

Известия РАН. Серия биологическая, 2020, № 3, стр. 310-317
Соматический и репродуктивный рост приморского гребешка в загрязненном Амурском заливе
А. В. Силина *
Национальный научный центр биологии моря ДВО РАН
690041 Владивосток, ул. Пальчевского, 17, Россия
* E-mail: allasilina@mail.ru
Поступила в редакцию 24.01.2018
После доработки 14.03.2018
Принята к публикации 15.03.2018
Аннотация
Исследована репродуктивная активность приморского гребешка Mizuhopecten yessoensis (Jay, 1857) в загрязненном Амурском зал. (зал. Петра Великого Японского моря) с учетом сезонного роста его гонады, мускула-аддуктора и пищеварительной железы. Установлено, что нерест гребешка в загрязненном заливе приводит к потере массы не только гонады, но и мускула-аддуктора, а также пищеварительной железы, откуда следует, что энергия, требуемая для гаметогенеза, обеспечивается, в частности, энергетическими резервами этих соматических тканей. Обнаружено, что в годы наиболее сильного загрязнения акватории при первом гаметогенезе гребешка снижение массы его органов незначительное в период нереста, что свидетельствует о неполноценном участии впервые нерестящихся двухлетних особей в процессе воспроизводства популяции. Отмечено, что при снижении уровня загрязнения среды гребешки этого же возраста значимо теряют массы гонады и соматических тканей при нересте, т.е. происходит полноценный нерест. Найдено, что в период половозрелости приросты массы соматических тканей снижаются с увеличением возраста, а репродуктивная активность повышается, позднее снижаясь в старческой стадии развития гребешка.
В водах умеренных широт морские двустворчатые моллюски могут ассимилировать из окружающей среды только сезонно ограниченное количество энергии, поэтому соматический и репродуктивный рост обычно разделяются во времени так, чтобы максимизировать репродуктивную продукцию (MacDonald, Thompson, 1985, 1986; Strohmeier et al., 2000). Различные способы, с помощью которых энергия приобретается, сохраняется и распределяется для этих процессов, варьируют между видами и даже между популяциями одного вида (Epp et al., 1988; Ramirez-Llodra, 2002; Saucedo, Southgate, 2008). Например, развитие гонады у устрицы Pinctada mazatlanica в Калифорнийском зал. осуществляется в основном за счет запасов энергии, хранящихся в течение зимы, во-первых, в мускуле аддукторе и, во-вторых, в пищеварительной железе (Vite-Garcıa, Saucedo, 2008). В то же время устрица Pteria sterna из того же залива вместо резервов ткани использует доступную пищу для развития гонады зимой и весной, а в течение позднего лета большое количество энергии для этого процесса мобилизуется, во-первых, из пищеварительной железы и, во-вторых, из мускула-аддуктора (Vite-Garcıa, Saucedo, 2008). На практике каждый вид использует различные комбинации стратегий, чтобы управлять доступной энергией и регулировать гаметогенез.
Созревание гонады и нерест приводят к мобилизации макромолекулярных резервов из соматических тканей у многих видов Pectinidae. У гребешков только мускул-аддуктор, пищеварительная железа и гонада рассматриваются как органы запаса энергоемких веществ (Barber, Blake, 2006). Уменьшение массы мускула-аддуктора во время гаметогенеза было выявлено у нескольких видов гребешков, таких как Chlamys septemradiata, C. opercularis, Placopecten magelanicus, Argopecten irradians concentricus, A. irradians irradians и A. purpuratus (Ansell, 1974; Taylor, Venn, 1979; Barber, Blake, 1981; Robinson et al., 1981; Epp et al., 1988; Martinez, Mettifogo, 1998). При этом предполагалось, что в мускуле-аддукторе запасаются вещества, обеспечивающие энергией процесс гаметогенеза, главным образом гликоген (Martinez, Mettifogo, 1998). В то же время в условиях обильного обеспечения пищей, масса мускула-аддуктора не уменьшается во время гаметогенеза P. magelanicus (Paon, Kenchington, 1995). Известно, что пищеварительная железа двустворчатых моллюсков играет существенную роль в накоплении липидов и меньшую в накоплении гликогена (Chang et al., 1989; Caers et al., 1999; Strohmeier et al., 2000; Rodriguez-Astudillos et al., 2005). Эти субстанции составляют резервы энергии, особенно для гаметогенеза, а также роста раковины (Ansell, 1974; Taylor, Venn, 1979; Barber, Blake, 1981; Robinson et al., 1981; Caers et al., 1999; Strohmeier et al., 2000; Le Pennec et al., 2001). Разнообразие данных о важности различных органов Pectinidae как места запаса энергоемких веществ и их пригодность в обеспечении гаметогенеза объясняют способностью популяций адаптировать способы хранения энергии и утилизационную стратегию к местным условиям (Barber, Blake, 2006).
Несмотря на явную сезонность всех важных событий в жизненном цикле коммерчески важного приморского гребешка Mizuhopecten yessoensis (Jay, 1857), нерестящегося 1 раз в год, результатов сравнения темпов роста его соматических и генеративных тканей в разные этапы гаметогенеза в литературе крайне мало. Ранее исследование такого рода было проведено только для молодых культивируемых гребешков из теплых вод о. Хонсю в Японии (Takahashi, Mori, 1971). Сравнительное исследование роста различных органов гребешков разного возраста, обитающих в естественных популяциях, ранее было затруднено невозможностью определения возраста каждой особи. В предлагаемой работе было запланировано изучение динамики темпов роста общей массы и массы отдельных органов разновозрастных гребешков из естественной популяции при различных стадиях развития гонад в условиях разной степени загрязнения окружающей среды. Эти исследования были предприняты для определения, существует ли энергетическое инвестирование в продукцию гонады и нерест у приморского гребешка из соматических органов для обеспечения его гаметогенеза.
МАТЕРИАЛЫ И МЕТОДЫ
Район исследований расположен у восточных берегов Амурского зал. (зал. Петра Великого) у Владивостока в районе устья Первой Речки. Этот район характеризуется химическим и органическим загрязнением воды и донных отложений, так как находится у выхода коллектора промышленных и бытовых сточных вод города (Tkalin et al., 1993). По экспертным оценкам вместе со сточными водами в течение, например, 2000 г. поступило ~44 643 т загрязняющих веществ 28 наименований, включая органические загрязнители, тяжелые металлы, пестициды и нефтеуглеводороды (Нигматулина, 2005). Кроме того, данная акватория попадает под воздействие стока р. Раздольной, несущей загрязнители, поступающие из городов Китая, г. Уссурийска, поселков и с сельхозугодий бассейна реки (Звалинский и др., 2012). До начала 1990-х гг. наблюдалось усиление загрязнения залива вследствие развития хозяйственной деятельности в прибрежье залива и реки. Позднее из-за значительного снижения промышленного производства и сельскохозяйственной деятельности в России загрязнение прибрежных акваторий снизилось, состояние бентосного сообщества улучшилось (Лукьянова и др., 2012; Силина, 2019). Сравнительные исследования соматического и репродуктивного роста гребешка были проведены для этих двух периодов развития биотопа. Сбор гребешка проводили водолазы в разные сезоны 1994–1995 и 2015–2016 гг. с заиленного грунта на глубине 6–8 м.
Динамика масс соматических тканей, мускула-аддуктора, пищеварительной железы и гонады приморского гребешка была исследована в зависимости от стадии развития его гонады в течение годового цикла гаметогенеза. Основные стадии развития гонады гребешка в зал. Петра Великого были выделены несколькими авторами, причем число стадий не всегда совпадало. Мной выбраны основные стадии, упомянутые в работах большинства авторов, и названы согласно классификации Дзюба (Дзюба, 1986). Гребешки были отобраны 01.06.1994, т.е., согласно С.М. Дзюба, в нерестовую стадию для исследованной акватории (70 экз.). Далее 26.08.1994 (стадия репродуктивной инертности, покоя, 64 экз.), 30.11.1994 (стадия начала развития гонад, 80 экз.) и 01.03.1995 (стадия активного гаметогенеза, 36 экз.). Через 20 лет гребешки были собраны в те же периоды: 22.06.2015 (нерестовая стадия, 24 экз.), 25.08.2015 (стадия покоя, 44 экз.), 01.12.2015 (начало развития гонад, 29 экз.) и 13.05.2016 (активный гаметогенез, 25 экз.). Чтобы избежать преждевременного нереста, гребешки были вскрыты и взвешены непосредственно после отлова. Вначале отделяли мягкие ткани от раковины, разделяли и взвешивали гонаду и соматические ткани с точностью 0.1 г на весах BL-620S (Shimadzu, Япония). Затем раздельно взвешивали мускул-аддуктор и пищеварительную железу. Известно, что приморский гребешок – раздельнополый моллюск и параметры для одновозрастных самцов и самок из одной и той же популяции могут несколько различаться (Silina, 2016). В то же время известно, что соотношение полов в популяции близко к 1 : 1, поэтому полученные данные усреднялись без разделения по половому признаку.
Возраст каждой особи был определен по ранее разработанной методике (Silina, 1996). Этот метод основан на способности приморского гребешка формировать микрослои, визуально различимые в микроскульптуре внешней поверхности его верхней створки. Внешний вид и ширина образованных в разные сезоны микрослоев различаются (Silina, 1996). По числу участков раковины, образованных летом, т.е. годовых колец, определяли возраст каждого гребешка.
Статистический анализ состоял из сравнения средних значений изучаемых параметров одновозрастных гребешков, отловленных на разных стадиях гаметогенеза, по критерию Стьюдента (t-test) с использованием программного обеспечения Statistica (Statsoft). Этот же метод был использован при сравнении параметров гребешков одного возраста, отловленных в разные годы. До статистического анализа все сравниваемые данные были протестированы на нормальность распределения по критерию Колмогорова–Смирнова. Были установлены уровни значимости P < 0.01 и P < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Рост гребешка в период интенсивного загрязнения среды. Установлено, что в 1994–1995 гг. каждая проба гребешков состояла из 2–9-летних особей. При сравнении массы тканей гребешка у разновозрастных особей установлено, что массы как соматических, так и репродуктивных тканей увеличивались в течение всей жизни гребешка, даже в неблагоприятных условиях загрязненной среды в 1994–1995 гг. (рис. 1, 2). В течение года их приросты были неравномерны (рис. 1). Наиболее значимые колебания масс гонады, а также соматических тканей и их составляющих могут быть по крайней мере частично связаны с определенными фазами репродуктивного цикла (рис. 1). В первую очередь обращает на себя внимание ярко выраженное, статистически значимое (P < 0.01) снижение показателей массы после нереста к периоду наступления репродуктивного покоя, за исключением генерации двухлеток (рис. 1). В это время в отличие от других возрастных классов для двухлетних особей характерен не отрицательный, а положительный прирост массы соматических тканей, а масса гонады снизилась незначительно (рис. 1). После нерестового периода гонады двухлеток, как и у старших особей, потеряли окраску, характерную для соответствующих полов, они стали полупрозрачными. Этот факт свидетельствует, скорее всего, о малопродуктивном нересте двухлеток в данном районе. По сравнению со средневозрастными особями, после нереста наблюдалось менее значимое в относительных показателях снижение массы гонады и у старой возрастной группы 7–9-леток (рис. 1г), т.е. нерест старых особей был сравнительно менее продуктивным в исследованном загрязненном районе. Однако и у этой группы гребешков после нереста значимо (P < 0.05) снизились массы мускула-аддуктора и пищеварительной железы (рис. 1б, г).
Рис. 1.
Сезонная динамика средних значений масс соматических тканей (а), мускула-аддуктора (б), гонады (в) и пищеварительной железы (г) приморского гребешка Mizuhopecten yessoensis в возрасте 2, 3, 4, 5, 6 и 7–9 лет (1–6 соответственно) в 1994–1995 гг. в северо-восточной части Амурского зал. Вертикальные линии – ошибки средних значений параметров; для рис. 1–4.
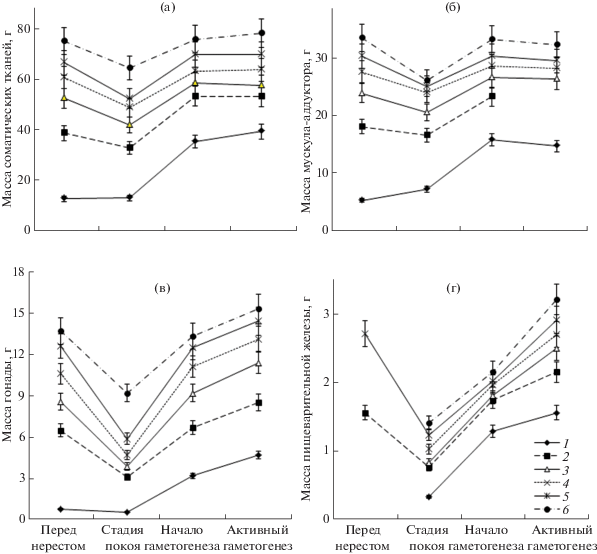
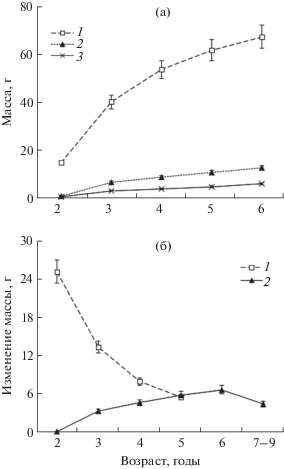
Рис. 2.
Рост масс соматических тканей (1), гонады перед нерестом (2) и гонады в стадии покоя (3) (а), годовые приросты массы соматических тканей (1) и годовая репродуктивная продукция (2) (б) приморского гребешка Mizuhopecten yessoensis в 1994–1995 гг. в северо-восточной части Амурского зал.
Наибольший за весь цикл гаметогенеза статистически значимый (P < 0.01) прирост масс соматических тканей и органов гребешка наблюдался в период начала гаметогенеза (рис. 1). Позднее, в период активного гаметогенеза, при низкой температуре воды, с конца ноября до конца марта, в массе мускула-аддуктора и в целом соматических тканей прироста не произошло, для массы мускула наблюдалось даже заметное снижение этого показателя (рис. 1а, б). Напротив, в это время гонада заметно развивалась у всех возрастных групп, для 2-летних (P < 0.01) и 3–5-летних (P < 0.05) особей статистически значимо (рис. 1в). Значимо (P < < 0.01–0.05) у особей старше двух лет в период активного гаметогенеза увеличилась и масса пищеварительной железы, но у двухлеток она практически не изменилась (рис. 1г).
В данных условиях среды массы соматических тканей и гонады увеличивались в течение жизни гребешка (рис. 2а), но скорости изменения массы соматических тканей и репродуктивного выхода имели противоположные направленности (рис. 2б). С наступлением половозрелости годовые приросты массы соматических тканей год от года значимо (P < 0.01) снижались, а репродуктивная продукция повышалась вплоть до 6-летнего возраста гребешка (рис. 2б). Позднее у 7–9-летних особей, фактически в старческой для исследованного района стадии онтогенеза гребешка, статистически значимо (P < 0.01) снизилась и репродуктивная активность (рис. 2б).
Рост гребешка при снижении уровня загрязнения среды. В 2015–2016 гг. каждая проба гребешков состояла из 2–4-летних особей. Только 2 раза были встречены 5-летние гребешки, по одной особи в пробе. Старых особей в этот период наблюдения не было найдено, поэтому не удалось изучить особенности изменения темпов роста тканей у старых гребешков при более благоприятных условиях среды их обитания. При сравнительно сниженной степени загрязнения исследуемой акватории в 2015–2016 гг. у особей всех возрастов статистически значимо (P < 0.01) повысились как массы соматических тканей и их составляющих, так и масса гонады по сравнению с аналогичными параметрами гребешка в 1994–1995 гг. (рис. 1 и 3).
Рис. 3.
Сезонная динамика средних значений масс соматических тканей (а), мускула-аддуктора (б), гонады (в) и пищеварительной железы (г) приморского гребешка Mizuhopecten yessoensis в возрасте 2, 3 и 4 лет (1–3 соответственно) в 2015–2016 гг. в северо-восточной части Амурского зал.
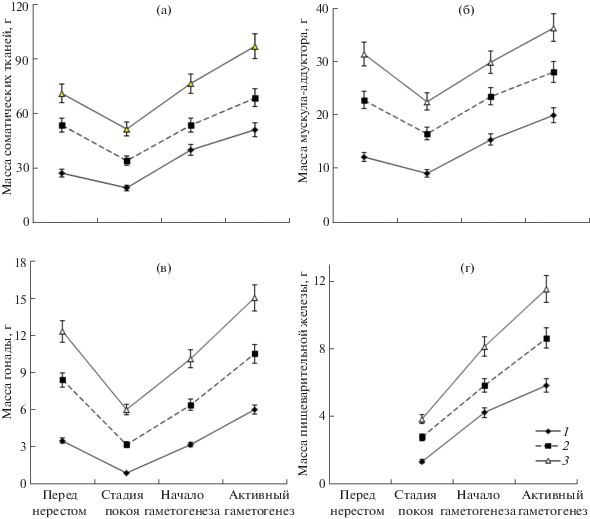
В целом характер сезонного изменения массы тканей остался прежним. Также обращает на себя внимание снижение масс всех органов после нереста гребешка. В то же время имеется существенное отличие от результатов наблюдения в 1994–1995 гг., выраженное в характере изменения средней массы гонады у генерации 2-летних особей. После нереста в 2015 г. масса их гонады значимо (P < 0.01) снизилась, в среднем в 4 раза, что, по-видимому, объясняется более продуктивным нерестом в этой возрастной категории в улучшенной по качеству среде по сравнению с нерестом 2‑леток в 1994 г.
При меньшем загрязнении среды в 2015–2016 гг. наблюдалось значительное увеличение масс соматических тканей и их составляющих не только в период начала гаметогенеза, но и в период активного гаметогенеза при низкой температуре воды (рис. 3). Проба в 2016 г. была взята во второй, а не в первой половине этой стадии, как в 1995 г., однако при сильном загрязнении среды в 1994–1995 гг. высоких темпов роста органов не наблюдалось и после активной стадии, перед нерестом (рис. 1). Поэтому в период сильного загрязнения заметного прироста, сравнимого с таковым в 2015–2016 гг., нельзя было ожидать и для второй половины стадии активного гаметогенеза. В отличие от 1995 г. в 2016 г. в период активного гаметогенеза масса пищеварительной железы значимо повысилась и у 2-леток (рис. 1г и 3г).
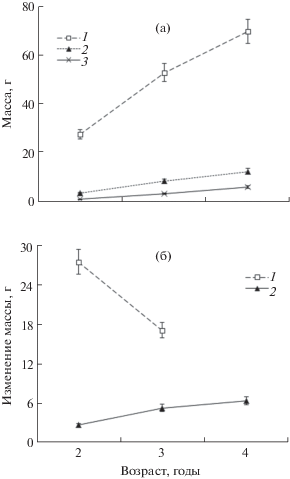
В 2015–2016 гг. абсолютные значения и годовые приросты масс соматических и репродуктивных тканей гребешка были значимо (P < 0.01) выше, чем в 1994–1995 гг. (рис. 2 и 4). Как и в предыдущие годы, с возрастом прирост массы соматических тканей снижался, а репродуктивная продукция росла по крайней мере до 4-летнего возраста (рис. 4б), притом более высокими темпами, чем 20 лет назад (рис. 2б и 4б).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Установлено, что в течение года темпы как соматического, так и репродуктивного роста приморского гребешка изменяются значительно. Главный период роста соматических тканей гребешка в Амурском зал. происходит в период начала гаметогенеза, а при более благоприятных условиях – и в период активного гаметогенеза моллюска. В эти этапы гаметогенеза наблюдаются также значительные приросты репродуктивных тканей гребешка. Противоположные тенденции в приросте тканей гребешка наблюдаются после нереста, когда массы соматических и репродуктивных тканей значимо снижаются. Можно сделать вывод, что колебания темпов роста гребешка и его различных органов определяются, в частности, таким эндогенным фактором, как репродуктивный цикл. Известно, что и интенсивность метаболизма приморского гребешка также зависит от стадии репродуктивного цикла (Седова, Викторовская, 2000).
Нерест гребешка в заливе приводит к большой потере массы и, соответственно, энергии не только в гонадах, но и в соматических тканях. Резерв гликогена мускула-аддуктора традиционно считается источником запасной энергии, доступным двустворчатому моллюску в период стресса, в частности, для такого энергозатратного процесса, как гаметогенез (Martinez, Mettifogo, 1998). Позже было показано, что липиды пищеварительной железы также могут быть источником запасенной энергии, особенно когда резерв мускула-аддуктора истощен (Le Pennec et al., 2001). Действительно, эксперименты с радиоэлементами с устрицей Crassostrea gigas показали существование гемолимфатического переноса липидов между органами моллюска посредством гемоцитов и плазмы (Allen, Conley, 1982; Pollero, Heras, 1989). Передача липидов от пищеварительной железы к созревающей гонаде самки была обнаружена и у гребешков, например у Argopecten irradians eoneentrieus и Chlamys hericia (Vassallo, 1973; Barber, Blake, 1985). Мной выявлено уменьшение массы как мускула-аддуктора, так и пищеварительной железы приморского гребешка в период нереста. Это может свидетельствовать о том, что запасов энергии только в одном из этих органов недостаточно для успешной репродукции вида в исследованном неблагоприятном для гребешка районе (Силина, Овсянникова, 1995; Силина, 2019). Таким образом, энергетические запасы организма перераспределяются в пользу репродуктивных усилий.
Наблюдения других авторов за ростом 1–3-летних гребешков в условиях его культивирования в теплых водах Японии показали, что в нерестовый период масса пищеварительной железы снижается, а масса мускула продолжает расти (Takahashi, Mori, 1971; Fuji, Hashizume, 1974). Первый результат соответствует, а последний факт противоположен данным, полученным для гребешков из Амурского зал. По-видимому, для исследуемого вида моллюсков характерно первоочередное использование в стрессовых ситуациях, например при нересте, энергетических запасов, накопленных в пищеварительной железе, а в условиях хронического стресса, при обитании в неблагоприятных условиях, для обеспечения воспроизводства популяции дополнительно используются запасы энергии и мускула-аддуктора. У устрицы P. sterna большое количество энергии для развития гонады может мобилизоваться, во-первых, из пищеварительной железы и, во-вторых, из мускула-аддуктора (Vite-Garcıa, Saucedo, 2008).
При сравнении изменений масс разных органов приморского гребешка в течение его репродуктивного цикла в одном и том же биотопе, но при разной степени загрязнения среды обращает на себя внимание значимая разница в показателях массы как соматических, так и репродуктивных тканей. Кроме высоких концентраций тяжелых металлов, пестицидов и нефтеуглеводородов в воде и донных осадках (Tkalin et al., 1993), ухудшающих состояние бентосных организмов, обитающих в Амурском зал. (Силина, 2018), важным фактором среды для приморского гребешка является низкая концентрация кислорода в придонном слое воды. Известно, что в заливе она может снижаться вплоть до критических значений 2–3 мл/л и 50–60%-ного насыщения (Рачков, 2002). Концентрация кислорода <6 мл/л оказывает неблагоприятное воздействие на приморский гребешок (Yamamoto, 1957). Понижение концентрации кислорода в воде происходит из-за его потребления микроорганизмами при разложении органических веществ во взвеси и донных осадках, поступающих туда с промышленными, сельскохозяйственными и бытовыми отходами (Нигматулина, 2005; Звалинский и др., 2012). Очевидно, что при более высокой степени загрязнения залива органическими и другими веществами в 1994–1995 гг. (Лукьянова и др., 2012) гребешку наносился больший урон, чем в 2015–2016 гг., что и отразилось на темпах его роста.
В условиях большего загрязнения акватории, в период активного гаметогенеза гребешка, отчетливо выражен преимущественный рост гонады, а не соматических тканей. Таким образом, полученные данные указывают на то, что в неблагоприятных условиях среды биоэнергетическая стратегия изучаемого вида направлена на первоочередное распределение доступных питательных веществ в гонады для обеспечения успешного воспроизводства популяции. Аналогичные результаты получены при исследовании роста и других видов моллюсков (MacDonald, Thompson, 1986).
Другое особенно заметное отличие выражено в темпах роста масс органов 2-летних особей. В условиях сильного загрязнения у молодых особей, нерестящихся впервые, практически не наблюдается снижения массы их органов после нереста. Даже, напротив, масса мускула несколько прирастает в это время. А в период начала гаметогенеза масса репродуктивного органа 2-леток прирастает не так интенсивно, как масса соматических тканей. Таким образом, в неблагоприятной среде обитания особенно страдает репродуктивная функция молодых особей. При снижении уровня загрязнения сезонные изменения масс гонады и соматических тканей особей, нерестящихся в первый раз, происходят практически так же, как и у средневозрастных гребешков.
Онтогенез вносит свои коррективы в изменения массы органов гребешка в течение его репродуктивного цикла. С наступлением половозрелости прирост массы соматических тканей снижается, а репродуктивная продукция (масса гонады) повышается, вплоть до старческой для исследованного района стадии онтогенеза, т.е. идет перераспределение энергии для первоочередного обеспечения репродуктивного успеха популяции. В старческом возрасте снижается не только скорость роста соматических тканей гребешка, но и его репродуктивная активность.
Список литературы
Дзюба С.M. Половая система и гаметогенез // Приморский гребешок. Владивосток: Дальнаука, 1986. Гл. 10. С. 118–130.
Звалинский В.И., Тищенко П.И., Михайлик Т.А., Тищенко П.Я. Эвтрофикация Амурского залива // Современное экологическое состояние залива Петра Великого Японского моря / Ред. Христофорова Н.К. Владивосток: Изд. дом ДВФУ, 2012. С. 76–114.
Лукьянова О.Н., Черкашин С.А., Симоконь М.В. Обзор современного экологического состояния залива Петра Великого (2000–2010 гг.) // Вест. ДВО РАН. 2012. № 2. С. 55–63.
Нигматулина Л.В. Воздействие сточных вод контролируемых выпусков на экологическое состояние Амурского залива (Японское море): Автореф. дис. канд. биол. наук. Владивосток: ТИНРО, 2005. 19 с.
Рачков В.И. Характеристика гидрохимических условий вод Амурского залива в теплый период года // Изв. ТИНРО. 2002. Т. 131. С. 65–77.
Седова Л.Г., Викторовская Г.И. Зависимость интенсивности обмена от репродуктивной активности морского гребешка Mizuhopecten yessoensis (Jay) // Изв. ТИНРО. 2000. Т. 127. С. 469–474.
Силина А.В. Популяция приморского гребешка при климатических и антропогенных изменениях среды в Амурском заливе Японского моря // Океанология. 2019. Т. 59. № 1. С. 82–92.
Силина А.В., Овсянникова И.И. Многолетние изменения в сообществе приморского гребешка и его эпибионтов в загрязненной части Амурского залива Японского моря // Биол. моря. 1995. № 1. С. 59–66.
Allen W.V., Conley H. Transport of lipids in the blood of the Pacific oyster, Crassostrea gigas (Thunberg) // Comp. Biochem. Physiol. 1982. V. 71B. P. 201–207.
Ansell A.D. Seasonal changes in biochemical composition of bivalve Chlamys septemradiata from Clyde Sea area // Mar. Biol. 1974. V. 25. № 2. P. 85–99.
Barber B.J., Blake N.J. Energy storage and utilization in relation to gametogenesis in Argopecten irradians concentricus (Say) // J. Exp. Mar. Biol. Ecol. 1981. V. 52. P. 121–134.
Barber B.J., Blake N.J. Intra-organ biochemical transformations associated with oogenesis in the bay scallop Argopecten irradians concentricus (Say) as indicated by 14C incorporation // Biol. Bull. 1985. V. 168. P. 39–49.
Barber B.J., Blake N.J. Reproductive physiology. Scallops: biology, ecology, and aquaculture // Develop. Aquacult. Fish. Sci . 2006. V. 35. P. 357–416.
Caers M., Coutteau P., Cure K., Morales V., Gajardo G., Sorgeloos P. The Chilean scallop Argopecten purpuratus (Lamarck, 1819). I. Fatty acid composition and lipid content of six organs // Comp. Biochem. Physiol. 1999. V. 123B. P. 89–96.
Chang Y.J., Sagawara Y., Nomura T. Structure and function of digestive diverticula in the scallop, Patinopecten yessoensis (Jay) // Tohoku J. Agr. Res. 1989. V. 39. P. 81–93.
Epp 1., Bricelj M., Malouf R.E. Seasonal partitioning and utilization of energy reserves in two age classes of the bay scallop Argopecten irradians irradians (Lamarck) // J. Exp. Mar. Biol. Ecol. 1988. V. 121. P. 113–136.
Fuji A., Hashizume M. Energy budget for a Japanese common scallop, Patinopecten yessoensis (Jay), in Mutsu Bay // Bull. Fac. Fish. Hokkaido Univ. 1974. V. 25. P. 7–19.
Le Pennec G., Le Pennec M., Beninger P.G. Seasonal digestive gland dynamics of the scallop Pecten maximus in the Bay of Brest (France) // J. Mar. Biol. Ass. UK. 2001. V. 81. P. 663–671.
MacDonald B.A., Thompson R.J. Influence of temperature and food availability on the ecological energetics of the giant scallop Placopecten magellanicus. II. Reproductive output and total production // Mar. Ecol. Prog. Ser. 1985. V. 25. P. 295–303.
MacDonald B.A., Thompson R.J. Influence of temperature and food availability on the ecological energetics of the giant scallop Placopecten magellanicus. III. Physiological ecology, the gametogenetic cycle and scope for growth // Mar. Biol. 1986. V. 93. P. 37–48.
Martinez G., Mettifogo L. Mobilization of energy from adductor muscle for gametogenesis of the scallop, Argopecten purpuratus Lamarck // J. Shellfish. Res. 1998. V. 17. P. 113–116.
Paon L.A., Kenchington E.L.R. Changes in somatic and reproductive tissues during artificial conditioning of the sea scallop, Placopecten magelanicus (Gmelin, 1791) // J. Shellfish. Res. 1995. V. 14. P. 53–58.
Pollero R.J., Heras H. Intra-organ hemolymphatic transport of free fatty acids, triacylglycerols and phospholipids in the freshwater bivalve Diplodon delodontus // Comp. Biochem. Physiol. 1989. V. 93A. P. 673–676.
Ramırez-Llodra E. Fecundity and life-history strategies in marine invertebrates // Adv. Mar. Biol. 2002. V. 43. P. 87–170.
Robinson W.E., Wehling W.E., Morse M.P., McLeod G.C. Seasonal changes in soft-body component indices and energy reserves in the Atlantic deep-sea scallop, Placopecten magelanicus // Fish. Bull. 1981. V. 79. P. 449–458.
Rodriguez-Astudillos S., Villalejo-Fuerte M., Garcia-Dominguez F., Guerrero-Caballero R. Biochemical composition and its relationship with the gonadal index of the black oyster Hyotissa hyotis (Linnaeus, 1758) at Espiritu Santo Gulf of California // J. Shellfish. Res. 2005. V. 24. P. 975–978.
Saucedo P., Southgate P. Reproduction, development and growth // Pearl oyster. Amsterdam: Elsevier, 2008. P. 131–186.
Silina A.V. Mortality of late juvenile and adult stages of the scallop Mizuhopecten yessoensis (Jay) // Aquaculture. 1996. V. 141. P. 97–105.
Silina A.V. Is sexual size dimorphism inherent in scallop Patinopecten yessoensis? // Scientifica. 2016. V. 2016. Art. ID 8653621.
Strohmeier T., Duinker A., Lie O. Seasonal variations in chemical composition of the female gonad and storage organs in Pecten maximus (L.) suggesting that somatic and reproductive growth are separated in time // J. Shellfish. Res. 2000. V. 19. P. 741–747.
Takahashi K., Mori K. Seasonal variation in the methabolism of lipids and glycogen in the scallop, Patinopecten yessoensis (Jay). I. Biochemical studies // Tohoku J. Agricult. Res. 1971. V. 22. № 2. P. 114–124.
Taylor A.C., Venn T.J. Seasonal variation in weight and biochemical composition of the tissues of the queen scallop, Chlamys opercularis, from the Clyde Sea area // J. Mar. Biol. Ass. UK. 1979. V. 59. № 3. P. 605–621.
Tkalin A.V., Belan T.A., Shapovalov E.N. The state of the marine environment near Vladivostok, Russia // Mar. Pollut. Bull. 1993. V. 26. № 8. P. 418–422.
Vassallo M.T. Lipid storage and transfer in the scallop Chlamys hericia Gould // Comp. Biochem. Physiol. 1973. V. 44A. P. 1169–1175.
Vite-Garcia M.N., Saucedo P.E. Energy storage and allocation during reproduction of pearl oyster Pteria sterna (Gould, 1851) at Bahia de La Paz, Baja California Sur, Mexico // J. Shellfish Res. 2008. V. 27. P. 375–383.
Yamamoto G. Tolerance of scallop spats to suspended silt, low oxygen tention, high and low salinity and sudden temperature changes // Sci. Rep. Tohoku Univ. Ser. 4. Biol. 1957. № 23. P. 73–82.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая