кн. изд-во,1979.134 с.
Демаков Ю.П. Структура и закономерности развития лесов Республики Марий Эл. Йошкар-Ола: Поволж. гос. техн. ун-т, 2018. 432 с.
Денисова Л.В., Вахрамеева М.Г. Род башмачок (Венерин башмачок) – Cypripedium L. // Биологическая флора Московской области. Вып. 4. М.: МГУ, 1978. С. 62–70.
Животовский Л.А. Популяционная структура вида: экогеографические единицы и генетическая дифференциация популяций // Биология моря. 2016. Т. 42. № 5. С. 323–333.
Животовский Л.А. Две ветви исследований популяционной структуры вида – экологическая и генетическая: история, проблемы, решения // Генетика. 2017. Т. 53. С. 1244–1253.
Животовский Л.А. Экогеографические единицы и единицы запаса вида в пресноводных экосистемах (на примере кеты о. Сахалин) // Принципы и способы сохранения биоразнообразия: Матер. VII Междунар. науч. конф. Йошкар-Ола: Вертикаль, 2019. С. 276–278.
Животовский Л.А., Османова Г.О. Эколого-географический подход к выявлению популяционной структуры вида у растений // Экология и география растений и растительных сообществ: Матер. IV Междунар. науч. конф. Екатеринбург: АНО ВО “Гуманитарный университет”, 2018. С. 282–285.
Животовский Л.А., Османова Г.О. Популяционная биогеография растений. Йошкар-Ола: Вертикаль, 2019а. 128 с.
Животовский Л.А., Османова Г.О. Экогеографические единицы и охрана внутривидового разнообразия // Изв. РАН. Сер. биол., 2019б. С. 12–24.
Животовский Л.А., Османова Г.О. Экогеографические единицы и популяционная структура вида // Принципы и способы сохранения биоразнообразия: Матер. VII Междунар. науч. конф. Йошкар-Ола: Вертикаль, 2019в. С. 10–13.
Заугольнова Л.Б. Иерархический подход к анализу лесной растительности малого речного бассейна (на примере Приокско-Террасного заповедника) // Бот. журн. 1999. Т. 84. № 8. С. 42–56.
Зверев А.А. Информационные технологии в исследованиях растительного покрова. Томск: ТМЛ-Пресс, 2007. 304 с.
Злобин Ю.А. Популяционная экология растений: современное состояние, точки роста. Сумы: Университет. книга, 2009. 263 с.
Злобин Ю.А., Скляр В.Г., Клименко А.А. Популяции редких видов растений (Теоретические основы и методика изучения). Сумы. Университет. книга, 2013. 439 с.
Карта лесного фонда Республики Марий Эл. URL: https://fs00.infourok. ru/images/doc/160/185032/img50.jpg (дата обращения: 11.01.2018).
Карта рельефа Республики Марий Эл. URL: https://commons.wikimedia. org/wiki/File:Relief_map_of_Mari_El.png (дата обращения: 11.01.2018).
Карта речных бассейнов Республики Марий Эл. URL: https://commons. wikimedia.org/wiki/File:Реки_Марий_Эл.svg (дата обращения: 11.01.2018).
Корытный Л.М. Бассейновая концепция: от гидрологии к природопользованию // География и природ. ресурсы. 2017. № 2. С. 5–16.
Красная книга Республики Марий Эл. Том “Растения. Грибы” / Сост. Богданов Г.А., Абрамов Н.В., Убранавичюс Г.П., Богданова Л.Г. Йошкар-Ола: Мар. гос. ун-т, 2013. 324 с.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978. 351 с.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
Марков М.В. Популяционная биология растений. М.: КМК, 2012. 387 с.
Мартыненко А.Б., Бочарников В.Н. Экологическое районирование Дальнего Востока // Изв. РАН. Сер. геогр. 2008. № 2. С. 76–84.
Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, 1998. 413 с.
Национальный атлас почв Российской Федерации. Информационные ресурсы факультета почвоведения МГУ. URL: https://soilatlas.ru/ (дата обращения: 04.01.2018).
Османова Г.О., Богданов Г.А., Животовский Л.А. Выделение многовидовых экогеографических агрегаций редких видов растений в целях организации охраняемых природных территорий (на примере флоры Республики Марий Эл) // Экология. 2019. № 5. С. 373–377.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 472 с.
Смыков А.Е. Закономерности пространственно-временной динамики основных параметров лесного фонда Республики Марий Эл: Автореф. дис. канд. с.-х. наук. Йошкар-Ола: Мар. гос. техн. ун-т, 2008. 23 с.
Ступишин А.В., Лаптева Н.Н., Васильева Д.П. Физико-географическое районирование территории Марийской АССР // Географический сборник, № 4. Казань: Изд-во КГУ, 1969. С. 3–8.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. М.: Наука, 1973. 277 с.
Ханина Л.Г., Заугольнова Л.Б., Смирнов В.Э., Глухова Е.М. Методика анализа биоразнообразия растительного покрова заповедников // Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России. М.: Науч. мир, 2000. С. 30–45.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 197 с.
Черешнев И.А. Биогеография пресноводных рыб Дальнего Востока. Владивосток: Дальнаука, 1998. 131 с.
Шварц С.С. Популяционная структура вида // Зоол. журн. 1967. Т. 46. С. 1456–1469.
Яблоков А.В. Популяционная биология. М.: Высш. шк., 1987. 303 с.
Allendorf F.W., Luikart G.H., Aitken S.N. Conservation and the genetics of populations. 2nd ed. West Sussex, UK: Wiley-Blackwell, 2012. 664 p.
COSEWIC. Guidelines for recognizing designatable units. 2015. URL: http://www.cosewic.gc.ca/eng/sct2/sct2_5_e.cfm
Dionne M., Caron F., Dodson J.J., Bernatchez L. Landscape genetics and hierarchical genetic structure in Atlantic salmon: the interaction of gene flow and local adaptation // Mol. Ecol. 2008. V. 17. P. 2382–2396.
Fraser D.J., Weir L.K., Bernatchez L., Hansen M.M., Taylor E.B. Extent and scale of local adaptation in salmonid fishes: review and meta-analysis // Heredity. 2011. V. 106. P. 404–420.
Funk W.C., McKay J.K., Hohenlohe P.A., Allendorf F.W. Harnessing genomics for delineating conservation units // Trends Ecol. Evol. 2012. V. 27. P. 489–496.
Gagnaire P.A., Normandeau E., Côté C., Hansen M.M., Bernatchez L. The genetic consequences of spatially varying selection in the panmictic American eel (Anguilla rostrata) // Genetics. 2012. V. 190. P. 725–736.
Manel S., Joost S., Epperson B.K., Holderegger R., Storfer A., Rosenberg M.S., Scribner K.T., Bonin A., Fortin M.J. Perspectives on the use of landscape genetics to detect genetic adaptive variation in the field // Mol. Ecol. 2010. V. 19. P. 3760–3772.
Moritz C. Strategies to protect biological diversity and the evolutionary processes that sustain it // Syst. Biol. 2002. V. 51. P. 238–254.
Pearse D.E., Miller M.R., Abadía-Cardoso A., Garza J.C. Rapid parallel evolution of standing variation in a single, complex, genomic region is associated with life history in steelhead/rainbow trout // Proc. Roy. Soc. London, Biol. Sci. 2014. V. 281. № 1783. ID 20140012.
Perrier C., Bourret V., Kent M.P., Bernatchez L. Parallel and nonparallel genome-wide divergence among replicate population pairs of freshwater and anadromous Atlantic salmon // Mol. Ecol. 2013. V. 22. P. 5577–5593.
Quantum GIS. 2017. URL: http://www.gisenglish.com/2017/12/ download-free-qgis-21815-for-windows.html
Sork V.L., Waits L. Contributions of landscape genetics approaches, insights, and future potential // Mol. Ecol. 2010. V. 19. P. 3489–3495.
Teramura A.H. Experimental ecological genetics in Plantago IX. Differences in growth and vegetative reproduction in Plantago lanceolata L. (Plantaginaceae) from adjacent habitat // Am. J. Bot. 1983. V. 70. № 1. P. 53–58.
Turkington R., Harper J.L. The growth distribution and neighubur relationships of Trifolium repens a permanent pasture. IV. Finescale biotic differentiation // J. Ecol. 1979. V. 67. P. 245–254.
Waples R.S. Pacific salmon, Oncorhynchus spp., and the definition of “species” under the Endangered Species Act. // Mar. Fish Rev. 1991. V. 53. P. 11–22.
Waples R.S. Distinct population segments // The Endangered Species Act at Thirty. V. 2. Conserving biodiversity in human-dominated landscapes. Washington, D.C.: Island Press, 2006. P. 127–149.
Waples R.S., Gaggiotti O. What is a population? An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity // Mol. Ecol. 2006. V. 15. P. 1419–1439.
Waples R.S., Gustafson R.G., Weitkamp L.A., Myers J., Johnson O., Busby P., Hard J., Bryant G., Waknitz F., Neely K., Teel D., Grant W., Winans G., Phelps S., Marshall A., Baker B. Characterizing diversity in Pacific salmon // J. Fish Biol. 2001. V. 59 (suppl. A). P. 1–41.
Yang J., Weedon M.N., Purcell S., Lettre G., Estrada K., Willer C.J., Smith A.V., Ingelsson E., O’Connell J.R., Mangino M., Mägi R., Madden P.A., Heath A.C., Nyholt D.R., Martin N.G., Montgomery G.W., Frayling T.M., Hirschhorn J.N., McCarthy M.I., Goddard M.E., Visscher P.M. GIANT Consortium. Genomic inflation factors under polygenic inheritance // Eur. J. Hum. Genet. 2011. V. 19. P. 807–812.
Zhivotovsky L.A., Yurchenko A.A., Nikitin V.D., Safronov S.N., Shitova M.V., Zolotukhin S.F., Makeev S.S., Weiss S., Rand P.S., Semenchenko A.Yu. Eco-geographic units, population hierarchy, and a two-level conservation strategy with reference to a critically end angered salmonid, Sakhalin taimen Parahucho perryi // Conserv. Genet. 2015. V. 16. P. 431–441.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая