

Известия РАН. Серия биологическая, 2020, № 4, стр. 423-433
Эмиссия СО2 с поверхности ствола сосны обыкновенной в разных условиях произрастания
А. Г. Молчанов 1, *, А. В. Ольчев 2, 3
1 Институт лесоведения РАН
143030 Московская обл., с. Успенское, ул. Советская, 21, Россия
2 МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы 1, Россия
3 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: a.georgievich@gmail.com
Поступила в редакцию 06.02.2018
После доработки 04.04.2018
Принята к публикации 04.04.2018
Аннотация
Представлены результаты экспериментальных исследований эмиссии СО2 с поверхности стволов сосны обыкновенной Pinus sylvestris L. в заболоченном пушицево-сфагновом сосняке в Ярославской обл. и в высокопродуктивном разнотравно-черничном сосняке на хорошо дренированных супесчаных почвах Серебряноборского лесничества Московской обл. Исследования проведены в течение нескольких вегетационных периодов методом экспозиционных камер при разных погодных условиях и глубинах залегания грунтовых вод. Количественно описана сезонная и внутрисуточная изменчивость эмиссии СО2 с поверхности стволов живых деревьев в лесах южно-европейской тайги и выявлена зависимость потоков СО2 от факторов внешней среды (температуры воздуха и уровня грунтовых вод). Для заболоченного сосняка обнаружены различия в дыхании стволов деревьев разного класса роста.
Современные изменения климата и наблюдаемый устойчивый рост глобальной температуры большинство исследователей связывают с увеличением содержания в атмосфере парниковых газов, и прежде всего диоксида углерода (СО2) (IPCC, 2013). Наземные экосистемы, и в частности леса, не только поглощают и аккумулируют СО2 из атмосферы, но и являются его активным источником (Luyssaert et al., 2007). Эмиссия СО2 с поверхности растительного и почвенного покрова определяется совокупностью биотических и абиотических факторов и изменяется в широких пределах в зависимости от типа растительных сообществ и условий произрастания (Malhi et al., 1999; Falge et al., 2002; Ibrom et al., 2006; Алферов и др., 2017; Замолодчиков и др., 2017; и др.).
Принято считать, что в лесных экосистемах основным источником естественного поступления CO2 в атмосферу служит эмиссия СО2 с поверхности почвы, включающая в себя дыхание почвенной биоты и корней растений, а также дыхание надземной фотосинтезирующей фитомассы (Lavigne et al., 1997; Law et al., 1999; Malhi et al., 1999). Существенный вклад в суммарное дыхание экосистемы может оказывать и эмиссия СО2 от разложения валежа (Молчанов и др., 2011; Сафонов и др., 2012). Вклад дыхания нефотосинтезирующих элементов живых деревьев в общую эмиссию надземной части фитомассы лесов во многих исследованиях довольно часто игнорируется, хотя может составлять от 5 до 22% (Цельникер и др., 1993; Zha et al., 2004). В качестве основных факторов, влияющих на скорость эмиссии СО2 с поверхности стволов живых деревьев, рассматриваются температура камбия и воздуха в подкроновом пространстве (Linder, Trong, 1981; Goulden et al., 1996). Также отмечается, что существенное влияние на дыхание стволов деревьев могут оказывать условия их произрастания (радиационный режим, структура почвенного покрова, глубина залегания грунтовых вод) и скорость прироста биомассы (Молчанов и др., 2011). Так, на основе анализа динамики роста сосны в разных условиях произрастания Эдвардсом и Золлинсом (Edwards, Sollins, 1973) было показано, что для сосны обыкновенной дыхание стволов деревьев IV класса роста примерно в 4 раза слабее, чем у деревьев I класса роста. Определенные различия в интенсивности эмиссии СО2 с поверхности ствола могут наблюдаться и в разные периоды вегетации. На примере сосны обыкновенной в Сибири была показана устойчивая зависимость скорости дыхания стволов деревьев в летние месяцы от фенофазы роста и развития побегов (Забуга, Забуга, 2013).
Сходные результаты были получены при описании внутрисуточной изменчивости дыхания стволов в сосновых лесах восточной Финляндии (Zha et al., 2004), а также в сосняках центральной части европейской территории России (Цельникер и др., 1993). Было показано, что если осенью скорость дыхания стволов деревьев мало меняется в течение суток и составляет ∼0.3 мкмоль СО2/(м2 · с), то летом различия ее ночных и дневных значений могут быть достаточно существенными, варьируя ночью от 0.8 до 1.1, а днем от 1.0 до 1.7 мкмоль СО2/(м2 · с) (Zha et al., 2004). Таким образом, в среднем за лето интенсивность дыхания стволов сосны достигает ∼1.2 мкмоль СО2/(м2 · с).
Достаточно широкий диапазон изменчивости дыхания нефотосинтезирующих частей древесных растений в различных условиях произрастания, а также существенный недостаток экспериментальных данных о структуре потоков СО2 в лесных экосистемах обуславливают необходимость получения новых данных о масштабах пространственно-временнóй изменчивости дыхания стволов разных видов древесных растений, а также их зависимости от факторов внешней среды. Они могут быть использованы не только для более детального изучения структуры баланса СО2 в лесных экосистемах, но и для интерпретации существующих данных по экосистемным потокам, получаемым с помощью современных мониторинговых систем, в том числе основанных на методе турбулентных пульсаций (eddy covariance) (Goulden et al., 1996; Ibrom et al., 1996; Falge et al., 2002; Luyssaert et al., 2007; Бурба и др., 2016). Кроме того, сохраняется потребность в адекватной параметризации зависимости скорости эмиссии СО2 древесных растений от внешних факторов для дальнейшего использования полученных соотношений в моделях углеродного цикла локального и регионального масштаба (De Pury, Farquhar, 1997; Ольчев и др., 2008, 2017; Kurbatova et al., 2008, 2009; Oltchev et al., 2008; Корзухин, 2015).
Цель исследования – получение новых данных о сезонной и внутрисуточной изменчивости эмиссии СО2 с поверхности стволов сосны обыкновенной Pinus sylvestris L. у деревьев разных классов роста, находящихся в разных условиях произрастания, а также построение модели, описывающей зависимость скорости дыхания стволов сосны от факторов внешней среды.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования временнóй изменчивости дыхания стволов сосны обыкновенной проводились в Серебряноборском опытном лесничестве Института лесоведения РАН в Московской обл. (с мая по октябрь 2011 г. и в мае и ноябре 2012 г.), а также на стационаре “Городище” Северной лесной опытной станции Института лесоведения РАН в Ярославской обл. (в мае и июне 2009 г., а также в июне и августе 2011 г.). В Серебряном Бору измерения проводились в 140-летнем сосняке разнотравно-черничном I бонитета, произрастающем на дерново-слабоподзолистой супесчаной почве при уровне почвенно-грунтовых вод (УПГВ) 2.0−2.5 м. Для измерений были выбраны деревья I класса роста высотой 26−29 м и диаметром 40−45 см.
На стационаре “Городище” исследования проводились в заболоченном сосняке пушицево-сфагновом Vб класса бонитета с мелкозападинным микрорельефом. Мощность торфяной залежи на исследуемом участке составляла ∼1 м. Корневые шейки старых сосен были погружены в торф до глубины 40−60 см. Пушицево-сфагновый сосняк состоял из деревьев двух поколений со средним возрастом 144 и 54 года и со средней сомкнутостью крон ∼30%. Для изучения газообмена СО2 с поверхности ствола на стационаре были выбраны деревья разного класса роста: деревья интенсивного роста (I класса роста) высотой 13 м и диаметром 18 см и деревья слабого роста (IV класса роста) высотой 7 м и диаметром 12 см.
Эмиссия СО2 с поверхности стволов измерялась методом экспозиционных камер по открытой схеме (Edwards, Sollins, 1973; Rayment, Jarvis, 1997; Pumpanen et al., 2004; и др.). Герметичные прямоугольные камеры с площадью основания 200−250 см2 и объемом 140−210 см3 из прозрачной полиэтиленовой пленки устанавливались на выбранных деревьях на высоте ∼1.3 м на северной стороне дерева для защиты камер от нагрева прямыми солнечными лучами (Молчанов, 2010). Крепление камер к деревьям осуществлялось с помощью пластилина толщиной 0.8−1.0 см. Входные и выходные отверстия для соединительных трубок располагались в обрамлении камер с их разных сторон, что обеспечивало равномерное перемешивание воздуха внутри камер. Положение каждой из камер на дереве не менялось в течение всего периода наблюдений. Через камеры в течение всего эксперимента поддерживался постоянный проток окружающего наружного воздуха со средней скоростью 1 л/мин. Скорость потока воздуха через каждую камеру измерялась и корректировалась с помощью поплавкового расходомера РС-3А (Россия).
Эмиссия СО2 с поверхности ствола дерева рассчитывалась как функция разности концентраций СО2 в выходящем из камеры и во входящем в камеру наружном воздухе, скорости воздушного потока через камеру и площади поверхности камеры, прилегающей к стволу (Молчанов, 2010; Молчанов и др., 2017). Измерения концентрации СО2 проводились с помощью портативного инфракрасного газоанализатора LI-820 (LI-COR Biosciences, США). Показания газоанализатора регистрировались каждые 5 с и сохранялись с 20-секундным осреднением на накопителе данных EMS (Чехия) вместе с данными о температуре почвы и воздуха, относительной влажности воздуха и приходящей суммарной солнечной радиации. Последовательное подключение измерительных камер к газоанализатору проводилось с помощью автоматической системы переключения каналов, построенной на основе трехходового переключателя, позволяющего осуществлять прокачку воздуха через камеры в течение всего периода измерений, препятствуя застаиванию воздуха в камерах в периоды, когда измерения газообмена не проводились (Молчанов, 2010, 2014).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Сезонная и суточная изменчивость дыхания стволов сосны. Результаты исследования дыхания стволов сосны свидетельствуют об их достаточно хорошо выраженной сезонной изменчивости. В разнотравно-черничном сосняке при оптимальной водообеспеченности сосны на протяжении вегетационного периода эмиссия СО2 с поверхности стволов характеризуется минимальными значениями весной и осенью и максимальными летом (табл. 1). Если весной при температуре 20−25°С средняя скорость дыхания стволов составляла 1.19 ± 0.41 мкмоль СО2/(м2 · с), то летом она несколько увеличилась, достигая 2.06 ± ± 0.43 мкмоль СО2/(м2 · с). Осенью наблюдалось устойчивое снижение эмиссии СО2 до 0.62 ± ± 0.11 мкмоль СО2/(м2 · с) (ноябрь).
Таблица 1.
Средние значения и стандартные отклонения эмиссии СО2 с поверхности стволов деревьев в разнотравно-черничном и пушицево-сфагновом сосняках
| Дата | УПГВ, м | Число измерений | Эмиссия CО2 с поверхности ствола, мкмоль СО2/(м2 · с) | |
|---|---|---|---|---|
| Сосняк разнотравно-черничный | ||||
| Дерево I класса роста | ||||
| 25−31.05.2011 | >2.0 | 120 | 1.19 ± 0.41 | |
| 27−31.07.2011 | >2.0 | 231 | 2.06 ± 0.43 | |
| 01−09.08.2011 | >2.0 | 408 | 1.72 ± 0.47 | |
| 06−15.10.2011 | >2.0 | 111 | 0.91 ± 0.38 | |
| 28.04−21.05.2012 | >2.0 | 1120 | 1.94 ± 0.58 | |
| 15−18.11.2012 | >2.0 | 127 | 0.62 ± 0.11 | |
| Сосняк пушицево-сфагновый | ||||
| Дерево IV класса роста | Дерево I класса роста | |||
| 27−28.05.2009 | 0.04 | 58 | 1.22 ± 0.21 | 1.34 ± 0.18 |
| 17−18.06.2009 | 0.07 | 48 | 0.91 ± 0.67 | 3.23 ± 0.72 |
| 24−25.06.2011 | 0.15 | 230 | 1.55 ± 0.32 | 2.76 ± 0.68 |
| 22−24.08.2011 | 0.43 | 60 | 0.58 ± 0.22 | 2.13 ± 0.49 |
В заболоченном пушицево-сфагновом сосняке на стационаре “Городище” сезонная динамика эмиссии СО2 менялась от года к году, достигая максимальных значений в мае−июне (для деревьев I класса роста – 1.3−3.2 мкмоль СО2/(м2 · с), а для деревьев IV класса роста – 0.9−1.8 мкмоль СО2/(м2 · с)) и постепенно уменьшаясь в июле−августе на фоне постепенного снижения УПГВ (табл. 1). Дыхание стволов для деревьев I класса роста снижалось в августе до 2.1 ± 0.5, а для деревьев IV класса роста – до 0.58 ± 0.2 мкмоль СО2/(м2 · с). При этом отмечалось отсутствие существенных различий между эмиссией СО2 с поверхности стволов деревьев I класса роста в сосняках пушицево-сфагновом и разнотравно-черничном.
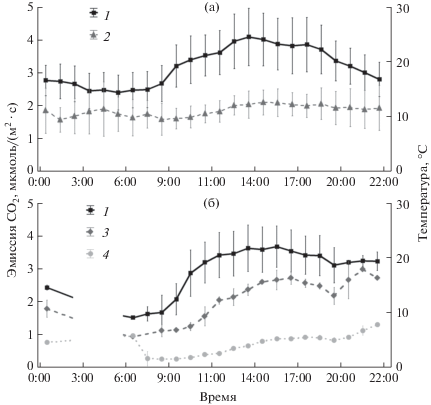
Внутрисуточная изменчивость эмиссии СО2 с поверхности стволов для обоих древостоев была выражена также достаточно хорошо. Для сосняка разнотравно-черничного минимальные значения скорости дыхания стволов наблюдались в ночные и утренние часы, а максимальные − в послеполуденное время (14:00) (рис. 1). Для сосняка пушицево-сфагнового минимальные значения скорости дыхания также были характерны для ночных и утренних часов, при этом максимум эмиссии был сдвинут на более позднее время (с первым максимумом в 15:00−17:00, и вторым максимумом в вечернее время). В суточном ходе для обеих экспериментальных площадей прослеживалась тесная связь дыхания стволов с изменением температуры воздуха, а для заболоченного сосняка пушицево-сфагнового отмечались также и существенные различия между скоростью дыхания стволов сосен, принадлежащих к разным классам роста. Так, в пушицево-сфагновом сосняке при УПГВ >40 см средняя скорость эмиссии СО2 с поверхности ствола сосны в дневное время у деревьев I класса роста была почти в 3 раза выше, чем у деревьев IV класса роста (2.4 ± 0.6 и 0.9 ± 0.2 мкмоль СО2/(м2 · с) соответственно) (рис. 1).
Рис. 1.
Суточный ход температуры воздуха и эмиссии СО2 с поверхности ствола в сосняках разнотравно-черничном (а) и пушицево-сфагновом (б) у деревьев I и IV классов роста при УПГВ 48 см. Приведены средние значения параметров и стандартные отклонения от среднего. 1 − температура воздуха, 2 − эмиссия СО2 с поверхности ствола в сосняке разнотравно-черничном, 3 − эмиссия СО2 с поверхности ствола в сосняке в пушицево-сфагновом I класса роста при УПГВ 48 см, 4 − эмиссия СО2 с поверхности ствола в сосняке в пушицево-сфагновом IV класса роста при УПГВ 48 см.
Зависимость дыхания стволов сосны от факторов внешней среды. Анализ зависимости дыхания стволов сосны обыкновенной от внешних условий показал, что если для сосняка разнотравно-черничного отмечается хорошо выраженная зависимость скорости дыхания стволов от температуры окружающего воздуха (рис. 2), то в заболоченном сосняке пушицево-сфагновом скорость эмиссии СО2 с поверхности стволов зависит не только от температуры воздуха, но и от УПГВ, который для заболоченного сосняка может быть рассмотрен в качестве индикатора степени влагообеспеченности древесного полога. При этом влияние УПГВ на дыхание стволов проявляется по-разному для деревьев разного класса роста (рис. 3). Минимальные значения скорости дыхания наблюдаются у деревьев I и IV классов роста при поверхностном залегании грунтовых вод. При увеличении УПГВ у деревьев I класса роста дыхание стволов растет, достигая максимума при УПГВ ∼10 см. При УПГВ 20−30 см наблюдается незначительное падение скорости дыхания с увеличением УПГВ, скорость которого варьирует в зависимости от температуры воздуха. В то же время для деревьев IV класса роста максимальные значения дыхания стволов сосны наблюдаются при УПГВ 15−20 см. При дальнейшем увеличении УПГВ у деревьев этого класса роста наблюдается устойчивое и более сильное снижение скорости дыхания стволов во всех температурных диапазонах (рис. 3).
Рис. 2.
Зависимость эмиссии СО2 с поверхности стволов сосны от температуры воздуха в сосняке разнотравно-черничном (июль−август 2011 г.) (а) и пушицево-сфагновом сосняке для деревьев I и IV классов роста (1 и 2 соответственно) при разных УГПВ (48 (б), 16 (в) и 2–5 см (г)). Штриховые кривые соответствуют модельным аппроксимациям (уравнение (1)), полученным с использованием параметров из табл. 2.
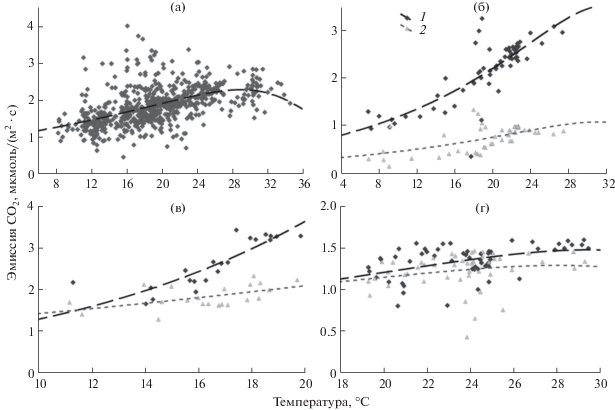
Рис. 3.
Зависимость эмиссии СО2 с поверхности стволов сосны от температуры воздуха и уровня почвенно-грунтовых вод (УПГВ) в пушицево-сфагновом сосняке у деревьев I (а) и IV (б) классов роста.
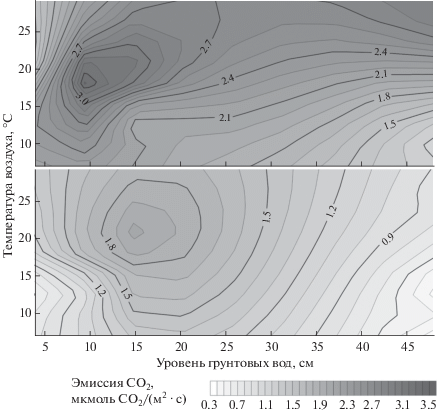
Анализ температурной зависимости скорости эмиссии СО2 с поверхности стволов как в разнотравно-черничном, так и в пушицево-сфагновом сосняках показал хорошо выраженный рост скорости дыхания при увеличении температуры от 6 до 28°С, а также относительно слабую зависимость скорости дыхания от температуры при температурах воздуха >28°С (рис. 3). При этом температурная зависимость дыхания ствола в пушицево-сфагновом сосняке для деревьев IV класса роста существенно отличалась от температурной зависимости для стволов деревьев I класса роста как при поверхностном (УПГВ 3−5 см), так и при более глубоком залегании грунтовых вод (УПГВ 45 см).
Параметризация зависимости дыхания стволов сосны от температуры и УПГВ. На основе анализа изменчивости скорости эмиссии СО2 с поверхности стволов сосны при изменении внешних условий была построена простая мультипликативная модель, учитывающая зависимость скорости дыхания от температуры воздуха и УПГВ, выбранного в качестве индикатора увлажнения корнеобитаемого слоя почвы.
Для описания изменения скорости дыхания стволов сосны при изменении температуры воздуха использовалась нелинейная функция, позволяющая описать как экспоненциальное увеличение скорости дыхания в диапазоне изменения температур до 25°C, так и ее возможное постепенное снижение при увеличении температуры выше некоторых критических значений, определяемых видоспецифическими особенностями деревьев в разных условиях произрастания. Вид подобной нелинейной функции близок к функциональной зависимости, используемой для описания отклика параметров фотосинтеза листьев растений на изменение температуры в биохимических моделях (De Pury, Farquhar, 1997). При построении графика зависимости скорости дыхания стволов сосны от УПГВ учитывалось снижение скорости дыхания при аномально высоком и низком УПГВ, выявленное в заболоченном сосняке пушицево-сфагновом (рис. 3). Для древостоев на хорошо дренированных участках (сосняк разнотравно-черничный) влияние УПГВ на скорость дыхания стволов не учитывалось.
В общем виде выражение для описания отклика дыхания ствола (Rstem) на изменение температуры воздуха (Ta) и УПГВ может быть записано так:
(1)
$\begin{gathered} R{}_{{{\text{stem}}}} = {{R}_{{{\text{stem}}\left( {{{T}_{{\text{a}}}} = 25^\circ {\text{C}}} \right)}}}{{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{T}_{{\text{a}}}}} \right){\kern 1pt} {{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{Z}_{W}}} \right), \\ {{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{T}_{{\text{a}}}}} \right) = \exp \left[ {\frac{{{{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}\left( {{{T}_{{\text{a}}}} - 298} \right)}}{{298R{{T}_{{\text{a}}}}}}} \right] \times \\ {{ \times \,\,\left[ {1 + \exp \left( {\frac{{298\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}} - {{H}_{{d,{{R}_{{{\text{stem}}}}}}}}}}{{298R}}} \right)} \right]} \mathord{\left/ {\vphantom {{ \times \,\,\left[ {1 + \exp \left( {\frac{{298\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}} - {{H}_{{d,{{R}_{{{\text{stem}}}}}}}}}}{{298R}}} \right)} \right]} {\left[ {1 + \exp \left( {\frac{{{{T}_{{\text{a}}}}\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}} - {{H}_{{d{\text{,}}{{R}_{{{\text{stem}}}}}}}}}}{{{{T}_{{\text{a}}}}\Re }}} \right)} \right]}}} \right. \kern-0em} {\left[ {1 + \exp \left( {\frac{{{{T}_{{\text{a}}}}\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}} - {{H}_{{d{\text{,}}{{R}_{{{\text{stem}}}}}}}}}}{{{{T}_{{\text{a}}}}\Re }}} \right)} \right]}}, \\ \end{gathered} $где ${{R}_{{{\text{stem}}\left( {{{T}_{{\text{a}}}}{\text{ = 25}}^\circ {\text{C}}} \right)}}}$ − значение Rstem при температуре воздуха 25°C в мкмоль/(м2 · с); ${{E}_{{{\text{a,}}{{R}_{{{\text{stem}}}}}}}}$ − энергия активации, определяющая скорость экспоненциального увеличения Rstem при росте температуры (при температурах ниже температурного оптимума для скорости дыхания), в Дж/моль; $\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}}$ − фактор, определяющий энтропию температурной зависимости Rstem, в Дж/(К · моль); ${{H}_{{d,\,{{R}_{{{\text{stem}}}}}}}}$ − параметр, определяющий скорость уменьшения Rstem при значениях температуры выше оптимума, в Дж/моль; $\Re $ − универсальная газовая постоянная ($\Re $ = 8.314 Дж/(моль · К); ${{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{T}_{{\text{a}}}}} \right)$ − функция, описывающая зависимость скорости дыхания от температуры; ${{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{Z}_{W}}} \right)$ − функция, описывающая зависимость скорости дыхания от УПГВ (ZW) и изменяющаяся от 0 до 1. Для разнотравно-черничного сосняка ${{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{Z}_{W}}} \right)$ принимается равной единице.
Выражение для описания лимитирующего влияния УПГВ на скорость дыхания стволов записывалось в следующем виде:
(2)
$\begin{gathered} {{f}_{{{{R}_{{{\text{stem}}}}}}}}\left( {{{Z}_{W}}} \right) = \\ = \left\{ {0.5\operatorname{th} \left[ {{{\left( {{{Z}_{W}} - {{{{d}_{1}}} \mathord{\left/ {\vphantom {{{{d}_{1}}} {2.5}}} \right. \kern-0em} {2.5}}} \right)} \mathord{\left/ {\vphantom {{\left( {{{Z}_{W}} - {{{{d}_{1}}} \mathord{\left/ {\vphantom {{{{d}_{1}}} {2.5}}} \right. \kern-0em} {2.5}}} \right)} {0.045}}} \right. \kern-0em} {0.045}}} \right] - } \right. \\ {{\left. { - \,\,0.5\operatorname{th} \left[ {{{\left( {{{Z}_{W}} - \left( {{{d}_{2}} + 0.2} \right)} \right)} \mathord{\left/ {\vphantom {{\left( {{{Z}_{W}} - \left( {{{d}_{2}} + 0.2} \right)} \right)} {0.1}}} \right. \kern-0em} {0.1}}} \right]} \right\}}^{m}}, \\ \end{gathered} $где d1 и d2 характеризуют интервал изменения УПГВ, в пределах которого он не является лимитирующим фактором для Rstem, a m определяет скорость ослабления скорости дыхания стволов деревьев при d2 > ZW. Значения d1 и d2 для деревьев I класса роста сосняка пушицево-сфагнового при температуре воздуха 25°С составляют по данным измерений 0.10 и 0.24 м, а для сосен IV класса роста − 0.12 и 0.22 м соответственно. Параметр m был принят равным 0.2 для деревьев I класса роста и 0.3 для деревьев IV класса роста.
Оценка параметров модели, описывающих зависимость скорости дыхания стволов от температуры и УПГВ, проводилась методом наименьших квадратов с использованием алгоритма Левенберга−Марквардта (Гилл и др., 1985). Параметры $\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}}$ и ${{H}_{{d,\,{{R}_{{{\text{stem}}}}}}}}$ определялись только на основе экспериментальных данных, полученных в разнотравно-черничном сосняке. Это было обусловлено наличием данных измерений скорости дыхания для достаточно широкого диапазона изменения температур, в том числе >30°С. Для параметризации дыхания стволов сосны в пушицево-сфагновом сосняке эти параметры принимались равными значениям, рассчитанным для разнотравно-черничного сосняка.
Сравнение ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ для разных типов сосняков показало значительные различия. Для сосняка пушицево-сфагнового IV класса роста отмечается устойчивый рост ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ с увеличением УПГВ, причем при УПГВ 48 см ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ = 36 878 Дж/моль, т.е. почти в 2 раза выше ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ при УПГВ 3−5 см (${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ = 18 733 Дж/моль). Для сосняка пушицево-сфагнового I класса роста отмечаются достаточно высокие значения ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ при УПГВ 15 см, существенно превышающие не только значения ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ при высоком и низком УПГВ (табл. 2), но также значения ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ для сосняка пушицево-сфагнового IV класса роста во всем диапазоне изменения УПГВ. Сравнивая значения ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ для разных типов сосняков, можно отметить их незначительные различия между сосняком разнотравно-черничным (${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ = 23 954 Дж/моль) и сосняками пушицево-сфагновым IV класса роста при среднем УПГВ (${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ = 27 361 Дж/моль) и I класса роста при высоком залегании грунтовых вод (${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$ = = 25 920 Дж/моль).
Таблица 2.
Значения параметров ${{R}_{{{\text{stem}}\left( {{{T}_{{\text{a}}}} = 25^\circ {\text{C}}} \right)}}}$ и ${{E}_{{{\text{a,}}\;{{R}_{{{\text{stem}}}}}}}}$ в уравнении (1) для расчета скорости эмиссии СО2 с поверхности ствола сосны в зависимости от температуры воздуха (при ${{H}_{{d,\;{{R}_{{{\text{stem}}}}}}}}$ = 202 000 Дж/моль, $\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}}$ = = 652 Дж/(К · моль))
| УПГВ, см | ${{R}_{{{\text{stem}}\left( {{{T}_{{\text{a}}}} = 25^\circ {\text{C}}} \right)}}}$, мкмоль СО2/(м2 · с) | ${{E}_{{{\text{a}},{{R}_{{{\text{stem}}}}}}}}$, Дж/моль |
|---|---|---|
| Сосняк разнотравно-черничный, деревья I класса роста | ||
| >200 | 2.18 | 23 954 |
| Сосняк пушицево-сфагновый, деревья I класса роста | ||
| 3−5 | 1.39 | 25 920 |
| 15 | 5.80 | 72 504 |
| 48 | 2.89 | 44 160 |
| Сосняк пушицево-сфагновый, деревья IV класса роста | ||
| 3−5 | 1.26 | 18 733 |
| 15 | 2.45 | 27 361 |
| 48 | 0.93 | 36 878 |
Примечание. ${{R}_{{{\text{stem}}\left( {{{T}_{{\text{a}}}} = 25^\circ {\text{C}}} \right)}}}$ − скорость дыхания ствола сосны при температуре 25°C; ${{E}_{{{\text{a,}}\;{{R}_{{{\text{stem}}}}}}}}$ − энергия активации, определяющая скорость экспоненциального увеличения дыхания ствола сосны при росте температуры (при температурах ниже температурного оптимума для скорости дыхания); ${{H}_{{d,\;{{R}_{{{\text{stem}}}}}}}}$ − параметр, определяющий скорость уменьшения скорости дыхания ствола сосны при значениях температуры выше оптимума;$\Delta {{S}_{{{{R}_{{{\text{stem}}}}}}}}$ − фактор, определяющий энтропию температурной зависимости дыхания ствола сосны.
Сравнение результатов модельных экспериментов показало что, несмотря на выявленные различия в зависимости скорости дыхания для разных типов сосняков от температуры и УПГВ, предложенная нелинейная модель (уравнения (1) и (2)) позволяет достаточно адекватно описать зависимость скорости дыхания от факторов внешней среды (рис. 3, табл. 2). Коэффициент детерминации (r2) между смоделированными и измеренными значениями скорости дыхания изменялся от 0.46 (p < 0.05) для разнотравно-черничного сосняка до 0.54 (p < 0.05) для сосняка пушицево-сфагнового.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ результатов экспериментальных исследований дыхания стволов сосны в разных условиях произрастания показывает их достаточно высокий диапазон изменчивости. При этом скорость дыхания стволов сосны в разных фенологических фазах развития может быть соизмерима со скоростями темнового дыхания охвоенных побегов (Zha et al., 2002). Этот факт подтверждается данными (Молчанов, 2007), показавшими, что ночное дыхание хвои, тонких корней и ствола сосны обыкновенной, а также почвы без корней составляет соответственно 8, 26, 23, 16% суммарного поглощения СО2 пологом соснового насаждения Iа бонитета. Таким образом, вклад эмиссии СО2 с поверхности стволов сосны в экосистемное дыхание соизмерим с вкладом автотрофного дыхания корней и превышает вклад гетеротрофного почвенного дыхания для исследуемой лесной экосистемы, что свидетельствует о важности учета данной составляющей при расчете интегральных экосистемных потоков СО2.
Выявленная в ходе исследования высокая чувствительность скорости дыхания стволов сосны к температуре отмечается многими авторами (Goulden et al., 1996; Zha et al., 2004; Забуга, 2006). Так, полученные данные измерений скорости дыхания в диапазоне изменения температур до 25°С хорошо согласуются с данными измерений (Молчанов и др., 2011), проведенных в Тверской обл. в спелом насаждении ели европейской и показавших двукратное увеличение скорости дыхания стволов ели в период с июня по начало августа на фоне роста температуры от 10 до 20°С. Ранее для сосны европейской в Финляндии была отмечена устойчивая сезонная изменчивость температурной чувствительности дыхания ствола на протяжении всего вегетационного периода (Zha et al., 2004). При этом чувствительность скорости дыхания к изменению температуры, определяемая температурным коэффициентом (Q10), была максимальна летом и составляла 1.98 (что соответствует росту скорости дыхания в 1.98 раза при увеличении температуры на 10°С). Сходные результаты были получены и Гулденом с соавт. (Goulden et al., 1996) для ряда лиственных и хвойных пород деревьев в Харвардском лесу, состоящем преимущественно из красного дуба и клена с участием канадской тсуги и белой сосны.
Значительные различия между скоростью эмиссии СО2 с поверхности стволов сосны в древостое I и IV классов роста сосняка пушицево-сфагнового могут быть связаны с влиянием комплекса факторов, среди которых существенную роль играют различия значений радиального прироста деревьев разных классов роста (Забуга, 2006; Забуга, Забуга, 2013, 2014). Радиальный прирост стволов деревьев IV класса роста априори значительно ниже такового у деревьев I класса роста, что обеспечивает более низкие скорости дыхания ствола, а также ее более слабую чувствительность к изменению факторов внешней среды (рис. 2). Определенное влияние на разные скорости дыхания стволов сосен разных классов роста могут оказывать и различия в водном режиме.
Важный момент, обращающий на себя внимание при анализе температурных зависимостей скорости эмиссии СО2 с поверхности живых стволов сосны – слабая чувствительность дыхания стволов к изменению температуры при ее значениях 26–28°С и выше. Данная особенность температурных зависимостей ранее отмечалась в основном при исследованиях температурного отклика ключевых параметров фотосинтеза, определяемых течением ферментативных реакций в листьях растений (Leuning, 1995). При рассмотрении температурных зависимостей эмиссии СО2 с поверхности ствола сосны данная закономерность может быть связана с замедлением течения ассимиляционных процессов сосны в послеполуденное время на фоне частичного закрытия устьиц из-за высоких температур воздуха, высокого дефицита упругости водяного пара и слабой обводненности побегов. Для пушицево-сфагнового сосняка данная тенденция может усиливаться также за счет падения УПГВ, приводящего к снижению влагообеспеченности корнеобитаемого слоя особенно для деревьев IV класса роста. Важно отметить, что при условии поверхностного залегания грунтовых вод различия в эмиссии СО2 с поверхности ствола у деревьев разного класса роста в сосняке пушицево-сфагновом почти полностью отсутствуют. Это может быть связано с недостатком кислорода в корнеобитаемом слое деревьев при высоком залегании грунтовых вод, что обычно ведет к снижению скорости ассимиляции (Молчанов, 1993) и, как следствие, к уменьшению скорости дыхания нефотосинтезирующих частей древесных растений.
Предлагаемая в рамках исследования параметризация зависимости скорости дыхания от температуры является развитием широко используемых в настоящее время модельных подходов, основанных на уравнении Аррениуса (Medlyn et al., 2002; Oltchev et al., 2008) и на правиле Вант-Гоффа, предполагающем увеличение скорости дыхания в 2−4 раза при росте температуры на 10°С (Lloyd, Taylor, 1994). Заложенное в обоих подходах бесконечное экспоненциальное увеличение скорости эмиссии СО2 с ростом температуры позволяет достаточно хорошо описать зависимость скорости дыхания нефотосинтезирующих частей растений от температуры 0–30°С. Однако при более высоких температурах (35−40°C) в условиях теплового стресса репрезентативность данного подхода вызывает сомнения, что подтверждается результатами проведенного исследования. Использованная в рамках данного исследования нелинейная функция (De Pury, Farquhar, 1997), позволяющая описать замедление и ослабление эмиссии СО2 при высоких температурах, с нашей точки зрения, более репрезентативная, если учитывать возможный значительный нагрев поверхности фотосинтезирующих и нефотосинтезирующих частей деревьев в летнее время и тесную связь скорости дыхания стволов с ассимиляционными процессами, протекающими в древесных растениях.
Включение в модельный алгоритм многопараметрической зависимости скорости дыхания стволов сосен от УПГВ позволило учесть разнонаправленные изменения эмиссии СО2 стволов сосен при повышении и значительном понижении УПГВ. Несмотря на относительную сложность построения модельного алгоритма, он опирается на использование нескольких ключевых параметров, описывающих скорость снижения дыхания стволов при подтоплении верхних горизонтов почвы и диапазон изменения УПГВ, при котором влияние УПГВ на скорость дыхания полностью отсутствует. Оценка этих параметров может быть легко выполнена на основании данных экспериментальных наблюдений в различных типах древостоев, расположенных на заболоченных территориях.
Выбор модельных параметризаций базировался пока на достаточно ограниченной выборке экспериментальных данных, полученных для двух типов сосняков в разных условиях произрастания, что, очевидно, нельзя считать достаточным для их широкого применения при проведении расчетов составляющих углеродного баланса лесных растительных сообществ в региональном масштабе. Очевидна необходимость дальнейшей проверки модели и ее оптимизации с привлечением новых данных о дыхании стволов деревьев разных пород в разных условиях произрастания.
ЗАКЛЮЧЕНИЕ
Результаты проведенных полевых экспериментов по исследованию временнóй изменчивости дыхания стволов сосны обыкновенной в разных типах леса при различной температуре воздуха и глубине залегания грунтовых вод показали, с одной стороны, наличие достаточно хорошо выраженной сезонной и внутрисуточной изменчивости, а с другой − устойчивую зависимость скорости дыхания стволов от температуры окружающего воздуха, а для заболоченного пушицево-сфагнового сосняка и от УПГВ. Существенные различия в скорости эмиссии СО2 с поверхности стволов деревьев в заболоченном сосняке были выявлены также и между деревьями разных классов роста. При этом деревья I класса роста оказались более чувствительными к изменению температуры и менее чувствительными к изменению УПГВ по сравнению с более угнетенными деревьями IV класса. Кроме того, у деревьев I класса роста при понижении УПГВ <40 см интенсивность дыхания была близкой по значениям к скорости дыхания стволов в разнотравно-черничном сосняке, что может быть обусловлено комплексом факторов, включая близкие значения скорости нетто фотосинтеза в данных типах сосняков. Все выявленные различия достаточно хорошо согласуются с имеющимися в литературе экспериментальными данными. Построенная на основании результатов экспериментов эмпирическая модель позволяет описать зависимость дыхания стволов сосны от температуры воздуха в сосняках разных типов и для деревьев различных классов роста в широком диапазоне изменения УПГВ. Полученная зависимость может найти применение для параметризации процессов СО2-обмена в моделях биогеохимического цикла земной поверхности локального и регионального масштабов. Ее дальнейшее уточнение возможно, в частности, с привлечением информации о вертикальном распределении корней деревьев разных классов роста по глубине, их водообеспеченности, минеральном составе почв и возрасте насаждений.
Работа выполнена при финансовой поддержке гранта РНФ 14-14-00956-П.
Список литературы
Алферов А.М., Блинов В.Г., Гитарский М.Л., Грабар В.А., Замолодчиков Д.Г., Зинченко А.В., Иванова Н.П., Ивахов В.М., Карабань Р.Т., Карелин Д.В., Калюжный И.Л., Кашин Ф.В., Конюшков Д.Е., Коротков В.Н., Кровотынцев В.А., Лавров С.А., Марунич А.С., Парамонова Н.Н., Романовская А.А., Трунов А.А., Шилкин А.В., Юзбеков А.К. Мониторинг потоков парниковых газов в природных экосистемах / Под ред. Замолодчикова Д.Г., Карелина Д.В., Гитарского М.Л., Блинова В.Г. Саратов: Амирит, 2017. 279 с.
Бурба Г.Г., Курбатова Ю.А., Куричева О.А., Авилов В.К., Мамкин В.В. Метод турбулентных пульсаций. Краткое практическое руководство. М.: Li-Cor Inc., 2016. 225 с.
Гилл Ф., Мюррей У., Райт М. Практическая оптимизация. Practical optimization. М.: Мир, 1985. 509 с.
Забуга В.Ф., Забуга Г.А. Дыхание сосны обыкновенной. Новосибирск: Наука, 2013. 208 с.
Забуга В.Ф., Забуга Г.А. Оценка дыхания сосны обыкновенной (Pinus sylvestris) на стадии кульминации ее текущего прироста в лесостепном Предбайкалье // Сиб. экол. журн. 2014. Т. 21. № 1. С. 93−107.
Забуга Г.А. Эколого-физиологические аспекты продукционного процесса сосны обыкновенной в лесостепном Предбайкалье: Автореф. дис. докт. биол. наук. Иркутск: Ангарск. гос. ун-т, 2006. 44 с.
Замолодчиков Д.Г., Грабовский В.И., Шуляк П.П., Честных О.В. Современное сокращение стока углерода в леса России // Докл. РАН. 2017. Т. 476. № 6. С. 719−721.
Корзухин М.Д. Расчетные оценки влияния изменения климата на продуктивность лесов (обзор подходов). Проблемы экол. мониторинга и моделирования экосистем. 2015. Т. 26. № 2. С. 33−58.
Молчанов А.Г. Сравнение интенсивности фотосинтеза сосны в разных эдафических условиях // Лесоведение. 1993. № 6. С. 76−80.
Молчанов А.Г. Баланс СО2 в экосистемах сосняков и дубрав в разных лесорастительных зонах. Тула: Гриф и К, 2007. 284 с.
Молчанов А.Г. Мониторинг эколого-физиологических показателей в экосистемах // Серебряноборское опытное лесничество: 65 лет лесного мониторинга. М.: КМК, 2010. С. 112−129.
Молчанов А.Г. СО2 древостоев в естественных условиях // Фотосинтетическая деятельность и продукционные процессы фитоценозов. Вып. 1. Орел: ГАУ, 2014. С. 63−88.
Молчанов А.Г., Курбатова Ю.А., Ольчев А.В. Влияние сплошной вырубки леса на эмиссию СО2 с поверхности почвы // Изв. РАН. Сер. биол. 2017. № 2. С. 190−196.
Молчанов А.Г., Татаринов Ф.А., Курбатова Ю.А. Эмиссия СО2 стволами живых деревьев и валежом в еловых лесах юга-запада Валдайской возвышенности // Лесоведение. 2011. № 3. С. 14−25.
Ольчев А.В., Курбатова Ю.А., Варлагин А.В., Выгодская Н.Н. Модельный подход для описания переноса СО2 между лесными экосистемами и атмосферой // Лесоведение. 2008. № 3. С. 3−13.
Ольчев А.В., Авилов В.К., Байбар А.С., Белотелов Н.В., Болондинский В.К., Иванов Д.Г., Кузьмина Е.В., Курбатова Ю.А., Левашова Н.Т., Мамкин В.В., Мангура П.А., Молчанов А.Г., Мухартова Ю.В., Никитин М.А., Новенко Е.Ю., Придача В.Б., Ривин Г.С., Розинкина И.А., Сазонова Т.А., Сандлерский Р.Б., Суркова Г.В., Холопцева Е.С. Леса Европейской территории России в условиях меняющегося климата. М.: КМК, 2017. 276 с.
Сафонов С.С., Карелин Д.В., Грабар В.А., Латышев Б.А., Грабовский В.И., Уварова Н.Е., Замолодчиков Д.Г., Коротков В.Н., Гитарский М.Л. Эмиссия диоксида углерода от разложения валежа в южнотаежном ельнике // Лесоведение. 2012. № 5. С. 44−49
Цельникер Ю.Л., Малкина И.С., Ковалев А.Г., Чмора С.Н., Мамаев В.А., Молчанов А.Г. Рост и газообмен СО2 у лесных деревьев. М.: Наука, 1993. 256 с.
De Pury D.G.G., Farquhar G.D. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models // Plant Cell Environ. 1997. V. 20. P. 537−557.
Edwards N.N., Sollins P. Continuous measurement of carbon dioxide evolution from partitioned forest floor components // Ecology. 1973. V. 54(2). P. 406−412.
Falge E., Baldocchi D., Tenhunen J., Aubinet M., Bakwin P., Berbigier P., Bernhofer C., Burba G., Clement R., Davis K.J., Elbers J.A., Goldstein A.H., Grelle A., Granier A., Guomundsson J., Hollinger D., Kowalski A.S., Katul G., Law B.E., Malhi Y., Meyers T., Monson R.K., Munger J.W., Oechel W., Paw U K.T., Pilegaard K., Rannik U., Rebmann C., Suyker A., Valentini R., Wilson K., Wofsy S. Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements // Agric. For. Meteorol. 2002. V. 113. P. 53−74.
Goulden M.L., Munger J.W., Fan S.-M., Daube B.C., Wofsy S.C. Measurements of carbon sequestration by long-term eddy covariance: methods and a critical evaluation of accuracy // Global Change Biol. 1996. V. 2. P. 169−182.
Ibrom A., Schütz C., Tworek T., Morgenstern K., Oltchev A., Falk M., Constantin J., Gravenhorst G. Eddy-correlation measurements of fluxes of CO2 and H2O above a spruce forest // J. Phys. Chem. Earth. 1996. V. 21(5−6). P. 409−414.
Ibrom A., Jarvis P.G., Clement R., Morgenstern K., Oltchev A., Medlyn B.E., Wang Y.P., Wingate L., Moncrieff J.B., Gravenhorst G. A comparative analysis of simulated and observed photosynthetic CO2 uptake in two coniferous forest canopies // Tree Physiol. 2006. V. 26. P. 845–864.
IPCC Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the IPCC. Cambridge: Cambr. Univ. Press, 2013. 1535 p.
Kurbatova J., Li C., Varlagin A., Xiao X., Vygodskaya N. Modeling carbon dynamics in two adjacent spruce forests with different soil conditions in Russia // Biogeosciences. 2008. V. 5. P. 969−980.
Kurbatova J., Li C., Tatarinov F., Varlagin A., Shalukhina N., Olchev A. Modeling of the carbon dioxide fluxes in European Russia peat bog // Environ. Res. Lett. 2009. V. 4. 045022.
Lavigne M.B., Ryan M.G., Anderson L. Comparing nocturnal eddy covariance measurements to estimates of ecosystem respiration made by scaling chamber measurements at six coniferous boreal sites // J. Geophys. Res. 1997. V. 102(28). P. 977–985.
Law B.E., Ryan M.G., Anthoni P.M. Seasonal and annual respiration of a ponderosa pine ecosystem // Global Change Biol. 1999. V. 5. P. 169–182.
Leuning R. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants // Plant Cell Environ. 1995. V. 18. P. 339–355.
Linder S., Trong E. The seasonal variation in stem and coarse root respiration of a 20-year scoots pine (Pinus sylvestris L.) // Mitt. Forstlichen Bund. Wien. 1981. H. 142. S. 125−139.
Lloyd J., Taylor Y. On the temperature dependence of soil respiration // Funct. Ecol. 1994. V. 8(3). P. 315−323.
Luyssaert S., Inglima I., Jung M., Richardson A.D., Reichsteins M., Papale D., Piao S.L., Schulze E.D., Wingate L., Matteucci G., Aragao L., Aubinet M., Beers C., Bernhoffer C., Black K.G., Bonal D., Bonnefond J.M., Chambers J., Ciais P., Cook B., Davis K.J., Dolman A.J., Gielen B., Goulden M., Grace J., Granier A., Grelle A., Griffis T., Grunwald T., Guidolotti G., Hanson P.J., Harding R., Hollinger D.Y., Hutyra L.R., Kolar P., Kruijt B., Kutsch W., Lagergren F., Laurila T., Law B.E., Le Maire G., Lindroth A., Loustau D., Malhi Y., Mateus J., Migliavacca M., Misson L., Montagnani L., Moncrieff J., Moors E., Munger J.W., Nikinmaa E., Ollinger S.V., Pita G., Rebmann C., Roupsard O., Saigusa N., Sanz M.J., Seufert G., Sierra C., Smith M.L., Tang J., Valentini R., Vesala T., Janssens I.A. CO2 balance of boreal, temperate, and tropical forests derived from a global database // Global Change Biol. 2007. V. 13. P. 2509−2537.
Malhi Y., Baldocchi D.D., Jarvis P.G. The carbon balance of tropical, temperate and boreal forests // Plant Cell Environ. 1999. V. 22. P. 715−740.
Medlyn B.E., Dreyer E., Ellsworth D., Forstreuter M., Harley P.C., Kirschbaum M.U.F., Le Roux X., Loustau D., Montpied P., Strassemeyer J., Walcroft A., Wang K. Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data // Plant, Cell Environ. 2002. V. 25. P. 1167–1175.
Oltchev A., Ibrom A., Ross T., Falk U., Rakkibu G., Radler K., Grote S., Kreilein H., Gravenhorst G. A modelling approach for simulation of water and carbon dioxide exchange between multi-species tropical rain forest and the atmosphere // J. Ecol. Modell. 2008. V. 212. P. 122–130.
Pumpanen J., Kolari P., Ilvesniemi H., Minkkinen K., Vesala T., Niinistö S., Lohila A., Larmola T., Morero M., Pihlatie M., Janssens I., Curiel Yuste J., Grünzweig J.M., Reth S., Subke J.-A., Savage K., Kutsch W., Østreng G., Ziegler W., Anthoni P., Lindroth A., Hari P. Comparison of different chamber techniques for measuring soil CO2 efflux // Agr. Forest Meteorol. 2004. V. 123. P. 159−176.
Rayment M.B., Jarvis P.G. An improved open chamber in the field // J. Geophys. Res. 1997. V. 102. P. 28779–28784.
Zha T.S., Wang K.Y., Ryyppö A., Kellomäki S. Needle dark respiration in relation to within-crown position in Scots pine trees grown in long-term elevation of CO2 concentration and temperature // New Phytologist. 2002. V. 156. P. 33−41.
Zha T., Kellomaki S., Wang K.-Y., Ryyppo A., Ninisto S. Seasonal and annual stem respiration of Scots pine trees under boreal conditions // Ann. Bot. 2004. V. 94(6). P. 889−896.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая