

Известия РАН. Серия биологическая, 2021, № 2, стр. 200-210
Радиальный прирост сосны обыкновенной Pinus sylvestris L. в северотаежных лишайниковых сосновых лесах и редколесьях
В. В. Горшков 1, 2, Н. И. Ставрова 1, П. Н. Катютин 1, *, А. Ю. Лянгузов 3
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Профессора Попова, 2, Россия
2 Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
194021 Санкт-Петербург, Институтский пер., 5, литер У, Россия
3 Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7–9, Россия
* E-mail: PauRussia@binran.ru
Поступила в редакцию 04.04.2018
После доработки 03.07.2018
Принята к публикации 03.07.2018
Аннотация
Исследованы виталитетная структура древостоев и годичный прирост стволов особей сосны обыкновенной разных категорий состояния в северотаежных средневозрастных лишайниковых сосновых лесах и редколесьях (Кольский п-ов). Установлено, что в древостоях сосновых лесов и редколесий по числу преобладают (60–75%) ослабленные и сильно ослабленные особи, доля здоровых не превышает 20%, но по запасу составляет не менее 30%. Отмечено, что через 30–85 лет после пожара радиальный прирост деревьев сосны в изученных сообществах снижается в 1.5–3.5 раза, а площадь годичного кольца увеличивается в 1.5–5 раз. Обнаружено, что дифференциация деревьев разных категорий состояния по приросту достоверно проявляется начиная с 50 лет после пожара. Зафиксировано, что соотношение значений площади годичного кольца здоровых, ослабленных, сильно ослабленных и усыхающих особей в сосновых лесах и редколесьях составляет соответственно 8 : 5 : 2.5 : 1 и 11 : 5.5 : 3.5 : 1. Показано, что различия ресурсных потенциалов сосновых редколесий и лесов (1 : 1.6) проявляются на уровне сообществ, но не проявляются на уровне отдельных особей.
Годичный прирост деревьев формируется под влиянием большого числа внутренних и внешних по отношению к особи факторов. В качестве основных внутренних факторов выступают видовая специфика, генетические различия, возраст, онтогенетическое состояние (Крамер, Козловский, 1983; Methods …, 1990; Ваганов, Шашкин, 2000; Vaganov et al., 2006). Среди внешних факторов выделяются внутриценотические, связанные с внутри- и межвидовой конкуренцией особей в лесных сообществах; экотопические, обусловленные характеристиками почвообразующих пород, почв и климата; природные катастрофические (массовые ветровалы, пожары, инвазии вредителей) и антропогенные (рубки, пожары, мелиорации, промышленное загрязнение) нарушения (Methods …, 1990; Schweingruber, 1996).
Ряд аспектов влияния внешних факторов на прирост древесных растений привлекает особое внимание исследователей. Это прежде всего касается климатических изменений (Битвинскас, 1974; Fritts, 1976; Бузыкин и др., 1986; Шиятов, 1986; Ваганов и др., 1996; Briffa et al., 1996; Monserud, Sterba, 1996; Шашкин, Ваганов, 2000; Моисеев, 2002; Алексеев, Сорока, 2003; Кузьмин и др., 2011; Николаева и др., 2015; Vacek et al., 2017) и различных видов антропогенного воздействия (Björkdahl, Eriksson, 1989; Ярмишко, 1997; Pärn, 2003; Ярмишко, Ярмишко, 2004; Wilczyński, 2006; Danek, 2007; Черненькова и др., 2012; Juknys et al., 2014; Махныкина и др., 2017; Ярмишко и др., 2017; Vacek et al., 2017).
В меньшей степени изучена проблема связи показателей роста с внутриценотическими факторами, конкурентными отношениями особей в составе древостоев и ценопопуляций древесных видов (Буяк, Карпов, 1983; Бузыкин и др., 1986; Nilsson, Gemmel, 1993; Monserud, Sterba, 1996; Bigler, Bugmann, 2003; Горшков, Катютин, 2009; Das, 2012), проявляющимися, в частности, в естественной дифференциации деревьев по степени развития и уровню жизненного состояния (Алексеев, 1989; Solberg, 1999; Бебия, 2000; Торлопова, Ильчуков, 2003; Dobbertin, 2005; Демидко, 2006; Демидко и др., 2010; Ставрова и др., 2010; Горшков и др., 2013).
Цель исследования – оценка возрастной динамики радиального прироста (по ширине и площади годичных слоев) деревьев сосны обыкновенной разных категорий жизненного состояния в средневозрастных северотаежных сосновых лесах и редколесьях.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование выполнено на территории Кольского п-ова в районе среднего течения р. Лива (67°30′–68°10′ с.ш., 33°57′–34°21′ в.д.) в лишайниковых сосновых лесах с суммой площадей сечений 14–16 м2/га и лишайниковых сосновых редколесьях с суммой площадей сечений 7–9 м2/га и средней плотностью соответственно 1200 и 650 шт./га (табл. 1), сформировавшихся после катастрофических пожаров 20-х годов прошлого века. На территории исследований не проводились лесохозяйственные мероприятия.
Таблица 1.
Характеристики древостоев Pinus sylvestris в лишайниковых сосновых лесах и редколесьях
| № ПП |
Размер ПП, га | Древесный ярус | ||||
|---|---|---|---|---|---|---|
| возраст, лет | D1.3, см | высота, м | число деревьев, экз./га | S, м2/га | ||
| Сосняк лишайниковый | ||||||
| 97 | 0.10 | 70 | 10.8 | 9.8 | 1670 | 15.29 |
| 82 | 0.15 | 313* | 35.6 | 17.7 | 40 | 3.98 |
| 76 | 13.5 | 12.6 | 667 | 9.54 | ||
| 81 | 0.10 | 315* | 48.1 | 14.0 | 10 | 1.82 |
| 76 | 13.0 | 10.1 | 1080 | 14.33 | ||
| Сосновое лишайниковое редколесье | ||||||
| 56 | 0.10 | 68 | 10.1 | 8.0 | 970 | 7.77 |
| 92 | 0.15 | 188* | 34.2 | 12.0 | 13 | 1.19 |
| 85 | 11.8 | 9.2 | 453 | 4.95 | ||
| 96 | 0.15 | 73 | 20.0 | 9.3 | 293 | 9.20 |
Лишайниковые сосновые леса и редколесья распространены на вершинах и склонах холмов и речных террасах, сложенных песчаными, ледниковыми и водно-ледниковыми отложениями. На этих высоководопроницаемых и бедных элементами минерального питания почвообразующих породах под сосновыми лесами и редколесьями формируются почвы, относящиеся к отделу Al–Fe-гумусовых почв: подзолы иллювиально-железистые ненасыщенные, мелкоподзолистые, иллювиально-малогумусовые, песчаные, среднескелетные (Шишов и др., 2004). Они отличаются коротким почвенным профилем (до 60 см) и накоплением оксалаторастворимых соединений Al и Fe в иллювиальном горизонте при низком содержании в нем (не более 1.5%) фульватного гумуса (Никонов, 1987; Переверзев, 2004).
В исследованных сообществах основу древесного яруса (70–100% по запасу) составляют особи Pinus sylvestris L. послепожарного происхождения. Допожарный компонент, возраст которого превышает давность последнего пожара, представлен единичными экземплярами только в трех сообществах (табл. 1). В травяно-кустарничковом ярусе преобладают Vaccinium vitis-idea L., Vaccinium myrtillus L., Empetrum hermaphroditum Hagerup. В мохово-лишайниковом покрове преобладают лишайники рода Cladonia (C. stellaris (Opiz.) Brodo, C. rangiferina (L.) Nyl., C. mitis (Sandst.) Hustich). Среди мхов встречаются в основном Pleurozium schreberi (Brid) Mitt. и Dicranum polysetum Sw.
Исследования выполнены на 6 постоянных пробных площадях (ППП) размером 0.10–0.15 га (3 ППП в лишайниковых лесах и 3 ППП в лишайниковых редколесьях). На каждой пробной площади у 30–100 деревьев сосны обыкновенной в возрасте 70–80 лет (первое послепожарное поколение, формирующее основу древостоя) буравом Пресслера отбирали керны на высоте 1.3 м. Измерение радиального прироста проводилось по общепринятым методикам (Шиятов, 1986; Methods …, 1990) с использованием микроскопа МБС-10 и полуавтоматической установки LINTAB-6 с точностью 0.01 мм.
Категории жизненного состояния 70–80-летних деревьев сосны обыкновенной определялись с учетом имеющихся рекомендаций (Алексеев В.А., 1989; Алексеев А.С., 1990; Ярмишко, 1997; Мето-ды …, 2002; Ярмишко и др., 2003; Санитарные …, 2006) на основе сравнения плотности кроны с плотностью кроны эталонной особи, принимаемой за единицу. Эталонная особь представляет собой растение с минимальным угнетением, которое максимально реализует потенциальные возможности роста и развития вида в данных условиях местопроизрастания. Плотность ее кроны максимальна для данного вида. Выделяли 5 категорий состояния: I – неугнетенные (здоровые) особи – относительная плотность кроны (CD) > > 0.75–1.0, II – умеренно угнетенные (умеренно ослабленные) особи – CD > 0.5–0.75, III – сильно угнетенные (сильно ослабленные) особи – CD > > 0.25–0.5, IV – усыхающие особи – CD > 0–0.25, V – сухие особи.
Оценка участия деревьев разного состояния в формировании древесного яруса проводилась как по числу особей, так и по запасу древесины. Доля участия каждого дерева в формировании общего запаса рассчитывалась на основе условной величины – произведения площади ствола на высоте 1.3 м на высоту дерева.
Для расчета площади годичного кольца деревьев могут быть использованы разные методические подходы (Волков, Тартаковский, 2011). В данной работе на основании измеренных величин рассчитывали средние показатели линейного радиального прироста и площади годичного кольца древесины у живых деревьев P. sylvestris, согласно разработанному нами методу (Лянгузов и др., 2017). Была проанализирована динамика радиального роста особей в интервале давности пожара 30–85 лет. Исходные данные были сгруппированы по типам леса (сосновое редколесье и сосняк лишайниковый) и категориям жизненного состояния особей.
Для каждого дерева вычисляли приближенные значения площади годичных колец, предполагая, что годичное кольцо представляет собой разность кругов с соседними радиусами. Годичный радиальный прирост (P) каждого дерева был представлен числовым рядом P1, …, Pn (мм) за период от условно первого года (первый радиус R1) до последнего. Радиусы колец (R) вычисляли последовательным суммированием R1 = P1, R2 = R1+ P2, R3 = R2+ P3, …, Rn = Rn – 1 + Pn (или обратной процедурой вычитания из полного радиуса ствола Rn – 1 = Rn – Pn …). Затем последовательно вычисляли площади кругов по формуле Si = $\pi R_{i}^{2}$. Для расчета площади годичного прироста (dSi) древесины из площади большего вычитали площадь меньшего круга (dSi = Si – Si – 1).
Для обработки и представления результатов использовали написанный нами скрипт на свободном языке программирования R (The Comprehensive…). Скрипт решал задачи предварительной сортировки данных, вычисления площади годичных колец, группировки значений по статусу деревьев и временны́м интервалам, оценивания параметров распределения показателей с помощью бутстрэп-метода, построения графиков.
Для выявления основных закономерностей изменения (или стабильности) исследуемых показателей значения массивов данных разделяли по временны́м интервалам (1945–1966–1982–1996–2015 гг.) на четыре группы. Для анализа результатов с помощью метода бутстрэпа оценивали медианное (непараметрическое среднее) значение показателя (толщина и площадь годичного кольца) в каждой группе и процентили (2.5, 17, 83, 97.5) выборки. Найденные значения представляли графически в виде “ящик-с-усами”. Значимость различий показателей в разных группах оценивали с использованием медианного теста.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
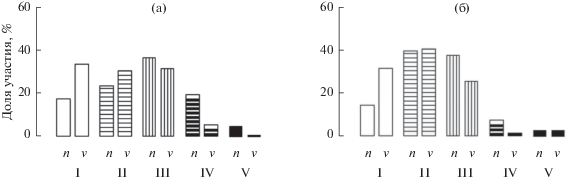
Распределения деревьев сосны обыкновенной по категориям жизненного состояния в двух разных условиях местопроизрастания достоверно различаются (медианный тест: MMT = 8.59, p < 0.01). В лишайниковых лесах в составе древесного яруса по числу преобладают сильно ослабленные (40%) особи и приблизительно одинаково (~20%) представлены здоровые, ослабленные и усыхающие (рис. 1а). Древесный запас в равной мере (~30%) распределен между здоровыми, ослабленными и сильно ослабленными. В лишайниковых редколесьях (рис. 1б) по числу преобладают ослабленные и сильно ослабленные особи (в сумме ~75%). Основной вклад в формирование запаса древостоя вносят ослабленные особи (40%), доли здоровых и сильно ослабленных составляют соответственно 30 и 25%. Особи крайних категорий состояния (здоровые и усыхающие) отличаются противоположным соотношением доли участия по числу и запасу. Это свидетельствует о том, что в древостоях средневозрастных северотаежных сосновых лесов продолжается интенсивный процесс дифференциации и отпада наиболее ослабленных особей. Аналогичные закономерности виталитетной структуры были выявлены и в 140-летних древостоях сосны кедровой в условиях южной тайги на территории Западной Сибири (Демидко и др., 2010).
Рис. 1.
Виталитетные спектры древостоев сосны обыкновенной в лишайниковых сосновых лесах (а) и редколесьях (б) Кольского п-ва с давностью пожара 85 лет. n и v – встречаемость различных категорий жизненного состояния, % общего числа особей и % запаса древесины соответственно. I–V – здоровые, ослабленные, сильно ослабленные, усыхающие и сухие соответственно.
В лишайниковых лесах у 70–80-летних деревьев независимо от уровня жизненного состояния 55 лет назад (через 30 лет после пожара) радиальный прирост не имел достоверных различий и составлял в среднем 1.5 мм/год. Через 30–85 лет после пожара у всех категорий особей происходило последовательное снижение радиального прироста, на фоне которого наблюдалось расслоение по скорости роста (рис. 2а). За указанный период радиальный прирост здоровых деревьев снизился в 1.8, сильно ослабленных и ослабленных – в 2.3–2.5, усыхающих – в 3.5 раза. Достоверные различия радиального прироста у особей, имеющих разный уровень жизненного состояния, начали проявляться через 50 лет после пожара. К концу анализируемого периода (через 85 лет после пожара) деревья всех категорий состояния имели достоверные (но не более чем двукратные) различия по значениям радиального прироста (рис. 2а).
Рис. 2.
Динамика радиального прироста деревьев сосны обыкновенной разных категорий состояния в процессе послепожарного восстановления лишайниковых сосновых лесов (а) и редколесий (б). I–IV – категории состояния: здоровые, ослабленные, сильно ослабленные, усыхающие соответственно; для рис. 2 и 3.
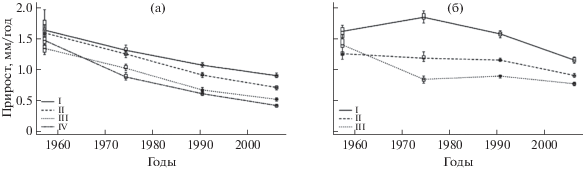
В южно-таежных кедровых лесах устойчивое соответствие между радиальным приростом и жизненным состоянием деревьев сосны кедровой сибирской было отмечено после 70 лет. За последние 30 лет в 140-летних древостоях скорость роста здоровых деревьев была выше, чем ослабленных и сильно ослабленных соответственно в 1.5 и 3 раза (Демидко и др., 2010).
Вариабельность радиального прироста деревьев сосны обыкновенной в лишайниковых лесах, оцененная с помощью коэффициента осцилляции (KD), в рассматриваемый период сукцессии у здоровых и ослабленных деревьев не изменялась, варьируя от 1.9 до 2.5, а у особей двух низших категорий виталитета она увеличилась от 2.5–3 до 3.7–5. При этом абсолютный диапазон значений радиального прироста у всех категорий особей уменьшился от 3.6–4 до 1.7–2.1 мм (табл. 2).
Таблица 2.
Абсолютный диапазон (D) и коэффициент осцилляции (KD) линейного радиального прироста особей сосны обыкновенной разных категорий состояния в лишайниковых сосновых лесах и редколесьях с разной давностью пожара
| Давность пожара, лет |
Категория состояния дерева | |||||||
|---|---|---|---|---|---|---|---|---|
| здоровые | ослабленные | сильно ослабленные | усыхающие | |||||
| D, мм | KD | D, мм | KD | D, мм | KD | D, мм | KD | |
| Лишайниковые сосновые леса | ||||||||
| 30 | 4.0 | 2.5 | 3.61 | 2.3 | 3.97 | 3.0 | 3.75 | 2.6 |
| 50 | 2.44 | 1.9 | 2.72 | 2.2 | 2.76 | 2.8 | 3.13 | 3.6 |
| 65 | 2.28 | 2.3 | 2.02 | 2.3 | 1.91 | 2.9 | 2.24 | 3.8 |
| 85 | 2.07 | 2.4 | 1.74 | 2.5 | 1.84 | 3.7 | 2.10 | 5.3 |
| Лишайниковые сосновые редколесья | ||||||||
| 30 | 4.19 | 2.7 | 3.85 | 3.2 | 8.94 | 6.7 | – | – |
| 50 | 4.68 | 2.6 | 3.96 | 3.5 | 3.00 | 3.8 | – | – |
| 65 | 3.13 | 2.1 | 3.21 | 2.9 | 2.95 | 3.5 | – | – |
| 85 | 2.89 | 2.6 | 2.60 | 3.0 | 2.32 | 3.2 | – | – |
Площадь годичного кольца 70–80-летних деревьев сосны обыкновенной в лишайниковых сосновых лесах через 30–85 лет после пожара имела принципиально иной характер динамики. При давности пожара 30 лет значения этого показателя у всех категорий особей достоверно не различались и составляли в среднем около ~1 см2/год. В период от 30 до 65 лет после пожара у здоровых, ослабленных и сильно ослабленных деревьев площадь годичного кольца увеличилась соответственно в 4, 1.7 и 1.4 раза, а у усыхающих уменьшилась в 2 раза, и в дальнейшем достоверно не изменялась (рис. 3а). Начиная с 50 лет после пожара, площадь годичного кольца у деревьев разных категорий состояния имела достоверные различия и к концу анализируемого периода, через 85 лет после пожара, соотношение площади годичного кольца здоровых, ослабленных, сильно ослабленных и усыхающих особей составило 8 : 5 : 2.5 : 1.
Рис. 3.
Динамика площади годичного кольца стволов сосны обыкновенной разных категорий состояния в процессе послепожарного восстановления лишайниковых лесов (а) и редколесий (б).
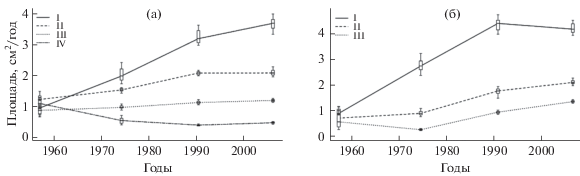
Диапазон значений площади годичного кольца у здоровых, ослабленных и сильно ослабленных особей сосны обыкновенной в период от 30 до 85 лет после пожара направленно не изменялся, варьируя в основном от ~10 до 14 см2/год (табл. 3). За этот же период значние коэффициента осцилляции площади годичного кольца уменьшилось у здоровых деревьев в 6, у ослабленных и сильно ослабленных – в 2–2.5 раза. У усыхающих особей коэффициент осцилляции площади годичного кольца в интервале 30–50 лет после пожара увеличился в 3.5–4 раза и в дальнейшем не изменялся, оставаясь на высоком уровне. При давности пожара 30 лет по значению коэффициента осцилляции выделялись только усыхающие особи, у которых оно было в среднем в 2 раза ниже, чем у здоровых, ослабленных и сильно ослабленных (табл. 3). На стадии средневозрастных древостоев (85 лет после пожара) различия этого показателя у разных виталитетных групп стали более отчетливыми. Наименьшая вариабельность площади годичного кольца (KD = 3) наблюдалась у здоровых деревьев, у ослабленных и сильно ослабленных она была примерно в 2, а у усыхающих – в 9 раз выше.
Таблица 3.
Абсолютный диапазон (D) и коэффициент осцилляции (KD) площади годичного кольца особей сосны обыкновенной разных категорий состояния в лишайниковых сосновых лесах и редколесьях с разной давностью пожара
| Давность пожара, лет |
Категория состояния дерева | |||||||
|---|---|---|---|---|---|---|---|---|
| здоровые | ослабленные | сильно ослабленные | усыхающие | |||||
| D, см2 | KD | D, см2 | KD | D, см2 | KD | D, см2 | KD | |
| Лишайниковые сосновые леса | ||||||||
| 30 | 14.15 | 15.6 | 13.68 | 11.5 | 12.21 | 14.8 | 7.41 | 7.0 |
| 50 | 10.97 | 5.6 | 12.70 | 8.5 | 11.26 | 12.2 | 13.13 | 26.7 |
| 65 | 10.95 | 3.4 | 14.33 | 7.0 | 10.90 | 10.1 | 9.28 | 27.2 |
| 85 | 9.43 | 2.6 | 11.23 | 5.5 | 6.92 | 6.0 | 10.07 | 23.9 |
| Лишайниковые сосновые редколесья | ||||||||
| 30 | 10.41 | 11.8 | 3.90 | 5.6 | 10.23 | 18.5 | – | – |
| 50 | 13.06 | 4.8 | 8.91 | 10.1 | 11.45 | 46.5 | – | – |
| 65 | 13.42 | 3.1 | 11.09 | 6.3 | 10.59 | 11.4 | – | – |
| 85 | 17.27 | 4.1 | 13.26 | 6.3 | 10.48 | 7.8 | – | – |
В лишайниковых сосновых редколесьях был проанализирован прирост деревьев сосны трех категорий состояния (здоровых, ослабленных и сильно ослабленных), поскольку доля усыхающих особей оказалась очень низкой. В начале анализируемого периода (через 30 лет после пожара) средний радиальный прирост деревьев разных категорий в сосновых редколесьях, также как в сосняках лишайниковых, практически не различался и составлял в среднем 1.4 мм/год. При увеличении давности пожара от 30 до 50 лет у здоровых особей значение прироста выросло до 1.8 мм/год, у ослабленных достоверно не изменилось (1.1 мм/год), у сильно ослабленных уменьшилось до 0.8 мм/год (рис. 2б). В конце указанного периода различия значений радиального прироста у особей разных категорий состояния были максимальными. В интервале от 50 до 85 лет после пожара радиальный прирост у здоровых и ослабленных особей снизился соответственно до 1.1 и 0.9 мм, а у сильно ослабленных оставался без изменения. В связи с этим к концу исследованного периода разброс значений приростов особей разных категорий состояния сократился: у крайних категорий он не превышает 1.5 раза.
Абсолютный диапазон значений радиального прироста у особей всех категорий состояния в лишайниковых редколесьях за изученный временной интервал снизился (табл. 2). У здоровых и ослабленных особей снижение было незначительным (примерно в 1.5 раза), а у сильно ослабленных – более существенным (в 4 раза). Коэффициент осцилляции радиального прироста у здоровых и ослабленных деревьев сосны в интервале 30–85 лет после пожара не изменялся, у сильно ослабленных уменьшился в 2 раза (табл. 2).
Площадь годичного кольца особей крайних категорий состояния в лишайниковых сосновых редколесьях в начале анализируемого периода при давности пожара 30 лет различалась не более чем в 1.5 раза: 0.88 и 0.55 см2/год у здоровых и сильно ослабленных деревьев соответственно (рис. 3б). В период от 30 до 65 лет после пожара у здоровых особей прирост по площади увеличился в 5 раз и затем достоверно не изменялся. У ослабленных особей значимый рост отмечался начиная с давности пожара 50 лет, и в целом за исследованный период площадь годичного кольца возросла примерно в 3 раза. У сильно ослабленных деревьев в интервале 30–50 лет наблюдалось снижение площади прироста, а в период от 50 до 85 лет после пожара он увеличился в 5.5 разa. Наибольшая дифференциация особей разных категорий состояния по площади годичного прироста отмечалась при давности пожара 65 лет. В конце исследуемого периода соотношение площадей годичных колец здоровых, ослабленных и сильно ослабленных особей по отношению к соответствующему значению у усыхающих особей в лишайниковых лесах составило соответственно 11 : 5.5 : 3.5.
Абсолютный диапазон значений площади годичного кольца в сосновых лишайниковых редколесьях у здоровых и ослабленных особей возрос за исследованный период соответственно в 1.7 и 3.5 раза, а у сильно ослабленных особей не изменился (табл. 3). Значения коэффициента осцилляции у всех категорий особей в первой половине исследованного периода (до 50 лет после пожара) были выше, чем во второй: у здоровых и ослабленных в 1.5–3, у сильно ослабленных в среднем в 4 раза.
Сравнение характера динамики линейного радиального прироста и площади годичного кольца у особей сосны обыкновенной при давности пожара 30–85 лет свидетельствует об их существенных различиях. В лишайниковых сосновых лесах линейный радиальный прирост за указанный временной интервал у особей всех категорий состояния снижался (в среднем в 2 раза). Аналогичное изменение в том же возрастном интервале было зарегистрировано в сосновых культурах на территории Брянской обл. (Марченко и др., 2014). Долговременная тенденция снижения радиального прироста деревьев, следующая за непродолжительным периодом его увеличения в первые годы жизни особей, отмечалась во многих исследованиях для разных лесообразующих видов бореальных лесов (Щекалев, Тарханов, 2007; Гончарова и др., 2012; Демаков, 2013; Полоскова и др., 2013; Демаков, Андреев, 2014; Николаева и др., 2015; Ярмишко и др., 2017).
В отличие от радиального прироста площадь годичного кольца у большей части особей (за исключением усыхающих) в изученных сообществах как минимум до 65 лет после пожара возрастала. Эта закономерность характерна как для лишайниковых сосновых лесов, так и для лишайниковых редколесий. Для северотаежных сосновых древостоев она отмечалась ранее (Ярмишко и др., 2017).
Дифференциация особей разных категорий состояния по значениям площади годичного кольца выражена значительно четче, чем по значениям линейного радиального прироста: в первом случае различие здоровых и сильно ослабленных особей 3-кратное, во втором – и 1.5–1.8-кратное. Следует отметить, что многие авторы (Антанайтис, Загреев 1969; Алексеев, Лайранд, 1993; Усольцев, 1997) указывали на более высокую объективность показателя площади годичного прироста для отражения динамики годичной продукции древесины.
Изученные типы северотаежных сообществ – лишайниковые сосновые леса и редколесья – заметно различаются по динамике площади годичного кольца особей сосны обыкновенной разных категорий жизненного состояния. У ослабленных и сильно ослабленных особей, составляющих по числу и запасу основу древостоев (60–75%), среднее значение площади годичного кольца в сосняках лишайниковых более высокое, чем в лишайниковых редколесьях. Максимальные различия (30–40%) наблюдаются при давности пожара 50 лет. Стабилизация площади годичного кольца в сосняках лишайниковых (65 лет после пожара) наступает раньше, чем в сосновых редколесьях, где до 85 лет после пожара продолжает увеличиваться прирост ослабленных и сильно ослабленных особей (рис. 2б). Однако к концу периода исследований средние значения площади годичного кольца у ослабленных (2.06 и 2.09 см2/год) и сильно ослабленных (1.15 и 1.34 см2/год) особей в двух типах сообществ достоверно не различались.
Характер динамики коэффициента осцилляции площади годичного кольца у ослабленных и сильно ослабленных особей в двух изученных типах сосновых сообществ одинаковый, во второй половине изученного временнóго интервала он снижается в 1.5–4 раза (табл. 3).
Динамика площади годичного кольца у здоровых особей в двух изученных типах сообществ также имеет ряд различий, которые начинают проявляться позже, чем у ослабленных и сильно ослабленных (с 50 лет после пожара и сохраняются до 85 лет после пожара). Через 50–65 лет после пожара площадь годичного кольца здоровых деревьев сосны в сосняках лишайниковых на 30% ниже, чем в лишайниковых редколесьях. Однако к концу исследованного периода (85 лет после пожара) различия, оставаясь статистически достоверными, сокращаются до 10%. Вариабельность площади годичного кольца у здоровых особей сопоставима в двух изученных типах сообществ и наиболее высокая до 30 лет после пожара (KD= 12–16), к 50 годам после пожара она снижается в 2–3 раза, а к 65 годам – в 4 раза и в дальнейшем стабилизируется.
Результаты выполненных сравнений позволяют сделать следующие основные заключения: до 50 лет после пожара, т.е. в период до восстановления основных средостабилизирующих функций лишайникового покрова, условия для роста подавляющего большинства особей (ослабленных и сильно ослабленных) в сосновых лишайниковых редколесьях менее благоприятные, чем в лишайниковых сосновых лесах. Это обусловлено тем, что почвы сосновых редколесий отличаются более низкой влагоудерживающей способностью в связи с повышенной долей фракции крупного песка (Ставрова и др., 2015).
Главные характеристики, определяющие стабилизацию режима влажности верхних горизонтов почв, – плотность и высота мохово-лишайникового покрова и толщина подстилки (Siren, 1955; Kershaw, 1975; Санников, 1992). Согласно имеющимся данным (Горшков и др., 2005) восстановление этих параметров в северотаежных лишайниковых сосновых лесах завершается в течение 70–100 лет после пожара. В результате увеличения высоты и плотности лишайникового покрова, восстановления толщины подстилки и стабилизации водного режима почв через 85 лет после пожара, площадь годичного прироста большинства деревьев сосны в лишайниковых лесах и редколесьях становится практически одинаковой.
Относительно небольшое число особей в сосновых редколесьях, которые на момент исследования относились к категории здоровых, по-видимому, развивается в микроместообитаниях, где различия гранулометрического состава не имеют решающего значения. Это могут быть, в частности, участки с недогоревшей подстилкой, на которых рост особей сосны в сосновых лесах и редколесьях может быть одинаковым или даже более успешным на отдельных этапах в условиях редколесий.
В целом можно заключить, что при давности пожара >80 лет различие в продуктивности местообитаний северотаежных сосновых лишайниковых лесов и редколесий проявляется на уровне сообществ (в плотности и сумме площадей сечений древостоев), но не проявляется на уровне отдельных особей.
На основе таких показателей, как средняя плотность особей на единицу площади, соотношение числа особей разных категорий состояния и среднее значение площади годичного кольца особей каждой категории, может быть оценен ресурсный потенциал древостоев (суммарная площадь годичного прироста всех деревьев). Установлено, что общий ресурсный потенциал лишайниковых сосновых лесов в 1.6 раза превышает ресурсный потенциал редколесий (табл. 4 ). Различие формируется в основном за счет здоровых и сильно ослабленных особей, ресурсный потенциал которых в лишайниковых лесах соответственно в 2.2 и 1.6 раза выше, чем в редколесьях. Ресурсный потенциал ослабленных деревьев в двух рассматриваемых типах сообществ практически одинаков.
На долю здоровых особей в лишайниковых сосновых лесах и редколесьях приходится соответственно 40 и 30% общего ресурсного потенциала, на долю ослабленных – 28 и 43%, сильно ослабленных – 27 и 26% (табл. 4).
Таблица 4.
Оценка ресурсного потенциала деревьев разных категорий состояния в лишайниковых сосновых лесах и редколесьях
| Категории состояния | Число стволов | Средний годичный прирост по площади, см2/год* | Суммарный годичный прирост, см2/год | Доля в общем годичном приросте, % |
|---|---|---|---|---|
| Лишайниковые сосновые леса | ||||
| Здоровые | 222 | 3.69 | 819 | 40.7 |
| Ослабленные | 270 | 2.06 | 556 | 27.6 |
| Сильно ослабленные | 468 | 1.15 | 538 | 26.7 |
| Усыхающие | 240 | 0.42 | 101 | 5.0 |
| Всего | 1200 | – | 2014 | 100 |
| Лишайниковые сосновые редколесья | ||||
| Здоровые | 91 | 4.16 | 379 | 30.3 |
| Ослабленные | 260 | 2.09 | 543 | 43.3 |
| Сильно ослабленные | 247 | 1.34 | 331 | 26.4 |
| Усыхающие | 52 | – | – | – |
| Всего | 650 | – | 1253 | 100 |
К наиболее продуктивным категориям деревьев относятся здоровые и умеренно ослабленные. В редколесьях здоровые деревья на фоне существенно более низких значений плотности (до 2–2.5 раза) при сходной средней площади годичного прироста имеют в 2 раза более низкий ресурсный потенциал по сравнению с потенциалом здоровых деревьев в лишайниковых лесах (табл. 5). Умеренно ослабленные особи в лишайниковых сосновых лесах и редколесьях имеют одинаковую плотность, одинаковую площадь годичного кольца и соответственно примерно равный ресурсный потенциал (табл. 5). Сильно ослабленные особи при сходном приросте по площади, но примерно в 2 раза более низкой плотности, как и здоровые деревья, отличаются существенно более низким ресурсным потенциалом в редколесьях по сравнению сосновыми лесами (табл. 5).
Таблица 5.
Соотношение числа, запаса древесины и прироста особей сосны обыкновенной разных категорий жизненного состояния в лишайниковых сосновых лесах и редколесьях с давностью пожара 85 лет
| Параметр | Редколесья/леса |
|---|---|
| Общая плотность | 0.54 |
| Здоровые особи | |
| Плотность | 0.41 |
| Доля по запасу | 0.91 |
| Радиальный прирост | 1.25 |
| Площадь годичного кольца | 1.13 |
| Ресурсный потенциал | 0.46 |
| Ослабленные особи | |
| Плотность | 0.96 |
| Доля по запасу | 1.33 |
| Радиальный прирост | 1.25 |
| Площадь годичного кольца | 1 |
| Ресурсный потенциал | 0.98 |
| Сильно ослабленные особи | |
| Плотность | 0.53 |
| Доля по запасу | 0.81 |
| Радиальный прирост | 1.46 |
| Площадь годичного кольца | 1.17 |
| Ресурсный потенциал | 0.62 |
ЗАКЛЮЧЕНИЕ
Выполненное исследование показало, что древостои постпирогенных средневозрастных северотаежных лишайниковых сосновых лесов и редколесий на 60–75% состоят из ослабленных и сильно ослабленных особей, на долю здоровых деревьев приходится не более 20%. Различия в скорости роста ствола у деревьев разных категорий состояния начинают достоверно проявляться с 50 лет после пожара. В период от 30 до 85 лет у всех особей в составе древостоев наблюдается снижение радиального прироста, более резко выраженное у сильно ослабленных и усыхающих (в 2.5–3.5 раза) по сравнению со здоровыми и умеренно ослабленными (в 1.5–2 раза). В отличие от радиального прироста величина прироста по площади годичного кольца в указанном возрастном интервале возрастает: более существенно (4–5-кратно) у здоровых деревьев, в меньшей степени (в среднем 2-кратно) – у сильно ослабленных. Наблюдаемые в период от 65 до 85 лет замедление или прекращение увеличения площади годичного кольца и снижение вариабельности годичных приростов отражают, по-видимому, с одной стороны переход к выраженному конкурентному взаимодействию особей в составе сосновых древостоев, с другой – восстановление средостабилизирующих функций сообществ. Различие в продуктивности местообитаний лишайниковых сосновых лесов и редколесий проявляется на уровне сообществ (в плотности и сумме площадей сечений древостоев), но не проявляется на уровне отдельных особей.
Работа выполнена при финансовой поддержке программы Президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России”, а также с привлечением ресурсов Ресурсного центра “Обсерватория экологической безопасности” Научного парка Санкт-Петербургского государственного университета.
Список литературы
Алексеев А.С. Анализ экологической структуры популяций Picea abies (Pinaceae) в условиях атмосферного загрязнения // Ботан. журн. 1990. Т. 75. № 9. С. 1277–1284.
Алексеев А.С., Лайранд Н.И. К методике дендроэкологического анализа // Ботан. журн. 1993. Т. 78. № 10. С. 103–107.
Алексеев А.С., Сорока А.Р. Анализ долговременных тенденций роста Pinus sylvestris на северо-западе Кольского полуострова // Ботан. журн. 2003. Т. 88. № 6. С. 59.
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Антанайтис В.В., Загреев В.В. Прирост леса. М.: Лес. пром-сть, 1969. 240 с.
Бебия С.М. Дифференциация деревьев в лесу, их классификация и определение жизненного состояния древостоев // Лесоведение. 2000. № 4. С. 35–43.
Битвинскас Т.Т. Дендроклиматические исследования. Л.: Гидрометеоиздат, 1974. 170 с.
Бузыкин А.И., Дашковская И.С., Черкашин В.П. Динамика радиального прироста хвойных разного ценотического положения в Приангарье // Дендрохронология и дендроклиматология. Новосибирск: Наука, 1986. С. 79–86.
Буяк А.В., Карпов В.Г. Сравнительный анализ динамики радиального прироста ели // Факторы регуляции экосистем еловых лесов. Л.: Наука, 1983. С. 65–78.
Ваганов Е.А., Шашкин А.В. Рост и структура годичных колец хвойных. Новосибирск: Наука, 2000. 232 с.
Ваганов Е.А., Шиятов С.Г., Мазепа В.С. Дендроклиматические исследования в Урало-Сибирской субарктике. Новосибирск: Наука, 1996. 246 с.
Волков Ю.В., Тартаковский В.А. Алгоритм анализа площадей годичных слоев деревьев. // Изв. Томск. политехн. ун-та. 2011. № 319(5). С. 112–117.
Гончарова О.А., Кузьмин А.В., Полоскова Е.Ю. Динамика годичного радиального роста старовозрастных деревьев сосны обыкновенной на Кольском полуострове (пос. Умба) // Теор. и прикл. экология. 2012. № 2. С. 118–122.
Горшков В.В., Катютин П.Н. Внутрипопуляционная дифференциация Picea obovata Ledeb. по радиальному приросту // Динамика лесных сообществ Северо-Запада России. СПб.: ВВМ, 2009. С. 185–196.
Горшков В.В., Ставрова Н.И., Баккал И.Ю. Динамика восстановления лесной подстилки в бореальных сосновых лесах после пожаров // Лесоведение. 2005. № 3. С. 37–45.
Горшков В.В., Ставрова Н.И., Катютин П.Н., Тумакова Е.А. Типы размерной и виталитетной структуры ценопопуляций Pinus sylvestris (Pinaceae) в условиях северной тайги (Кольский п-ов) // Растит. ресурсы. 2013. Т. 49. № 4. С. 512–531.
Демаков Ю.П. Динамика радиального прироста деревьев в сосняках лишайниково-мшистых заповедника “Большая Кокшага” // Науч. труды гос. Природ. заповедника “Большая Кокшага”. 2013. № 6. С. 143–162.
Демаков Ю.П., Андреев Н.В. Закономерности радиального прироста деревьев сосны в приозерных биотопах национального парка “Марий Чодра” // Экопотенциал. 2014. № 3(7). С. 48–58.
Демидко Д.А. Виталитетная структура ненарушенных древостоев кедра сибирского в субальпийском подпоясе и на верхней границе леса в горном Алтае // Экология. 2006. № 5. С. 394–397.
Демидко Д.А., Кривец С.А., Бисирова Э.М. Связь радиального прироста и жизненного состояния у деревьев кедра сибирского // Вестн. Томск. гос. ун-та. Биология. 2010. № 4. С. 68–80.
Крамер П.Д., Козловский Т.Т. Физиология древесных растений. М.: Лec. Промышленность, 1983. 464 с.
Кузьмин А.В., Полоскова Е.Ю., Гончарова О.А. Определение временных интервалов влияния температурных факторов на формирование радиального прироста в сосновых древостоях Кольского региона // Вестн. Мурманск. гос. техн. ун-та. 2011. Т. 14. № 3. С. 565–576.
Лянгузов А.Ю., Ярмишко В.Т., Лянгузова И.В. Новый метод оценки годичного прироста стволов древесных растений // Растит. ресурсы. 2017. № 4. С. 580–593.
Марченко С.И., Умнов Е.Г., Панина Н.Б. Особенности роста по диаметру рядовых и групповых культур сосны обыкновенной // Актуальные проблемы лес. комплекса. 2014. № 38. С. 34–38.
Махныкина А.В., Верховец С.В., Кошурникова Н.Н. Воздействие нарушающих факторов различного происхождения на радиальный прирост в сосновых насаждениях центральной Сибири // Изв. РАН. Сер. биол. 2017. № 4. С. 476–482.
Методы изучения лесных сообществ. СПб.: НИИхимии СПбГУ, 2002. 240 с.
Моисеев П.А. Влияние изменений климата на радиальный прирост и формирование возрастной структуры высокогорных лиственничников Кузнецкого Алатау // Экология. 2002. № 1. С. 10–17.
Николаева С.А., Савчук Д.А., Бочаров А.Ю. Влияние различных факторов на прирост деревьев Pinus sibirica в высокогорных лесах центрального Алтая // Журн. Сиб. фед. ун-та. Сер.: Биология. 2015. Т. 8. № 3. С. 299–318.
Никонов В.В. Почвообразование на северном пределе сосновых биогеоценозов. Л.: Наука, 1987. 142 с.
Переверзев В.Н. Лесные почвы Кольского полуострова. М.: Наука, 2004. 232 с.
Полоскова Е.Ю., Гончарова О.А., Кузьмин А.В. Возрастные особенности и динамика радиального роста сосны обыкновенной на Кольском полуострове // Вестн. МГТУ. 2013. Т. 16. № 3. С. 519–525.
Санитарные правила в лесах Российской Федерации. 2006 // Интернет-портал лесной отрасли wood.ru / www.wood.ru.
Санников С.Н. Экология и география естественного возобновления сосны обыкновенной. М.: Наука,1992. 263 с.
Ставрова Н.И., Горшков В.В., Катютин П.Н. Динамика виталитетной структуры ценопопуляций Picea obovata и Betula pubescens в процессе послепожарных сукцессий северотаежных еловых лесов // Ботан. журн. 2010. Т. 95. № 11. С. 1550–1565.
Ставрова Н.И., Горшков В.В., Баккал И.Ю., Катаева М.Н. Типологическое разнообразие сосновых лесов и его связь с гранулометрическим составом почв в условиях северной тайги // Материалы III международной научно-практической конференции, посвященной 110-летию со дня рождения академика Н.В. Смольского. Минск: ЗАО “Кондифо”, 2015. С. 196–200.
Торлопова Н.В., Ильчуков С.В. Жизненное состояние коренных сосняков Печоро-Илычского биосферного заповедника // Лесоведение. 2003. № 3. С. 34–40.
Усольцев В.А. Биологические аспекты таксации фитомассы деревьев. Екатеринбург: УрО РАН, 1997. 216 с.
Черненькова Т.В., Бочкарев Ю.Н., Фридрих М., Беттгер Т. Воздействие природно-антропогенных факторов на радиальный прирост деревьев Кольского Севера // Лесоведение. 2012. № 4. С. 3–15.
Шашкин Е.А., Ваганов Е.А. Динамика прироста площадей сечения стволов у деревьев в разных районах Сибири в связи с глобальными изменениями температуры // Лесоведение. 2000. № 3. С. 3–11.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Шиятов С. Г. Дендрохронология верхней границы леса на Урале. М.: Наука, 1986. 136 с.
Щекалев Р.В., Тарханов С.Н. Радиальный прирост сосны обыкновенной при аэротехногенном загрязнении в бассейне Северной Двины // Лесоведение. 2007. № 2. С. 45–50.
Ярмишко В.Т. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере. СПб.: Изд-во НИИхимии СПбГУ, 1997. 210 с.
Ярмишко В.Т., Ярмишко М.А. Радиальный прирост Pinus sylvestris (Pinaceae) на северном пределе распространения // Ботан. журн. 2004. Т. 89. № 7. С. 1992–1110.
Ярмишко В.Т., Горшков В.В., Ставрова Н.И. Виталитетная структура Pinus sylvestris L. в лесных сообществах с разной степенью и типом антропогенной нарушенности (Кольский полуостров) // Растит. ресурсы. 2003. Т. 39. Вып. 4. С. 1–19.
Ярмишко В.Т., Лянгузова И.В., Лянгузов А.Ю. Изменение годичного прироста стволов Pinus sylvestris (Pinaceae) при снижении аэротехногенного загрязнения // Растит. ресурсы. 2017. № 4. С. 527–542.
Bigler C., Bugmann H. Growth-dependent tree mortality models based on tree rings // Can. J. For. Res. 2003. V. 33(2). P. 210–221.
Björkdahl G., Eriksson H. Effects of crown decline on increment in Norway spruce (Picea abies (L.) Karst) in Southern Sweden / Eds Brække H., Bjor K., Halvorsen B. Air pollution as stress factor in the Nordic forests // Comm. Norw. For. Res. 1989. V. 42. P. 19–36.
Briffa K.R., Jones P.D., Schweingruber F.H. et al. Tree-ring variables as proxy-climate indicator: Problems wiht low-frecuency signals // Eds Jones P.D., Bradley R.S., Jouzel J. Climatic variations and mechanisms of the last 2000 years. Berlin; Heidelberg: Springer-Verlag, 1996. P. 9–41.
Danek M. The influence of industry on Scots pine stands in the South-eastern Part of the Silesia-Krakow Upland (Poland) on the basis of dendrochronological analysis // Water, Air, Soil Pollut. 2007. V. 185. P. 265.
Das A. The effect of size and competition on tree growth rate in old-growth coniferous forests // Can. J. For. Res . 2012. V. 42(11). P. 1983–1995.
Dobbertin M. Forest growth as indicator of tree vitality and tree reaction to environmental stress: a review // Eur. J. For. Res. 2005. V. 124(4). P. 319–333.
Fritts H.C. Tree rings and climate. L.: Acad. Press., 1976. 567 p.
Juknys R., Augustaitis A., Vencloviene J., Kliučius A., Vitas A., Bartkevičius E. Jurkonis N. Dynamic response of tree growth to changing environmental pollution // Eur. J. For. Res. 2014. V. 133(4). P. 713–724.
Kershaw K.A. Studies of lichen-dominated systems. XV. The temperature and humidity profiles in a Cladina alpestris mat // Can. J. Bot. 1975. V. 53(22). P. 2614–2620.
Methods of Dendrochronology. Applications in environmental sciences // Eds Cook E., Kairiukstis L. Dordrecht: Kluwer Acad. Press., 1990. 394 p.
Monserud R.A., Sterba H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria // For. Ecol. Manag. 1996. V. 80. P. 57–80.
Nilsson, U., Gemmel P. Changes in growth and allocation of growth in young Pinus sylvestris and Picea abies due to competition // Scand. J. For. Res. 1993. V. 8. P. 213–222.
Pärn H. Radial growth response of Scots pine to climate under dust pollution in Northeast Estonia // Water, Air, Soil Pollut. 2003. V. 144. P. 343–355.
Siren G. The development of spruce forest on raw humus sites in northern Finland and its ecology // Acta For. Fenn. 1955. V. 62. P. 1–363.
Solberg S. Crown condition and growth relationships within stands of Picea abies // Scand. J. For. Res. 1999. V. 14. P. 320–327.
Schweingruber F.H. Tree-Rings and environment. Dendroecology. Berne; Stuttgart; Vienna: Swiss Fed. Inst. For. Snowand Landscape Res., 1996. 609 p.
Vacek S., Vacek Z., Remeš J., Bílek L., Hůnová I., Bulušek D., Putalová T., Král J., Simon J. Sensitivity of unmanaged relict pine forest in the Czech Republic to climate change and air pollution // Trees. 2017. V. 31. P. 1599–1617.
Vaganov E.A., Hughes M.K., Shashkin A.V. Growth dynamics of conifer tree rings. Images of past and future environments. Ecological Studies. Berlin; Heidelberg; New York: Springer Verlag. 2006. V. 183. 354 p.
Wilczyński S. The variation of tree-ring widths of Scots pine (Pinus sylvestris L.) affected by air pollution // Eur. J. For. Res. 2006. V.125(3). P. 213–219.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая