

Известия РАН. Серия биологическая, 2021, № 1, стр. 93-102
Распределение запасов фитомассы и биогенных элементов в древостоях лиственницы Гмелина в Центральной Эвенкии (на примере малого водосборного бассейна)
С. Г. Прокушкин 1, *, О. А. Зырянова 1, А. С. Прокушкин 1
1 Институт леса им. В.Н. Сукачева СО РАН – обособленное подразделение Федерального исследовательского центра “Красноярский научный центр СО РАН”
660036 Красноярск, Академгородок, 50, стр. 28, Россия
* E-mail: stanislav@ksc.krasn.ru
Поступила в редакцию 28.09.2018
После доработки 17.10.2019
Принята к публикации 06.11.2019
Аннотация
Впервые для водосборного бассейна в Центральной Эвенкии методом модельных деревьев выявлено изменение состава, структуры и запасов фитомассы и биогенных элементов в лиственничниках в зависимости от возраста древостоев. Определено, что основные массообразующие компоненты в фитомассе всех насаждений – стволовая и корневая фракции, причем с возрастом увеличивается доля стволовой фитомассы и снижаются доли ветвей и хвои. Установлено, что содержание корней мало изменяется с возрастом древостоя, что подтверждает гипотезу о ежегодном преобладающем инвестировании углерода в корни деревьев лиственницы в криолитозоне. Показано, что наибольшие запасы углерода аккумулированы в перестойных насаждениях, азота – в хвое молодняков.
Лиственничные леса севера Евразии относятся к бореальным экосистемам высоких широт, растущим на многолетнемерзлых почвах. Они выполняют почвозащитные и климаторегулирующие функции глобального масштаба, сохраняя нетронутой многолетнюю мерзлоту, таяние даже 10% которой может к концу XXI в. дополнительно увеличить концентрацию углекислого газа в атмосфере на 80 ppm и температуру воздуха на планете на 0.7°С (Raupach, Canadell, 2008), ускорив процесс потепления климата.
Роль высокоширотных лесов в цикле углерода в условиях климатических изменений до сих пор до конца не выяснена (Schulze et al., 1999; Chapin et al., 2000; Jarvis et al., 2001; и др.). Подчеркивается лишь неопределенность, существующая в отношении запасов углерода в северных лесах и их почвах (Вудвелл, Хафтон, 1997), а также тот факт, что их отличительная черта по сравнению с другими лесами – высокая доля (в процентах) корней в общей фитомассе экосистем, что характерно для олиготрофных (особенно по запасам азота) почв (Ruess et al., 1996). Поэтому оценка фактических запасов фитомассы лесов высоких широт и ее распределения по отдельным фракциям важна для понимания изменения потоков углерода и других биогенных элементов при глобальном потеплении. Подобные данные также необходимы для более полного учета лесных ресурсов и обоснования различных лесохозяйственных мероприятий, направленных на выращивание лесов разного назначения.
Оценка фитомассы древостоев по составляющим фракциям – трудоемкий процесс. К настоящему времени в системе Государственного учета лесного фонда (ГУЛФ) накоплен огромный объем данных в основном о запасах стволовой древесины, составлены многочисленные региональные таблицы хода роста (ТХР), сведения же о запасах и продуктивности фитомассы лесов отрывочны и малочисленны (Исаев и др., 1993; Ваганов и др., 2005; Жукова и др., 2008; Демаков и др., 2015; и др.). Ликвидировать эту диспропорцию предложил В.А. Усольцев (Усольцев, 2002) путем сопряжения банков данных ГУЛФ по запасам стволовой древесины и традиционных нормативов (ТХР) с данными о фитомассе лесов на основе многофакторных регрессионных моделей. Последние предусматривают оценку фитомассы или ее переводных коэффициентов по основным массообразующим показателям, входящим в состав ГУЛФ и ТХР. На основе предложенного подхода была дана оценка биологической продуктивности лесных экосистем Северной Евразии, а также был проведен ее географический анализ (Усольцев, 2001, 2013, 2016).
Для лиственничных лесов среднесибирского сектора криолитозоны, сформированных лиственницей Гмелина Larix gmelinii (Rupr.) Rupr., данные о запасах и продуктивности фитомассы древостоев и ее структуре единичны (Kajimoto et al., 1999, 2006; Плешиков и др., 2002; Прокушкин и др., 2008; Данилин, 2009) и не могут адекватно характеризовать особенности фитомассы лесных экосистем региона в целом. С помощью результатов данного исследования мы попытались хотя бы частично заполнить информационное “белое пятно”.
На обширных территориях при характеристике насаждений, оценке запасов фитомассы древостоев и отдельных ее компонентов целесообразно не только использовать классический метод их определения на пробных площадях (ПП), но и совмещать его с бассейновым подходом (Трифонова, 2005). Данный подход позволяет выявить эколого-лесоводственные особенности территории и оценить миграционно-вещественные потоки всего биогеохимического комплекса водосборного бассейна (Путилин, Шкаруба, 2011). В гористых условиях Средней Сибири с развитой сетью водосборных бассейнов малых рек и ручьев применение этого подхода позволяет получать количественные оценки территории и применять ГИС-технологии.
Цель работы – в модельном бассейне на основании лесоустроительных материалов и выполненных наземных исследований провести инвентаризацию лесных фитоценозов, выявить особенности орографического распределения и возрастной структуры древостоев, оценить запас фитомассы лиственничных древостоев и отдельных ее компонентов (корней, стволовой древесины, ветвей, хвои) и определить в них запасы биогенных элементов.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились на территории Нижнетунгусского округа лиственничных и лиственнично-темнохвойных северотаежных лесов Ангаро-Тунгусской таежной провинции Среднесибирской лесорастительной области (Углерод…, 1994). Климат района резко-континентальный со среднегодовой температурой воздуха –9.5°С, средней температурой января –36.7°С, средней температурой июля 16.3°С (Справочник…, 1967, 1969). Годовое количество осадков 322 мм. Почвы, положительные температуры в верхнем 20-сантиметровом слое которых держатся <4 мес. в году, относятся к мерзлотным сезонно-талым или очень холодным и по трофическим свойствам характеризуются как олиготрофные (Абаимов и др., 1996).
Объектами исследования были чистые насаждения лиственницы Гмелина в бассейне руч. Кулингдакан (приток р. Кочечум, 64°19′ с.ш., 100°15′ в.д.), русло которого делит водосборную площадь на две примерно равные части северо-западной и юго-восточной ориентации. Для определения площади выделов и всего водосбора использовали крупномасштабную топографическую карту М 1 : 25 000, оцифрованную с помощью ГИС-технологий, в результате чего были получены векторные слои с привязкой к масштабу (рис. 1).
Рис. 1.
Распределение насаждений и их площади на водосборном бассейне руч. Кулингдакан; числитель – номер выдела, знаменатель – площадь, га. 1 – поймы ручья, 2 – молодняки (<40 лет), 3 – спелые (120–140 лет), 4 – перестойные (>140 лет), 5 – гари, 6 – не покрытые лесом.
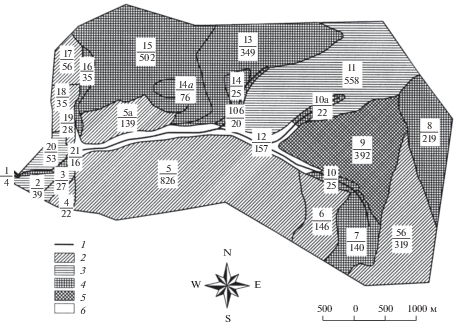
С помощью имеющихся лесоустроительных материалов (Лесохозяйственный…, 2001), дополнительно заложенных топоэкологических профилей и девяти ПП были уточнены таксационные показателей древостоев и дана лесотаксационная характеристика лиственничников во всех выделах с указанием средней высоты (Н, м), диаметра (D, см), полноты и запаса древостоев.
Запасы фитомассы и ее распределение по отдельным компонентам дерева у лиственницы были определены в древостоях разных возрастных групп: молодняках (<40 лет), средневозрастных и приспевающих (50–100 лет), спелых (101–140 лет) и перестойных (>140 лет) (табл. 1). Для учета надземной фитомассы использовали 57 модельных деревьев (табл. 2), у которых отдельно взвешивали стволовую часть и ветви вместе с хвоей. Затем брали каждую пятую охвоенную ветку, у которой хвою отделяли и взвешивали отдельно.
Таблица 1.
Распределение площадей и запасов древостоев лиственницы по возрастным группам в водосборном бассейне
| Возрастные группы | Число выделов | $\frac{{{\text{Площадь, га,}}}}{\begin{gathered} {\text{доля}}\,\,{\text{общей}} \\ {\text{площади,}}\,\,{\text{\% }} \\ \end{gathered} }$ | $\frac{{{\text{Запас*, }}{{{\text{м}}}^{{\text{3}}}}{\text{,}}}}{\begin{gathered} {\text{доля общего}} \\ {\text{запаса, \% }} \\ \end{gathered} }$ | $\frac{{{\text{Запас фитомассы**,}}\,\,{\text{т а}}{\text{.с}}{\text{.м}}{\text{.}}}}{{{\text{доля общего запаса, \% }}}}$ |
|---|---|---|---|---|
| Молодняки (<40 лет) |
9 | $\frac{{2129}}{{57.8}}$ | $\frac{{28{\kern 1pt} 184.9}}{{26.4}}$ | $\frac{{14{\kern 1pt} 797}}{{26.6}}$ |
| Спелые (120–140 лет) |
4 | $\frac{{123}}{{3.4}}$ | $\frac{{6992.4}}{{6.5}}$ | $\frac{{3671}}{{6.6}}$ |
| Перестойные (>140 лет) |
11 | $\frac{{1429}}{{38.8}}$ | $\frac{{71{\kern 1pt} 593.5}}{{67.1}}$ | $\frac{{37{\kern 1pt} 114}}{{66.8}}$ |
| Итого | 24 | $\frac{{3681}}{{100}}$ | $\frac{{106{\kern 1pt} 770.8}}{{100}}$ | $\frac{{55{\kern 1pt} 582}}{{100}}$ |
Примечание. * – данные приведены по лесоустроительным материалам (Лесохозяйственный регламент Эвенкийского лесничества от 01.01.2001), уточненным по таблицам хода роста (Бондарев, 1995) при сплошном перечете на пробных площадях; ** – запасы получены на основании плотности древесины лиственницы 525 кг/м. http://brigadeer.ru/svojstva-drevesiny/plotnost-drevesiny-tablica-i-drugie-fizicheskie-svojstva.html
Таблица 2.
Таксационная характеристика древостоев лиственницы на пробных площадях (ПП) и распределение надземной фитомассы по отдельным компонентам модельных деревьев
| № ПП |
Таксационные показатели древостоев | Число моделей, шт. |
Среднее содержание фитомассы по фракциям* | ||||||
|---|---|---|---|---|---|---|---|---|---|
| возраст, лет |
$\frac{{D{\text{,}}\,\,{\text{см}}}}{{H,\,\,{\text{м}}}}$ | полнота | запас, м3/га |
стволовая | ветви | хвоя | сумма | ||
| Молодняки (<40 лет) | |||||||||
| 1 | 25–40 | $\frac{{2.3}}{{2.7}}$ | 0.6 | 10.0 | 8 | $\frac{{142.7 \pm 55.65}}{{64.5 \pm 5.1}}$ | $\frac{{57.4 \pm 21.93}}{{21.7 \pm 3}}$ | $\frac{{40.3 \pm 17.04}}{{13.8 \pm 2.2}}$ | $\frac{{240.3 \pm 93.73}}{{100}}$ |
| Средневозрастные и приспевающие (50–100 лет)** | |||||||||
| 1а | 50 | $\frac{{6.8}}{{9.5}}$ | 0.6 | 72.8 | 8 | $\frac{{15.8 \pm 6.54}}{{84.6 \pm 0.98}}$ | $\frac{{2.4 \pm 1.17}}{{10.8 \pm 0.78}}$ | $\frac{{0.9 \pm 0.39}}{{4.3 \pm 0.40}}$ | $\frac{{19.0 \pm 8.08}}{{100}}$ |
| 6 | 70–100 | $\frac{{5.8}}{{7.3}}$ | 0.4 | 35.5 | 6 | $\frac{{15.1 \pm 5.88}}{{86.9 \pm 3.59}}$ | $\frac{{0.9 \pm 0.24}}{{10.5 \pm 2.96}}$ | $\frac{{0.3 \pm 0.09}}{{2.8 \pm 0.86}}$ | $\frac{{16.3 \pm 6.2}}{{100}}$ |
| 5 | 90 | $\frac{{6.5}}{{7.2}}$ | 0.4 | 27.7 | 7 | $\frac{{13.5 \pm 5.31}}{{88.9 \pm 1.03}}$ | $\frac{{1.7 \pm 0.8}}{{7.6 \pm 0.83}}$ | $\frac{{0.7 \pm 0.29}}{{6.4 \pm 0.32}}$ | $\frac{{10.7 \pm 3.59}}{{100}}$ |
| 2 | 100 | $\frac{{10.9}}{{11.2}}$ | 0.6 | 79.2 | 9 | $\frac{{8.4 \pm 1.2}}{{87.2 \pm 1.72}}$ | $\frac{{1.1 \pm 0.3}}{{9.6 \pm 1.53}}$ | $\frac{{0.3 \pm 0.08}}{{3.0 \pm 0.37}}$ | $\frac{{9.7 \pm 1.54}}{{100}}$ |
| 3 | 100 | $\frac{{10.5}}{{11.5}}$ | 0.6 | 97.5 | 4 | $\frac{{33.3 \pm 0.55}}{{88.2 \pm 1.56}}$ | $\frac{{4.2 \pm 0.54}}{{8.7 \pm 1.2}}$ | $\frac{{1.5 \pm 0.2}}{{3.1 \pm 0.44}}$ | $\frac{{39.1 \pm 0.19}}{{100}}$ |
| Среднее содержание фитомассы по группе | $\frac{{2.0 \pm 0.59}}{{9.44 \pm 1.03}}$ | $\frac{{0.7 \pm 0.23}}{{3.92 \pm 1.14}}$ | $\frac{{19.0 \pm 5.31}}{{100}}$ | ||||||
| Спелые (100–140 лет) | |||||||||
| 4 | 130 | $\frac{{4.1}}{{4.3}}$ | 0.5 | 22.3 | 9 | $\frac{{7.6 \pm 3.21}}{{85.1 \pm 2.8}}$ | $\frac{{0.7 \pm 0.24}}{{9.7 \pm 1.5}}$ | $\frac{{0.4 \pm 0.12}}{{5.4 \pm 1.26}}$ | $\frac{{8.7 \pm 3.53}}{{100}}$ |
| Перестойные (≥140 лет) | |||||||||
| 7 | 150–260 | $\frac{{11.0}}{{8.6}}$ | 0.4 | 44.1 | 14 | $\frac{{33.4 \pm 11.73}}{{83.7 \pm 4.48}}$ | $\frac{{2.6 \pm 0.64}}{{11.7 \pm 3.16}}$ | $\frac{{1.0 \pm 0.23}}{{4.7 \pm 1.34}}$ | $\frac{{37.0 \pm 12.25}}{{100}}$ |
| 8 | 180 | $\frac{{12.1}}{{10.6}}$ | 0.5 | 72.1 | 10 | $\frac{{71.7 \pm 25.9}}{{90.4 \pm 1.77}}$ | $\frac{{3.7 \pm 1.04}}{{7.6 \pm 1.42}}$ | $\frac{{1.0 \pm 0.24}}{{2.2 \pm 0.43}}$ | $\frac{{76.4 \pm 26.62}}{{100}}$ |
| Среднее содержание фитомассы по группе | $\frac{{52.5 \pm 19.14}}{{87.05 \pm 3.35}}$ | $\frac{{3.1 \pm 0.57}}{{9.65 \pm 2.05}}$ | $\frac{{1.0 \pm 0.01}}{{3.45 \pm 1.25}}$ | $\frac{{56.7 \pm 19.72}}{{100}}$ | |||||
В криолитозоне лиственница Гмелина формирует поверхностную корневую систему. В связи с этим запасы подземной фитомассы определяли путем непосредственной раскопки всей корневой системы у восьми деревьев в молодняках и по одному дереву у лиственниц старших возрастов. Для учета брали все фракции корней: скелетные, проводяшие и физиологически активные окончания. Корни отмывали от почвы и высушивали. Средний образец каждой фракции (древесины стволов, ветвей, хвои и корней) высушивали до абсолютно сухой массы (а.с.м.) и взвешивали.
Оценку запасов фитомассы различных фракций (компонентов) древостоев на территории водосборного бассейна проводили в разных возрастных группах лиственничников на основе регрессионного анализа и соотношений этих компонентов (в процентах) у модельных деревьев (Шмидт, 1984). Основным массообразующим параметром в обоих вариантах расчетов была стволовая древесина. Выбор относительных, а не абсолютных значений был обусловлен высокой индивидуальной изменчивостью отдельных морфологических параметров у деревьев (Kv = 43–74%, где Kv – коэффициент вариации) (Данилин, 2009). При этом возникают существенные трудности для интерпретации данных и обсуждения результатов. Подобный подход позволил точнее оценить общие и специфические закономерности распределения фитомассы отдельных компонентов дерева в зависимости от возраста.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Структура надземной фитомассы древостоев. На территории бассейна все древостои сформировались после сильных низовых пожаров конца XIX–начала XX в. В основном это одновозрастные древостои с полнотой 0.4–0.6 и максимальными запасами 80–100 м3/га (табл. 2). Только в пойме ручья и пониженных местообитаниях встречаются сохранившиеся отдельные деревья и участки лиственничников допожарной генерации в возрасте 180–260 лет. В целом наиболее продуктивные древостои (IV–V бонитеты) сформировались в средней и верхней частях коренного склона юго-западной экспозиции. В 1960–1980-х гг. значительная часть древостоев бассейна была повторно уничтожена катастрофическими лесными пожарами.
Анализ пространственного распределения насаждений лиственницы разных возрастных групп показал, что молодняки занимают 57.8, перестойные древостои – 38.8, а спелые – лишь 3.4% общей площади бассейна (табл. 1). При этом запасы фитомассы стволовой древесины в молодняках составляют 14 797, спелых и перестойных лиственничниках – 3671 и 37 114 т соответственно (табл. 1). Общий запас фитомассы стволовой древесины в рассматриваемом бассейне составил 55 582 т а.с.м.
Исследование распределения запасов фитомассы модельных деревьев лиственницы по отдельным фракциям (корни, стволовая древесина, ветви, хвоя) позволило установить, что во всех возрастных группах в надземной части древостоев преобладает стволовая фракция (табл. 2). При этом с увеличением возраста древостоев происходит существенное увеличение и ее доли в общей фитомассе дерева: если в молодняках стволовая древесина в среднем составляет 64.6 ± 5.1% всей надземной фитомассы дерева, то в спелых и перестойных древостоях она достигает 85.2 ± 1.8% и 89.4 ± 1.4% соответственно. Подобная тенденция была отмечена и в объединенной средневозрастно-приспевающей возрастной группе, где с увеличением возраста древостоев (от 50 до 100 лет) также наблюдалось увеличение доли стволовой древесины (от 83.7 ± 4.48% до 88.2 ± 1.56%). Одновременно в надземной фитомассе древостоев резко уменьшались доли фракций ветвей и хвои (табл. 2). В частности, фитомасса ветвей была снижена в среднем от 21.7 ± 3% в молодняках до 7.7 ± 0.9% в спелых и перестойных древостоях, в то время как фитомасса хвои – основного ассимилирующего аппарата дерева – снижалась от 13.7 ± 2.2% в молодняках до 3.1 ± 0.7% в спелых и перестойных древостоях. Сходная структура фитомассы древостоя в 30-летнем послепожарном молодняке в Центральной Эвенкии была отмечена ранее (Данилин, 2009).
Оценка запасов фитомассы отдельных компонентов лиственницы и общих запасов древостоев. Распределение фитомассы деревьев по отдельным фракциям (ствол, ветви, хвоя, корни) было подробно исследовано ранее (Кузьмичев, 1977; Кофман, 1986; Суховольский, 1996; Бузыкин и др., 2002; Усольцев, 2013, 2016). При этом в связи с большой трудоемкостью измерения фитомассы прямые детальные и систематические исследования фракционного состава были крайне малочисленны, а запасы, как правило, вычисляются на основе установленных закономерностей. Связи между фитомассами различных фракций деревьев обычно описываются регрессионными и аллометрическими уравнениями, наиболее надежная основа для разработки которых – информация о фактической структуре фитомассы модельных деревьев, полученная так называемым деструктивным методом на ПП. Поскольку в разных регионах фракционная структура фитомассы равновеликих деревьев лиственницы различна, а обобщенные (усредненные) ее оценки увеличивают стандартные оценки определения массы отдельных фракций на 19–46% (Усольцев и др., 2013), возникает необходимость получения локальной нормативно-справочной информации о подеревной фитомассе лиственницы.
К основным предикторам в оценке фитомассы дерева относятся такие его таксационные показатели, как диаметр и высота (Кофман, 1986; Уткин и др., 1996; Демаков и др., 2015). К ним добавляют еще протяженность кроны, густоту и возраст древостоев (Богачев, Свалов, 1978) и обусловленные развитием техники дистанционного зондирования диаметр (ширину) кроны, а также диаметр кроны и высоту дерева (Усольцев и др., 2017). Однако в некоторых случаях основной массообразующий параметр – стволовая древесина (Бузыкин и др., 2002; Соловьев и др., 2012).
В наших исследованиях стволовая древесина выбрана в качестве исходного массообразующего показателя как наиболее инертного показателя в повыдельных данных используемой лесоустроительной информации. Были установлены регрессионные зависимости запасов различных фракций фитомассы модельных деревьев лиственницы от стволовой древесины в разных возрастных группах древостоев (рис. 2а–г). Анализ уравнений показывает, что к наиболее приемлемым и соответствующим экспериментальным данным относятся зависимости, установленные для молодняков и средневозрастных древостоев лиственницы (R2 ≥ 0.5, где R – коэффициент корреляции). В спелых и перестойных насаждениях уравнения нельзя признать адекватными фактическим данным: доля необъясненной дисперсии в них составляет 52–78% при коэффициентах детерминации R2 ≥ 0.22–0.48 (рис. 2). Вероятно, это связано с недостаточным числом взятых для анализа модельных деревьев в этих возрастных группах (5 в спелых и 8 в перестойных древостоях) (табл. 2) или с влиянием неучтенного разнообразия широкого спектра эндогенных факторов деревьев и лесорастительных условий. Коэффициенты корреляции (R = 0.69–0.79) указывают на высокую (тесную) зависимость запасов фитомассы ветвей и хвои от стволовой древесины в первых двух группах древостоев (рис. 2а, б). Аналогичные зависимости в спелых и перестойных древостоях демонстрируют слабую (умеренную) и среднюю (значительную) тесноту связи (R = 0.47–0.69).
Рис. 2.
Зависимость запасов фитомассы отдельных компонентов дерева от стволовой фитомассы у лиственницы в молодняках (а), средневозрастных (б), спелых (в) и перестойных (г) насаждениях. 1 – ветви, 2 – хвоя, 3 – корни.
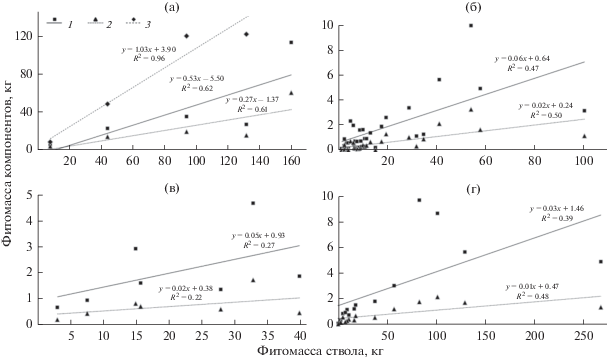
Особо следует отметить очень высокую корреляцию (R = 0.98) между запасами фитомассы хвои и физиологически активных корней диаметром ≥1 мм в молодняках (рис. 2а). Такая взаимосвязь фитомассы основных органов обеспечения жизнедеятельности деревьев вполне объяснима в период интенсивного роста и развития молодых лиственничников.
Высокая и очень высокая корреляция (0.70 ≤ R ≤ ≤ 0.99) между морфологическими показателями деревьев, их фитомассой, а также между отдельными фракциями фитомассы в молодом послепожарном лиственничнике в Центральной Эвенкии была отмечена ранее (Данилин, 2009).
Все вышеизложенное позволяет заключить, что установленные регрессионные зависимости для оценки различных фракций фитомассы модельных деревьев лиственницы от стволовой древесины применимы только для определения запасов корней, ветвей и хвои в молодняках и ветвей и хвои в средневозрастных древостоях. Использование полученных регрессионных уравнений для определения аналогичных показателей в спелых и перестойных лиственничниках не корректно. Поэтому в дальнейших расчетах нами применен следующий методический прием: для модельных деревьев каждой возрастной группы лиственничников впервые выявлены характерные для них определенные соотношения содержания фитомассы отдельных компонентов деревьев в зависимости от стволовой массы, которая принимается за 100% (табл. 3). Подобные соотношения известны для некоторых древесных пород. В частности, между объемами пней и стволов они составляют в среднем у сосны 18–25, ели 25–30, березы и осины 22–24% (Жукова и др., 2008), соотношение корней и стволовой фитомассы сосны обыкновенной в зависимости от условий произрастания варьирует от 25 до 34% (Орлов, Кошельков, 1971).
Таблица 3.
Распределение фитомассы отдельных компонентов у лиственницы разного возраста в зависимости от массы ствола на пробных площадках (ПП)
| № ПП |
Характеристика ПП | Среднее содержание фитомассы, доля массы ствола, % | ||||
|---|---|---|---|---|---|---|
| возрастная группа древостоев |
число моделей, шт. | ствол | ветви | хвоя | корни | |
| 1 | Молодняки | 9 | 100 | 38.7 ± 7.6 | 23.9 ± 5.1 | 101.9 ± 11.2 |
| 2 | Средневозрастные и приспевающие | 30 | 100 | 12.8 ± 1.7 | 4.6 ± 0.7 | 86.3* |
| 3 | Спелые | 9 | 100 | 12.7 ± 2.6 | 4.5 ± 0.78 | 93.7* |
| 4 | Перестойные | 14 | 100 | 8.9 ± 1.21 | 3.7 ± 0.87 | 93.7* |
В лиственничниках Центральной Эвенкии доли фитомассы ветвей и хвои в фитомассе стволовой древесины в молодняках составляют 38.7 и 23.9%, снижаются в спелых и перестойных насаждениях до 12.7–8.9 и 4.5–3.7% соответственно (табл. 3). Соотношение фитомасса корней/фитомасса стволов с возрастом древостоев снижается незначительно: со 101.9% в молодняках до 86–93% в спелых и перестойных насаждениях (табл. 3), т.е. содержание корней во всех случаях остается достаточно высоким. Вероятно, справедливо предположение о ежегодном преобладающем инвестировании углерода в корни деревьев лиственницы в условиях лимита элементов минерального питания растений на многолетнемерзлых почвах Сибири (Kanazawa et al., 1994; Kajimoto et al., 1999). В связи с этим у лиственницы Гмелина в процессе онтогенеза формируется мощная корневая система, определяющая ее устойчивость к экстремальным эдафическим условиям криолитозоны.
Установленные в нашем случае соотношения в целом повторяют выявленные выше изменения фракционного состава фитомассы модельных деревьев с возрастом. Это логично, поскольку запас фитомассы стволовой древесины, преобладающей фракции всех возрастных групп, изменяется пропорционально общему запасу надземной фитомассы.
С использованием процентных соотношений фитомассы отдельных компонентов в модельных деревьях (табл. 3) и имеющихся данных о запасах фитомассы стволовой древесины (табл. 1) проведена оценка запасов основных компонентов надземной и подземной фитомассы древостоев в изучаемом бассейне (табл. 4). Выявлено, что общий запас фитомассы всех компонентов древостоев составляет 123 174.5 т, из них 45.1% приходится на стволовую древесину, 43.1% – на корни, 7.7% – на ветви и 4.1% – на хвою. Надземная фитомасса составляет 56.9% общей фитомассы древостоев. Наши результаты находятся в хорошем соответствии с данными, полученными ранее для данного региона (Kajimoto et al., 1999). Согласно результатам расчетов, проведенных с использованием аллометрических взаимосвязей, стволовая древесина составляет 46.4%, ветви – 8.2%, хвоя – 2.6% общей фитомассы древостоев, соотношение надземная/подземная фитомасса – 57.2%/42.8% (Kajimoto et al., 1999). В нашем случае расчеты выполнялись на основе процентных соотношений между стволовой древесиной и компонентами (ветви, хвоя) модельных деревьев лиственницы. Следовательно, этот подход не уступает по точности аллометрическим расчетам и может применяться даже при средней (R = 0.47–0.69) корреляционной зависимости между признаками.
Таблица 4.
Запасы фитомассы отдельных компонентов древостоев лиственницы на территории бассейна, т а.с.м.: сравнение двух вариантов расчета
| Фракция | Группа | |||
|---|---|---|---|---|
| молодняки | спелые | перестойные | итого | |
| Корни* | $\frac{{15{\kern 1pt} 244.8}}{{15{\kern 1pt} 078.2}}$ | $\frac{ - }{{3168.1}}$ | $\frac{ - }{{34{\kern 1pt} 775.8}}$ | $\frac{{15{\kern 1pt} 244.8}}{{53{\kern 1pt} 022.1}}$ |
| Хвоя* | $\frac{{3993.8}}{{3536.4}}$ | $\frac{{73.8}}{{165.1}}$ | $\frac{{371.6}}{{1373.2}}$ | $\frac{{4439.2}}{{5074.7}}$ |
| Ветви* | $\frac{{7836.9}}{{5726.4}}$ | $\frac{{184.5}}{{466.2}}$ | $\frac{{1114.9}}{{3303.1}}$ | $\frac{{9136.3}}{{9495.6}}$ |
| Ствол** | $\frac{{14{\kern 1pt} 797.0}}{{14{\kern 1pt} 797.0}}$ | $\frac{{3671.0}}{{3671.0}}$ | $\frac{{37{\kern 1pt} 114.0}}{{37{\kern 1pt} 114.0}}$ | $\frac{{55{\kern 1pt} 582.0}}{{55{\kern 1pt} 582.0}}$ |
| Итого | $\frac{{41{\kern 1pt} 872.5}}{{39{\kern 1pt} 138.0}}$ | $\frac{{3929.3}}{{7470.4}}$ | $\frac{{38{\kern 1pt} 600.5}}{{76{\kern 1pt} 566.1}}$ | $\frac{{84{\kern 1pt} 402.3}}{{123{\kern 1pt} 174.5}}$ |
Отметим, что в лиственничниках криолитозоны запас подземной фитомассы сопоставим с запасом стволовой древесины (табл. 4), а общий запас фитомассы древостоя в биогеоценозах значительно меньше суммарных запасов напочвенного покрова, которые в бассейне составляют 42 234 т (травяно-кустарничковый и мохово-лишайниковый покровы) и 133 661 т (подстилка) (Прокушкин, Зырянова, 2017).
Сравнительная оценка запасов фитомассы различных компонентов древостоев лиственницы, относящихся к разным возрастным группам (табл 4), позволила сделать вывод, что оба метода расчета наиболее адекватно “работают” в молодняках (R2 = 0.61–0.96, 0.78 < R < 0.98): расчетные значения фитомассы для корней, хвои и ветвей, полученные с помощью регрессионных уравнений, превышают таковые для процентных соотношений на 1, 13 и 37% соответственно, а итоговые цифры различаются всего на 7% (табл. 4). В спелых (R2 = 0.22–0.27, R = 0.47–0.52) и перестойных (R2 = 0.39–0.48, R = 0.62–0.69) лиственничниках, наоборот, значения показателей фитомассы для хвои и ветвей, рассчитанные с помощью регрессионных уравнений, ниже значений, рассчитанных с помощью процентных соотношений. При этом разница весьма существенная: в 2.2–3.7 раза для хвои, в 2.5–2.9 раза для ветвей и 1.9–2 раза в итоговых цифрах спелых и перестойных древостоев соответственно (табл. 4). Общие запасы фитомассы всех компонентов древостоев на территории бассейна, вычисленные с помощью уравнений регрессии, в 1.5 раза ниже значений, вычисленных на основе процентных соотношений.
Оценка запасов биогенных элементов. Для оценки запасов углерода, азота, фосфора, калия в фитомассе древостоев использовали сведения о содержании этих элементов в отдельных компонентах лиственницы (Митрофанов, 1977; Прокушкин и др., 2008, 2010, 2014; Matsuura et al., 2010). С помощью этих данных выполнили расчет содержания биогенных элементов в стволах, ветвях, хвое и корнях лиственницы в различных возрастных группах древостоев и определили их общий запас на территории исследуемого бассейна (рис. 3).
Рис. 3.
Запасы биогенных элементов в фитомассе древостоев лиственницы в водосборном бассейне, т а.с.м. А – молодняки, Б – спелые, В – перестойные, Г – итого в бассейне. 1 – корни, 2 – хвоя, 3 – ветви, 4 – ствол, 5 – итого.
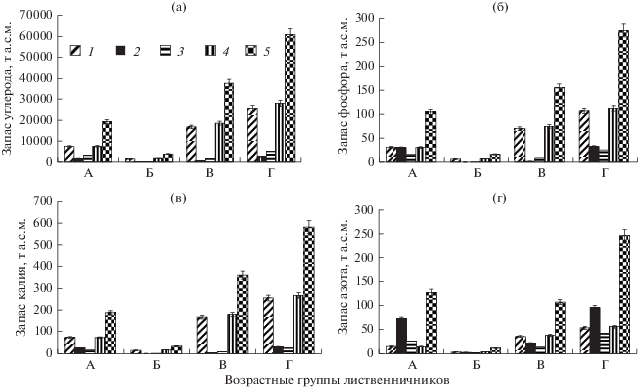
Анализ полученных данных показал, что в фитомассе древостоев на территории бассейна аккумулировано >60.7 тыс. т углерода (рис. 3а). Основные его запасы находятся в стволовой древесине и корнях – 48.8 и 42.2% соответственно. Запасы углерода в хвое и ветвях на порядок ниже и составляют 4.1 и 4.9% общего его содержания. Среди возрастных групп лиственничников максимальные запасы углерода сосредоточены в перестойных древостоях (рис. 3а).
По сравнению с запасами углерода запасы фосфора и калия в фитомассе древостоев бассейна значительно меньше и составляют 273.4 и 580.4 т соответственно (рис. 3б, в). Однако накопление этих элементов в компонентах древостоев аналогично структуре запасов углерода, а градиент распределения запасов имеет следующий вид: стволовая древесина > корни > хвоя > ветви. Максимальный вклад в накопление фосфора и калия в древостоях бассейна также вносят перестойные насаждения.
Иные закономерности отмечены в отношении запасов азота. Локализация азота происходит в основном в хвое, где его количество достигает 94.9 т (рис. 3г). При этом наибольшее его накопление отмечено в молодняках. Запас азота в скелетных корнях, стволовой древесине и ветвях составляет 55.6, 53.0, 41.8 т соответственно. В целом в лиственничных древостоях данного бассейна содержится 60 687.9 т углерода, 245.2 т азота, 273.4 т фосфора и 580.4 т калия.
ЗАКЛЮЧЕНИЕ
В результате исследования, проведенного на территории экспериментального водосборного бассейна, получена количественная оценка распределения его площадей по возрастным группам древостоев лиственницы. Для каждой группы определены состав, структура, запасы стволовой древесины и фитомассы модельных деревьев по фракциям (хвоя, ветви, стволы, корни). На основании результатов регрессионного анализа и процентных соотношений изучаемых фракций выявлены запасы фитомассы этих компонентов для древостоев всего бассейна.
Общий запас надземной и подземной фитомассы древостоев по сравнению с живым напочвенным покровом существенно меньше и составляет 123 174.5 т а.с.м. При этом основными массообразующими компонентами являются стволовая и корневая фракции – 45.1 и 43.1% соответственно – с максимальным накоплением их фитомассы в древостоях старших возрастных групп. Запасы других фракций (ветвей и хвои) в древостоях невелики.
В целом структура фитомассы деревьев с возрастом меняется: с его увеличением растет доля стволовой и корневой массы с одновременным снижением доли ветвей и особенно хвои. Максимальные запасы фитомассы стволов, корней и ветвей выявлены в перестойных древостоях, а хвои – в молодняках. Запасы подземной фитомассы в спелых и перестойных лиственничниках сопоставимы с запасами стволовой древесины, в то время как в молодняках они их превышают.
Среди изучаемых биогенных элементов в фитомассе древостоев преобладает углерод, запасы которого аккумулированы в основном в стволовой древесине и скелетных корнях спелых и перестойных насаждений. Запасы фосфора и калия распределены в фитомассе аналогично запасу углерода, а запасы азота сосредоточены преимущественно в хвое и мелких корнях.
Часть этих элементов, и особенно азота, в результате ежегодного отпада хвои, мелких ветвей и физиологически активных корней возвращается в почву и включается в биологический круговорот. Значительная часть других элементов законсервирована на многие десятилетия в стволах, скелетных корнях и крупных ветвях.
Список литературы
Абаимов А.П., Прокушкин С.Г., Зырянова О.А. Эколого-фитоценотическая оценка воздействия пожаров на леса криолитозоны Средней Сибири // Сиб. экол. журн. 1996. № 1. С. 51‒60.
Богачев А.В., Свалов Н.С. Методы таксации лесного и лесосечного фонда // Итоги науки техники. Сер. Лесоведение и лесоводство. М.: ВИНИТИ, 1978. Т. 2. С. 7–209.
Бондарев А.И. Строение и нормативы таксации притундровых лесов северо-востока Красноярского края: Автореф. дис. канд. биол. наук. Красноярск, ИЛ СО РАН, 1995. 19 с.
Бузыкин А.И., Пшеничникова Л.С., Суховольский В.Г. Густота и продуктивность древесных ценозов. Новосибирск: Наука, 2002. 152 с.
Ваганов Е.А., Ведрова Э.Ф., Верховец С.В., Ефремов С.П., Ефремова Т.Т., Круглов В.Б., Онучин А.А., Сухинин А.И., Шибистова О.Б. Леса и болота Сибири в глобальном цикле углерода // Сиб. экол. журн. 2005. № 4. С. 631‒649.
Вудвелл Д.М., Хафтон Р.Э. Тайна великого северного леса. Устойчивое развитие бореальных лесов, Москва. М.: Федеральная служба лес. хоз-ва РФ, 1997. С. 39.
Данилин И.М. Структурно-функциональная организация лиственничного фитоценоза послепожарной восстановительной сукцессии на севере Средней Сибири // Сиб. экол. журн. 2009. № 1. С. 77‒90.
Демаков Ю.П., Пуряев А.С., Черных В.Л., Черных Л.В. Использование аллометрических зависимостей для оценки фитомассы различных фракций деревьев и моделирования их динамики // Вестн. Поволж. гос. технол. ун-та. Сер: Лес. Экология. Природопользование. 2015. № 2(26). С. 19‒36.
Жукова А.И., Григорьев И.В., Григорьева О.И., Ледяева А.С. Лесное ресурсоведение. СПб.: ГЛТА, 2008. 215 с.
Исаев А.С., Коровин Г.Н., Уткин А.И., Пряжников А.А., Замолодчиков Д.Г. Оценка запасов и годичного депонирования углерода в фитомассе лесных экосистем России // Лесоведение. 1993. № 5. С. 3‒10.
Кофман Г.Б. Рост и форма деревьев. Новосибирск: Наука, 1986. 211 с.
Кузьмичев В.В. Закономерности роста древостоев. Новосибирск: Наука, 1977. 160 с.
Лесохозяйственный регламент Эвенкийского лесничества от 01.01 2001 г. Красноярск: Красноярск. управление лес. хоз-ва, 2001. 250 с.
Митрофанов Д.П. Химический состав лесных растений Сибири. Новосибирск: Наука, 1977. 220 с.
Орлов А.Я., Кошельков С.П. Почвенная экология сосны. М.: Наука, 1971. 332 с.
Плешиков Ф.И., Каплунов В.Я., Токмаков С.В. Структура фитомассы и годичная продукция северных лесов. Лесные экосистемы Енисейского трансекта. Новосибирск: Изд-во СО РАН, 2002. С. 73‒83.
Прокушкин С.Г., Зырянова О.А. Оценка запасов фитомассы в лиственничниках Центральной Эвенкии (на примере малого водосборного бассейна) // Вестн. КрасГАУ. 2017. № 9. С. 122‒128.
Прокушкин С.Г., Абаимов А.П., Прокушкин А.С. Структурно-функциональные особенности лиственницы Гмелина в криолитозоне Центральной Эвенкии. Красноярск. Ин-т леса им. В.Н. Сукачева СО РАН, 2008. 161 с.
Прокушкин С.Г., Прокушкин А.С., Сорокин Н.Д. Интенсивность разложения отдельных компонентов фитодетрита в лиственничниках криолитозоны Средней Сибири // Изв. РАН. Сер. биол. 2014. № 1. С. 76‒92.
Прокушкин С.Г., Бугаенко Т.Н., Прокушкин А.С., Шкикунов В.Г. Сукцессионная трансформация растительного и почвенного покрова на солифлюкционных площадях в криолитозоне Центральной Эвенкии // Изв. РАН. Сер. биол. 2010. № 1. С. 93‒104.
Путилин А.Ф., Шкаруба А.М. Водосборный бассейн как геосистемная основа изучения миграционно-вещественных потоков // Сиб. экол. журн. 2011. № 5. С. 673‒676.
Соловьев В.А., Чан Тхи Тху Нян, Шорохова Е.В. Распределение углерода по фракциям фитомассы различных древостоев и лесного массива // Изв. СПб. лесотехн. академии. 2012. № 198. С. 33‒40.
Справочник по климату СССР. Вып. 21. Л.: Гидрометеоиздат, 1967. Ч. II; 1969. Ч. III.
Суховольский В.Г. Фракционная структура и продуктивность фитомассы деревьев и насаждений // Лесоведение. 1996. № 1. С. 30‒40.
Трифонова Т.А. Развитие бассейнового подхода в почвенных и экологических исследованиях // Почвоведение. 2005. № 9. С.1054‒1061.
Углерод в экосистемах лесов и болот России / Ред. Алексеев В.А., Бердси Р.А. Красноярск: ИЛ СО РАН, 1994. 225 с.
Усольцев В.А. Фитомасса лесов Северной Евразии: база данных и география. Екатеринбург: УрО РАН, 2001. 708 с.
Усольцев В.А. Фитомасса лесов Северной Евразии: нормативы и элементы географии. Екатеринбург: УрО РАН, 2002. 763 с.
Усольцев В.А. Вертикально-фракционная структура фитомассы деревьев. Исследование закономерностей. Екатеринбург: УрО РАН, 2013. 602 с.
Усольцев В.А. Фитомасса модельных деревьев лесообразующих пород Евразии: база данных, климатически обусловленная география, таксационные нормативы. Екатеринбург: УрО РАН, 2016. 335с.
Усольцев В.А., Колчин К.В., Маленко А.А. Смещения всеобщих аллометрических моделей при локальной оценке фитомассы деревьев лиственницы // Вестн. Алтайск. гос. аграрн. ун-та. 2017. № 4 (150). С. 85‒90.
Уткин А.И., Замолодчиков Д.Г., Гульбе Т.А., Гульбе Я.Н. Аллометрические уравнения для фитомассы по данным деревьев сосны, ели, березы и осины в европейской части России // Лесоведение. 1996. № 6. С. 36‒46.
Шмидт В.М. Математические методы в ботанике. Л.: Изд-во ЛГУ, 1984. С. 87‒105.
Chapin F.S., III, Mcguire A.D., Randerson J., Pielke S.R.R., Baldocchi D., Hobbie S.E., Roulet N., Eugster W., Kasischke E., Rastetter E.B., Zimov S.A., Running S.W. Arctic and boreal ecosystems of western North America as components of the climate system // Glob. Change Biol. 2000. V. 6. P. 211‒223.
http://brigadeer.ru/svojstva-drevesiny/plotnost-drevesiny-tablica-i-drugie-fizicheskie-svojstva.html
Jarvis P.G., Saugier B., Schulze E.-D. Productivity of boreal forests // Terrestrial global productivity / Eds Roy J., Saugier B., Mooney H.A. San Diego: Acad. Press, 2001. P. 211‒244.
Kajimoto T., Matsuura Y., Sofronov M.A., Volokitina A.V., Mori S., Osawa A., Abaimov A.P. Above- and belowground biomass and net primary productivity of a Larix gmelinii stand near Tura, central Siberia // Tree Physiol. 1999. № 19. P. 815‒822.
Kajimoto T., Matsuura Y., Osawa A., Abaimov A.P., Zyryanova O.A., Isaev A.P., Yefremov D.P., Mori S., Koike T. Size-mass allometry and biomass allocation of two larch species growing on the continuous permafrost region in Siberia // Forest Ecol. Manage. 2006. V. 222. P. 314‒325. https://doi.org/10.1016/j.foreco.2005.10.031
Kanazawa Y., Osawa A., Ivanov B.I, Maximov T.C. Biomass of a Larix gmelinii (Rupr.) Litv. stand in Spaskayapad, Yakutsk // Proc. 2nd Sympos. on joint Siberian permafrost studies between Japan and Russia in 1993. Tsukuba, Japan, 1994. P. 153.
Matsuura Y., Hirobe M. Soil carbon and nitrogen, and characteristics of soil active layer in Siberian permafrost region // Permafrost ecosystems: Siberian larch forests. Ecological studies 209 / Eds Osawa A., Zyryanova O.A., Matsuura Y., Kajimoto T., Wein R.W. Dordrecht; Heidelberg; London; New York: Springer, 2010. P. 149‒163.
Raupach M.R., Canadell J.G. Observing a vulnerable carbon cycle // The continental-scale greenhouse gas balance of Europe / Eds Dolman H., Valentini R., Freibauer A. Berlin: Springer, 2008. P. 5–32.
Ruess R.W., Van Cleve K., Yarie J., Viereck L.A. Contributions of fine root production and turnover to the carbon and nitrogen cycling in taiga forests of the Alaskan interior // Can. J. Forest Res. 1996. V. 26. P. 1326‒1336.
Schulze E.-D., Lloyd J., Kelliher F.M., Wirth C., Rebmann C., Luhker B., Mund M., Knohl A., Milyukova I.M., Schulze W., Ziegler W., Varlagin A.B., Cogachev A.F., Valentini R., Dore S., Grigoriev S., Kolle O., Panfyorov M.I., Tchebakova N.M., Vygodskaya N.N. Productivity of forests in Eurosiberian boreal region and their potential to act as a carbon sink – a synthesis // Glob. Change Biol. 1999. V. 5. P. 703‒722.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая