

Известия РАН. Серия биологическая, 2020, № 5, стр. 507-511
Изменчивость признаков черепа кабарги Moschus moschiferus L. в разных фазах популяционного цикла
В. И. Приходько *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: pvi-1949@mail.ru
Поступила в редакцию 19.01.2018
После доработки 20.01.2019
Принята к публикации 20.01.2019
Аннотация
Исследована краниальная изменчивость четырех (трех краниометрических и одного неметрического – дополнительные косточки) признаков 105 черепов кабарги из предгорий Восточного Саяна в разных фазах (депрессии и роста численности) популяционного цикла вида. Отмечена продолжительность цикла 50 лет, что соответствует времени 10 поколений животных. Выявлена разнонаправленная связь линейных размеров черепа у особей разного пола с фазами численности популяции. Установлено, что изменчивость неметрического признака черепа характеризуется наибольшей частотой проявления и максимальным числом косточек в фазе депрессии у животных обоего пола, постепенным убыванием их числа в фазе роста, затем полной утратой косточек по мере нарастания численности популяции. Обнаружено, что черепные фены ключевые в краниальной изменчивости и могут быть использованы как критерии для мониторинга природных популяций редких и исчезающих подвидов кабарги.
Важнейшее проявление внутривидовой изменчивости – морфологическая неоднородность природных популяций при преобразованиях их структур (Одум, 1975; Шварц, 1977, 1980). В большинстве случаев морфологическая изменчивость обусловлена периодическими флуктуациями численности, изменениями пространственно-этологической структуры географических популяций в разных фазах популяционных циклов (Шварц, 1977, 1980; Шилов, 1977).
Изменчивость популяций мелких млекопитающих в процессе динамики численности связывается с влиянием абиотических факторов среды или сезонной доступностью кормов в определенные периоды роста животных (Krebs, 1964; Krebs, Myers, 1974; Mihok, Fuller, 1981; Lidicker, Ostfeld, 1991; Wolff, 1993; Васильев и др., 2004). В ходе циклических колебаний численности выживают и отбираются генетически разнокачественные особи, адаптированные к высоким или низким значениям плотности населения (Krebs, 1978; Шилов, 1991).
Одна из проблем, с которой сталкиваются исследователи, – трудности выявления корреляции между динамикой численности и изменчивостью морфологических признаков для сопоставимых фаз популяционного цикла копытных. Флуктуации численности у диких копытных доказаны, как и их причины, связанные с комплексом средовых условий (Ломанов, 1995; Шилов, 1997) и антропогенными (Данилкин, 2009) факторами. Динамика населения диких копытных влияет на экологическую структуру и на феногенотипическую изменчивость популяций. Для сайгака Saiga tatarica L. на основе статистического анализа признаков экстерьера животных показано наличие фенотипической специфичности у разных поколений в изолированной популяции в ходе изменения ее численности (Проняев, Аксенов, 1988).
Опубликованные ранее данные свидетельствуют о быстром снижении численности природных популяций кабарги в северной части ареала в конце XIX в. (Туркин, Сатунин, 1902; Щербаков, 1959; Приходько, 2008), когда вид был на грани вымирания. Причиной катастрофического сокращения населения вида было истребление животных человеком. Депрессия численности, длившаяся на протяжении десятилетий, по-видимому, повлияла не только на пространственно-этологическую структуру, но и на параметры морфологических признаков популяций в границах всего ареала вида.
Цель работы – исследование изменчивости краниологических признаков в разных фазах популяционного цикла кабарги, населявшей в Восточном Саяне одни и те же местообитания в условиях воздействия антропогенного фактора на структуру популяций.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использованы музейные коллекции (Зоологического музея МГУ, Москва, серия “S”; Зоологического института РАН, Санкт-Петербург) черепов кабарги из популяции бассейна р. Манна Восточного Саяна. Всего обработано 105 черепов разнополых особей, возраст которых превышал 1 год. Возраст определяли по стадии развития зубной системы (Приходько, 2004). К выборке, характеризующей депрессивную фазу численности популяции, отнесены черепа животных, добытых с 1885 по 1894 г. (сборы М.Е. Киборта, 53 экз.). Вторая выборка черепов (сборы А.Н. Щербакова и других исследователей, 52 экз.) отнесена к середине фазы роста численности той же географической популяции, а период сбора образцов 1950–1953 гг. При анализе выборок в табл. 1 используются датировки 1893 и 1950 гг. Временной период между выборками одного популяционного цикла составляет 50 лет, что соответствует времени 10 поколений кабарги.
Таблица 1.
Средние значения основных промеров черепа кабарги и частоты встречаемости дополнительных черепных косточек
| Признак | Выборка черепов 1893 г. (депрессия численности) |
Выборка черепов 1950 г. (подъем численности) |
||
|---|---|---|---|---|
| самцы (n = 39) |
самки (n = 14) |
самцы (n = 33) |
самки (n = 19) |
|
| МД | 154.21 ± 0.58 | 158.26 ± 0.85 | 155.65 ± 0.67 | 156.43 ± 0.86 |
| ДЛО | 76.7 ± 0.43 | 78.07 ± 0.72 | 76.8 ± 0.49 | 77.07 ± 0.48 |
| ШМК | 46.13 ± 0.17 | 46.15 ± 0.25 | 46.17 ± 0.19 | 45.74 ± 0.19 |
| ЧДК | 0.18 | 0.07 | 0.087 | 0 |
Три абсолютных измерения и один неметрический признак черепа использовали в качестве переменных для статистических исследований. При анализе морфологической изменчивости использовали общепринятые наиболее важные признаки черепа кабарги: максимальную длину (МД) – от наиболее выступающих назад точек на затылочных мыщелках до краевого конца верхнечелюстных костей; орбитальную длину лицевого отдела (ДЛО) – от переднего края глазницы до краевого конца верхнечелюстной кости; ширину мозговой капсулы (ШМК) – между внешними боковыми краями мозговой капсулы; частоту встречаемости дополнительных косточек (ЧДК), расположенных на границах срастания лобных и теменной костей (рис. 1). Фенотипический (фен) черепной признак характеризуется наличием на крыше мозговой капсулы двух дополнительных косточек, одной косточки и отсутствием косточек. Мы анализируем лишь частоту их встречаемости в выборках. Промеры проводили с точностью до 0.1 мм.
Рис. 1.
Основные промеры и полиморфность неметрического признака черепа кабарги. МД – максимальная длина, ДЛО – орбитальная длина лицевого отдела, ШМК – ширина мозговой капсулы. Цифрами обозначены музейные номера черепов – с одной (а) и с двумя (б) косточками.
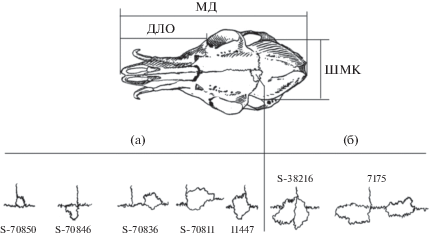
Для метрических признаков рассчитывали средние значения и оценивали достоверность различий по t-критерию. Результаты половых различий в исследуемых выборках черепов кабарги приведены в табл. 1.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изменчивость краниометрических признаков. По средним значениям длины черепа самки превосходят самцов в выборках 50-летнего популяционного цикла. Наблюдаемая закономерность особенно проявляется между самцами и самками в фазе депрессии численности. От фазы спада к фазе подъема численности прослеживается незначительное увеличение максимальной длины черепа самцов. По мере нарастания численности размеры черепа самок существенно уменьшаются. Одновременно у самок наблюдается также направленное к фазе роста уменьшение средних размеров мозговой капсулы. Вместе с тем статистический анализ средних значений черепа не выявил достоверных различий между однополыми особями из разных фаз цикла.
Особенности краниометрической изменчивости в разных фазах численности свидетельствуют о специфичности популяционного цикла кабарги, длившегося с конца XIX до конца ХХ в. Наличие противоположно направленного изменения размеров черепа у полов в зависимости от численности согласуется с данными автора, полученными в полевых условиях по избирательной добыче человеком животных среди половозрастных групп (Приходько, 2018). Обычно снижению численности вида предшествует изъятие из популяции взрослых самцов и частично молодых самок с пониженной реакцией на опасность, что приводит к нарушению соотношения полов в пользу старых самок, раннему участию молодых самцов в размножении и, возможно, проявлению инбредной депрессии на протяжении всей фазы спада. В фазе подъема численности по мере восстановления пространственно-этологической структуры популяции в размножение вступают в основном средневозрастные особи, что обеспечивает нивелировку размеров черепов у разнополых особей. Выявленный же половой диморфизм в размерах черепов объясняется развитием у самцов длинных клыков; их рост оказывает влияние на укорочение межчелюстных и верхнечелюстных костей в онтогенезе.
Изменчивость неметрического признака. Неметрические признаки часто сопряжены с общей морфологической изменчивостью, зависящей от колебаний численности и связанной с ней плотностью населения, как показано на грызунах (Васильев и др., 2004). Наличие на крыше мозговой капсулы кабарги одной или двух косточек рассматривается не как аномалия в строении черепа, а как проявление морфологического разнообразия в условиях флуктуирующей численности.
Анализ частоты встречаемости неметрического признака показал его преобладание в выборках самцов и самок в фазе депрессии численности. Более того, у самцов в фазе спада чаще, чем у самок, проявляется наличие пары косточек. Частота встречаемости черепного фена при низкой численности снижается с 18% у самцов до 7% у самок. Сходное смещение частот фена отмечено также для разнополых особей в фазе подъема численности, причем самки в этот период характеризуются отсутствием неметрического черепного признака. Практически равные частоты проявления этого признака обнаруживаются у самок в период депрессии и у самцов в фазе подъема численности. Отмеченное смещение фена в разных фазах популяционного цикла кабарги свидетельствует о его неустойчивом состоянии, обусловленном, по-видимому, уровнем генетических различий среди животных при существенных колебаниях численности.
Выделенный полиморфный признак проявляется у животных разного возраста, имеющих как широкую, так и узкую мозговую капсулу. Следовательно, черепной фен может иметь несколько пороговых состояний (две дополнительные косточки > одна > 0), которые соответствуют убывающим нормам генетического разнообразия популяции по мере роста численности населения. Можно предположить, что популяции с нарушенной человеком структурой стремятся перейти от полиморфности морфологических структур (признаков) к устойчивой фенотипической мономорфности. Развитие нескольких косточек на своде черепа обуславливает все фенотипические варианты, проявляющиеся при локальной депопуляции на основе одной и той же нормы реакции при разной плотности населения. Таким образом, фаза депопуляции – поставщик обширного разнообразия морфологических признаков кабарги для естественного отбора в ходе микроэволюции вида.
Ранее на большой выборке черепов (349 экз.) кабарги было показано возрастание проявления рассматриваемого фена у животных в направлении от центра видообразования (Внутренняя Монголия) к северной части ареала вида (Приходько, 2017; Prikhod’ko, 2017). Выявленная наибольшая внутривидовая изменчивость фенотипа черепа приурочена к центру современного ареала (Забайкалье, Саяны), где в прошлом происходили значительные изъятия ресурсов животных этого вида человеком. По этой причине наблюдались периодические флуктуации как локальной численности, так и экологической плотности населения в природных популяциях.
Таким образом, географическое положение популяции и циклы ее численности влияют на проявление фенотипичных признаков черепа кабарги, которые могут быть специфичными для каждого цикла. Вероятность реализации той или иной морфогенетической схемы определяется естественным отбором и действиями генов, контролирующими морфологические модификации. Естественный отбор, как свидетельствуют наши данные, повышает частоту встречаемости аллелей “дополнительные косточки черепа” (ДКЧ) у кабарги от 7 до 18% в зависимости от пола за относительно короткий временной период (50 лет). Генетические варианты аллелей ДКЧ постепенно утрачиваются в фазе подъема, а на пике численности животные могут передавать исходные гены фенотипической мономорфности без ДКЧ следующим поколениям. В этой связи можно предположить, что процесс формирования и утраты черепных фенов – периодическое явление в природных флуктуирующих популяциях кабарги.
Выявленные неметрические черепные фены – ключевые в комплексе признаков фенотипической изменчивости в фазах популяционного цикла – могут быть морфологическими критериями для мониторинга состояния природных популяций редких и находящихся под угрозой исчезновения подвидов кабарги.
Список литературы
Васильев А.Г., Фалеев В.И., Галактионов Ю.К., Ковалева В.Ю., Ефимов В.М., Епифанцева Л.Ю., Поздняков А.А., Дупал Т.А., Абрамов С.А. Реализация морфологического разнообразия в природных популяциях млекопитающих. 2-е изд., испр. Новосибирск: Изд-во СО РАН, 2004. 232 с.
Данилкин А.А. Динамика населения диких копытных России: гипотезы, факторы, закономерности. М.: КМК, 2009. 310 с.
Ломанов И.К. Закономерности динамики численности и размещения населения лося в Европейской части России. М.: Изд-во ЦНИЛ Охотдепартамента МСХиП РФ, 1995. 60 с.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
Приходько В.И. Закономерности роста и развития кабарги, Moschus moschiferus (Artiodactyla) // Зоол. журн. 2004. Т. 83. № 2. С. 252–260.
Приходько В.И. Разведение кабарги. Научно-практические рекомендации. М.: КМК, 2008. 142 с.
Приходько В.И. Изменчивость неметрических признаков черепа кабарги Moschus moschiferus L. (Moschidae, Cetartiodactyla) // Изв. РАН. Сер. биол. 2017. № 2. С. 142–148.
Приходько В.И. Динамика численности кабарги (Moschus moschiferus L.) в России // Вестн. охотоведения. 2018. Т. 15. № 1. С. 26–32.
Проняев А.В., Аксенов С.В. Фенотипическая изменчивость изолированной популяции сайгака в ходе изменения численности // Хронологические изменения численности охотничьих животных в РСФСР. М.: ЦНИЛ Главохоты РСФСР, 1988. С. 143–151.
Туркин Н.В., Сатунин К.А. Звери России. М.: Изд-во Н.В. Туркина, 1902. 506 с.
Шварц С.С. Внутривидовая изменчивость и видообразование. Эволюционный и генетический аспекты проблемы // Успехи соврем. териологии. М.: Наука, 1977. С. 279–290.
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 277 с.
Шилов И.А. Эколого-физиологические основы популяционных отношений у животных. М.: Изд-во МГУ, 1977. 261 с.
Шилов И.А. Динамика популяций и популяционные циклы // Структура популяций у мелких млекопитающих. М.: Наука, 1991. С. 151–172.
Шилов И.А. Экология. М.: Высш. шк., 1997. 512 с.
Щербаков А.Н. Кабарга и ее мускус // Уч. зап. Хабаров. пед. ин-та, 1959. Т. 4. С. 79–87.
Krebs C.J. Cyclic variation in skull-body regression of lemmings // Can. J. Zool. 1964. V. 42. P. 631–643.
Krebs C.J. A review of the Chitty hypothesis of population regulation // Can. J. Zool. 1978. V. 56. P. 2463–2480.
Krebs C.J., Myers J.H. Population cycles in small mammals // Adv. Ecol. Res. 1974. V. 8. P. 267–399.
Lidicker W.Z., Ostfeld R.S. Extralarge body size in California voles: causes and fitness consequences // Oikos. 1991. V. 61. № 1. P. 108–121.
Mihok S., Fuller W. Morphometric variation in Clethrionomys gapperi; are all voles created equal // Can. J. Zool. 1981. V. 59. P. 2275–2283.
Prikhod’ko V.I. Variability of nonmetric traits of musk deer Moschus moschiferus L. (Moschidae, Cetartiodactyla) cranium // Biol. Bull. 2017. V. 44. № 2. P. 172–178.
Wolff J.O. Does the “Chitty effect” occur in Peromyscus? // J. Mammal. 1993. V. 74. № 4. P. 846–851.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая