

Известия РАН. Серия биологическая, 2020, № 5, стр. 534-547
Панцирные клещи – обитатели лишайников в таежной зоне европейского Северо-Востока: биотопическая приуроченность, экологические группы видов
Е. Н. Мелехина *
Институт биологии Коми научного центра Уральского отделения Российской академии наук
167982 г. Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: melekhina@ib.komisc.ru
Поступила в редакцию 12.11.2018
После доработки 18.11.2019
Принята к публикации 14.02.2020
Аннотация
В качестве местообитаний панцирных клещей обследованы напочвенные (Cladina arbuscula, C. rangiferina, C. stellaris, Cetraria islandica) и эпифитные (Hypogymnia physodes и Bryoria subcana) лишайники в таежных лесах европейского Северо-Востока. Всего в лишайниках разных видов обнаружено 55 видов орибатид из 30 семейств. Выделено пять экологических групп панцирных клещей – обитателей лишайников. Установлено, что специфику населения эпифитов составляют виды арборикольной доминирующей и арборикольной немногочисленной групп. Показано, что для напочвенных лишайников характерны гемиэдафические доминирующие и гемиэдафические немногочисленные виды. Выделена арборикольно-гемиэдафическая группа, которая включает в себя виды, обитающие как в напочвенных, так и в эпифитных лишайниках. Выявлены экологически викарирующие виды по отношению к эпифитам – представители родов Phauloppia, Mycobates, Carabodes, Oribatula.
Панцирные клещи – постоянные и многочисленные обитатели лишайникового и мохового покрова на деревьях (Trave, 1963; Бязров, 1988; Мелехина, Бязров, 1997), почве, камнях и прибрежных скалах (Gjelstrup, Søchting, 1979; Coloff, 1983; Ниеми, Вилкамаа, 1988; Тарба, 1992). Определены комплексы видов, связанных с лишайниками как местообитаниями, в различных регионах (Strenzke, 1952; Pschorn-Walcher, Gunhold, 1957; Andre, 1975, 1979, 1984, 1985; Seyd, 1988; Seyd, Seaward, 1984). Орибатиды рассматривались в качестве компонентов лишайниковых консорций (Trave, 1963; Бязров и др., 1971). Исследованы биотопические предпочтения панцирных клещей, обитающих в лишайниках разных жизненных форм, произрастающих на разных субстратах, в различных растительных сообществах (Бязров, Мелехина, 1992, 1994; Мелехина, Бязров, 1997; Штанчаева, 1997). Панцирные клещи – обитатели эпифитных лишайников – были представлены как биоиндикаторы радиоактивного загрязнения природных экосистем (Мелехина, Криволуцкий, 1993).
В таежной зоне лишайники принимают заметное участие в сложении лесных биоценозов и отличаются разнообразием видов (Пыстина, 2003). Получены данные о таксономическом составе и особенностях распределения панцирных клещей в лишайниках разных видов и жизненных форм (Мелехина, 2000, 2001).
Цель работы – анализ таксономического разнообразия и биотопической приуроченности панцирных клещей, обитающих в лишайниках; выявление комплексов видов, ассоциированных с напочвенными и эпифитными лишайниками в хвойных лесах таежной зоны европейского Северо-Востока.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования были проведены в окрестностях пос. Кажым Койгородского р-на Республики Коми (60°19′58′′ с.ш., 51°32′00′′ в.д.). Климат в районе исследований умеренно-континентальный, с продолжительной зимой и коротким прохладным летом. Среднемесячные температуры января и июля –15, +17°C соответственно. Продолжительность безморозного периода ~100 сут, годовое количество осадков до 700 мм, средняя максимальная высота снежного покрова в лесу 100 см (Атлас…, 1997).
Преобладающие растительные формации – хвойные таежные фитоценозы. В подзоне средней тайги на плакорах господствуют еловые леса, древесный ярус в которых состоит из ели сибирской Picea obovata Lebed., иногда с примесью березы и пихты. Наиболее типичные ассоциации – ельники чернично-зеленомошные; в травянисто-кустарничковом ярусе преобладают мелкие лесные кустарнички, главным образом черника и брусника (Юдин, 1954; Леса…, 1999). На боровых террасах и междуречных равнинах развиты сосновые леса, которые образованы сосной обыкновенной Pinus silvestris L.; широко распространены сосняки лишайниковые. Древостой чаще всего чисто сосновый, иногда с примесью березы, реже встречаются лиственницы и ели. Травянисто-кустарничковый ярус разрежен, кустарнички представлены брусникой, черникой, вороникой; в напочвенном покрове преобладают лишайники, по затененным местам встречаются мхи (Юдин, 1954; Леса…, 1999). Доминантами напочвенного покрова обычно выступают лишайники рода Cladina – C. stellaris (Opiz.) Brodo, C. rangiferina (L.) Nyl. и C. arbuscula (Wallr.) Hale & W. L. Culb, часто отмечается Cetraria islandica (L.) Ach. (Пыстина, 2003).
Наблюдения были проведены в двух типах сосновых сообществ (сосняке лишайниково-зеленомошном и сосняке черничном) и двух типах еловых сообществ (ельнике черничном и ельнике зеленомошном). В качестве местообитаний панцирных клещей обследованы популяции лишайников разных видов и жизненных форм: напочвенных Cladina arbuscula, C. rangiferina, C. stellaris и Cetraria islandica, эпифитных Hypogymnia physodes (L.) Nyl. и Bryoria subcana (Nyl. ex Stiz.) Brodo et D. Hawksw. (табл. 1). Видовая принадлежность собранных лишайников была определена Л.Г. Бязровым. Таксономия лишайников приведена по классификации Сантессона (Santesson, 1993), а жизненные формы – по классификации Голубковой и Бязрова (Голубкова, Бязров, 1989).
Таблица 1.
Обследованные местообитания панцирных клещей в хвойных растительных сообществах
| Лишайники | Растительное сообщество | |||
|---|---|---|---|---|
| сосняк | ельник | |||
| лишайниково-зеленомошный | черничный | черничный | зеленомошный | |
| Напочвенные радиально-лопастные кустистые прямостоячие | ||||
| Cladina arbuscula | + | + | + | − |
| Cladina rangiferina | + | + | + | + |
| Cladina stellaris | + | + | − | + |
| Напочвенный плосколопастной кустистый | ||||
| Cetraria islandica | + | + | + | − |
| Эпифитный листоватый | ||||
| Hypogymnia physodes | + | + | + | − |
| Эпифитный кустистый повисающий | ||||
| Bryoria subcana | − | − | − | + |
Сбор материала проводили в июле–августе 1989–1992 гг. Слоевища H. physodes собирали на высоте 1.5–2.0 м со стволов сосен в сосняках и со стволов и ветвей елей в ельнике черничном, B. subcana – со стволов и ветвей елей; сборы эпифитов с десяти стволов составляли среднюю пробу. Пробы напочвенных лишайников и почвы, площадью 100 см2 каждая, отбирали в десятикратной повторности в каждом растительном сообществе дважды за период наблюдения (в 1989 и 1992 гг.). Почву отбирали на глубину до 10 см отдельно от лишайников. Всего было проанализировано 80 почвенных проб, 80 проб напочвенных лишайников и 32 пробы эпифитных лишайников объемом 2 л каждая.
Было идентифицировано >30 тыс. экземпляров половозрелых панцирных клещей. Группировки панцирных клещей сравнивали по видовому составу и структуре населения, используя методы анализа фаунистического материала (Песенко, 1982). Определяли долю каждого вида в структуре населения (Pij, %), степень участия местообитания в размещении вида (qij, %), степень относительной биотопической приуроченности видов (F). На основании этих показателей определяли принадлежность вида к той или иной экологической группе. При описании структуры доминирования использовалась классификация Энгельмана (Потапов, Кузнецова, 2011): эудоминант – 39.4–100, доминант – 12.4–39.3, субдоминант – 3.9–12.3, малочисленный (рецедент) – 1.3–3.8, редкий (субрецедент) – <1.3%. Таксономия панцирных клещей приведена по классификации Субиаса (Subias, 2019).
Было проанализировано распространение видов в лишайниковых группировках различных регионов (Strenzke, 1952; Trave, 1963; Andre, 1975, 1979, 1984, 1985; Gjelstrup, Sǿchting, 1979; Solhǿy, Koponen, 1981; Seyd, Seaward, 1984; Ниеми, Вилкамаа, 1988; Stary, 1988; Бязров, Мелехина, 1992, 1994; Тарба, 1992; Штанчаева, 1997; Мелехина, Бязров, 1997; и др.).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Видовой состав, структура доминирования. В обследованных местообитаниях в общей сложности было обнаружено 90 видов панцирных клещей, представителей 33 семейств; в лишайниках четырех растительных сообществ было найдено 55 видов из 30 семейств. Наибольшей видовой насыщенностью отличались семейства Oribatulidae (5 видов), Suctobelbidae (7), Crotoniidae (3), Phthiracaridae (3), Carabodidae (3). Большинство семейств было представлено 1–2 видами каждое. Наибольшее число таксонов отмечалось, как правило, в напочвенных лишайниках (рис. 1). В табл. 2–5 представлены виды, которые проявили положительную или отрицательную биотопическую приуроченность к тому или иному местообитанию.
Рис. 1.
Число таксонов панцирных клещей в напочвенных и эпифитных лишайниках сосновых и еловых растительных сообществ (C. arb. – Cladina arbuscula, C. ran. – Cladina rangiferina, C. stell. – Cladina stellaris, C. isl. – Cetraria islandica, H. phys. – Hypogymnia physodes, B. subc. – Bryoria subcana). а – сосняк лишайниково-зеленомошный, б – сосняк черничный, в – ельник черничный, г – ельник зеленомошный; 1–3 – число видов, родов и семейств соответственно.
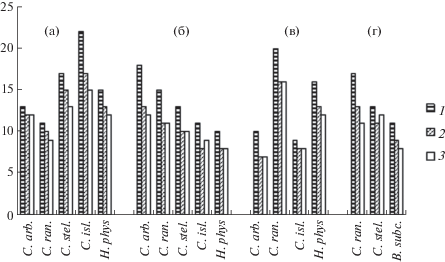
Таблица 2.
Распределение панцирных клещей по местообитаниям в сосняках
| Вид/местообитание/ показатели биотопической приуроченности | Cladina arbuscula | Cladina rangiferina | Cladina stellaris | Cetraria islandica | Hypogimnia physodes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P | q | F | P | q | F | P | q | F | P | q | F | P | q | F | |
| Phthiracarus laevigatus | – | – | – | – | – | – | 1.0 | 60.0 | 0.52 | 0.5 | 20.0 | –0.04 | – | – | – |
| P. longulus | – | – | – | 0.9 | 70.0 | 0.80 | – | – | – | 0.4 | 30.0 | 0.23 | – | – | – |
| Trhypochthonius cladonicolus | – | – | – | 44.4 | 57.4 | 0.68 | 20.4 | 42.1 | 0.21 | 0.4 | 0.5 | –0.98 | – | – | – |
| Camisia biurus | – | – | – | – | – | – | 0.3 | 25.0 | –0.17 | 1.2 | 75.0 | 0.83 | – | – | – |
| Furcoppia dentata | – | – | – | – | – | – | – | – | – | – | – | – | 1.2 | 100 | 1.0 |
| Ceratoppia quadridentata | – | – | – | – | – | – | – | – | – | 1.0 | 88.9 | 0.93 | 0.4 | 11.1 | 0.27 |
| Adoristes ovatus poppei | 0.3 | 3.0 | –0.77 | 1.8 | 19.7 | –0.01 | 1.6 | 28.8 | –0.08 | 4.1 | 48.5 | 0.55 | – | – | – |
| Eueremaeus oblongus silvestris | 0.3 | 8.0 | –0.47 | – | – | – | 1.9 | 92.0 | 0.92 | – | – | – | – | – | – |
| Graptoppia foveolata | – | – | – | – | – | – | – | – | – | – | – | – | 0.9 | 100 | 1.0 |
| Suctobelbella acutidens duplex | – | – | – | – | – | – | 1.5 | 90.0 | 0.90 | 0.3 | 10.0 | –0.41 | – | – | – |
| Carabodes labyrinthicus | – | – | – | – | – | – | – | – | – | – | – | – | 32.1 | 100 | 1.0 |
| C. marginatus | 7.7 | 30.5 | 0.29 | 6.5 | 26.7 | 0.18 | 4.7 | 30.5 | –0.04 | 2.7 | 11.7 | –1.0 | 0.4 | 0.6 | –0.85 |
| C. subarcticus | 50.4 | 24.0 | 0.13 | 27.8 | 13.6 | –0.23 | 49.5 | 38.7 | 0.14 | 46.1 | 23.7 | 0.07 | – | – | – |
| Tectocepheus velatus | 0.7 | 5.9 | –0.59 | 5.0 | 43.5 | 0.51 | 1.6 | 28.8 | –0.08 | 2.6 | 23.5 | 0.07 | – | – | – |
| Trichoribates berlesei | 0.3 | 13.3 | –0.22 | 0.7 | 33.3 | 0.33 | – | – | – | – | – | – | 3.3 | 53.4 | 0.88 |
| Chamobates pusillus | 0.1 | 33.3 | 0.35 | – | – | – | – | – | – | – | – | – | 0.8 | 66.7 | 0.93 |
| Diapterobates humeralis | – | – | – | – | – | – | – | – | – | – | – | – | 6.1 | 100 | 1.0 |
| D. oblongus | – | – | – | – | – | – | – | – | – | 0.5 | 50.0 | 0.57 | 1.6 | 50.0 | 0.86 |
| Oribatula (Z.) propinqua | – | – | – | – | – | – | – | – | – | – | – | – | 45.5 | 100 | 1.0 |
| Phauloppia nemoralis | – | – | – | – | – | – | 0.1 | 9.1 | –0.65 | – | – | – | 4.1 | 90.9 | 0.98 |
| Scheloribates laevigatus | 38.0 | 35.3 | 0.38 | 11.8 | 11.3 | –0.33 | 13.8 | 21.1 | –0.28 | 31.6 | 31.8 | 0.27 | 1.6 | 0.5 | –0.86 |
| Pergalumna nervosa | 0.6 | 50.0 | 0.61 | – | – | – | 0.1 | 12.5 | –0.54 | 0.3 | 25.0 | 0.11 | 0.4 | 12.5 | 0.33 |
| Сосняк черничный | |||||||||||||||
| P. longulus | – | – | – | 2.2 | 65.0 | 0.69 | 1.2 | 35.0 | 0.24 | – | – | – | – | – | – |
| T. cladonicolus | 10.3 | 34.3 | 0.03 | 10.1 | 25.7 | 0.02 | 15.7 | 40.0 | 0.33 | – | – | – | – | – | – |
| C. biurus | 0.3 | 33.0 | 0.01 | – | – | – | 0.7 | 67.0 | 0.71 | – | – | – | – | – | – |
| Heminothrus longisetosus | 0.6 | 28.0 | –0.12 | 2.2 | 72.0 | 0.77 | – | – | – | – | – | – | – | – | – |
| F. dentata | – | – | – | – | – | – | – | – | – | – | – | – | 0.4 | 100 | 1 |
| A. ovatus poppei | 1.5 | 18.5 | –0.36 | 6.0 | 55.4 | 0.57 | – | – | – | 9.7 | 26.1 | 0.63 | – | – | – |
| E. oblongus silvestris | 1.4 | 91.7 | 0.91 | 0.2 | 8.3 | –0.57 | – | – | – | – | – | – | – | – | – |
| S. acutidens duplex | – | – | – | 2.5 | 65.3 | 0.69 | 1.2 | 30.4 | 0.14 | – | – | – | – | – | – |
| C. labyrinthicus | 1.9 | 5.8 | –0.78 | 11.9 | 27.4 | 0.06 | 6.4 | 14.7 | –0.32 | 2.9 | 1.9 | –0.60 | 55.1 | 50.2 | 0.80 |
| C. marginatus | 15.0 | 38.6 | 0.13 | 16.3 | 32.0 | 0.17 | 14.1 | 27.7 | 0.07 | 2.9 | 1.7 | –0.65 | – | – | – |
| C. subarcticus | 31.9 | 30.0 | –0.06 | 35.2 | 25.3 | 0.01 | 43.8 | 31.4 | 0.16 | 63.2 | 13.3 | 0.32 | – | – | – |
| T. velatus | 12.7 | 62.3 | 0.54 | 2.0 | 7.5 | –0.61 | – | – | – | 13.2 | 14.5 | 0.36 | 0.4 | 0.6 | –0.89 |
| T. berlesei | 1.4 | 100 | 1.0 | – | – | – | – | – | – | – | – | – | – | – | – |
| Oribatula tibialis | 8.4 | 59.6 | 0.50 | 2.2 | 12.0 | –0.42 | – | – | – | – | – | – | – | – | – |
| O. (Zygoribatula) exilis | 0.6 | 83.3 | 0.82 | – | – | – | – | – | – | – | – | – | 0.4 | 16.7 | 0.29 |
| O. (Z.) propinqua | – | – | – | – | – | – | – | – | – | – | – | – | 8.5 | 100 | 1 |
| P. nemoralis | – | – | – | – | – | – | – | – | – | – | – | – | 33.1 | 100 | 1 |
| S. laevigatus | 9.0 | 62.0 | 0.54 | 2.3 | 12.3 | –0.40 | – | – | – | 1.7 | 2.7 | –0.49 | 0.4 | 0.9 | –0.85 |
Примечание. P – доля вида в структуре населения (%); q – степень участия местообитания в размещении вида (%); F – индекс относительной биотопической приуроченности вида; «–» – вид не был зарегистрирован; для табл. 2–4.
Таблица 3.
Распределение панцирных клещей по местообитаниям в ельнике черничном
| Вид/местообитание/ показатели биотопической приуроченности | Cladina arbuscula | Cladina rangiferina | Cetraria islandica | Hypogimnia physodes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P | q | F | P | q | F | P | q | F | P | q | F | |
| Phthiracarus laevigatus | – | – | – | 0.3 | 14.3 | –0.50 | 3.7 | 85.7 | 0.95 | – | – | – |
| Camisia biurus | – | – | – | 2.0 | 100 | 1 | – | – | – | – | – | – |
| Ceratoppia quadridentata | – | – | – | 0.3 | 11.1 | –0.60 | – | – | – | 3.0 | 88.9 | 0.92 |
| Adoristes ovatus poppei | – | – | – | 0.3 | 2.0 | –0.92 | 30.5 | 98.0 | 0.99 | – | – | – |
| Suctobelbella acutidens duplex | 3.5 | 84.6 | 0.87 | – | – | – | – | – | – | 0.7 | 15.4 | –0.27 |
| Carabodes labyrinthicus | 0.9 | 37.5 | 0.20 | – | – | – | – | – | – | 2.0 | 62.5 | 0.68 |
| C. marginatus | 10.7 | 30.3 | 0.05 | 15.6 | 51.8 | 0.37 | 12.4 | 17.9 | 0.13 | – | – | – |
| C. subarcticus | 56.6 | 52.2 | 0.47 | 35.0 | 37.7 | 0.10 | 21.8 | 10.1 | –0.20 | – | – | – |
| Tectocepheus velatus | 15.1 | 38.4 | 0.22 | 10.5 | 31.2 | –0.04 | 23.6 | 30.4 | 0.44 | – | – | – |
| Cymbaeremaeus cymba | – | – | – | – | – | – | – | – | – | 0.7 | 100 | 1 |
| Trichoribates berlesei | – | – | – | 0.5 | 50.0 | 0.34 | 0.6 | 25.0 | 0.33 | 0.4 | 25.0 | 0.03 |
| Chamobates pusillus | 3.8 | 32.4 | 0.09 | 4.3 | 43.3 | 0.21 | – | – | – | 3.4 | 24.3 | 0.08 |
| Diapterobates oblongus | – | – | – | – | – | – | – | – | – | 0.4 | 100 | 1 |
| Oribatula tibialis | 4.1 | 48.2 | 0.40 | 3.8 | 51.9 | 0.37 | – | – | – | – | – | – |
| O. (Zygoribatula) exilis | – | – | – | – | – | – | – | – | – | 3.0 | 100 | 1 |
| O. (Z.) propinqua | – | – | – | – | – | – | – | – | – | 1.1 | 100 | 1 |
| Phauloppia nemoralis | – | – | – | 0.3 | 0.4 | –0.98 | – | – | – | 81.0 | 99.6 | 0.99 |
| Scheloribates laevigatus | 1.9 | 7.8 | –0.65 | 16.4 | 79.2 | 0.77 | 6.2 | 13.0 | –0.06 | – | – | – |
Таблица 4.
Распределение панцирных клещей по местообитаниям в ельнике зеленомошном
| Вид/местообитание/показатели биотопической приуроченности | Cladina rangiferina | Cladina stellaris | Bryoria subcana | ||||||
|---|---|---|---|---|---|---|---|---|---|
| P | q | F | P | q | F | P | q | F | |
| Trhypochthonius cladonicolus | 7.1 | 58.5 | 0.73 | 3.6 | 41.5 | 0.35 | – | – | – |
| Ceratoppia quadridentata | – | – | – | 0.4 | 50.0 | 0.49 | 0.2 | 50.0 | –0.12 |
| Adoristes ovatus poppei | 4.1 | 73.7 | 0.85 | 1.1 | 26.3 | 0.02 | – | – | – |
| Eueremaeus oblongus silvestris | 4.4 | 31.2 | 0.34 | 7.0 | 68.8 | 0.73 | – | – | – |
| Graptoppia foveolata | – | – | – | – | – | – | 0.1 | 100 | 1 |
| Suctobelbella acutidens duplex | 2.9 | 100 | 1 | – | – | – | – | – | – |
| Carabodes labyrinthicus | 2.7 | 90.0 | 0.95 | – | – | – | 0.1 | 10.0 | –0.84 |
| C. marginatus | 0.6 | 4.4 | –0.66 | 9.3 | 95.6 | 0.97 | – | – | – |
| C. subarcticus | 53.1 | 48.5 | 0.61 | 40.2 | 51.2 | 0.51 | 0.1 | 0.3 | –0.99 |
| Tectocepheus velatus | 9.4 | 69.6 | 0.82 | 2.9 | 30.4 | 0.12 | – | – | – |
| Trichoribates berlesei | 0.9 | 20.0 | 0.05 | – | – | – | 1.1 | 80.0 | 0.51 |
| Diapterobates humeralis | – | – | – | – | – | – | 12.5 | 100 | 1 |
| D. oblongus | – | – | – | – | – | – | – | – | – |
| Mycobates tridactylus | – | – | – | – | – | – | 0.2 | 100 | 1 |
| Oribatula tibialis | 5.0 | 100 | 1 | – | – | – | – | – | – |
| O. (Zygoribatula) exilis | – | – | – | – | – | – | 0.8 | 100 | 1 |
| O. (Z.) propinqua | – | – | – | – | – | – | 1.1 | 100 | 1 |
| Phauloppia nemoralis | 0.6 | 0.2 | –0.98 | – | – | – | 83.5 | 99.8 | 0.99 |
| Scheloribates laevigatus | 1.2 | 2.9 | –0.77 | 27.9 | 94.9 | 0.96 | 0.3 | 2.2 | –0.97 |
Таблица 5.
Видовой состав и относительное обилие (%) панцирных клещей в почве различных растительных сообществ
| Вид/растительное сообщество | Сосняк | Ельник | ||
|---|---|---|---|---|
| лишайниково-зеленомошный | черничный | черничный | зеленомошный | |
| Trhypochthonius cladonicolus | 0.3 | – | – | – |
| Damaeus bituberculatus | 0.3 | 1.9 | 1.3 | 3.3 |
| Adoristes ovatus poppei | 0.8 | 1.8 | 3.6 | 3.9 |
| Dissorhina ornata s. str. | – | – | 4.4 | – |
| D. ornata globosa | 2.5 | – | – | 7.8 |
| Oppiella (Moritzoppiella) neerlandica | 0.9 | 1.4 | 1.7 | – |
| Oppiella nova | 2.1 | 22.3 | 21.4 | 11.4 |
| Quadroppia quadricarinata | – | 3.5 | 2.4 | 3.0 |
| Suctobelbella acutidens s. str. | 4.6 | – | 6.7 | – |
| S. longicuspis s. str. | 1.7 | 4.7 | – | – |
| S. singularis | 0.9 | 1.3 | – | 3.7 |
| S. baloghi | – | 6.2 | – | – |
| Carabodes marginatus | 3.6 | 1.5 | 0.6 | 0.9 |
| C. subarcticus | 20.9 | 3.1 | 3.6 | 3.9 |
| Tectocepheus velatus | 28.0 | 36.6 | 5.5 | 27.8 |
| Ceratozetella sellnicki | – | – | 5.1 | – |
| Chamobates pusillus | 3.0 | 1.7 | 17.4 | 9.8 |
| Oribatula tibialis | 0.2 | 1.1 | 1.3 | – |
| Scheloribates laevigatus | 17.4 | 2.1 | 7.1 | 5.8 |
В напочвенных лишайниках сосняка лишайниково-зеленомошного доминирующими были орибатиды Carabodes subarcticus Trägårdh, 1902, Scheloribates laevigatus (Koch, 1835), Trhypochthonius cladonicolus (Willmann, 1919); последний из них входил в число доминантов только в лишайниках рода Cladina, выступая в C. rangiferina эудоминантом (табл. 2). Группу субдоминантов составляли C. marginatus (Michael, 1884) и Tectocepheus velatus (Michael, 1880). В напочвенном Cetraria islandica располагалась наибольшая доля особей Ceratoppia quadridentata (Haller, 1882), Camisia biurus (Koch, 1839) и Adoristes ovatus poppei (Oudemans, 1906). Население панцирных клещей эпифитного лишайника H. physodes отличалось специфичностью. Доминировали по обилию Oribatula (Z.) propinqua (Oudemans, 1902) (эудоминант) и Carabodes labyrinthicus (Michael, 1879), субдоминантами выступали Diapterobates humeralis (Hermann, 1804) и Phauloppia nemoralis (Berlese, 1916). Большинство видов характеризовалось положительной биотопической приуроченностью к эпифиту; 7 видов из 15 были встречены только в этом лишайнике, в том числе доминанты и один субдоминант (D. humeralis), а также редкие и малочисленные виды Furcoppia dentata (Willmann, 1950), Graptoppia foveolata (Paoli, 1908), Suctobelbella acutidens s. str. (Forsslund, 1941), Liebstadia pannonica (Willmann, 1951). В слоевищах H. physodes обитала наибольшая доля особей таких видов, как P. nemoralis, Chamobates pusillus (Berlese, 1895) и Trichoribates berlesei (Jacot, 1929). Особи Diapterobates oblongus (L. Koch, 1879) были распределены равномерно между эпифитом и напочвенным лишайником C. islandica.
Группировки орибатид в напочвенных лишайниках рода Cladina в сосняке черничном характеризовались полидоминантной структурой населения. Устойчиво доминировали виды C. subarcticus и C. marginatus, большая доля особей всегда принадлежала к C. subarcticus (табл. 2). Вид T. cladonicolus в лишайнике C. stellaris был доминантом, в C. arbuscula и C. rangiferina – субдоминантом. В ядро доминантов в C. arbuscula входил также T. velatus. В качестве субдоминантов выступали S. laevigatus и Oribatula tibialis (Nicolet, 1855) – в C. arbuscula и A. ovatus poppei и C. labyrinthicus – в C. rangiferina. В Cetraria islandica в отличие от лишайников рода Cladina группу доминантов составляли лишь C. subarcticus и T. velatus, первый из них выступал в роли эудоминанта. Субдоминантом был A. ovatus poppei. Вид T. cladonicolus здесь не был найден. Только в слоевищах C. islandica были обнаружены четыре вида, в их числе C. quadridentata. Наиболее специфичным было население эпифитного лишайника H. physodes. Основу структуры населения здесь составляли C. labyrinthicus (эудоминант), P. nemoralis (доминант) и O. (Z.) propinqua (субдоминант), два последних в напочвенных лишайниках не были отмечены. Только в талломах H. physodes присутствовал вид F. dentata – субрецедент. В эпифите были встречены единичные экземпляры Suctobelbella acutidens duplex (Strenzke, 1950) с отрицательным индексом F.
В группы доминантов и субдоминантов в напочвенных лишайниках ельника черничного входили C. subarcticus, C. marginatus и T. velatus, при этом в лишайниках рода Cladina первым по обилию был C. subarcticus (в C. arbuscula – эудоминант). В лишайнике C. islandica первым доминантом был A. ovatus poppei, который в C. rangiferina был редким, в C. arbuscula отсутствовал. Вид S. laevigatus выступал доминантом в C. rangiferina и субдоминантом – в C. islandica. В талломах H. physodes было обнаружено 16 видов, абсолютно доминировал один вид – P. nemoralis (>80% всех особей; F = 0.99). В этом лишайнике находилась наибольшая доля особей C. quadridentata (F = 0.92) и C. labyrinthicus (F = 0.68). Почти для всех видов, за исключением одного – S. acutidens duplex, была определена положительная приуроченность к эпифиту. Только в H. physodes было обнаружено девять видов, в том числе Cymbaeremaeus cymba (Nicolet, 1855), Oribatula (Z.) exilis (Nicolet, 1855), O. (Z.) propinqua и D. oblongus, редкие или малочисленные.
В ельнике зеленомошном в слоевищах C. rangiferina эудоминантом был C. subarcticus. Группу субдоминантов составляли пять видов: Eueremaeus oblongus silvestris (Forsslund, 1956), A. ovatus poppei, T. cladonicolus, T. velatus и O. tibialis. В талломах C. stellaris два вида – C. subarcticus и S. laevigatus – были эудоминантом и доминантом соответственно и два вида – C. marginatus и E. oblongus silvestris – субдоминантами. В эпифитном лишайнике B. subcana преобладали по обилию два вида – P. nemoralis и D. humeralis, первый из них – эудоминант. Все остальные виды были редкими. Только в эпифите были обнаружены O. (Z.) propinqua, O. (Z.) exilis, D. humeralis, G. foveolata, Mycobates tridactylus Willmann, 1929 и Micreremus brevipes (Michael, 1888). Для видов C. quadridentata, C. labyrinthicus, C. subarcticus, S. laevigatus был определен отрицательный индекс F.
В напочвенных лишайниках сосняков и ельников кроме видов, перечисленных в табл. 2–4, были отмечены Euphthiracarus cribrarius (Berlese, 1904), Trhypochthonius tectorum s. str. (Berlese, 1896), Galumna lanceata (Oudemans, 1900), Parachipteria punctata (Nicolet, 1855).
В почве сосняка лишайниково-зеленомошного, сосняка черничного, ельника черничного и ельника зеленомошного было найдено 30, 28, 30 и 26 видов соответственно. Доминирующими были 2 или 3 вида из числа следующих: T. velatus, Oppiella nova (Oudemans, 1902), C. pusillus, S. laevigatus, C. subarcticus; выделялись группы субдоминантов из 1–6 видов (табл. 5). Вид S. laevigatus относился к доминантам или субдоминантам. Вид C. subarcticus, первый доминант в напочвенных лишайниках, был одним из доминантов в почве сосняка лишайниково-зеленомошного. Во всех сообществах были выделены группы субдоминантов, которые были составлены видами C. pusillus, Dissorhina ornata globosa (Paoli, 1908), Suctobelbella baloghi (Forsslund, 1958), S. acutidens s. str. (Forsslund, 1941), Ceratozetella sellnicki (Rajski, 1958) и S. laevigatus.
В почве наиболее часто встречались виды семейства Oppiidae: Dissorhina ornata s. str. (Oudemans, 1900), D. ornata globosa, Oppiella (Moritzoppiella) neerlandica (Oudemans, 1900), Rhinoppia subpectinata (Oudemans, 1900), Microppia minus (Paoli, 1908), O. nova, а также виды семейства Suctobelbidae: Suctobelbella acutidens s. str., S. acutidens sarekensis (Forsslund, 1941), S. baloghi, S. longicuspis Jacot, 1937, S. perforata (Strenzke, 1950), S. subtrigona (Oudemans, 1900). Обычным в почве был вид семейства Quadroppiidae Quadroppia quadricarinata (Michael, 1885). Представители указанных семейств обычны в почвах зональных и интразональных сообществ на Европейском Севере (Мелехина, 2011).
В почве были найдены виды, которые не встречались в лишайниках, – это Phthiracarus globosus (Koch, 1841), Acrotritia ardua s. str. (Koch, 1841), Mesotritia flagelliformis (Ewing, 1909), Oribotritia fennica Forsslund et Märkel, 1963, Heminothrus (P.) peltifer (Koch, 1839), Cepheus cepheiformis (Nicolet, 1855), Ceratozetes gracilis (Michael, 1884), Diapterobates notatus (Thorell, 1871), Fuscozetes fuscipes (Koch, 1844), Edwardzetes edwardsi (Nicolet, 1855), Eupelops occultus (Koch, 1835), E. plicatus (Koch, 1835), Neoribates roubali (Berlese, 1910), Achipteria coleoptrata s. str. (Linnaeus, 1758), A. italica (Oudemans, 1914).
Следовательно, разные группы местообитаний отличались составом доминирующих видов: в напочвенных лишайниках доминировали C. subarcticus, C. marginatus, T. velatus, T. cladonicolus, S. laevigatus, A. ovatus poppei; в эпифитных лишайниках – P. nemoralis, O. (Z.) propinqua, C. labyrinthicus, D. humeralis; в почве – O. nova, C. pusillus, C. sellnicki, T. velatus. В эпифитах сосняков первыми по обилию были O. (Z). propinqua и C. labyrinthicus, в ельниках (как в кустистых, так и в листоватых эпифитах) абсолютным доминантом был P. nemoralis.
Биотопическая приуроченность. Выявлены виды, которые проявили биотопическую приуроченность к серии местообитаний или к конкретному местообитанию. Так, вид C. subarcticus, который был доминирующим в напочвенных лишайниках, показал приуроченность к лишайникам рода Cladina. В отношении к C. islandica индекс F был положительным в сосняке черничном и отрицательным в ельнике черничном. В эпифитах C. subarcticus, как правило, не встречался; был отмечен единично в B. subcana, причем индекс F был отрицательным. В сосновой роще в окрестностях ст. Кево (Финляндия) C. subarcticus был приурочен к напочвенным Stereocaulon paschale и Cladina stellaris, при этом в эпифите не был обнаружен (Бязров, Мелехина, 1994). Он был найден в напочвенных лишайниках Северной Норвегии (Бязров, Мелехина, 1992), в лишайниковых пустошах Северной Финляндии (Solhǿy, Koponen, 1981), в эпифитах Московской обл. (Мелехина, Бязров, 1997).
Вид Carabodes labyrinthicus проявил приуроченность к эпифитам в сосняках и ельнике черничном (F = 0.68–1.0). В сосняке лишайниково-зеленомошном этот вид был найден исключительно в эпифите, в сосняке черничном также и в напочвенных лишайниках четырех видов, индекс F для трех из них (C. stellaris, C. arbuscula и C. islandica) был отрицательным. В ельниках C. labyrinthicus отмечался в напочвенных лишайниках рода Cladina, с низким обилием. C. labyrinthicus был одним из доминантов в слоевищах H. physodes в лесах Московской обл. (Мелехина, Бязров, 1997), присутствовал в эпифите и в напочвенных лишайниках в сосновой роще ст. Кево, Финляндия (Бязров, Мелехина, 1994), в напочвенном Cetraria nivalis в сухой мохово-лишайниковой пустоши в Норвегии (Бязров, Мелехина, 1992). Вилльманн (Willmann, 1933) назвал C. labyrinthicus ксерофильным видом, типичным для лишайников и коры деревьев, Сейд и Сюард (Seyd, Seaward, 1984) отнесли его к группе беспозвоночных, которые предпочитают лишайники как местообитания и как источник пищи. C. labyrinthicus был одним из наиболее многочисленных в эпифитах в Южной Бельгии (Andre, 1975, 1979, 1984, 1985); отмечался как характерный обитатель лишайникового покрова на деревьях в Северной Германии (Strenzke, 1952), Польше (Недбала, 1970), Дании (Gjelstrup, Sǿchting, 1979), в лесах Средних Альп (Pschorn-Walcher, Gunhold, 1957).
Для вида Carabodes marginatus в сосняках определена положительная приуроченность к видам рода Cladina и отрицательная – к Cetraria islandica. Единичные экземпляры C. marginatus обнаружены в эпифите H. physodes сосняка лишайниково-зеленомошного, при этом индекс F был отрицательным. В почве C. marginatus характеризовался невысоким обилием. Этот вид ранее отмечался как обитатель лишайниковых и мохово-лишайниковых группировок (Trave, 1963; Solhǿy, Koponen, 1981; Seyd, Seaward, 1984; Бязров, Мелехина, 1992; Мелехина, Бязров, 1997).
Вид Trhypochthonius cladonicolus проявил приуроченность к лишайникам рода Cladina в сосняках, наиболее высокий индекс F был определен для C. rangiferina. Индекс F по отношению к C. islandica был отрицательным. В эпифитах он не был найден. Вид T. cladonicolus Сейд и Сюард (Seyd, Seaward, 1984) включили в группу видов, которые предпочитают лишайники в качестве местообитания и источника пищи, а также обитают на других растениях.
Вид Phauloppia nemoralis был приурочен к эпифитам как в сосняках (F = 0.98–1.0), так и в ельниках (F = 0.99). В ельниках этот вид был абсолютным доминантом (>80% всех особей). Особей P. nemoralis мы находили в эпифитах среднетаежных лесов в разные сезоны года. Ранее отмечалось, что этот вид – характерный обитатель лишайников (Seyd, Seaward, 1984).
Вид Oribatula (Z.) propinqua, найденный исключительно в эпифитах, был эудоминантом и субдоминантом в H. physodes в сосняках и редким в ельниках. Вид O. (Z.) propinqua был одним из доминирующих в слоевищах H. physodes в сосновом лесу Брянской обл. (Штанчаева, 1997), был найден на коре сосен в Польше (Недбала, 1970).
Вид Scheloribates laevigatus проявил положительную приуроченность к лишайникам рода Cladina, иногда был отмечен в эпифитах как малочисленный или редкий, с отрицательным индексом F. Он был обычным в почве, иногда выступал доминантом. Известно, что S. laevigatus – эвритопный вид; ранее его находили в эпифитных лишайниках (Stary, 1988; Тарба, 1992; Мелехина, 1999; Мелехина, Бязров, 2007).
Вид Oribatula tibialis показал приуроченность к C. arbuscula в сосняке черничном и к C. rangiferina в ельнике зеленомошном. Он встречался в почве, где был рецедентом или субрецедентом. Вид O. tibialis известен как эвритопный, его часто обнаруживали в наскальных и эпифитных лишайниках Абхазии (Тарба, 1992).
Вид Tectocepheus velatus был приурочен к напочвенным лишайникам; в эпифитах встречался крайне редко, с отрицательным значением индекса F. В почве он был доминантом или субдоминантом. Вид T. velatus находили в эпифитных лишайниках в Московской обл. (Мелехина, Бязров, 1997) и Южной Чехии (Stary, 1988), а также в моховом покрове на деревьях и каменных стенах в Германии (Pschorn-Walcher, Gunhold, 1957).
Вид Adoristes ovatus poppei был приурочен к лишайнику C. islandica, в эпифитах не найден. В почве ельника зеленомошного он был субдоминантом, в других растительных сообществах был редким и малочисленным. В Польше вид A. poppei находили преимущественно в лесной подстилке, иногда на стволах деревьев (Недбала, 1970).
Вид Diapterobates humeralis был приурочен к эпифитным лишайникам как в сосняках, так и в ельниках, будучи доминантом и субдоминантом. Вид Trichoribates berlesei встречался как в напочвенных, так и в эпифитных лишайниках; показал приуроченность к эпифитам в сосняке лишайниково-зеленомошном и ельнике зеленомошном (F = 0.88 и 0.51 соответственно). Этот вид (назван как Trichoribates trimaculatus) мы находили в слоевищах H. physodes на стволах сосен в окрестностях Киева (Мелехина, Криволуцкий, 1993). В Бельгии он относился к группе доминантов в эпифитных лишайниках, как листоватых, так и кустистых (Andre, 1976, 1979, 1984, 1985), в Польше отмечался на стволах дуба (Недбала, 1970), в Германии часто встречался в мохово-лишайниковом покрове деревьев и был отнесен к группе ведущих форм с четким ксерофильным характером (Pschorn-Walcher, Gunhold, 1957).
Вид Ceratoppia quadridentata отмечался как в напочвенных, так и в эпифитных лишайниках, был приурочен к эпифиту H. рhysodes, в B. subcana встречался c низким обилием. В эпифитах хвойных лесов средней тайги мы обнаруживали C. quadridentata и зимой (Мелехина, 2001). Этот вид находили на стволах деревьев в Абхазии (Тарба, 1992).
Вид Cymbaeremaeus cymba был встречен исключительно в эпифите H. physodes ельника черничного. Ранее он отмечался в эпифитных лишайниках (H. physodes, Parmelia sulcata) и зимой (Мелехина, 2001). Вид C. cymba находили в эпифитах в окрестностях Киева (Мелехина, Криволуцкий, 1993), в Московской обл. (Бязров и др., 1971), Польше (Недбала, 1970), Бельгии (Andre, 1984); он был назван как типичный древеснообитающий вид в Абхазии (Тарба, 1992). Вилльманн (Willmann, 1931) назвал вид C. cymba арборикольным ксерофильным.
Немногочисленный вид Scapheremaeus palustris (Sellnick, 1924) был приурочен к эпифитному H. рhysodes ельника черничного. Этот вид отмечался в эпифитном покрове и зимой (Мелехина, 2001). В ряде случаев S. palustris находили в напочвенных лишайниках, единично; в почве он не был обнаружен.
Вид Oribatula (Z.) exilis был приурочен к эпифитам. Этот вид неоднократно отмечался в эпифитных лишайниках (Бязров и др., 1971; Мелехина, Криволуцкий, 1992; Штанчаева, 1997). В Московской обл. наибольшим обилием он отличался в листоватых эпифитах Parmelia sulcata и H. physodes (Мелехина, Бязров, 1997), в окрестностях Познани (Польша) присутствовал на коре дуба и ели (Недбала, 1970). В Абхазии O. (Z.) exilis отнесен к арборикольным видам (Тарба, 1992), Пшорн-Вальхер и Гунхольд (Pschorn-Walcher, Gunhold, 1957) включили его в группу ведущих видов с четким ксерофильным характером.
Вид Eueremaeus oblongus silvestris показал приуроченность к лишайникам рода Cladina, встречался в разных видах с разным обилием. Вид E. oblongus был назван типичным древеснообитающим в Абхазии (Тарба, 1992), был многочисленным в листоватых и кустистых эпифитах в Бельгии (Andre, 1979, 1984), часто встречался в эпифитах в Германии (Pschorn-Walcher, Gunhold, 1957). Штренцке (Strenzke, 1952) отнес его к группе ксерофильных видов.
Вид Diapterobates oblongus проявил приуроченность к эпифиту в сосняке лишайниково-зеленомошном (F = 0.86) и ельнике черничном (F = 1.0), к напочвенному C. islandica в сосняке лишайниково-зеленомошном (F = 0.57). Для ряда видов с небольшим обилием была выявлена биотопическая приуроченность к напочвенным или эпифитным лишайникам. Так, виды Phthiracarus longulus (Koch, 1841) и Heminothrus longisetosus (Willmann, 1925) показали приуроченность к C. rangiferina и C. stellaris в сосняках, Phthiracarus laevigatus (Koch, 1844) и Neoribates aurantiacus (Oudemans, 1914) – к C. islandica в ельнике и сосняке соответственно.
Экологические группы. Относительно исследованной серии местообитаний, типичных для еловых и сосновых лесов европейского Северо-Востока, были выделены экологические группы панцирных клещей. При определении принадлежности вида к той или иной экологической группе учитывали значения индексов F, q и P. В названиях экологических групп отражены приуроченность видов к определенной группе местообитаний и их численные характеристики. Пять групп видов связаны с лишайниками, две группы – с почвой.
К группе арборикольных доминирующих видов были отнесены C. labyrinthicus, O. (Z.) propinqua, P. nemoralis и D. humeralis, которые были доминантами и субдоминантами в эпифитах и проявили к ним положительную биотопическую приуроченность (F = 0.99–1.0). Арборикольные немногочисленные виды были обнаружены только в эпифитах с небольшим обилием: F. dentata, M. tridactylus, C. cymba, M. brevipes.
Группу арборикольно-гемиэдафических составили виды, которые обитали как в эпифитных, так и напочвенных лишайниках, везде были немногочисленны, это виды C. quadridentata, O. (Z.) exilis, T. berlesei, D. oblongus, S. palustris, Pergalumna nervosa (Berlese, 1914).
В группу гемиэдафических доминирующих видов вошли C. subarcticus, C. marginatus, T. velatus, T. cladonicolus, S. laevigatus, A. ovatus poppei, O. tibialis, которые были доминантами и субдоминантами в напочвенных лишайниках и проявили к ним биотопическую приуроченность; очень редко встречались в эпифитах. Гемиэдафические немногочисленные виды были найдены в напочвенных лишайниках, с положительным индексом F. Они были встречены также в почве с небольшим обилием. Некоторые из них были отмечены в эпифитах единично, с отрицательным индексом F. Это виды P. longulus, P. laevigatus, E. cribrarius, C. biurus, H. longisetosus, T. tectorum s. str., N. dorsalis, E. oblongus silvestris, S. acutidens duplex, G. lanceata, P. punctata, N. aurantiacus.
Виды, которые были обнаружены только в почве, были подразделены на две экологические группы: эпиэдафических и эуэдафических видов. В группу эпиэдафических вошли виды, которые по системе жизненных форм Криволуцкого (Криволуцкий и др., 1995) относятся главным образом к обитателям поверхности почвы и верхних горизонтов подстилки, а частично – к обитателям толщи подстилки. Это виды P. globosus, A. ardua s. str., M. flagelliformis, O. fennica, H. (P.) peltifer, C. cepheiformis, C. gracilis, C. sellnicki, D. notatus, F. fuscipes, E. edwardsi, E. occultus, E. plicatus, N. roubali, A. coleoptrata s. str., A. italica. В группу эуэдафических включены виды, которые относятся к жизненной форме обитателей мелких почвенных скважин. Это виды D. ornata s. str., D. ornata globosa, O. (O.) nova, O. (M.) neerlandica, R. subpectinata, M. minus, Q. quadricarinata, S. acutidens s. str., S. acutidens sarekensis, S. baloghi, S. longicuspis, S. perforata, S. subtrigona.
Разные авторы выделяли комплексы видов панцирных клещей, ассоциированных с лишайниками. Штренцке (Strenzke, 1952) назвал две синузии орибатид, характерные для мхов и лишайников, растущих на камнях: гигрофильную и ксерофильную. Ведущим видом этих двух синузий был Oribatula (Z.) exilis. В Германии Пшорн-Вальхер и Гунхольд (Pschorn-Walcher, Gunhold, 1957) выделили две синузии панцирных клещей, характерные для эпифитного покрова деревьев: ксерофильного гемиэдафона открытых ландшафтов и ксерофильного гемиэдафона закрытых ландшафтов. Каждая синузия складывалась из представителей нескольких экологических групп. Авторы выделили четыре экологические группы: ведущих видов с четким ксерофильным характером, ксерофильных форм открытых ландшафтов, ксерофильных форм закрытых ландшафтов и эвриэкологических видов, или видов-спутников. Колоф (Coloff, 1983) назвал две группы видов панцирных клещей, связанных с лишайниками на прибрежных скалах в Англии: растущими в супралиторальной зоне и непосредственно на берегу. Для орибатид, обитающих в эпифитных лишайниках Бельгии, Андрэ (Andre, 1984, 1985) выделил группы видов, которые он назвал зоосоциологическими классами. Видовой состав классов определяется, по мнению автора, морфологическим типом эпифита. Следовательно, в разных регионах для лишайников как местообитаний выделяются специфические комплексы видов панцирных клещей.
Викарирующие виды. Выявлены экологически викарирующие виды – представители одного рода, которые являются обитателями эпифитных лишайников в разных географических регионах. К викарирующим отнесены представители родов Phauloppia, Mycobates, Carabodes, Oribatula (Zygoribatula). Так, вид P. nemoralis, который доминировал в эпифитах в наших сборах, Сейд и Сюард (Seyd, Seaward, 1984) включили в группу беспозвоночных, предпочитающих лишайники как местообитания и как источник пищи. Другой вид этого рода – P. lucorum (Koch, 1841) – был характерным для эпифитов в Бельгии (Andre, 1984), отмечался на коре стволов дуба и сосны в Польше (Недбала, 1970). По данным З. Тарба (1992 г.), в Абхазии вид P. lucorum (назван как P. longiporosa) входил в число доминантов в эпифитных лишайниках на стволах бука и был отнесен к типичным древеснообитающим видам. Этот же вид, названный как P. conformis, был многочисленным в эпифитных лишайниках Германии (Pschorn-Walcher, Gunhold, 1957), наиболее часто встречался в слоевищах Cladonia spp. В южной Чехии в эпифитном покрове деревьев доминирующими были виды P. lucorum и P. nemoralis (назван как P. coineaui) (Stary, 1988). Вид Phauloppia sp. был первым по обилию в лишайнике H. physodes, собранном со стволов деревьев в Московской обл. (Мелехина, Бязров, 1997). Можно заключить, что представители рода Phauloppia типичны для эпифитов в разных регионах; виды P. nemoralis и P. lucorum являются экологически викарирующими.
К викарирующим можно отнести виды рода Mycobates – M. parmeliae (Michael, 1884) и M. tridactylus Willmann, 1929. В наших сборах для эпифитов был характерен вид M. tridactylus. В литературе наиболее часто в качестве обитателя лишайников упоминается вид M. parmeliae. Зелльник (Sellnick, 1949) указывал, что этот вид предпочитает лишайники в качестве среды обитания. Особей M. parmeliae находили в лишайниковом покрове парковых деревьев в Германии (Pschorn-Walcher, Gunhold, 1957), в лишайниках пяти видов, растущих на прибрежных скалах Шотландских островов (Coloff, 1983).
Представители семейства Carabodidae были многочисленными в кустистых эпифитах Южной Бельгии (Andre, 1979). В разных регионах разные виды рода Carabodes были приурочены к лишайникам как местообитаниям. Они численно доминировали в эпифитах (Hypogymnia physodes, Cladonia coniocraea) на разных древесных породах в елово-широколиственных лесах Подмосковья (Бязров и др., 1971; Бязров, 1988; Мелехина, Бязров, 1997). В наших сборах характерным обитателем эпифитов был C. labyrinthicus. Этот вид отличался высоким обилием в эпифитных лишайниках Южной Бельгии (Andre, 1975, 1979, 1984, 1985) и других регионов. В зоне хвойно-широколиственных лесов для эпифитов наряду с C. labyrinthicus были характерны C. marginatus и C. subarcticus (Мелехина, Бязров, 1997). В группы доминантов в эпифитных лишайниках Абхазии входил вид C. rugosior Berlese, 1916 (назван как C. femoralis) (Тарба, 1992). Таким образом, ряд видов C. labyrinthicus–C. marginatus–C. rugosior–C. subarcticus – пример экологического викариата.
К экологическим викариатам можно отнести виды рода Oribatula (Zygoribatula): O. (Z.) propinqua и O. (Z.) exilis. Вид O. (Z.) propinqua относился к числу доминантов в эпифитах в наших сборах; его находили в этом типе местообитаний и другие авторы (Недбала, 1970; Штанчаева, 1997). В эпифитах неоднократно отмечался другой вид этого рода – O. (Z.) exilis (Pschorn-Walcher, Gunhold, 1957; Бязров и др., 1971; Тарба, 1992; Мелехина, Криволуцкий, 1992; Мелехина, Бязров, 1997).
ЗАКЛЮЧЕНИЕ
Определены комплексы видов панцирных клещей, ассоциированных с напочвенными и эпифитными лишайниками в таежных лесах европейского Северо-Востока. Относительно исследованной серии местообитаний выделено пять экологических групп панцирных клещей, характерных для лишайников, и две группы собственно почвенных обитателей. Две группы – арборикольных доминирующих и арборикольных немногочисленных видов – отражают специфику видового состава орибатид в эпифитных лишайниках. К ним отнесены виды, отмеченные исключительно или главным образом в эпифитах; в напочвенных лишайниках, как правило, не найдены. В группы гемиэдафических доминирующих и гемиэдафических немногочисленных включены виды, которые были характерны для напочвенных лишайников. Одну группу арборикольно-гемиэдафических составили виды, обитающие как в напочвенных, так и в эпифитных лишайниках. Виды, которые встречались только в почве и не были отмечены в лишайниках, подразделены на группы эпиэдафических и эуэдафических.
Установлено, что таксономический состав комплексов орибатид, связанных с лишайниками (экологических групп, зоосоциологических классов, синузий, выделяемых разными авторами), отличается в разных природно-климатических условиях. В то же время выявлены виды, характерные для лишайников, которые определяют сходство видового состава панцирных клещей в данном типе местообитаний в различных регионах Северной Палеарктики. Так, специфику населения эпифитов составляют такие виды, как Carabodes labyrinthicus, Cymbaeremaeus cymba, Oribatula (Z.) propinqua, Micreremus brevipes, виды рода Mycobates и др. Общие для напочвенных лишайников в разных регионах виды Carabodes marginatus, C. subarcticus, Trhypochthonius cladonicolus. Как в напочвенных, так и в эпифитных лишайниках встречаются Oribatula (Z.) exilis, Eueremaeus oblongus silvestris, Trichoribates berlesei, Scheloribates laevigatus и др. Такие семейства, как Carabodidae, Oribatulidae, Punctoribatidae, Cymbaeremaeidae, Ceratozetidae, отличаются постоянством участия в сложении группировок панцирных клещей в лишайниках.
Сопоставление полученных результатов с литературными данными позволило выявить примеры экологического викариата видов орибатид в эпифитных лишайниках как местообитаниях. К викарирующим отнесены представители родов Phauloppia (P. nemoralis, P. lucorum), Mycobates (M. parmeliae, M. tridactylus), Carabodes (C. labyrinthicus, C. marginatus, C. rugosior, C. subarcticus), Oribatula (Zygoribatula) (O. (Z.) propinqua, O. (Z.) exilis). Следовательно, для лишайников как местообитаний характерен определенный набор видов панцирных клещей. Наибольшей специфичностью отличается население эпифитных лишайников.
Работа выполнена по теме государственного задания Института биологии ФИЦ Коми НЦ УрО РАН № AAAA-A17-117112850235-2 при частичной финансовой поддержке РФФИ (грант 18-29-05028 мк).
Список литературы
Атлас Республики Коми по климату и гидрологии. М.: Дрофа, 1997. 116 с.
Бязров Л.Г. Беспозвоночные животные в эпифитных лишайниках разных жизненных форм в лесах Подмосковья // Биология почв Северной Европы. М.: Наука, 1988. С. 149–154.
Бязров Л.Г., Мелехина Е.Н. Панцирные клещи в лишайниковых консорциях Северной Скандинавии (на примере Варангер-фьорда) // Бюл. МОИП. Отд. биол. 1992. Т. 97. Вып. 3. С. 73–79.
Бязров Л.Г., Мелехина Е.Н. Панцирные клещи в лишайниковых консорциях лесотундры Северной Лапландии (Финляндия) // Бюл. МОИП. Отд. биол. 1994. Т. 99. Вып. 3. С. 40–45.
Бязров Л.Г., Медведев Л.Н., Чернова Н.М. Лишайниковые консорции в широколиственно-хвойных лесах Подмосковья // Биогеоценологические исследования в широколиственно-еловых лесах. М., 1971. С. 252–270.
Голубкова Н.С., Бязров Л.Г. Жизненные формы лишайников и лихеносинузии // Ботан. журн. 1989. Т. 74. № 6. С. 794–805.
Криволуцкий Д.А., Лебрен Ф., Кунст М., Акимов И.А., Баяртогтох Б., Василиу Н., Голосова Л.Д., Гришина Л.Г., Карппинен Э., Крамной В.Я., Ласкова Л.М., Лакстэн М., Маршалл В., Матвеенко А.А., Нетужилин И.А., Нортон Р., Ситникова Л.Г., Смрж Я., Стари И., Тарба З.М., Шалдыбина Е.С., Эйтминавичуте И.С. Панцирные клещи: Морфология, развитие, филогения, экология, методы исследования, характеристика модельного вида Nothrus palustris C.L. Koch, 1839. М.: Наука, 1995. 224 с.
Леса Республики Коми / Под ред. Козубова Г.М. и Таскаева А.И. М.: Издат. центр «Дизайн. Информация. Картография», 1999. 332 с.
Мелехина Е.Н. Разнообразие панцирных клещей лишайниковых группировок таежной зоны Республики Коми // Изучение и охрана разнообразия фауны, флоры и основных экосистем Евразии / Под ред. Павлова Д.С., Шатуновского М.И. М.: Ин-т проблем экологии и эволюции им. А.Н. Северцова РАН, 2000. С. 184 –191.
Мелехина Е.Н. Биоразнообразие панцирных клещей – обитателей эпифитных лишайников таежной зоны Республики Коми // Фауна и экология беспозвоночных животных европейского Северо-Востока. Сыктывкар: Коми науч. центр УрО РАН, 2001. № 166. С. 111–120.
Мелехина Е.Н. Таксономическое разнообразие и ареалогия орибатид (Oribatei) европейского Севера России // Изв. Коми науч. центра УрО РАН. 2011. № 2 (6). С. 30–37.
Мелехина Е.Н., Бязров Л.Г. Многолетние изменения биоразнообразия панцирных клещей в эпифитных лишайниках Подмосковья // Динамика биоразнообразия животного мира: Сб. докл. совещания. М., 1997. С. 120–123.
Мелехина Е.Н., Криволуцкий Д.А. Многолетняя динамика населения микроартропод эпифитных лишайников в районе Чернобыльской АЭС // Радиоэкологические исследования в 30-километровой зоне аварии на Чернобыльской АЭС. Сыктывкар: Коми науч. центр УрО РАН, 1993. № 127. С. 60–72.
Недбала В.Я. Фауна древесных панцирных клещей окрестностей Познани // Орибатиды (Oribatei), их роль в почвообразовательных процессах. Вильнюс: Москлас, 1970. С. 103–112.
Ниеми Р., Вилкамаа П. Микроартроподы, обитающие на двух видах лишайников прибрежных скал финского архипелага // Биология почв Северной Европы. М.: Наука, 1988. С. 145–148.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
Потапов М.Б., Кузнецова Н.А. Методы исследования сообществ микроартропод: пособие для студентов и аспирантов. М.: КМК, 2011. 84 с.
Пыстина Т.Н. Лишайники таежных лесов европейского Северо-Востока (подзоны южной и средней тайги). Екатеринбург: УрО РАН, 2003. 239 с.
Тарба З.М. Микроартроподы скальных и эпифитных лишайников Абхазии // Вестн. зоологии. 1992. № 2. С. 10–14.
Штанчаева У.Я. Панцирные клещи (Аcariformes, Oribatida) напочвенных и эпифитных лишайников соснового леса // Acarina. 1997. Т. 5. № 1–2. С. 79–85.
Юдин Ю.П. Производительные силы Коми АССР. Растительный мир. М.; Л.: Изд-во АН СССР, 1954. Т. 3. Ч. 1. С. 332–360.
Andre H. Observations on Belgian corticolous mites // Found Univ. Luxemb. Ser. Notes Rech. 1975. V. 4. P. 1–31.
Andre H. Notes on the ecology of corticolous epiphytic dwellers. 1. The mite fauna of fruticose lichens // Recent Adv. Acarol. 1979. V. 1. P. 551–557.
Andre H. Notes on the ecology of corticolous epiphytic dwellers. 3. Oribatida // Acarologia. 1984. V. 25. № 4. P. 385–395.
Andre H. Associations between corticolous microarthropod communities and epiphytic cover on bark // Holarctic Ecol. 1985. V. 8. № 2. P. 113–119.
Coloff M.J. Oribatid mites associated with marine and maritime lichens on the Islands of Great Cumbrae // Glasg. Nat. 1983. V. 20. P. 347–359.
Gjelstrup P., Søchting U. Criptostigmatid mites (Acarina) associated with Ramalina siliquosa (Lichens) on Bornholm in the Baltic // Pedobiologia. 1979. Bd 19. S. 237–245.
Pschorn-Walcher H., Gunhold P. Zur Kenntnis der Tiergemeinschaft in Moos- und Flechtenrasen an Park- und Waldbäumen // Z. Morph. Ökol. Tiere. 1957. Bd 46. H. 3. S. 343–354.
Santesson R. The lichens and lichenicolous fungi of Sweden and Norway. Lund: ISBN, 1993. 240 p.
Seyd E.L. The moss mites of the Cheviot (Acari: Oribatei) // Biol. J. Linn. Soc. 1988. V. 34. № 4. P. 349–369.
Seyd E.L., Seaward M.R.D. The association of oribatid mites with lichens // Zool. J. Linn. Soc. 1984. V. 80. № 4. P. 369–420.
Solhøy T., Koponen S. Oribatei fauna (Acari) on alpine heath at Kevo, Finland // Repts Kevo Subarctic Res. Stat. 1981. V. 17. № 1. P. 41–43.
Stary J. Pancirnici (Acari: Oribatida) nekterych vrchovist na Sumave, jizni Cechy // Sbor. Jihoces. Muz. v Ces. Budejovicich Prir. Vedy. 1988. № 28. S. 99–107.
Strenzke K. Untersuchungen über die Tiergemeinschaften des Bodens: die Oribatiden und ihre Synusien in den Böden Norddeutschlands // Zoologica. 1952. Bd 37. H. 104. S. 81–173.
Subías L.S. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Graellsia, 60 (número extraordinario). 2004. 305 p. Online version accessed in march 2019, 536 p. http://bba.bioucm.es/cont/docs/RO_1.pdf
Trave J. Ecologie et biologie des Oribates (Acariens) saxicoles et arboricoles // Vie et Milleu (Suppl.). 1963. V. 14. 267 p.
Willmann C. Acari aus dem Moosebruch // Z. Morph. Okol. 1933. V. 27. № 2. P. 373–383.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая