

Известия РАН. Серия биологическая, 2021, № 1, стр. 44-54
Модульная структура соцветий двух видов рода Hydrangea L.
И. М. Кокшеева 1, *, Д. В. Нехайченко 1
1 Ботанический сад-институт ДВО РАН
690024 Владивосток, ул. Маковского, 142, Россия
* E-mail: koksheeva@yandex.ru
Поступила в редакцию 17.05.2019
После доработки 25.10.2019
Принята к публикации 06.12.2019
Аннотация
Исследована структура сложных соцветий Hydrangea petiolaris Siebold et Zucc. и Hydrangea paniculata Sieb. с использованием физиономического, структурного и фрактального подходов. Установлено, что физиономически различные соцветия имеют общую структуру – плейотирс и равное число цветков. Отмечено, что строение соцветий в целом упорядоченное и имеет модульную организацию. Обнаружено, что изменения в структуре соцветий закономерны и происходят параллельно во всех модулях за счет редукции или недоразвития цветков в каждом из модулей.
Представители рода Hydrangea L. (Hydrangeaceae Dumort.) популярны в культуре во всем мире, благодаря чему довольно хорошо изучены в различных аспектах (Hufford et al., 2001; Hufford, 2004; Jacobs, 2010; Samain et al., 2010; Granados Mendoza et al., 2013; De Smet et al., 2015). Особый интерес для исследования представляют дальневосточные виды H. petiolaris Siebold et Zucc. и H. paniculata Sieb., произрастающие на территории России. H. petiolaris высоко ценится как декоративно-цветущая деревянистая лиана. H. paniculata – один из перспективных морозостойких видов, на основе которого выведено большое число популярных сортов, которые отличаются размерами куста, формами, размерами и окраской соцветий. Известно, что форма и размер соцветия зависят от его структуры, причем структура соцветий может изменяться за счет ограничений в развитии. Такие изменения, как правило, зависят от климатических условий в период формирования (Wisniewska, Zawadzka, 1962; Galopin et al., 2008). В связи с этим исследование структуры соцветий актуально и может быть использовано для моделирования различных адаптивных стратегий.
С точки зрения структуры соцветия играют важную роль для решения задач систематики и филогении (Claβen-Bockhoff, Bull-Hereñu, 2013; Kirchoff, Claβen-Bockhoff, 2013). Однако соцветия Hydrangea имеют противоречивые морфологические трактовки из-за использования различных подходов. Чаще всего в таксономических описаниях используют физиономический подход, в рамках которого соцветия Hydrangea характеризуются как щитковидные, зонтиковидные, округлой или пирамидальной формы (McClintock, 1957; Тахтаджян, 1966; Bertrand, 1992; Jintang et al., 2001). Однако физиономический подход отдельно используется крайне редко, так как содержит в себе элементы структурного подхода. В рамках структурного подхода соцветия Hydrangea характеризуются как метельчатые (Troll, 1964, 1969), цимы (Engler, 1891), дендрически разветвленный кластер (McClintock, 1957), тирс (Kuhlen, 1956), цимозные (Тахтаджян, 1966), плейохазий (Wisniewska, Zawadzka, 1962), цим или тирс (Jintang et al., 2001), сложное соцветие дихазиального типа (Galopin et al., 2008), цимозная метелка (Rabeler, 2016). Таким образом, соцветия Hydrangea типифицированы с точки зрения внешней формы и с точки зрения структуры осевой системы. Но этого недостаточно для понимания закономерностей изменений, происходящих как в общей структуре соцветия, так и в отдельных ее элементах.
Морфология растений – это область ботаники, которая напрямую нуждается в применении математики, точнее фрактальной геометрии, где рассматриваются пространственные структуры. Многие растения демонстрируют самоподобие, т.е. сходство части и целого, что указывает на их принадлежность к фракталам. В связи с этим для морфологии растений фрактальный подход представляет особый интерес. В рамках этого подхода сложное соцветие Hydrangea можно рассматривать как фрактал, структура которого повторяется не в точности и не в полной мере (квазисамоподобие) (Мандельброт, 2002). Строение сложных многоцветковых соцветий в целом упорядоченное и закономерное. Как правило, для них характерна модульная организация. Под модулем подразумевается однотипная структура, формирующаяся в результате одного цикла формообразования и закономерно повторяющаяся во времени и пространстве (Савиных, Мальцева, 2008). МакКлинток (McClintock, 1957) описывает соцветие Hydrangea как дендрически разветвленный, терминальный кластер, конечные элементы которого состоят из небольших кластеров. Кластер в данном определении сходен по значению с модулем, так как он обозначает объединение нескольких однородных элементов, которое может рассматриваться как самостоятельная единица. Однако отсутствие подробного описания структуры кластеров (модулей) и их распределения по осям соцветия Hydrangea (McClintock, 1957) не раскрывает закономерности изменений структуры соцветия даже в рамках фрактального подхода.
Цель работы – выявление закономерностей изменения структуры соцветия Hydrangea на основании анализа детальных схем, отражающих расположение цветков.
МАТЕРИАЛЫ И МЕТОДЫ
Для исследования использованы сформированные соцветия H. petiolaris Siebold et Zucc. (гортензия черешчатая) и H. paniculata Sieb. (гортензия метельчатая). Материал собран в 2015 г. с растений, произрастающих в условиях открытого грунта в экспериментальной зоне Ботанического сада-института ДВО РАН (43°13′27.48′′ с.ш., 131°59′36.32′′ в.д.). С каждого вида отбирали по 30 соцветий в разных частях куста/лианы. У соцветия измеряли диаметр и высоту, подсчитывали число ярусов, число порядков ветвления, общее число цветков и число краевых цветков. Для соцветия составлялась полная схема ветвления с учетом положения каждого отдельного цветка.
Данные обрабатывались вариационно-статистическими методами с использованием пакетов Microsoft Excel и StatSoft Statistica 6.0. В тексте и табл. 1 приведены среднее арифметическое ± ± стандартное отклонение. Проверку на нормальность распределения выборок значений морфометрических параметров проводили с помощью W-теста Шапиро–Уилка. В результате проверки у выборок отклонения от нармального распределения не были выявлены и для их сравнения использовали t-критерий Стьюдента.
Таблица 1.
Морфометрическая характеристика соцветий
| Критерий | Hydrangea petiolaris | Hydrangea paniculata | p |
|---|---|---|---|
| Высота, см | 13.3 ± 3.85 | 4.0 ± 0.62 | 7.1** |
| Диаметр, см | 17.3 ± 2.62 | 11.4 ± 2.0 | 5.1** |
| Число ярусов, шт. | 7 ± 0.31 | 11.3 ± 0.96 | 6.4** |
| Число порядков ветвления, шт. | 8 ± 1.06 | 6.4 ± 0.6 | 10.9** |
| Общее число цветков, шт. | 799 ± 188.6 | 862 ± 253.6 | 0.2 |
| Число краевых цветков, шт. | 7 ± 1.74 | 15 ± 8.6 | 2.6* |
Фотографии были сделаны с помощью фотокамеры Sony Cyber-shot HX400 и стереомикроскопа Stemi 2000-C (Carl Zeiss, Германия) с цветной цифровой фотокамерой Axiocam ICc 3. При обработке изображений, полученных на стереомикроскопе, использовали программу AxioVision 13.
Расшифровку структуры соцветия проводили с учетом филлотаксиса генеративных и репродуктивных побегов, углов дивергенции и длины междоузлий (Prusinkiewicz et al., 2007; Prusinkiewicz, Reuille, 2010). Все боковые ответвления на главной оси соцветия рассматривали как боковые оси первого порядка ветвления. На осях первого порядка выделяли оси второго, третьего и последующих порядков ветвления. Число ярусов на главной оси соцветия и порядков ветвления боковых осей учитывали по числу супротивно или поочередно расположенных осей. Под ярусом подразумевали пару супротивно или поочередно расположенных боковых осей, имеющих угол дивергенции 180°. Порядок ярусов устанавливали в акропетальном направлении, т.е. нижний ярус рассматривали как первый. Подобную структуру ярусов наблюдали и на боковых осях соцветия. Поэтому при анализе архитектуры боковых ответвлений порядок ярусов определяли также в акропетальном порядке, т.е. от основания оси к ее верхушке. Так как соцветие трехмерное и его оси расположены накрест супротивно, для удобства восприятия при составлении общей схемы длину междоузлий не учитывали, а положение осей отображали супротивно в одной плоскости. Чтобы не перегружать схему лишними элементами, прилистники на ней не отображены. При описании соцветий использованы физиономический и структурный подходы (Troll, 1964, 1969; Кузнецова и др., 1992; Endress, 2010).
При выделении в общей схеме повторяющихся элементов (модулей) использовали фрактальный подход (Мандельброт, 2002; Савиных, Мальцева, 2008). Под модулем подразумевали однотипную структуру в соцветии, закономерно повторяющуюся во времени и в пространстве (Савиных, Мальцева, 2008). Модули в соцветии выделяли по следующим признакам: боковые модули повторяют строение терминального модуля; в основании модуля листья уменьшаются в размере и преобразуются в прицветники (брактеи); ветвление модуля начинается из пазух прицветников и число разветвлений фиксировано; изменение филлотаксиса с супротивного у основных модулей на очередной происходит в более мелких модулях; более длинное междоузлие отмечено перед терминальным модулем и основными боковыми модулями, уменьшение длины междоузлий – при переходе в более мелкие модули; обнаружeны также редукция или недоразвитие латеральных элементов модуля (Кузнецова, 1991).
Для выявления последовательности изменений отдельных модулей в структуре соцветия для мелких модулей зарисовывали схемы расположения цветков и для наглядности использовали формулы.
Термины и формулы, используемые для описания структуры соцветий (Hufford, 2001; Endress, 2010):
терминальный цветок (tf) – цветок на верхушке главной оси соцветия,
tf1..n – цветок соподчиненного порядка ветвления;
цимоиды (с) – соцветия, главная ось которых заканчивается терминальным цветком и имеет не более двух боковых осей второго порядка ветвления;
дихазий (d) – соцветие, состоящее из терминального цветка и двух цветков соподчиненного порядка ветвления d = [tf + 2tf1];
монохазий (m) – соцветие, состоящее из терминального цветка и одного цветка соподчиненного порядка ветвления m = [tf + tf1];
сложный дихазий (cd) – соцветие, главная ось которого заканчивается терминальным цветком и имеет две боковые оси соподчиненного порядка ветвления, структура которых повторяет структуру главной оси, один или несколько раз cd = [tf + + 2(tf1 + 2tf2) …];
сложный монохазий (cm) – завиток и извилина – соцветие, главная ось которого заканчивается терминальным цветком и имеет одну боковую ось соподчиненного порядка ветвления, повторяющую структуру главной оси, один или несколько раз cm = [tf + tf1 + tf2 +…+ tfn];
тирс (T) – сложное соцветие, главная ось которого заканчивается терминальным цветком и имеет более двух боковых осей второго порядка ветвления, представленных цимоидами, при этом оси второго порядка ветвления могут ветвиться несколько раз сходным образом;
плейотирс (PT) – множественный тирс – сильно разветвленное соцветие, структурной единицей которого является тирс;
краевые цветки (mf) – цветки с крупным венчиковидным околоцветником, которые образуются на конечных ответвлениях боковых осей соцветия.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Характеристика вегетативных и репродуктивных побегов. Вегетативные и репродуктивные побеги H. paniculata и H. petiolaris имеют накрест супротивное листорасположение. Обычно в одном узле формируются два листа. Однако на репродуктивных побегах H. paniculata могут формироваться три листа. Угол дивергенции между листьями одного узла обычно равен 180°. Сходным образом располагаются прицветники и оси соцветий. Размеры листовых пластинок и черешков на вегетативных и репродуктивных побегах варьируют. На репродуктивных побегах размеры листьев сокращаются по направлению снизу вверх. При переходе побега из вегетативной части в репродуктивную размеры прицветников в составе соцветий также уменьшаются (до 1 мм) по направлению снизу вверх.
Характеристика структуры соцветий. Соцветия Hydrangea крупные, сложные и представляют собой систему соподчиненных ответвлений. Ветвление главной оси соцветий боковое, моноподиальное с очередным и супротивным расположением боковых осей, у конечных ответвлений – симподиальное. Каждая ось соцветия заканчивается терминальным цветком. По наличию и характеру прицветных листьев соцветия фрондулозно-брактеозные, т.е. в основании соцветия и основных модулей имеют зеленые верхушечные листья меньших размеров (рис. 1а) и видоизмененные листья – брактеи (прицветники) в более мелких модулях (рис. 1б). Средние числа цветков в соцветиях у растений, выращенных в одинаковых условиях среды, достоверно не различаются: у H. petiolaris в среднем 799 цветков, из которых 0.9% краевыe; у H. paniculata в среднем 862 цветкa, из которых 1.7% краевыe (табл. 1). Различия между видами выявлены по соотношению высоты и диаметра соцветия (Н. petiolaris – 1 : 2.85; H. paniculata – 1 : 1.3) и распределению цветков по осям соцветия. Боковые оси соцветий располагаются попарно. На главной оси соцветия H. petiolaris формируется до 7 пар боковых осей и до 8 порядков ветвлений на нижней паре боковых осей (рис. 2а (4a, 4б)). На главной оси соцветия H. paniculata формируется в среднем 12 пар боковых осей и до 6 порядков ветвления на нижней паре боковых осей (рис. 3б (4a, 4б)). Боковые оси H. petiolaris располагаются накрест супротивно, длина междоузлий главной оси соцветия и длина боковых осей, а также число ответвлений на боковых осях сокращаются по направлению снизу вверх (рис. 2а (1б, 4a, 4б)). Удлинением боковых осей нижних ярусов Н. petiolaris обуславливается расположение всех цветков в одной плоскости, в результате чего соцветие приобретает щитковидную форму (рис. 2а (1a, 1б)). Краевые цветки формируются на боковых осях двух нижних ярусов (рис. 2а (4a)). Оси соцветия нижних ярусов Н. paniculata располагаются накрест супротивно, с середины соцветия по направлению вверх оси переходят в поочередное расположение, при этом число цветков на них имеет парную симметрию и угол дивергенции между осями сохраняется (рис. 2б (1б)). Начиная со второго порядка ветвления в верхних ярусах и с третьего в нижних ярусах, междоузлия редуцируются, в результате чего боковые оси H. paniculata сближены (рис. 2б (4a)). Краевые цветки формируются на боковых осях пяти нижних ярусов (рис. 2б (4a)). Таким образом, на основе совокупности признаков выделены два физиономических типа соцветия: щитковидное (H. petiolaris) (рис. 2а (1a, 1б)) и метельчатое (Н. paniculata) (рис. 2б (1a, 1б)). При расшифровке структуры соцветий для каждого вида составлены подробные схемы, отражающие расположения всех цветков (рис. 2а (4б), 2б (4б)).
Рис. 1.
Типы листьев в соцветии Hydrangea. a и б – стрелки указывают на зеленые верхушечные листья меньших размеров и брактеи соответственно.

Рис. 2.
Соцветия Hydrangea petiolaris (а) и H. paniculata (б). 1a – вид сверху, 1б – вид сбоку, 2a – общий вид цимоида, 2б – схема цимоида, 2в – схематическая диаграмма цимоида, 2г – условное обозначение цимоида, 3a – общий вид тирса, 3б – схема тирса, 3в – схематическая диаграмма тирса, 3г – условное обозначение тирса, 4a – пары боковых осей соцветия; 4б – общая схема строения соцветия.
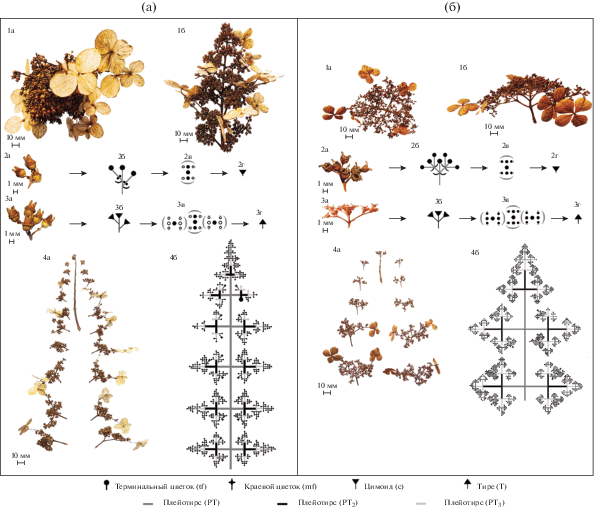
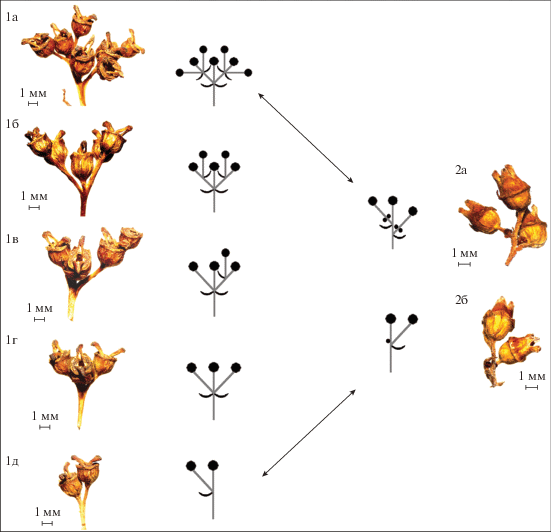
Рис. 3.
Цимоиды Hydrangea. 1a–1д – Hydrangea petiolaris, 1a–1в – сложный дихазий (cd), 1г – дихазии (d), 1д – монохазий (m), 2a, 2б – Hydrangea paniculata, 2a – сложный дихазий (cd), 2б – монохазий (m).
Модульная структура соцветия. При анализе подробных схем в структуре соцветия выделены следующие модули: основной модуль (I) – плейотирс (PT), промежуточный (II) – тирс (T), исходный (III) – цимоид (c) (рис. 2а, 2б). Плейотирс в структуре соцветия Hydrangea повторяется несколько раз (PT, PT1, PT2) (рис. 2а (4б), 2б (4б)). Вся структура соцветия характеризуется как плейотирс (PT), который образован плейотирсами PT1. Различия между видами наблюдаются по количеству и распределению плейотирсов (PT1) в общей структуре соцветия. Модуль PT H. petiolaris состоит из пяти модулей PT1: PT = [PT1 + 2PT1 + + 2PT1] = 5PT1 (рис. 2а (4б)). Один из модулей PT1 расположен терминально, а остальные четыре – накрест супротивно по отношению к главной оси. Модуль PT H. paniculata состоит в среднем из 13 модулей PT1: PT = 15PT1 или PT = 13PT1 или PT = 11PT1, при этом один из модулей PT1 расположен терминально, остальные – парно накрест супротивно или накрест поочередно (рис. 2б (4б)). Накрест поочередное расположение осей связано с их смещением, так как изначально они закладываются супротивно, а угол дивергенции при смещении сохраняется прежним (180°).
Модуль PT1 образован несколькими модулями PT2, и его структура может быть развита в большей или меньшей степени. При полном развитии модуль PT1H. petiolaris состоит из девяти модулей PT2 и имеет следующее строение: PT1 = [3(PT2 + + PT2 + PT2)] = 9PT2. Однако чаще всего часть модулей PT2 может быть недоразвита и представлена тирсами. Также число модулей PT2 может уменьшаться до семи или даже до трех. В случае если модуль PT1 образован семью модулями, его структура представлена как модулями PT2, так и тирсами (Т). Если модуль PT1 образован тремя модулями, то эти модули представлены только модулями PT2: PT1 = [PT2 + PT2 + PT2] = 3PT2.
Модуль PT1H. paniculata, как правило, имеет большее число модулей PT2. Если модуль PT1 состоит из 5, 7, 9 или 11 модулей, его структура представлена как модулями PT2, так и тирсами (Т). Если модуль PT1 состоит из 13 или 15 модулей, то часть модулей PT2 образована не только тирсами (T), но и цимоидами.
Модуль PT2 у обоих видов образован тирсами (T), один из которых расположен терминально, а остальные накрест супротивно или накрест поочередно: PT2 = [T + T + T] = 3T. В случае недоразвития некоторых тирсов модуль PT2 образован тирсами и цимоидами: PT2 = [T + 2c] или PT2 = [T + T + c] (их расположение при этом сохраняется).
Тирсы (T) Hydrangea состоят из цимоидов (с), элементы которых могут быть развиты в разной степени в зависимости от яруса и уровня ветвления осей соцветия. Боковые оси тирса в основании располагаются супротивно, а на верхушке – поочередно. Длина боковых осей, число порядков ветвления в его структуре, увеличивается по направлению сверху вниз. Обычно тирс образован тремя цимоидами: T = [c1 + 2c2] (рис. 2а (3a–3г)). При максимальном недоразвитии, у обоих видов тирс может состоять из пяти цветков (T = [tf + 2tf1 + + 2tf1]), а у H. paniculata – из четырех (T = [tf + tf1 + + 2tf1]) (рис. 2б (3a–3г)).
Соцветия исходного уровня у Hydrangea представлены цимоидами различной структуры: сложный дихазий (cd) (рис. 3 (1a–1в, 2a)), сложный монохазий (cm), дихазии (d) (рис. 3, (1г)) и монохазий (m) (рис. 3 (1д, 2б)). Боковые оси цимоидов располагаются супротивно (H. petiolaris) (рис. 2б (2a–2г), 3 (1a–1г)) или поочередно (H. paniculata) (рис. 3 (2a)). Сложный дихазий у H. petiolaris обычно имеет 7 цветков: cd = [tf + 2tf1 + 4tf2] или cd = [tf + 2(tf1 + 2tf2)] = [tf + 2d] (рис. 3 (1a)). Его элементы могут быть развиты в разной степени. В результате редукции сложный дихазий может быть представлен дихазием d = [tf + 2tf1], сложным монохазием cm = [tf + tf1 + tf2 + tf3], простым монохазием m = [tf + tf1] или терминальным цветком [tf]. Цимоиды H. paniculata также имеют 7 цветков, 3 из которых развиты, а 4 недоразвиты и расположены по 2 в основании боковых осей (рис. 2а (2a–2г), 3 (2a), 4 (1a–1в)). В связи с этим такие цимоиды могут быть охарактеризованы как недоразвитые сложные дихазии или как простые дихазии. В случае дальнейшего недоразвития элементов дихазия он может быть представлен монохазием или терминальным цветком.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Для понимания изменений, происходящих в структуре сложных соцветий, необходимо изначально правильно классифицировать саму структуру. Основу любой классификации составляет разделение соцветий на два противопоставляемых типа ветвления: рацемозное и цимозное. Известно, что в рацемозной структуре главная ось имеет неограниченное число боковых осей, но не имеет осей более высокого порядка. В цимозной структуре на главной оси образуются только две боковые оси, которые в дальнейшем могут ветвиться несколько раз сходным образом. Ветвление в соцветиях обычно начинается рацемозно, а оси более высокого порядка ветвятся цимозно. Такая модель ветвления характерна для тирса. Если основная модель ветвления в соцветии повторяется несколько раз, то цимозные соцветия называют плейотирсами, а рацемозные – составной кистью (Endress, 2010). Анализ соцветий H. petiolaris и H. paniculata показал, что ветвление в соцветии сходно с ветвлением тирса и повторяется несколько раз. В результате у обоих видов формируется соцветие плейотирс или множественный тирс, конечные ответвления которого представлены цимоидами. Для увеличения числа цветков в рацемозной модели главная ось удлиняется и образует больше боковых цветков, а в цимозной увеличивается число порядков ветвления. На главной оси соцветия H. petiolaris формируется до 7 пар боковых осей и до 8 порядков ветвления на нижней паре боковых осей (рис. 2а). На главной оси соцветия H. paniculata формируется в среднем 12 пар боковых осей и до 6 порядков ветвления на нижней паре боковых осей (рис. 2б). Увеличение или уменьшение структуры соцветия обоих видов закономерно и происходит за счет увеличения или сокращения числа боковых осей и порядков ветвления.
Тролль (Troll, 1964, 1699) рассматривает соцветия Hydrangeaceae как метелки, при этом отмечая нетипичное для этого семейства соцветие – малоцветковый щитковидный тирс (т.е. цимозное) у Deinanthe. Метелка при этом, как и тирс, описывается как промежуточная модель между цимозной и рацемозной структурой, так как на главной оси формируется более двух боковых осей первого порядка ветвления (не цимозная), каждая из которых имеет боковые оси более высокого порядка (не рацемозная). В метелке нет ограничений ни в числе порядков ветвления, ни в числе цветков в пределах одного порядка ветвления. Кроме того, конечные ответвления метелки представлены кистями (т.е. рацемозные), а по нашим данным, конечные ответвления гортензии представлены цимоидами, т.е. соцветия Hydrangeaceae не могут характеризоваться как метелка.
В соцветии Hydrangea отличить дихазий от кисти можно только с помощью анализа взаиморасположения всех элементов соцветия в трехмерном пространстве и при изучении изменений, происходящих в онтогенезе. Это связано с тем, что не всегда оси дихазия имеют супротивное расположение, а могут располагаться и поочередно (Тахтаджян, 2007). В соцветиях Hydrangea в направлении от основания к верхушке наблюдается смещение боковых осей. В результате дихазий с супротивным расположением осей преобразуется в дихазий с очередным расположением осей.
В сформированном соцветии H. paniculata часть осей, которые на ранней стадии закладывались изначально супротивно, по мере удлинения междоузлий расходятся и располагаются в верхней части соцветия поочередно по отношению одна к другой (рис. 5a, б). Тирсы H. paniculata из-за сильной цветочной редукции состоят из 4–5 цветков и по внешнему облику становятся похожими на кисти (рис. 2б (4б), 3 (2a, 2б)). В результате структура плейотирса Н. paniculata внешне становится похожей на метелку, что часто вносит путаницу в классификацию соцветия. На сходство соцветия Н. paniculata с метелкой также указывает порядок распускания цветков. Цветение Н. paniculata начинается в нижней паре боковых осей с распускания терминальных цветков. Затем начинает зацветать вторая снизу пара боковых осей, далее третья и т.д. (рис. 5в, г), т.е. последовательность цветения отмечается от основания к верхушке, что также было описано ранее (McClintock, 1957).
Рис. 4.
Пример недоразвития структуры соцветия Hydrangea. Стрелки указывают на недоразвитые цветки, расположенные по два в основании развитого цветка.

Рис. 5.
Соцветие Hydrangea paniculata. а и б – стрелки указывают на расположение боковых осей: супротивно (а), поочередно (б); в и г – порядок распускания цветков в соцветии.

Сложности при разграничении тирса и метелки возникают из-за того, что эти термины могут иметь несколько значений. Изначально термины “тирс” и “метелка” трактовались чисто габитуально, но в дальнейшем их характеристика претерпела изменения и стала основываться на признаках ветвления осей. При этом старое физиономическое значение терминов не исчезло. В результате такие термины в настоящее время имеют двойной смысл: с одной стороны, они отражают определенный внешний облик собрания цветков, а с другой – структуру осевой системы (Кузнецова, 1991; Кузнецова, Тимонин, 2017). Поэтому следует различать общий вид соцветия и тип разветвления составных частей (Buys, Hilger, 2003). Поскольку выбранный морфологами язык описания напрямую влияет на качество описания и его результаты, то необходимо использовать однозначно понимаемые, непротиворечивые термины, универсальные для специалистов как в области ботаники, так и в области смежных областей знаний. Более ценны в таких исследованиях иллюстрации, так как схемы лучше отображают сравнительные данные, чем описания, и облегчают сравнение отдельных частей соцветия (Kellogg, 2015).
Для морфологического анализа соцветий целесообразно использовать концепцию модульной организации растений, основанную на выделении однотипных закономерно повторяющихся единиц (т.е. модулей) (Нотов, 2001). Проблему повторяющихся единиц в соцветиях попытались решить на примере семейства Eriocaulaceae Martinov (Stützel, Trovó, 2013). Вместо классификации всех видов сильно разветвленных цветочных систем авторы описали основную репродуктивную единицу, общую для всех цветочных систем. Далее они описали уровни повторения в каждой системе. В структуре соцветий гортензии также были выделены повторяющиеся единицы – модули. Закономерное повторение модели ветвления на разных уровнях сложности привело к образованию сложного соцветия, состоящего из трех видов модулей: основного модуля (I) – плейотирса, промежуточного (II) – тирса, исходного (III) – цимоида. Несмотря на внешние физиономические различия между метелковидным (Н. paniculata) и щитковидным (H. petiolaris) плейотирсами, виды имеют сходную структуру и одинаковый набор модулей. Это согласуется с идеей Эймера (Eimer, 1897) о том, что у организмов одного таксона имеется общая последовательность онтогенетических стадий, но развитие может быть ограничено на разных стадиях. Веберлинг (Weberling, 1989), напротив, считал, что у видов близкородственных таксонов обнаруживаются совершенно различные соцветия, что не позволяет понять, какие же черты строения соцветия характерны для всего таксона. Результаты исследования показали, что физиономически различные соцветия H. petiolaris (щитковидное) и H. paniculata (метелковидное) имеют схожую структуру плейотирс.
Поскольку соцветия обладают модификационной пластичностью, то в ответ на различия в условиях произрастания, наследственно закрепленная, характерная для данного вида структура соцветия может внешне изменяться. Такие изменения практически всегда связаны с боковыми осями и носят не качественный, а количественный характер. Они часто связаны с числом цветков и относительной длиной осей соцветий, тогда как основная модель ветвления в основном не затрагивается (Endress, 2010). В зависимости от условий произрастания структура соцветий Hydrangea может быть развита в большей или меньшей степени за счет сокращения или увеличения числа ярусов и порядков ветвления боковых осей. Преобразование соцветия закономерно и происходит параллельно на всех уровнях ветвления. Структура плейотирса может быть недоразвита и представлена тирсами PT ⟶ T. Структура тирса зависит от структуры цимоида и при недоразвитии представлена цимоидом T ⟶ с. При недоразвитии цветков в сложном дихазии его структура может быть представлена дихазием, сложным монохазием, монохазием или терминальным цветком cd → d/cm → m → tf (рис. 3). Соцветия могут видоизменяться вследствие недоразвития или чрезмерного развития, а изменения структуры закономерны и могут быть симметричными или асимметричными.
Типы модулей и последовательность их формирования в структуре растения заложены генетически и определяют морфологический критерий вида (Савиных, 2013). Число модулей (число первичных, вторичных и третичных ответвлений) также контролируется генетически и у разных видов варьирует. При этом число первичных ответвлений на главной оси варьирует независимо от числа вторичных ответвлений, а отдельные виды часто имеют свой характерный диапазон значений (Kellogg, 2015). Структура соцветий обоих видов Hydrangea (Н. paniculata и Н. petiolaris) формируется сходным образом по типу дихазия. На начальных этапах онтогенеза в соцветии Н. petiolaris закладываются пять основных модулей, формирующих первый ярус, и последующее развитие общей структуры соцветия уже происходит параллельно в каждом из них по типу дихазия. Наличие пяти модулей в структуре соцветия также характерно и для других видов Hydrangea (например, H. macrophylla (Thunb.) Ser., (Wisniewska, Zawadzka, 1962; Galopin et al., 2008)). В соцветии Н. paniculata по сравнению с Н. petiolaris образуется больше ярусов на главной оси соцветия, но меньше уровней ветвления на боковых осях, за счет чего число основных модулей (в среднем 13) увеличивается, а число исходных модулей уменьшается. Удлинение осей отличается от ветвления соцветия, так как оно происходит после окончания разветвления и контролируется разными генами (Kellogg, 2015). Удлинение осей каждого порядка контролируется независимо от других. Из-за этого соцветия с одинаковым числом порядков ветвления и одинаковым числом ветвей в каждом порядке могут выглядеть совершенно по-разному в зависимости от того, какие оси удлиняются (Kellogg, 2015), что характерно и для изученных видов гортензии. Все оси в соцветии Н. petiolaris удлиняются таким образом, чтобы цветки расположились в одной плоскости, в результате чего соцветие приобретает щитковидную форму – формируется щитковидный плейотирс. В соцветии H. paniculata длина осей первого порядка уменьшается от основания к верхушке соцветия, а междоузлия на осях следующих порядков ветвления сближены до такой степени, что создается впечатление, что оси выходят из одной точки. В результате соцветие H. paniculata приобретает метелковидную форму – формируется метелковидный плейотирс.
Таким образом совместное использование физиономического, структурного и фрактального подходов позволило точнее описать структуру сложных соцветий Hydrangea. На основе совокупности признаков для Hydrangea выделено два физиономических типа соцветия: щитковидное (H. petiolaris) и метельчатое (Н. paniculata). В рамках структурного подхода структура соцветий обоих видов характеризуется как плейотирс. С использованием фрактального подхода в соцветии выделены три типа модулей: основной (I) – плейотирс, промежуточный (II) – тирс, исходный (III) – цимоид. Различия между видами выявлены по числу и распределению модулей в структуре соцветия. Структура соцветия изменяется за счет редукции или недоразвития цветков в каждом из модулей, при этом изменения закономерны и происходят параллельно во всех модулях.
Авторы благодарят В.Е. Харченко (Луганский национальный аграрный университет) за идеи и ценные рекомендации при выполнении работы.
Список литературы
Кузнецова Т.В. Морфология соцветий: современное состояние // Современные методы систематики высших растений. Морфология и экология репродуктивных органов цветковых растений. Итоги науки и техники. Ботаника. М.: ВИНИТИ, 1991. Т. 12. С. 51–124.
Кузнецова Т.В., Тимонин А.К. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М.: КМК, 2017. 183 с.
Кузнецова Т.В., Пряхина Н.И., Яковлев Г.П. Соцветия. Морфологическая классификация. СПб.: Химико-фармацевтический ин-т, 1992. 127 с.
Мандельброт Б.Б. Фрактальная геометрия природы. (Перевод с англ.) М.: Ин-т компьютерных исследований, 2002. 656 с.
Нотов А.А. Концепция модульной организации и проблема организационного полиморфизма на разных уровнях структурной иерархии живых организмов // Гомологии в ботанике: опыт и рефлексия: Тр. IX школы по теоретической морфологии растений “Типы сходства и принципы гомологизации в морфологии растений”. СПб.: Санкт-Петербургский союз ученых, 2001. С. 119–128.
Савиных Н.П. Модульная организация цветковых растений и ее последствия // Modern Phytomorphol. 2013. № 3. С. 139–145.
Савиных Н.П., Мальцева Т.А. Модуль у растений как структура и категория // Вестн. ТВГУ. Сер. биол. и экол. 2008. № 9. С. 227–234.
Тахтаджян А.Л. Система и филогения цветковых растений. Л.: Наука, 1966. 611 с.
Тахтаджян А.Л. Грани эволюции. Статьи по теории эволюции (1943–2006 гг.). СПб.: Наука, 2007. 326 с.
Bertrand H. Identification of Hydrangea macrophylla Ser. cultivars // Acta Hortic. 1992. V. 320. P. 209–212.
Buys M.H., Hilger H.H. Boraginaceae cymes are exclusively scorpiod and not helicoids // Taxon. 2003. V. 52(4). P. 719–724.
Claßen-Bockhoff R., Bull-Hereñu K. Towards an ontogenetic understanding of inflorescence diversity // Ann. Bot. 2013. V. 112(8). P. 1523–1542.
De Smet Y., Granados Mendoza C., Wanke S., Goetghebeur P., Samain M.S. Molecular phylogenetics and new (infra)generic classification to alleviate polyphyly in tribe Hydrangeeae (Cornales: Hydrangeaceae) // Taxon. 2015. V. 64(4). P. 741–753.
Eimer T. On orthogenesis and the importance of natural selection in species-formation. Chicago: The Open Court Publ. Company, 1897. 68 p.
Endress P.K. Disentangling confusions in inflorescence morphology: Patterns and diversity of reproductive shoot ramification in angiosperms // J. Syst. Evol. 2010. V. 48(4). P. 225–239.
Engler A. Hydrangeoideae – Hydrangeea // Die Naturlichen Pflanzenfamilien / Eds Engler A., Prantl A. Leipzig: Verlag Wilhelm Engelmann, 1891. V. 3. 2A. 142 p.
Galopin G., Codarin S., Viemont J.D., Morel P. Architectural development of inflorescence in Hydrangea macrophylla cv. Hermann Dienemann // Hort Sci. 2008. V. 43(2). P. 361–365.
Granados Mendoza C., De Vos P., Goetghebeur P., Samain M. Tree top climbing: towards an evolutionary tree of Hydrangea section Cornidia. Ghent: Ghent Univ., 2013. 203 p.
Hufford L. The families and genera of vascular plants, flowering plants, Dicotyledons: Celastrales, Oxalidales, Rosales, Cornales, Ericales // Hydrangeaceae / Ed. Kubitzki K. Berlin: Springer-Verlag Publ., 2004. V. 6. 489 p.
Hufford L., Moody M.L., Soltis D.E. A phylogenetic analysis of Hydrangeaceae based on sequences of the plastid gene matK and their combination with rbcL and morphological data // Int. J. Plant Sci. 2001. V. 162(4). P. 835–846.
Jacobs S. Flag flower morphology and phylogeny of Hydrangeaceae tribe Hydrangeeae. Washington: Wash. State Univ., 2010. 207 p.
Jintang P., Cui zhi G., Shumei H., Chaofen W., Shuying J., Lingdi L., Ohba H., Gornall R.J., Soltis D., Cullen J., Hultgård U.-M., Akiyama S., Bartholomew B., Alexander C. Flora of China // Sasifragaceae / Eds Wu Z., Raven P.H. Beijing: Šci. Press & St. Louis: Missouri Botanical Garden, 2001. V. 8. P. 269–452.
Kellogg E.A. Flowering Plants. Monocots: Poaceae // The Families and Genera of Vascular Plants / Ed. Kubitzki K. Berlin: Springer, 2015. V. 13. 416 p.
Kirchoff B., Claßen-Bockhoff R. Inflorescences: concepts, function, development and evolution // Ann. Bot. 2013. V. 112(8). P. 1471–1476.
Kuhlen H. Die Hortensien. Zur morphologie und anatomie der gattung Hydrangea. Aachen: Verlag Deutsche Gartnerborse, 1956. S. 36–47.
McClintock E. A monograph of the genus Hydrangea // Proc. California Acad. Sci. 1957. V. 29. P. 147–256.
Prusinkiewicz P., Erasmus Y., Lane B., Harder L.D., Coen E. Evolution and development of inflorecence architecture // Science. 2007. V. 316. P. 1452–1456.
Prusinkiewicz P., Reuille P.B. Constraints of space in plant development // J. Exp. Bot. 2010. V. 61(8). P. 2117–2129.
Rabeler R.K. Flora of North America North of Mexico // Hydrangeaceae / Ed. Freeman C.C. N.Y.: Oxford Univ. Press, 2016. V. 12. 632 p.
Samain M.S., Wanke S., Goetghebeur P. Unraveling extensive paraphyly in the genus Hydrangea s.l. with implications for the systematics of tribe Hydrangeeae // Syst. Bot. 2010. V. 35(3). P. 593–600.
Stützel T., Trovó M. Inflorescences in Eriocaulaceae: taxonomic relevance and practical implications // Ann. Bot. 2013. V. 112(8). P. 1505–1522.
Troll W. Die Infloreszenzen. Jena: VEB Gustav Fischer Verlag, 1964. V. I. 615 p.
Troll W. Die Infloreszenzen. Typologie und Stellung im Aufbau desVegetationskörpers. Jena: VEB Gustav Fischer Verlag, 1969. V. II. 630 p.
Weberling F. Structure and evolutionary tendencies of inflorescences in the Leguminosae // Monographs in Systematic Botany from the Missouri Botanical Garden. 1989. V. 29. P. 35–58.
Wisniewska E., Zawadzka Z. Przebieg Ksztaltowania sie Kwiatostanu u Hydrangea macrophylla Ser. Cv. Altona // Acta Agrobot. 1962. V. 11. P. 157–165.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая