

Известия РАН. Серия биологическая, 2020, № 6, стр. 596-603
Строение репродуктивных органов Aristolochia tomentosa (Aristolochiaceae)
О. В. Наконечная 1, *, Г. В. Белова 1, Т. Ю. Горпенченко 1, В. М. Казарин 1, В. М. Локтионов 1
1 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
690022 Владивосток, просп. 100-летия Владивостока, 159, Россия
* E-mail: markelova@biosoil.ru
Поступила в редакцию 10.12.2019
После доработки 24.12.2019
Принята к публикации 02.02.2020
Аннотация
Изучены динамика цветения, строение цветка, гинецея, андроцея редкой реликтовой лекарственной лианы Aristolochia tomentosa Sims. Установлено, что развитие от стадии едва заметного бутона до открытия цветка длится 20, цветения одного цветка – 5 сут. Отмечено, что цветки протогиничные, пыльцевые зерна A. tomentosa безапертурные, сфероидально-сплюснутые, полярный диаметр 30.72 ± 1.89, экваториальный – 33.21 ± 0.56 мкм. Обнаружено, что анатомическое строение гинецея и андроцея A. tomentosa близко к таковому у азиатского вида A. manshuriensis и отражает консервативность признаков, сохранившихся у видов после отклонения от предковой формы.
Представители рода Aristolochia (Aristolochiaceae) – реликты третичного периода – найдены в отложениях с эоцена, в плиоцене на территориях Европы, Северной Америки, Восточной Азии (Колаковский, Дорофеев, 1963). Виды Aristolochia представлены разными жизненными формами, в частности лианами, кустарниками и корневищными травами (Razzak et al., 1992; Endress, 1994). Большинство видов – источники лекарственных веществ, которые используют в восточной медицине (Zhou et al., 2011).
Ранее была выдвинута гипотеза о расхождении предков травянистых восточно-североамериканских и древесных видов рода Aristolochia во время олигоцена (González et al., 2014). Эти авторы выделили два кластера: неотропический и североамериканский/восточно-азиатский. Всплеск видообразования у представителей Aristolochia в каждом из двух кластеров произошел в миоцене. При расхождении континентов произошло разделение видов внутри этих кластеров. В результате два генетически наиболее близких один к другому представителя рода Aristolochia секции Siphisia (A. tomentosa Sims. и A. manshuriensis Kom.) оказались в разных точках земного шара, на разных континентах (González, Stevenson, 2002; Ohi-Toma et al., 2006; González et al., 2014).
A. tomentosa – древесная лиана, обитающая в 14 штатах США (Алабама, Арканзас, Джорджия, Иллинойс, Индиана, Канзас, Кентукки, Луизиана, Миссури, Оклахома, Северная Каролина, Теннесcи, Техас, Флорида) (Barringer, Whitteremore, 1997). Несмотря на широкий ареал, этот вид редкий (Ward et al., 2003), что связано с приуроченностью к определенным местообитаниям. Растения A. tomentosa произрастают на аллювиальных почвах вдоль рек и ручьев на высоте до 500 м над уровнем моря (Pfeifer, 1966; Joyner, Chester, 1994; Barringer, Whitteremore, 1997) и на открытых участках под пологом леса вдоль дорог (Burk, Zebryk, 2001).
A. manshuriensis – древесная лиана, эндемик Маньчжурского флористического района с ограниченным фрагментированным ареалом (Куренцова, 1968). Она произрастает в Китае, на Корейском п-ове; в России небольшие популяции находятся на юго-западе Приморского края (Воробьев, 1968; Oh, Pak, 2001). Вид занесен в Красную книгу РСФСР (1988) как “исчезающий”. Условия обитания данного вида во многом сходны с условиями обитания A. tomentosa на территории Северной Америки.
При анализе данных о генетической близости A. tomentosa и A. manshuriensis (González, Stevenson, 2002; Ohi-Toma et al., 2006; González et al., 2014), произрастающих на разных континентах, возникает вопрос о сохранении от предковой формы черт морфологически сходных или эволюционировавших с течением времени у исследуемых видов. Существует гипотеза, что у видов секции Siphisia присутствуют черты стазиса (черты стазиса – определенные характеристики, которые мало изменились или не изменились совсем в течение тысяч лет) или что некоторые характеристики видов претерпели значительные изменения (выработали адаптации), так как растения переместились в новые климатические и растительные зоны (Adams et al., 2005b). Этими авторами было показано, что в прорастании семян видов в секции Siphisia обнаружены и черты стазиса, и адаптационные изменения, которые проявляются в особенностях покоя и прорастания семян, а также в разных требованиях к температурным режимам, которые необходимы для развития зародыша (Adams et al., 2005а, b).
Данные о биологии размножения A. tomentosa фрагментарны. Исследования особенностей опыления цветков, строения репродуктивных органов в известной нам литературе отсутствуют. Полученные характеристики репродуктивных органов A. tomentosa могут быть полезны для уточнения вопросов систематики растений семейства Aristolochiaceae.
Цель работы – исследовать цветение, морфологические и анатомические особенности строения гинецея и андроцея A. tomentosa в условиях интродукции.
МАТЕРИАЛЫ И МЕТОДЫ
A. tomentosa – одревесневающая лиана, высота которой достигает 25 м (рис. 1) (Barringer, Whitteremore, 1997). Листья сердцевидные, заостренные или притупленные на вершине, мелкосердцевидные в основании (шириной 8–15, длиной 9–20 см), сильноопушенные сверху и снизу. Цветки одиночные, расположены в пазухах листа, по форме коленчатые (Pfeifer, 1966), пурпурные, желтые и зеленые, цветут в конце весны и летом (Barringer, Whitteremore, 1997). Камера (мешочек) околоцветника шаровидная или цилиндрическая, 0.5–1 × 0.5–0.8 см; сиринкс отсутствует; трубка изогнутая, 1–3 (1.5) × 0.5 см (Barringer, Whitteremore, 1997). Отгиб околоцветника 3-лопастной, лопасти треугольные (2 × 2 см). Гиностемий 3-лопастной, высотой 3 и шириной 6 мм, пыльников 6 в трех группах (Pfeifer, 1966).
Плоды A. tomentosa были собраны с нескольких лиан, произрастающих на берегах р. Камберленд и Лонг-Понд в округе Монтгомери (штат Теннесси), в ноябре 2000 г. и октябре 2001 г. Семена были пророщены в лабораторных условиях и перенесены в условия закрытого грунта в частной коллекции с неконтролируемыми условиями микроклимата – отоплением в осенне-зимний период, регулярным ручным поливом. Температура воздуха летом и зимой 20–28 и 10–15°С соответственно. Растения выращивали в контейнерах с почвенной смесью. Одна лиана достигла генеративного состояния. Ее цветки использовали для исследования динамики цветения, анатомического строения гинецея и андроцея. Систематическое положение A. tomentosa и других видов, использованных в обсуждении результатов, соответствует опубликованным ранее данным (González, Stevenson, 2002). Для определения сроков цветения отмечали даты раскрывания и увядания 50 цветков. Для получения морфометрических данных измеряли по 30 цветков A. tomentosa в 2017 и 2018 гг. и 20 цветков A. manshuriensis.
Для гистологического исследования использовали 10 цветков в женской фазе цветения (пыльники закрыты). Цветки и завязи фиксировали в смеси, содержащей формалин, уксусную кислоту и этиловый спирт (FAA) (7, 7 и 100 мл соответственно), затем заливали в парафин по общепринятой методике (Паушева, 1970). Срезы толщиной 8–10 мкм окрашивали гематоксилином и альциановым синим по модифицированным методикам (Камелина и др., 1992; Жинкина, Воронова, 2000). Для определения фертильности пыльцевых зерен использовали окраску ацетокармином (5 цветков в женской и 5 цветков в мужской фазах цветения (пыльники открыты)) (Паушева, 1970). При интерпретации результатов применили антморфологический метод изучения пыльцы (Хохловым, 1978). Препараты анализировали под микроскопом Axioscop 40 (Zeiss, Германия). Для проведения электронного сканирующего микроскопического исследования поверхности пыльцевого зерна пыльники и пыльцевые зерна фиксировали в глютаровом альдегиде в течение 3 сут, перед проведением исследования их отмывали от фиксатора проточной водой. Далее образцы высушивали, монтировали на специальные подложки, напыляли углеродом и исследовали под микроскопом EVO (Zeiss). Восприимчивость рыльца определяли по методике Робинсон (Robinsohn, 1924).
Для получения фотографий репродуктивных органов и семян использовали микроскоп Zeiss SteREO Discovery.V12 с цифровой видеокамерой AxioCam MRc и программное обеспечение CombineZM. Микроскопическое оборудование было предоставлено Центром коллективного пользования “Биотехнология и генетическая инженерия” ФНЦ биоразнообразия ДВО РАН, Владивосток. Полученные данные обрабатывали с использованием компьютерного пакета стандартных программ Excel. Результаты представлены как средние значения со стандартной ошибкой.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Растения A. tomentosa в условиях закрытого грунта достигают раннего генеративного состояния через 17 лет после посадки проростков. К генеративному периоду длина лианы увеличивается более чем на 10 м. Растения A. tomentosa листопадные (рис. 1а). Разворачивание листьев происходит одновременно с цветением. Листья сердцевидные длиной 13.89 ± ± 0.46, шириной 12.25 ± 0.39, глубиной выемки пластинки листа 2.20 ± 0.07, длиной черешка 3.85 ± ± 0.11 см.
В условиях закрытого грунта цветение начинается в конце февраля, заканчивается в середине мая. В пазухе листа развивается цветок и/или укороченный генеративный побег, несущий цветок. Развитие от стадии едва заметного бутона до стадии открытия цветка происходит за 20 сут. Цветки на цветоножках без прицветников, цветоножки длиной 20.83 ± 1.78 мм. Околоцветник светло-зеленый, зигоморфный (рис. 1б, табл. 1); в основании находится вздутие (камера), трубка изогнута, диаметр зева 4.98 ± 0.17 мм. Отгиб околоцветника ярко-желтый. С внутренней стороны трубка околоцветника окрашена бордовыми пятнами на белом фоне (рис. 1в–д); в области камеры окраска темно-бордовая, вокруг гиностемия околоцветник внутри почти белый (рис. 1е).
Рис. 1.
Aristolochia tomentosa. а – растение с цветками; б – цветок на третий день цветения; в – части околоцветника (часть отгиба, зева, трубки); г – участок трубки с восковым налетом; д – трубка в области сгиба и часть камеры; е – камера с волосками внутри; ж – гиностемий в женской фазе цветения; з – гиностемий в мужской фазе цветения; и – завязь с белым опушением; к – семена. Размер линейки: а – 100, б – 5, в – и – 1, к – 2 мм.

Цветки открываются в течение всего дня, чаще в утренние часы. Период цветения одного цветка – 5 сут. За весь период цветения на лиане открылось >100 цветков. Цветки обоеполые, протогиничные. Репродуктивные органы (пыльники, рыльце и пестик) срастаются вместе с образованием гиностемия (рис. 1ж, з, 2), его размеры приведены в табл. 1. Секрет на рыльце пестика появляется в день открытия цветка; цветок готов к опылению. Пыльники раскрываются через 2.5–3 сут после раскрытия цветка. К этому времени лопасти рыльца смыкаются (рис. 1з), секрет на них отсутствует; опыления не происходит. Через 5 сут после открытия цветка околоцветник высыхает и опадает.
Таблица 1.
Характеристики цветков Aristolochia tomentosa и A. manshuriensis
| Характеристика цветков | A. tomentosa | A. manshuriensis |
|---|---|---|
| Период от появления бутона до распускания околоцветника, сут | 20 | 18–20* |
| Период цветения отдельного цветка, сут | 5 | 7–9* |
| Высота отгиба околоцветника, мм | 18.31 ± 0.73 | 27.70* |
| Ширина отгиба околоцветника, мм | 19.63 ± 0.73 | 28.70* |
| Длина трубки околоцветника, мм | 17.27 ± 0.74 | 73.40* |
| Ширина трубки околоцветника, мм | 4.81 ± 0.24 | 10.70* |
| Длина камеры околоцветника, мм | 8.27 ± 0.23 | 17.10* |
| Ширина камеры околоцветника, мм | 8.03 ± 0.17 | 15.30* |
| Высота гиностемия, мм | 2.25 ±0.09 | 4.86 ± 0.15 |
| Ширина гиностемия, мм | 3.43 ± 0.13 | 4.61 ± 0.12 |
| Длина завязи, мм | 14.61 ± 0.90 | 11.78 ± 0.38 |
| Ширина завязи, мм | 3.63 ± 0.09 | 3.52 ± 0.12 |
| Полярный диаметр пыльцевого зерна, мкм | 30.72 ± 1.89 | 44.00** |
| Экваториальный диаметр пыльцевого зерна, мкм | 33.21 ± 0.56 | 48.50** |
Рыльце A. tomentosa трехлопастное. Эпидерма рыльца представлена округлыми мелкими (до 15 мкм) плотно прижатыми одна к другой клетками (рис. 2б). Участки, воспринимающие пыльцу, находятся на апикальной и латеральной частях рыльца в области, прикрывающей пыльники (рис. 2б, в). Эти участки представлены одним слоем вытянутых в высоту слизеобразующих клеток (до 50 мкм), заполненных секреторными каплями. Нижние слои рыльца состоят из крупных паренхимных (до 70 мкм) многоугольных клеток. Столбик открытый, его составляют стилодии шести плодолистиков. Канал столбика выстлан стигматоидными клетками. Проводящие пучки столбика расположены по два в каждом из стилодиев, проходят вдоль столбика. В средней части столбика проводящие пучки разделяются и отходят к пыльникам.
Рис. 2.
Строение репродуктивных органов Aristolochia tomentosa. а – поперечный срез завязи; б – продольный срез части рыльца и пыльника; в – участок рыльца с волосками, воспринимающими пыльцу; г – участок завязи с проводящим пучком; д – поперечный срез пыльников с пыльцевыми зернами; е, ж – пыльцевые зерна; з – поверхность пыльцевого зерна; и, к – семязачаток. Масштаб: а – 500; б, в, д, и – 100; г, к – 50; е – 10; ж – 2; з – 1 мкм.
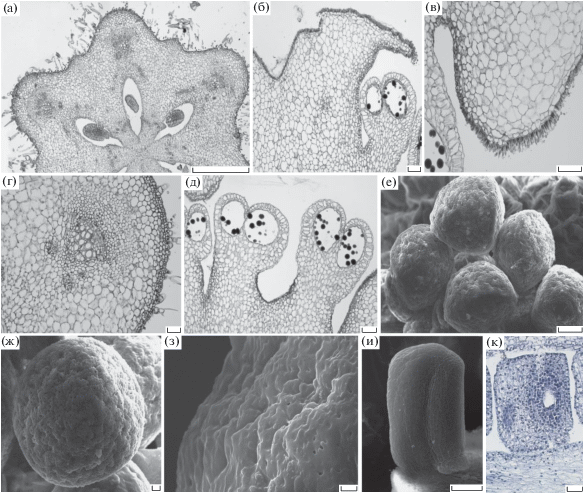
Завязь синкарпная 6-гнездная, нижняя, цилиндрическая (рис. 1и, 2а, табл. 1). Мелкие низкопризматические клетки составляют эпидерму завязи, ниже расположен слой клеток субэпидермы, соответствующий по размеру клеткам эпидермы (рис. 2а, г). В завязи длина клеток паренхимы достигает 50 мкм (рис. 2г). Проводящая система состоит из трех проводящих пучков в дорсальной части каждого плодолистика и небольших проводящих тяжей в вентральной части на границе срастания плодолистиков. Проводящие пучки A. tomentosa коллатеральные закрытые. Внутри завязи располагаются многочисленные семязачатки размером ~300 мкм (рис. 2а). Они анатропные, крассинуцеллятные, битегмальные, с массивным фуникулусом длиной 274.28 ± 1.80 мкм, шириной 104.17 ± 1.45 мкм (рис. 2и, к). Наружный интегумент короче внутреннего, состоит из двух слоев клеток, внутренний – из трех слоев. Нуцеллус эллипсоидной формы, длиной 163.69 ± 7.51, шириной 88.09 ± 2.13 мкм.
Рис. 3.
Пыльцевые зерна Aristolochia tomentosa. а–г – фертильные (ф) и дефектные (д) пыльцевые зерна; д–ж – прорастание (пр) пыльцевой трубки. Масштаб: а–е – 20, ж – 100 мкм.
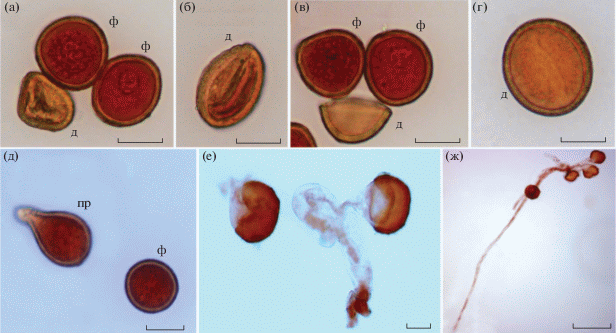
Пыльники биспорангиатные (рис. 2д), по 4 шт. на каждой из трех лопастей рыльца (рис. 1ж, з). Пыльники трехслойные. Наружный слой (эпидерма) состоит из мелких клеток, в среднем слое клетки крупные высокопризматические. Полость пыльника выстилают мелкие клетки тапетума (рис. 2б, в, д). Пыльцевые зерна округлой формы, безапертурные (рис. 2е, ж, табл. 1). Отношение длины полярного диаметра к экваториальному (коэффициент P/E) составляет 1.10. Согласно данному значению коэффициента по классификации Уокера и Дойла (Walker, Doyle, 1975) по форме такие зерна сфероидально-сплюснутые. Экзина бугорчатая с мелкими перфорациями (рис. 2з). В пыльниках 24 560 ± 990 пыльцевых зерен. Доля фертильности пыльцевых зерен в пыльниках в женскую фазу цветения цветка составила 97.79 ± 0.71, в мужской фазе – 60.34 ± 14.12%. Доля дефектности в женской фазе цветения низкая (2.20 ± 0.71%). Варианты дефектных пыльцевых зерен представлены на рис. 3а–г. Пророщенных пыльцевых зерен в пыльниках в женской фазе цветения цветка нет, в то время как мужская фаза цветения характеризовалась наличием пророщенных пыльцевых зерен с пыльцевыми трубками разной длины (29.24 ± 9.07%) (рис. 3д–ж).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Согласно приведенным ранее данным (Barringer, Whitteremore, 1997) в природных условиях A. tomentosa цветет поздней весной и летом. Более ранние сроки цветения в нашем эксперименте могут быть связаны с оптимальными условиями произрастания лианы в частной коллекции. Другие представители секции Siphisia, например североамериканский вид A. macrophylla Lam. и азиатский вид A. manshuriensis, в природных популяциях цветут также поздней весной, в начале лета (Barringer, Whitteremore, 1997; Наконечная и др., 2014). При этом у A. manshuriensis период цветения короткий – 3 нед (Наконечная и др., 2014).
Длительность развития бутона от момента появления в пазухе листа до момента открывания цветка у видов рода Aristolochia разная. Так, развитие бутона у травянистых видов A. fimbriata Cham. и A. elegans Mast. (секция Gymnolobus) происходит быстрее, чем у древесного вида A. tomentosa (10 и 14 сут соответственно) (Наконечная и др., 2015, 2016). В то же время у древесного вида A. manshuriensis длительность периода развития бутона такая же, как у A. tomentosa (Наконечная и др., 2014). Скорость развития бутона не зависит от размера цветка, но, вероятно, у травянистых форм она выше, и бутон открывается быстрее.
По окраске отгиба цветков A. tomentosa был выявлен полиморфизм. Ранее было отмечено (Pfeifer, 1966), что цветки данного вида пурпурные, желтые и зеленые. На нашей лиане цветки имели желтый отгиб околоцветника, как и цветки, описанные ранее (Barringer, Whitteremore, 1997). Был выявлен полиморфизм окраски отгиба околоцветника у A. manshuriensis, A. anguicida Jacq., A. oblongata Jacq. и A. pilosa Kunth (Наконечная, Нестерова, 2013; González, Pabón-Mora, 2015). Разнообразная окраска, вероятно, имеет приспособительный характер для привлечения бóльшего видового разнообразия потенциальных опылителей.
Размеры частей околоцветника цветков (камеры, трубки, отгиба) A. tomentosa, раскрывшихся на лиане в условиях интродукции, находятся в диапазоне значений, приведенных ранее для растений из природных популяций (Pfeifer, 1966; Barringer, Whitteremore, 1997). Размеры частей околоцветника будут ограничивать потенциальных опылителей, так как ширина зева в верхней части трубки и ее диаметр не превышают 5 мм. Опылители должны быть достаточно мелкими, чтобы пролезть в отверстие. Ранее на цветках A. tomentosa были отмечены жуки Canifa pallipes (Melsheimer) (Scraptiidae) (Graham, 2010), мухи Megaselia fungicola (Coquillett) (Phoridae) (Robertson, 1928). На цветках других видов Aristolochia также были отмечены мухи, участвующие в опылении цветков (Berjano et al., 2009; Наконечная и др., 2014).
Cтроение репродуктивных органов у представителей рода Aristolochia сходно. У видов секции Siphisia, к которой относится и A. tomentosa, рыльце трехлопастное, сформированное срастанием боковых поверхностей стилодий плодолистиков. Завязь A. tomentosa, как и все растение, имеет густое опушение, отсутствующее у других видов рода. Вероятно, данный признак можно использовать как систематический.
Морфологическое и анатомическое строения гинецея и андроцея A. tomentosa сходны с таковыми у A. manshuriensis (Наконечная и др., 2014). Строение проводящей системы завязи A. tomentosa имеет черты, сходные с отмеченными ранее для A. manshuriensis (Наконечная и др., 2006), за исключением строения центрального дорсального проводящего пучка плодолистика, который коллатеральный открытый. Строение семязачатка A. tomentosa – систематический признак, совпадающий с таковым у других видов рода (A. elegans, A. erecta L., A. fimbriata, A. grandiflora Sw. и т.д.) (Nair, Narayanan, 1962; Igersheim, Endress, 1998; González, Rudall, 2003).
Пыльцевые зерна A. tomentosa безапертурные, как у других представителей рода (Nair, Narayanan, 1962; González et al., 2001; Lu et al., 2015; Nakonechnaya, Kalachev, 2018; и др.). Между тем наблюдаемая сплюснутость пыльцевых зерен A. tomentosa отличает их от большинства исследованных видов Aristolochia, за исключением A. adalica Tosunoğlu & Malyer, A. hirta H. Buek (Tosunoglu, Malyer, 2014) и A. manshuriensis (Nakonechnaya, Kalachev, 2018), у которых можно наблюдать то же соотношение P/E.
По размеру пыльцевые зерна A. tomentosa меньше, чем у большинства исследованных видов рода Aristolochia, но близки по размерам к пыльце A. contorta (27.0–30.0) (Lu et al., 2015), A. fimbriata (25.0) (González et al., 2001). При этом у генетически наиболее близкого к A. tomentosa азиатского вида A. manshuriensis, произрастающего на территории России, пыльцевые зерна крупнее в 1.5 раза (Nakonechnaya, Kalachev, 2018).
Для A. tomentosa выявлены высокая фертильность (~100%) и, соответственно, низкая дефектность пыльцевых зерен (~2%). Согласно мнению С.С. Хохлова (Хохлова, 1968), дефектной считается пыльца, степень дефектности которой >11%. Для исследованного вида дефектность пыльцевых зерен в цветках женской фазы цветения низкая, что свидетельствует о полноценном функционировании андроцея. В то же время изменение показателей фертильности и дефектности в мужской фазе цветения, когда многие пыльцевые зерна прорастают, свидетельствует о быстром изменении в морфологическом состоянии пыльцевых зерен.
В целом некоторые особенности A. tomentosa (одновременное разворачивание листьев и цветение, срок развития бутона от появления до раскрывания цветка, форма цветка с трехлопастным отгибом, наличие трехлопастного рыльца, анатомическое строение репродуктивных органов, форма пыльцевых зерен) близки к таковым у A. manshuriensis (табл. 1), что может отражать черты предковой формы, сохранившиеся в неизменном состоянии на протяжении тысячелетий. Вероятно, выявленные характеристики – черты эволюционного стазиса у североамериканского и азиатского представителей рода Aristolochia. В то же время по размерам околоцветника, гиностемия, завязи, пыльцевых зерен виды различаются (табл. 1). Кроме того, наличие густого опушения цветков и завязей A. tomentosa отличает вид от других представителей рода. Данные характеристики могут быть результатом адаптации к измененным условиям существования и наличия иного состава опылителей.
ЗАКЛЮЧЕНИЕ
Изучены динамика цветения, строение цветка и репродуктивных органов. Отмечено, что общий план строения репродуктивных органов A. tomentosa характерен для других представителей рода. Сходное строение репродуктивных органов у видов рода отражает консервативность данных признаков, сохранившихся у видов после отклонения от предковой формы, и может отражать черты стазиса у A. tomentosa. Между тем различия в динамике цветения, форме околоцветника, размерах частей цветка и репродуктивных органов, пыльцевых зерен позволяют безошибочно идентифицировать A. tomentosa среди других видов рода.
Работа выполнена при финансовой поддержке ДВО РАН (грант № 18-4-049).
Список литературы
Воробьев Д.П. Дикорастущие деревья и кустарники Дальнего Востока. Л.: Наука, 1968. 275 с.
Жинкина Н.А., Воронова О.Н. О методике окраски эмбриологических препаратов // Ботан. журн. 2000. Т. 85. № 6. С. 168–171.
Камелина О.П., Проскурина О.Б., Жинкина Н.А. К методике окраски эмбриологических препаратов // Ботан. журн. 1992. Т. 77. № 4. С. 93–96.
Колаковский А.А., Дорофеев П.И. Порядок 3. Aristolochiales // Основы палеонтологии. Справочник для палеонтологов и геологов СССР. Голосеменные и покрытосеменные. М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1963. С. 439.
Красная книга РСФСР. Растения. М.: Росагропромиздат, 1988. 590 с.
Куренцова Г.Э. Реликтовые растения Приморья. Л.: Наука, 1968. 72 с.
Наконечная О.В., Нестерова С.В. Примитивные признаки и адаптации реликтовой лианы Aristolochia manshuriensis // Вестн. КрасГАУ. 2013. Вып. 1. С. 40–47.
Наконечная О.В., Нестерова С.В., Воронкова Н.М. Биология размножения Aristolochia elegans (Aristolochiaceae) в условиях ex situ // Ботан. журн. 2016. Т. 101. № 10. С. 1190–1201.
Наконечная О.В., Горпенченко Т.Ю., Корень О.Г., Журавлев Ю.Н. Строение гинецея и андроцея Aristolochia manshuriensis (Aristolochiaceae) // Растит. ресурсы. 2006. Т. 42. Вып. 3. С. 37–42.
Наконечная О.В., Нестерова С.В., Воронкова Н.М., Холина А.Б. Онтоморфогенез Aristolochia fimbriata Cham. (Aristolochiaceae) в условиях защищенного грунта // Вестн. ДВО. 2015. № 1. С. 50–57.
Наконечная О.В., Журавлев Ю.Н., Булгаков В.П., Корень О.Г., Сундукова Е.В. Род Кирказон на Дальнем Востоке России (Aristolochia manshuriensis Kom. и A. contorta Bunge). Владивосток: Дальнаука, 2014. 153 с.
Нестерова С.В., Наконечная О.В. Цветение и плодоношение представителей рода Aristolochia L. в условиях культуры // Биологическое разнообразие. Интродукция растений: Матер. Шестой Междунар. науч. конф. СПб.: СИНЭЛ, 2016. С. 356–359.
Паушева З.П. Практикум по цитологии растений. М.: Колос, 1970. 254 с.
Хохлов С.С. Апомиксис и цитоэмбриология растений. Саратов: Наука, 1968. 152 с.
Хохлов С.С. Выявление апомиктичных форм во флоре цветковых растений СССР. Саратов: Наука, 1978. 223 с.
Adams C.A., Baskin J.M., Baskin C.C. Comparative morphology of seeds of four closely related species of Aristolochia subgenus Siphisia (Aristolochiaceae, Piperales) // Bot. J. Linn. Soc. 2005a. V. 148. № 4. P. 433–436.
Adams C.A., Baskin J.M., Baskin C.C. Trait stasis versus adaptation in disjunct relict species: evolutionary changes in seed dormancy-breaking and germination requirements in a subclade of Aristolochia subgenus Siphisia (Piperales) // Seed Sci. Res. 2005b. V. 15. № 2. P. 161–173.
Barringer K., Whitteremore A.T. Aristolochiaceae Jusseu // Flora of North America, North of Mexico. N.Y.: Oxford Univ. Press, 1997. V. 3. P. 44–53.
Berjano R., Ortiz P.L., Arista M., Talavera S. Pollinators, flowering phenology and floral longevity in two Mediterranean Aristolochia species, with a review of flower visitor records for the genus // Plant Biol. 2009. V. 11. № 1. P. 6–16.
Burk C.J., Zebryk T.M. Woody Aristolochia species in western Massachusetts // Rhodora. 2001. V. 103. № 916. P. 427–430.
Endress P.K. Floral structure and evolution of primitive angiosperms: recent advances // Plant Syst. Evol. 1994. V. 192. P. 79–97.
González F., Pabón-Mora N. Trickery flowers: the extraordinary chemical mimicry of Aristolochia to accomplish deception to its pollinators // New Phytol. 2015. V. 206. P. 10–13.
González F., Rudall P.J. Structure and development of the ovule and seed in Aristolochiaceae, with particular reference to Saruma // Plant Syst. Evol. 2003. V. 241. № 3–4. P. 223–244.
González F., Stevenson D.W. A phylogenetic analysis of the subfamily Aristolochioideae (Aristolochiaceae) // Revista de la Academia Colombiana de Ciencias Exactas. Físicas y Naturales. 2002. V. 26. P. 25–60.
González F., Rudall P.J., Furness C.A. Microsporogenesis and systematics of Aristolochiaceae // Bot. J. Linn. Soc. 2001. V. 137. № 3. P. 221–242.
González F., Wagner S.T., Salomo K., Symmank L., Samain M.-S., Isnard S., Rowe N.P., Neinhuis Ch., Wanke S. Present trans–Pacific disjunct distribution of Aristolochia subgenus Isotrema (Aristolochiaceae) was shaped by dispersal, vicariance and extinction // J. Biogeogr. 2014. V. 41. P. 380–391.
Graham E.E. Host plant relationships and chemical communication in the Cerambycidae: Doct. diss. Urbana: Univ. Illinois Urbana-Champaign, 2010. 116 p.
Igersheim A., Endress P.K. Gynoecium diversity and systematics of the paleoherbs // Bot. J. Linn. Soc. 1998. V. 127. № 4. P. 289–370.
Joyner J.M., Chester E.W. The vascular flora of cross creeks national wildlife refuge, Stewart County, Tennessee // Castanea. 1994. V. 59. № 2. P. 117–145.
Lu L., Wortley A.H., Li D., Wang H., Blackmore S. Evolution of angiosperm pollen. 2. The basal angiosperms // Ann. Mo. Bot. Gard. 2015. V. 100. № 3. P. 227–269.
Nair N.C., Narayanan K.R. Studies on the Aristolochiaceae. I. Nodal and floral anatomy // Proc. Natl Inst. Sci. India. 1962. V. 28. P. 211–227.
Nakonechnaya O.V., Kalachev A.V. Pollen ultrastructure in Aristolochia manshuriensis and A. contorta (Aristolochiaceae) // Protoplasma. 2018. V. 255. № 5. P. 1309–1316.
Oh S.-Y., Pak J.-H. Distribution maps of vascular plants in Korea. Seul: Academybook Publ. Co, 2001. 997 p.
Ohi-Toma T., Sugawara T., Murata H., Wanke S., Neinhuis C., Murata J. Molecular phylogeny of Aristolochia sensu lato (Aristolochiaceae) based on sequences of rbcL, matK, and phyA genes, with special reference to differentiation of chromosome numbers // Syst. Bot. 2006. V. 31. № 3. P. 481–492.
Pfeifer H.W. Revision of the North and Central American hexandrous species of Aristolochia (Aristolochiaceae) // Ann. Mo. Bot. Gard. 1966. V. 53. № 2. P. 115–196.
Razzak M.A., Ali T., Ali S.I. The pollination biology of Arislohchia bracteolata Lamk (Aristolochiaceae) // Pak. J. Bot. 1992. V. 24. № 1. P. 79–87.
Robertson C. Flowers and Insects. Carlinville, Illinois, 1928. 221 p.
Robinsohn I. Die Farbungsreaction der Narben, Stigmatochromie, als morphologische Blutenuntersuchungenmethode // Sitsungsber. Akad. Wiss. 1924. Bd 133. H. 6. S. 181–211.
Tosunoglu A., Malyer H. Aristolochia adalica (Aristolochiaceae), a New Species from Anatolia, Turkey // Ann. Bot. Fenn. 2014. V. 51. № 3. P. 161–166.
Walker J.W., Doyle J.A. The bases of angiosperm phylogeny: palynology // Ann. Mo. Bot. Gard. 1975. V. 62. P. 664–723.
Ward D.B., Austin D.F., Coile N.C. Endangered and threatened plants of Florida, ranked in order of rarity // Castanea. 2003. V. 68. № 2. P. 160–17.
Zhou J., Xie G., Yan X. Encyclopedia of traditional chinese medicines: molecular structures, pharmacological activities, natural sources and applications. Berlin; Heidelberg: Springer-Verlag, 2011. V. I. 557 p.; V. II. 525 p.; V. III. 669 p.; V. IV. 636 p.; V. V. 601 p.; V. VI. 730 p.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая