

Известия РАН. Серия биологическая, 2021, № 2, стр. 211-224
К экологии размножения рыжей полевки Myodes (Clethrionomys) glareolus Schreb. на северной периферии ареала. Сообщение 2. Половое созревание, плодовитость взрослых и молодых полевок
1 Петрозаводский государственный университет
185910 Петрозаводск, просп. Ленина, 33, Россия
2 Карельский научный центр РАН
185000 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 01.02.2019
После доработки 01.11.2019
Принята к публикации 01.11.2019
Аннотация
В итоге многолетних исследований (1958–2017) определено, что в условиях северной периферии ареала скорость полового созревания сеголеток рыжей полевки зависит не столько от абсолютного возраста, сколько от календарного срока рождения. Отмечено, что отдельные особи могут размножаться уже через 25–30 сут после рождения, при этом чем раньше родились зверьки, тем быстрее они растут и развиваются и тем раньше созревают и вступают в размножение. Установлено, что самцы и самки, родившиеся позднее июня, в массе не созревают в год рождения и размножаются только следующей весной, после зимовки, и хотя среди них встречаются половозрелые особи (в основном июльских выводков), число их не превышает 21%. Обнаружено, что быстро развивающиеся сеголетки ранних выводков, почти поголовно участвующие в размножении в год рождения, обеспечивают рост населения к осени текущего года, а родившиеся во второй половине лета служат резервом популяции для следующего сезона размножения. Зафиксированы возрастная разнокачественность популяции и четкое “распределение обязанностей” между отдельными генерациями, которые достаточно лабильны и служат одним из важнейших механизмов регуляции численности. Отмечено, что плодовитость взрослых и размножающихся прибылых самок зависит не только от экологических условий, но и от их численности весной.
В данном сообщении – продолжении предыдущего – рассматриваются не затронутые ранее аспекты проблемы репродукции рыжей полевки (возраст и условия полового созревания зверьков, плодовитость и ее зависимость от возраста и времени рождения зверьков, участие в размножении прибылых особей и связи всего процесса воспроизводства популяции с эндогенными и экзогенными факторами). При этом характеристика материала, а также методы его получения и анализа достаточно подробно описаны в предыдущей статье и не требуют дублирования.
УЧАСТИЕ В РАЗМНОЖЕНИИ ПРИБЫЛЫХ ПОЛЕВОК
Согласно полученным нами данным в условиях Восточной Фенноскандии, служащей для изучаемого вида северной периферией ареала, отдельные его особи, причем как самцы, так и самки, могут размножаться уже через 25–30 сут после рождения. Их размеры и масса составляют 73–84 мм и 14–18.5 г соответственно, но состояние гонад и придаточных желез свидетельствует о достижении половой зрелости и участии в размножении. У самцов увеличены семенники (длина 9–11 мм, масса 450–650 мг), хорошо развиты придатки и семенные пузырьки (7–11.6 мм). В извитых канальцах появляются все клеточные элементы (от клеток Сертоли до зрелых сперматозоидов), которые обнаруживаются и в придатках семенника. Яичники самок демонстрируют полную картину овогенеза. Характерно наличие большого числа близких к разрыву граафовых пузырьков диаметром 300–500 мкм. В яичниках беременных самок образуются желтые тела. Масса яичников достигает 18–25 мг и более. Матка гиперемирована, стенки ее утолщены и имеют складчатое строение; длина рогов 18–26 мм. Минимальная масса активных в половом отношении сеголеток была равна 12–14 г при длине тела 73–81.6 мм. Это близко к ранее опубликованным данным (Наумов, 1948; Попов, 1960; и др.), согласно которым возраст подавляющего большинства исследованных ими половозрелых сеголеток 45–50 сут.
ВОЗРАСТНОЙ ПОЛИМОРФИЗМ РЕПРОДУКТИВНОЙ ЧАСТИ ПОПУЛЯЦИИ
В условиях таежного северо-запада России, а также в других частях ареала скорость полового созревания сеголеток зависит не столько от абсолютного возраста, сколько от календарного срока рождения. Чем раньше родились зверьки, тем быстрее проходит их рост и развитие и тем раньше они созревают и вступают в размножение. Это в равной мере относится и к самцам, и к самкам и находит косвенное подтверждение в изменении начальной массы размножающихся особей разного времени рождения. Так, полевки майских выводков начинают размножаться уже с массой 12–14, июньского – 16–19, июльского – 20–21 г. Параллельно этому изменяется и доля участия прибылых особей в размножении. Среди молодых самок майских выводков, достигших двухмесячного возраста, размножалось в среднем за все годы 88.3% (с колебаниями по годам от 45 до 100%), из июньских – 76% (29–83), июльских – 6.8% (0–10.5). Интенсивность полового созревания (сперматогенез) прибылых самцов ранних майско-июньских выводков отражена в табл. 1. У большинства из них начиная с середины июня наблюдается активный сперматогенез. Во второй декаде он отмечен у 47% самцов, в третьей – у 86%. В первой половине июля численность самцов с активным сперматогенезом несколько снижается за счет появления в уловах полевок июньских выводков, не достигших еще половозрелости, но в конце июля и в августе их численность снова увеличивается и достигает 89–100%. Начиная с середины сентября у самцов ранних выводков наступают затухание сперматогенеза и дегенерация зародышевого эпителия до однорядного слоя сперматогониев и клеток Сертоли. Семенники становятся дряблыми и приобретают темную коричневатую окраску, их масса составляет 20–90 мг; сокращаются в размерах и семенные пузырьки. Самцы, родившиеся позднее июня, в основном не созревают в год рождения и размножаются только следующей весной, после зимовки (табл. 2), и хотя среди них встречаются половозрелые особи (в основном июльских выводков), число их не превышает 21%.
Таблица 1.
Состояние половой системы и сперматогенез у прибылых самцов рыжей полевки
| Месяц | Декада | Общее число исследованных самцов | Масса тела, г | Длина, мм | Масса двух семенников, мг | Доля самцов со сперматогенезом, % от всех исследованных | |
|---|---|---|---|---|---|---|---|
| семенника | семенных пузырьков | ||||||
| Прибылые ранних выводков | |||||||
| Июнь | I | 17 | $\frac{{6.5{\kern 1pt} - {\kern 1pt} 20.2}}{{14.9}}$ | $\frac{{1.5{\kern 1pt} - {\kern 1pt} 11.5}}{{8.6}}$ | $\frac{{1.5{\kern 1pt} - {\kern 1pt} 10.0}}{{7.2}}$ | $\frac{{75.0{\kern 1pt} - {\kern 1pt} 124.0}}{{113.7}}$ | 0 |
| II | 134 | $\frac{{4.8{\kern 1pt} - {\kern 1pt} 28.3}}{{16.2}}$ | $\frac{{1.5{\kern 1pt} - {\kern 1pt} 12.6}}{{7.5}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 12.7}}{{7.3}}$ | $\frac{{30.0{\kern 1pt} - {\kern 1pt} 980.0}}{{372.1}}$ | 48.1 | |
| III | 85 | $\frac{{8.5{\kern 1pt} - {\kern 1pt} 23.8}}{{17.9}}$ | $\frac{{3.2{\kern 1pt} - {\kern 1pt} 13.7}}{{10.1}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 12.5}}{{7.8}}$ | $\frac{{150.0{\kern 1pt} - {\kern 1pt} 900.0}}{{501.1}}$ | 84.3 | |
| Июль | I | 155 | $\frac{{8.5{\kern 1pt} - {\kern 1pt} 34.1}}{{17.7}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 14.0}}{{9.5}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 13.6}}{{8.3}}$ | $\frac{{12.0{\kern 1pt} - {\kern 1pt} 1300.0}}{{504.1}}$ | 72.5 |
| II | 298 | $\frac{{7.9{\kern 1pt} - {\kern 1pt} 35.1}}{{17.6}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 17.5}}{{9.3}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 14.5}}{{8.2}}$ | $\frac{{2.5{\kern 1pt} - {\kern 1pt} 1100.0}}{{392.5}}$ | 48.9 | |
| III | 249 | $\frac{{4.5{\kern 1pt} - {\kern 1pt} 42.1}}{{17.5}}$ | $\frac{{1.8{\kern 1pt} - {\kern 1pt} 17.5}}{{8.2}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 17.3}}{{9.4}}$ | $\frac{{6.0{\kern 1pt} - {\kern 1pt} 1100.0}}{{424.2}}$ | 79.1 | |
| Август | I | 158 | $\frac{{10.8{\kern 1pt} - {\kern 1pt} 27.3}}{{17.9}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 13.7}}{{8.1}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.3}}{{9.9}}$ | $\frac{{40.0{\kern 1pt} - {\kern 1pt} 1550.0}}{{591.3}}$ | 87.9 |
| II | 123 | $\frac{{7.9{\kern 1pt} - {\kern 1pt} 26.6}}{{17.9}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 14.7}}{{8.0}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.8}}{{9.1}}$ | $\frac{{27.0{\kern 1pt} - {\kern 1pt} 1040.0}}{{474.7}}$ | 88.1 | |
| III | 151 | $\frac{{10.8{\kern 1pt} - {\kern 1pt} 23.4}}{{18.3}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 14.7}}{{5.9}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 15.8}}{{7.9}}$ | $\frac{{200.0{\kern 1pt} - {\kern 1pt} 1040.0}}{{545.3}}$ | 100 | |
| Сентябрь | I | 19 | $\frac{{10.8{\kern 1pt} - {\kern 1pt} 22.9}}{{16.6}}$ | $\frac{{3.0{\kern 1pt} - {\kern 1pt} 7.8}}{{5.8}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 10.8}}{{7.3}}$ | – | 39.8 |
| II | 11 | $\frac{{9.5{\kern 1pt} - {\kern 1pt} 25.6}}{{19.4}}$ | $\frac{{2.5{\kern 1pt} - {\kern 1pt} 7.0}}{{4.2}}$ | – | $\frac{{38.0{\kern 1pt} - {\kern 1pt} 120.0}}{{79.0}}$ | 0 | |
| III | 17 | $\frac{{14.9{\kern 1pt} - {\kern 1pt} 26.5}}{{20.9}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 10.2}}{{5.5}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 10.2}}{{4.3}}$ | $\frac{{47.9{\kern 1pt} - {\kern 1pt} 518.0}}{{93.6}}$ | 10.6 | |
| Oктябрь | I | 25 | $\frac{{9.2{\kern 1pt} - {\kern 1pt} 24.0}}{{15.7}}$ | $\frac{{1.2{\kern 1pt} - {\kern 1pt} 5.5}}{{2.8}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 4.0}}{{2.1}}$ | $\frac{{25.0{\kern 1pt} - {\kern 1pt} 93.0}}{{39.5}}$ | 0 |
| II | 33 | $\frac{{14.5{\kern 1pt} - {\kern 1pt} 18.9}}{{16.5}}$ | $\frac{{1.5{\kern 1pt} - {\kern 1pt} 4.2}}{{2.8}}$ | – | – | 0 | |
| III | 47 | $\frac{{12.2{\kern 1pt} - {\kern 1pt} 17.4}}{{14.6}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 4.1}}{{2.6}}$ | – | $\frac{{12.0{\kern 1pt} - {\kern 1pt} 25.0}}{{15.3}}$ | 0 | |
| Прибылые поздних выводков | |||||||
| Июль | I | 71 | $\frac{{8.5{\kern 1pt} - {\kern 1pt} 34.1}}{{16.7}}$ | $\frac{{3.0{\kern 1pt} - {\kern 1pt} 12.7}}{{8.6}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 13.3}}{{7.8}}$ | $\frac{{15.0{\kern 1pt} - {\kern 1pt} 1200.0}}{{403.6}}$ | 0 |
| II | 135 | $\frac{{7.9{\kern 1pt} - {\kern 1pt} 35.1}}{{16.8}}$ | $\frac{{2.9{\kern 1pt} - {\kern 1pt} 14.1}}{{7.7}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 12.3}}{{7.2}}$ | $\frac{{7.5{\kern 1pt} - {\kern 1pt} 904.0}}{{272.6}}$ | 6.1 | |
| III | 114 | $\frac{{4.5{\kern 1pt} - {\kern 1pt} 26.5}}{{15.2}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 12.4}}{{6.9}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 14.5}}{{8.1}}$ | $\frac{{8.0{\kern 1pt} - {\kern 1pt} 980.0}}{{316.0}}$ | 18.3 | |
| Август | I | 218 | $\frac{{8.0{\kern 1pt} - {\kern 1pt} 25.7}}{{16.9}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 13.2}}{{7.2}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.3}}{{8.8}}$ | $\frac{{20.0{\kern 1pt} - {\kern 1pt} 952.0}}{{469.1}}$ | 22.5 |
| II | 185 | $\frac{{7.9{\kern 1pt} - {\kern 1pt} 26.6}}{{16.8}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 13.0}}{{6.9}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.8}}{{8.2}}$ | $\frac{{15.0{\kern 1pt} - {\kern 1pt} 1040.0}}{{337.9}}$ | 14.5 | |
| III | 255 | $\frac{{1.6{\kern 1pt} - {\kern 1pt} 175.0}}{{16.8}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 12.9}}{{4.6}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.4}}{{7.0}}$ | $\frac{{14.0{\kern 1pt} - {\kern 1pt} 800.0}}{{157.0}}$ | 10.1 | |
| Сентябрь | I | 131 | $\frac{{7.3{\kern 1pt} - {\kern 1pt} 24.5}}{{15.9}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 13.0}}{{3.8}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 15.0}}{{5.0}}$ | $\frac{{20.0{\kern 1pt} - {\kern 1pt} 1000.0}}{{221.5}}$ | 0 |
| II | 84 | $\frac{{8.8{\kern 1pt} - {\kern 1pt} 22.6}}{{16.4}}$ | $\frac{{1.5{\kern 1pt} - {\kern 1pt} 10.0}}{{3.4}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 7.8}}{{3.7}}$ | $\frac{{17.5{\kern 1pt} - {\kern 1pt} 500.0}}{{124.3}}$ | 0 | |
| III | 89 | $\frac{{13.5{\kern 1pt} - {\kern 1pt} 23.5}}{{17.1}}$ | $\frac{{1.0{\kern 1pt} - {\kern 1pt} 9.3}}{{3.0}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 8.7}}{{0.9}}$ | $\frac{{9.0{\kern 1pt} - {\kern 1pt} 470.0}}{{65.2}}$ | 2.5 | |
| Октябрь | I | 40 | $\frac{{11.3{\kern 1pt} - {\kern 1pt} 20.5}}{{17.2}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 5.0}}{{3.1}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 4.7}}{{2.9}}$ | $\frac{{4.0{\kern 1pt} - {\kern 1pt} 30.0}}{{12.0}}$ | 0 |
| II | 31 | $\frac{{12.7{\kern 1pt} - {\kern 1pt} 25.0}}{{16.2}}$ | $\frac{{2.1{\kern 1pt} - {\kern 1pt} 3.1}}{{2.6}}$ | $\frac{{0{\kern 1pt} - {\kern 1pt} 4.7}}{{0.9}}$ | $\frac{{6.0{\kern 1pt} - {\kern 1pt} 20.0}}{{11.1}}$ | 0 | |
| III | 26 | $\frac{{12.0{\kern 1pt} - {\kern 1pt} 24.0}}{{16.5}}$ | $\frac{{2.0{\kern 1pt} - {\kern 1pt} 4.3}}{{2.9}}$ | – | $\frac{{10.0{\kern 1pt} - {\kern 1pt} 25.0}}{{14.7}}$ | 0 | |
| Ноябрь | I–III | 17 | $\frac{{9.4{\kern 1pt} - {\kern 1pt} 19.1}}{{14.1}}$ | $\frac{{2.7{\kern 1pt} - {\kern 1pt} 9.0}}{{4.8}}$ | – | $\frac{{8.0{\kern 1pt} - {\kern 1pt} 15.0}}{{12.1}}$ | 0 |
| Декабрь | I–III | 20 | $\frac{{13.0{\kern 1pt} - {\kern 1pt} 18.0}}{{15.0}}$ | $\frac{{2.2{\kern 1pt} - {\kern 1pt} 2.8}}{{2.5}}$ | – | $\frac{{5.0{\kern 1pt} - {\kern 1pt} 9.0}}{{6.9}}$ | 0 |
| Январь | I–III | 6 | $\frac{{16.0{\kern 1pt} - {\kern 1pt} 16.3}}{{16.1}}$ | $\frac{{2.8{\kern 1pt} - {\kern 1pt} 5.0}}{{3.9}}$ | – | $\frac{{14.0{\kern 1pt} - {\kern 1pt} 48.0}}{{26.7}}$ | 0 |
Примечание. В числителе – пределы колебаний, в знаменателе – средние значения, “–” – отсутствие данных; для табл. 1–5, 7.
Таблица 2.
Интенсивность размножения прибылых самок рыжей полевки (по данным отлова)
| Месяц | Декада | Общее число исследованных самок | Масса тела, г. | Относительное число самок (% общего числа) | |||||
|---|---|---|---|---|---|---|---|---|---|
| в течке | беременных | кормящих | беременных и одновременно кормящих | рожавших | без следов размножения | ||||
| Прибылые ранних выводков | |||||||||
| Июнь | I | 6 | $\frac{{8.8{\kern 1pt} - {\kern 1pt} 39.8}}{{18.0}}$ | – | 16.7 | – | – | – | 83.3 |
| II | 59 | $\frac{{8.3{\kern 1pt} - {\kern 1pt} 38.0}}{{15.4}}$ | 1.7 | 18.6 | 5.1 | – | 1.7 | 72.9 | |
| III | 43 | $\frac{{10.1{\kern 1pt} - {\kern 1pt} 30.8}}{{17.3}}$ | – | 20.9 | – | – | 2.3 | 76.7 | |
| Июль | I | 77 | $\frac{{10.8{\kern 1pt} - {\kern 1pt} 34.6}}{{18.6}}$ | 6.5 | 39.0 | 1.3 | 1.3 | 5.2 | 48.1 |
| II | 187 | $\frac{{8.9{\kern 1pt} - {\kern 1pt} 36.1}}{{18.5}}$ | 1.1 | 27.8 | 10.7 | 4.3 | 11.2 | 44.9 | |
| III | 159 | $\frac{{5.9{\kern 1pt} - {\kern 1pt} 35.2}}{{19.8}}$ | 0.6 | 42.8 | 10.7 | 3.1 | 12.6 | 30.2 | |
| Август | I | 144 | $\frac{{6.9{\kern 1pt} - {\kern 1pt} 35.9}}{{18.4}}$ | 0.7 | 40.3 | 6.9 | 1.4 | 12.5 | 37.5 |
| II | 133 | $\frac{{7.6{\kern 1pt} - {\kern 1pt} 35.4}}{{20.0}}$ | – | 42.1 | 8.3 | 5.3 | 18.0 | 25.6 | |
| III | 108 | $\frac{{10.6{\kern 1pt} - {\kern 1pt} 35.7}}{{17.3}}$ | – | 19.0 | 7.0 | 1.0 | 39.0 | 21.0 | |
| Сентябрь | I | 73 | $\frac{{9.9{\kern 1pt} - {\kern 1pt} 30.6}}{{17.3}}$ | – | 12.3 | 23.3 | – | 38.4 | 26.0 |
| II | 92 | $\frac{{11.0{\kern 1pt} - {\kern 1pt} 40.8}}{{19.0}}$ | – | 1.1 | 7.6 | – | 34.8 | 56.5 | |
| III | 74 | $\frac{{14.2{\kern 1pt} - {\kern 1pt} 25.8}}{{19.2}}$ | – | 1.4 | 8.1 | – | 45.9 | 41.9 | |
| Октябрь | I | 20 | $\frac{{18.4{\kern 1pt} - {\kern 1pt} 29.2}}{{23.4}}$ | – | – | 5.9 | – | 88.2 | 5.9 |
| II | 12 | $\frac{{19.8{\kern 1pt} - {\kern 1pt} 28.1}}{{24.4}}$ | – | – | 18.2 | – | 63.2 | 18.2 | |
| III | 4 | $\frac{{21.0{\kern 1pt} - {\kern 1pt} 26.6}}{{22.7}}$ | – | – | – | – | 75.0 | 25.0 | |
| Прибылые поздних выводков | |||||||||
| Июль | I | 36 | $\frac{{10.5{\kern 1pt} - {\kern 1pt} 34.6}}{{16.0}}$ | – | 8.3 | – | – | – | 91.7 |
| II | 78 | $\frac{{4.2{\kern 1pt} - {\kern 1pt} 21.4}}{{14.5}}$ | – | 3.8 | – | – | – | 96.2 | |
| III | 100 | $\frac{{5.0{\kern 1pt} - {\kern 1pt} 28.8}}{{15.5}}$ | 1.0 | 4.0 | – | – | – | 95.0 | |
| Август | I | 85 | $\frac{{6.9{\kern 1pt} - {\kern 1pt} 26.4}}{{15.5}}$ | – | 15.3 | – | – | 4.7 | 80.0 |
| II | 85 | $\frac{{5.6{\kern 1pt} - {\kern 1pt} 32.7}}{{16.8}}$ | – | 4.7 | – | – | 5.9 | 17.6 | |
| III | 145 | $\frac{{8.2{\kern 1pt} - {\kern 1pt} 35.7}}{{9.1}}$ | – | 6.9 | 2.8 | – | 3.4 | 86.9 | |
| Сентябрь | I | 127 | $\frac{{8.1{\kern 1pt} - {\kern 1pt} 27.7}}{{16.7}}$ | – | 3.9 | 3.1 | – | 7.1 | 85.0 |
| II | 133 | $\frac{{5.6{\kern 1pt} - {\kern 1pt} 40.8}}{{17.3}}$ | – | 0.8 | 3.8 | – | 6.8 | 89.5 | |
| III | 140 | $\frac{{10.0{\kern 1pt} - {\kern 1pt} 24.0}}{{17.6}}$ | – | 4.3 | – | – | 8.6 | 87.9 | |
| Октябрь | I | 133 | $\frac{{9.9{\kern 1pt} - {\kern 1pt} 23.4}}{{16.5}}$ | – | – | 0.8 | – | 7.5 | 91.7 |
| II | 87 | $\frac{{10.2{\kern 1pt} - {\kern 1pt} 21.7}}{{14.6}}$ | – | – | – | – | 3.4 | 96.6 | |
| III | 39 | $\frac{{12.2{\kern 1pt} - {\kern 1pt} 18.1}}{{14.9}}$ | – | – | – | – | 12.8 | 87.2 | |
| Ноябрь | I–III | 20 | $\frac{{9.3{\kern 1pt} - {\kern 1pt} 28.2}}{{15.2}}$ | – | 5.0 | – | – | 5.0 | 90.0 |
| Декабрь | I–III | 18 | $\frac{{13.1{\kern 1pt} - {\kern 1pt} 16.5}}{{15.5}}$ | – | – | – | – | – | 100.0 |
| Январь | III | 3 | $\frac{{13.6{\kern 1pt} - {\kern 1pt} 18.5}}{{15.3}}$ | – | – | – | – | – | 100.0 |
| Февраль | I | 2 | 15.2, 17.6 | – | – | – | – | 50.0 | 50.0 |
Размножение самок майского и июньского приплода начинается в июне (беременных и кормящих 31.5%), но особенно интенсивно проходит в июле и августе (60–100%). В сентябре размножение постепенно затухает, a в октябре прекращается. Последние встречи беременных и кормящих самок приходятся на конец сентября–середину октября (27, 28.09. и 13.10.1958, 20.09.1960, 26.09.1968, 3.10.1970, 22.09.1987 и 15.10.2014). Еще два случая (20.09.1948 и 27.09.1953) отмечены в сохранившихся архивах лаборатории зоологии КарНЦ РАН. Хорошо заметны половые различия в интенсивности размножения прибылых полевок. Во второй и третьей декадах июня в репродукции участвует 47–86% молодых самцов и только 10–40% прибылых самок, что связано с более поздним созреванием самок. В условиях Карелии половая зрелость наступает у них приблизительно на полмесяца позже, чем у самцов, а в Финляндии разница достигает 3–4 недель (Kaikusalo, 1972).
Данные о частоте рождения выводков прибылыми самками представлены в табл. 2. Отметим, что большинство самок, родившихся в мае–июне, в течение второго месяца жизни успевает принести первый помет, в течение третьего – второй, а некоторые – и третий выводок (обычно это зверьки майского рождения). К концу сезона размножения по одному выводку дают 23.6, по два – 64.3, но три – 8.1% молодых самок весенне-летних пометов. Число выводков, которых успевает принести за лето каждая прибылая самка, зависит от времени ее рождения и продолжительности жизни. Родившиеся в начале мая и дожившие до осени при благоприятных условиях приносят не менее трех пометов, а появившиеся на свет позднее или погибшие летом – только один или два. Прибылые позднего (июльского) рождения, если и размножаются в текущем году, то дают за сезон не более одной генерации. Только в сентябре мы наблюдали обратную картину – более интенсивное размножение сеголеток в годы высокой весенней численности.
Как и у зимовавших самок, интенсивность размножения прибылых полевок (скорость созревания и степень участия в репродукции) варьирует по годам в обратной зависимости от весенней плотности популяции (Ивантер, 1975). В годы низкой весенней численности молодые самки не только более интенсивно созревают, в большем количестве и раньше приступают к размножению, но и приносят больше выводков, вследствие чего их “занятость” в размножении намного выше, чем в годы средней и высокой исходной численности. Так, при низкой плотности исходного поголовья в июне размножалось в момент поимки 50% исследованных самок, а в годы высокой численности 14.3%. В июле эти показатели составили 65 и 40%, в августе – 87 и 53%. Обратно пропорциональную зависимость между числом размножающихся прибылых самок ранних выводков и числом перезимовавших зверьков наблюдали в искусственно созданной в природе популяции рыжей полевки в Польше (Bujalska, 1970; Petrusewicz et al., 1971). Авторы объясняют эту связь территориальным поведением взрослых самок.
Однако это исключение лишь подтверждает правило, согласно которому при достижении популяцией определенной критической плотности размножение ее затормаживается. При низкой весенней численности репродукция, как правило, проходит более интенсивно и это приводит к раннему насыщению территории зверьками. Отсюда и раннее прекращение их размножения. Таким образом, позднелетне-осеннее размножение прибылых ориентировано не на весенний, а на летний уровень плотности популяции.
В южно-таежных лесах Волжско-Камского междуречья были проведены наблюдения за полевками, рожденными в мае и июне, пережившими зиму и повторно размножавшимися на втором году жизни (Тупикова, Коновалова, 1971). В Карелии мы этого не обнаружили. Здесь, очевидно, все прибылые ранних выводков вымирают еще осенью или зимой и имеют, следовательно, всего один сезон размножения.
Весенние и летне-осенние сезонные генерации грызунов, роль которых в воспроизводстве популяции будет описана ниже, различаются не только сроками и интенсивностью размножения (табл. 1 и 2), но и темпами линейного и весового роста (рис. 1 и 2). Полевки майских и июньских выводков растут очень быстро, особенно в первые три месяца жизни. Наоборот, зверьки летне-осеннего рождения характеризуются низким темпом роста. К концу осени они имеют минимальные размеры и массу, перезимовывают в таком состоянии и лишь весной следующего года, составляя группу зимовавших, резко увеличивают скорость роста (“весенний скачок”).
Рис. 1.
Линейный рост (по длине тела) зимовавших и прибылых рыжих полевок разного времени рождения. 1 – зимовавшие самки, 2 – зимовавшие самцы, 3 – самки ранних выводков, 4 – самцы ранних выводков, 5 – самки поздних выводков, 6 – самцы поздних выводков. I–III – декады; для рис. 1 и 2.
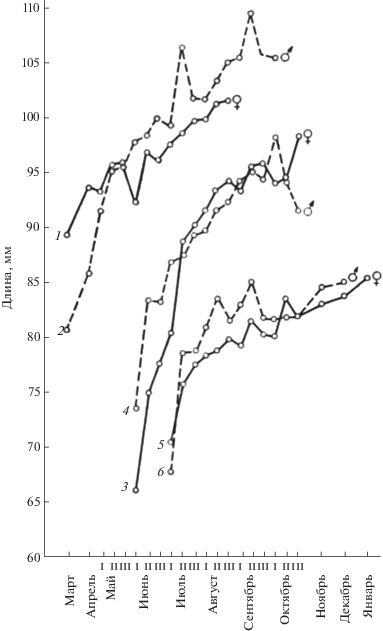
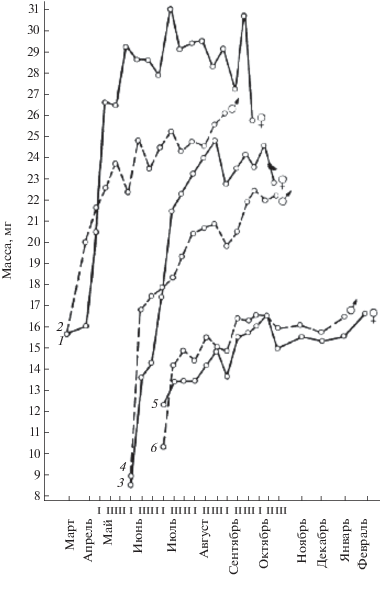
Различия в темпах роста помимо графиков иллюстрируют следующие цифры: средние масса и длина тела у зверьков ранних выводков в первый месяц их появления в уловах (в июне) составили 13.2 г и 76.4 мм, а у сеголеток поздних выводков (в июле) – 13.0 г и 76.4 мм. Следовательно, скорость роста в гнезде у них приблизительно одинаковая. Однако в дальнейшем они все более различаются, но на втором месяце жизни (в июле) прибылые раннего рождения имеют среднюю массу 19.3 г и среднюю длину тела 87.1 мм, тогда как сеголетки поздних выводков (в августе) – только 14.4 г и 80.3 мм; на третьем месяце жизни у прибылых ранних выводков – 22.2 и 92.2, а на поздних – 15.3 г и 81.6 мм соответственно. Если самцы и самки ранних генераций за первый месяц самостоятельной жизни (по отношению к исходному весу) прибавляют 6.3 г, или 46.2%, то прибылые поздних выводков – только 1.4 г (10.7%). Увеличение массы за второй месяц у ранних прибылых – 2.9 г (15%), у поздних – 1.1 г (7.6%). К третьему месяцу жизни масса прибылых ранних выводков составила по отношению к массе при выходе из гнезда 168% (привес 9 г), а у прибылых поздних рождений – 116% (привес 2.3 г).
Непосредственной причиной различий в темпах роста и развития у зверьков разных генераций может быть прямое воздействие среды. У полевок, родившихся весной, период послегнездового роста приходится на середину лета, т.е. на наиболее благоприятное время, отсюда и высокий темп их роста и созревания. Полевки же из поздних выводков развиваются на фоне менее благоприятных условий конца лета–начала осени (резкие перепады температуры, ненастье, недостаток сочного зеленого корма), поэтому рост их замедлен. Косвенным подтверждением этого служит тот факт, что при изменении экологических условий (например, в ситуациях высокой и низкой численности) интенсивность роста и развития сеголеток меняется. В годы депрессии численности, совпадающие с неблагоприятной экологической ситуацией, она минимальна, а в годы “пика” (при наиболее благоприятных экологических условиях) максимальна. К тому же и упитанность (а не только темпы роста и развития) оказывается у зверьков осенней генерации заметно ниже, чем у зимовавших и прибылых ранних выводков (Ивантер, 1975).
Однако нельзя не согласиться и с мнением С.С. Шварца с соавторами (1964), не исключающими роль генетических факторов, поскольку “весенние” и “осенние” генерации имеют разное происхождение: первые – прямые потомки (дети) зимовавших, а вторые – в большинство своем их внуки.
Но какова бы ни была причина этих различий, адаптивная их роль несомненна. Быстро развивающиеся сеголетки ранних выводков, почти поголовно участвующие в размножении в год рождения, обеспечивают рост населения к осени текущего года, а родившиеся во второй половине лета служат резервом популяции для следующего сезона размножения. Прибылые ранних выводков уже в июле несут основную нагрузку, связанную с воспроизводством популяции (табл. 3). Из общего числа размножающихся в этом месяце самок на долю сеголеток приходится 62%. В августе этот показатель еще выше – 78.2, в сентябре – 92.7, в октябре – 100%.
Таблица 3.
Возрастной состав размножающихся самок рыжей полевки
| Месяц | Общее число размножающихся самок | Относительное число самок, % общего числа | ||
|---|---|---|---|---|
| зимовавшие | прибылые выводков | |||
| ранних | поздних | |||
| Апрель | 3 | 100 | – | – |
| Май | 50 | 100 | – | – |
| Июнь | 144 | 81.2 | 11.8 | – |
| Июль | 297 | 43.8 | 52.5 | 3.7 |
| Август | 307 | 17.6 | 65.5 | 14.7 |
| Сентябрь | 203 | 8.4 | 66.0 | 25.6 |
| Октябрь | 25 | – | 44.0 | 56.0 |
| Всего | 1029 | 33.1 | 55.1 | 11.8 |
Если из 100 взрослых самок у 11 за сезон будет 1 выводок, у 60 – 2 выводка, у 25 – 3 выводка и у 3 – 4 выводка (согласно нашим данным), то всего они произведут 1247 молодых особей. Среди последних 1061 будут принадлежать к первым двум выводкам и не менее 90% из них (т.е. 477 самок) созреют в год рождения. С учетом частоты рождения пометов и средней их величины эти самки произведут ~3500 молодых. Общее число прибылых составит, следовательно, ~4750, из которых 72% воспроизвели сеголетки ранних выводков. Этот расчет подчеркивает значение прибылых полевок в воспроизводстве популяции.
Созревание ранних и несозревание поздних прибылых и вообще существование в популяции двух разнокачественных возрастных генераций сеголеток служит важнейшим условием для нормальной жизни популяции, обусловливает ее устойчивость, равномерное и успешное воспроизводство.
Главная задача ранних прибылых – обеспечение прироста численности в текущем году. Решая ее, они вынуждены быстро мобилизовывать все физиологические ресурсы и затем расплачиваются за это ранним старением и смертью. Они сыграли свою роль и больше не нужны популяции, так что их исчезновение к осени вполне оправдано и имеет глубокий экологический смысл. Прибылые поздних выводков не размножаются в год рождения, их организм не подорван интенсивной репродукцией, рост замедлен, так что они уходят в зиму при более мелких размерах и с не растраченными на размножение энергетическими ресурсами. И дело не только в том, что при более мелких размерах зверьков становится возможной более высокая их численность при той же биомассе: это создает более выгодное соотношение между биомассой животных и ресурсами местообитания (Кошкина, 1967). Мелкие размеры и связанная с этим незрелость, очевидно, способствуют более успешной перезимовке зверьков и по другим причинам. В частности, потребности у таких незрелых с замедленным ростом и пониженным обменом животных меньше, а устойчивость к неблагоприятным воздействиям выше. Биологический смысл этого явления близок к эффекту Денеля у землероек, но реализуется иначе.
По данным Я. Зейды (Zejda, 1971), рыжие полевки наиболее приспособлены к перенесению неблагоприятных условий зимы в Центральной Европе при массе 20, в Польше – 18, в Швеции – 16 г. Таким образом, по направлению к северу масса, благоприятная для зимовки, закономерно снижается. Напомним, что в Карелии она составляет 15–16, на Урале – 14–17 г. Следовательно, правило подтверждается. В Карелии, как и в Финляндии (Kaikusalo, 1972), имеются два сезонных минимума массы, соответствующих критическим периодам в жизни прибылых полевок: осенний (вторая половина октября), когда снежный покров еще не установился и недостаточно защищает зверьков, энергетические траты которых вследствие холодной погоды возрастают, и среднезимний (в декабре), связанный с наступлением сильных длительных холодов. Этот последний минимум менее выражен, очевидно, в связи с хорошей снежной защитой и завершением перестройки терморегуляции на зимний вариант.
Возрастная разнокачественность популяции и четкое “распределение обязанностей” между отдельными генерациями достаточно лабильны и служат одним из важнейших механизмов регуляции численности. Выше было отмечено, что интенсивность созревания молодняка и участие его в репродукции популяции обратно пропорциональны его численности. Это означает, что при низкой плотности популяция способна мобилизовывать все воспроизводительные способности и максимально реализовывать весь потенциал размножения, а при высокой – затормаживать репродукцию. Такой способ авторегуляции гораздо совершеннее и экономнее, чем контроль путем изменения численности помета или смертности, и в популяциях рыжей полевки играет, очевидно, главную роль.
ПЛОДОВИТОСТЬ ЗИМОВАВШИХ САМОК
Средняя численность выводка у зимовавших самок на основании подсчета эмбрионов и плацентарных пятен оказалась равной 5.82 ± 0.06 (табл. 4). При этом обращает на себя внимание подтверждение выявленной нами ранее (Ивантер, 1975) достоверной положительной асимметрии (косости) распределения самок по числу эмбрионов в выводке (А = 0.54 ± 0.17). Это свидетельствует о продолжающемся в северных популяциях процессе отбора на повышенную плодовитость и хорошо согласуется с известной тенденцией к увеличению среднего размера помета по направлению с юга на север (Наумов, 1948; Шварц, 1959, 1963). Наибольший размер выводка наблюдается на Урале и в таежных областях европейской части бывшего СССР (Кировская и Вологодская области, Республики Коми и Карелия), а наименьший – в южной части ареала (Украина, Белоруссия, Молдавия, Воронежская обл.). В то же время ряд исключений (например, сравнительно низкая плодовитость рыжих полевок в Лапландии и Ленинградской обл. и, наоборот, высокая – в Тульской обл. и Волжско-Камском крае) свидетельствует о несколько ином направлении тенденции. По-видимому, плодовитость увеличивается не с юга на север, а с юго-запада на северо-восток.
Таблица 4.
Размер выводка у зимовавших рыжих полевок
| Месяц и порядок выводка | Общее число беременных самок | Число самок (экз.) с числом эмбрионов (3–11) | Среднее число эмбрионов на 1 самку* | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
| Май | 35 | 4 | 7 | 14 | 6 | 2 | 1 | – | – | – | 4.94 ± 0.22 (2.7 + 2.3) |
| Июнь | 72 | 5 | 6 | 21 | 29 | 9 | 2 | – | – | – | 5.51 ± 0.16 (2.82 + 2.75) |
| Июль | 168 | 1 | 20 | 49 | 51 | 36 | 16 | 3 | 1 | 1 | 5.97 ± 0.12 (2.9 + 2.9) |
| Август | 51 | 9 | 6 | 4 | 17 | 9 | 4 | 2 | – | – | 5.60 ± 0.21 (2.9 + 3.1) |
| Сентябрь | 6 | – | – | – | 5 | 1 | – | – | – | – | 6.14 ± 0.22 (3.7 + 2.6) |
| Всего | 332 | 19 | 38 | 88 | 108 | 57 | 23 | 5 | 1 | 1 | 5.63 ± 0.08 (3.0 + 2.7) |
| I выводок | 60 | 4 | 9 | 22 | 14 | 8 | 3 | – | – | – | 5.36 ± 0.15 |
| II выводок | 91 | – | 10 | 22 | 33 | 13 | 10 | 2 | – | 1 | 6.03 ± 0.14 |
| III выводок | 42 | – | 6 | 7 | 15 | 10 | 4 | – | – | – | 5.98 ± 0.18 |
| IV выводок | 3 | – | – | – | 3 | – | – | – | – | – | 6.0 |
Примечание. * – в скобках указано число эмбрионов в правом и левом рогах матки; для табл. 4 и 5.
Размер выводка у зимовавших рыжих полевок, как и у большинства других мелких млекопитающих, закономерно изменяется по сезонам (табл. 5). В мае–июне он равен 5.4, в июле–августе – 6.1, в сентябре – 6.2. Согласно этому первые выводки бывают несколько меньше повторных (5.4 и 6.0 соответственно). Интересно, что в южно-таежных лесах Кировской обл. отмечена обратная закономерность (Тупикова, Коновалова, 1971).
Таблица 5.
Изменение численности выводка у прибылых самок рыжей полевки
| Месяц | Общее число беременных самок | Число самок, экз., с числом эмбрионов 1–10 | Среднее число эмбрионов на 1 самку* | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Прибылые ранних выводков | ||||||||||||
| Июнь | 21 | 1 | – | 1 | 1 | 7 | 5 | 2 | 1 | 3 | – | 6.0 ± 0.4 (3.3 + 3.0) |
| Июль | 154 | – | 1 | 2 | 23 | 52 | 42 | 23 | 8 | 3 | – | 5.6 ± 0.1 (2.7 + 2.9) |
| Август | 134 | – | – | 2 | 16 | 40 | 48 | 20 | 8 | – | – | 5.7 ± 0.1 (2.8 + 2.7) |
| Сентябрь | 74 | – | 1 | 4 | 12 | 13 | 15 | 11 | 12 | 2 | 4 | 6.2 ± 0.2 (3.4 + 2.7) |
| Июнь–сентябрь | 383 | 1 | 2 | 9 | 52 | 112 | 110 | 56 | 29 | 8 | 4 | 5.7 ± 0.1 (3.1 + 2.8) |
| Прибылые поздних выводков | ||||||||||||
| Август–сентябрь | 83 | – | – | 2 | 12 | 25 | 23 | 17 | 2 | 1 | 1 | 5.5 ± 0.2 (2.6 + 2.9) |
| Всего | 466 | 1 | 2 | 11 | 64 | 137 | 133 | 73 | 31 | 9 | 5 | 5.8 ± 0.1 (3.0 + 2.8) |
| I выводок | 243 | 1 | 1 | 3 | 38 | 65 | 72 | 41 | 15 | 5 | 2 | 6.5 ± 0.06 |
| II выводок | 187 | – | 1 | 6 | 23 | 60 | 50 | 22 | 15 | 4 | 2 | 5.6 ± 0.04 |
| III выводок | 19 | – | – | 1 | 3 | 7 | 6 | 3 | 2 | – | 1 | 5.8 ± 0.1 |
К сожалению, нельзя проанализировать возрастную изменчивость плодовитости взрослых самок, поскольку разница в месяц не всегда улавливается по развитию корней М2, но можно судить о ней по косвенным данным, оценивая возраст по длине тела зверьков (Ивантер, 1975). При этом мы сознательно отказались от такого критерия, как масса, хотя им часто пользуются в подобных случаях. Масса беременных самок сильно зависит от числа вынашиваемых эмбрионов и их размеров (стадии беременности), а также от упитанности, тогда как длина тела свободна от влияния этих факторов и поэтому точнее отражает возраст.
Наибольший средний выводок вынашивают самки длиной 100–109 мм (Ивантер, 1975). Более мелкие (молодые) отличаются меньшей плодовитостью, несколько ниже она и у самок старых (110 мм и более). Закономерность эта довольно отчетлива, а достоверность ее подтверждается дисперсионным анализом (Ивантер, 1975).
Плодовитость взрослых, как и размножающихся прибылых самок, зависит также от уровня весенней численности и интенсивности нарастания ее к осени. В южно-таежных лесах Вятско-Камского междуречья эту связь не удалось обнаружить (Тупикова, Коновалова, 1971), но согласно полученным нами данным она достаточно отчетлива: при высокой весенней численности средний размер выводка у зимовавших полевок составил 6.0, при средней – 5.1, при низкой же – 5.2. Подтверждается она и результатами дисперсионного анализа (Ивантер, 1975). Таким образом, обнаруживается достоверная обратная зависимость плодовитости взрослых самок от численности популяции. Возможно, это результат популяционного стресса, увеличивающего эмбриональную смертность, снижающего овуляцию и имплантацию.
ПЛОДОВИТОСТЬ СЕГОЛЕТОК
Данные о размерах выводка у сеголеток представлены в табл. 5. В среднем, согласно всем полученным данным, численность равна 5.63 ± 0.01 (с колебаниями от 1 до 10), т.е. совершенно такая же, как и у взрослых самок. Здесь также выявляются и ясные возрастные отличия. С возрастом средний размер выводка постепенно увеличивается. Сезонные изменения менее отчетливы. Можно судить лишь об увеличении плодовитости в сентябре.
Наибольший размер помета у зимовавших самок (табл. 4) наблюдается по второму и третьему выводкам (6.0 и 5.98), первый же выводок демонстрирует наименьшие показатели (5.36), что можно связать с его появлением в характеризующийся наименее благоприятными экологическими условиями весенне-летний период. У молодых же отмечается закономерное снижение индивидуальной плодовитости от первого выводка к последующим (от 6.5 к 5.6–5.8), что, как и авторы опубликованной работы (Тупикова, Коновалова, 1971), мы склонны объяснять постепенным истощением молодых самок в результате частых беременностей. У молодых самок, как и у зимовавших, наблюдается асимметрия в распределении выводков по числу эмбрионов (А = 0.30 ± 0.14), что подтверждает наше предположение о существовании в северных популяциях рыжей полевки отбора на повышенную плодовитость. При этом изменение размера выводка по годам не позволяет выявить четкой зависимости плодовитости молодых самок от численности популяции.
Проведенное нами сопоставление размера выводка у зимовавших и прибылых самок из разных биотопов (табл. 6) подтвердило выявленную ранее закономерность (Попов, 1960). Наименьший средний размер выводка у взрослых полевок отмечается в сосняках-зеленомошниках (5.3), далее следуют вырубки (5.7), ельники травяно-зеленомошные (5.8), лиственные (6.0) и смешанные приречные леса (6.1). Если перечислить эти местообитания в порядке улучшения их экологических свойств, то они расположатся в той же последовательности. Значит, плодовитость взрослых самок в известной мере определяется качествами окружающих условий. При их улучшении плодовитость самок достоверно увеличивается. У прибылых полевок картина менее четкая, вероятно, в связи с их большой подвижностью и менее тесной связью с определенным биотопом. Однако и здесь наименьшая плодовитость (5.5 против 6.0 в остальных местообитаниях) отмечена в бедных ресурсами сосняках (разница достоверна при Р > 0.98).
Таблица 6.
Плодовитость самок рыжей полевки в разных биотопах
| Биотоп | Зимовавшие | Прибылые | ||||
|---|---|---|---|---|---|---|
| n | lim | M ± m | n | lim | M ± m | |
| Сосняки | 20 | 4–9 | 6.5 ± 1.1 | 55 | 2–9 | 5.6 ± 0.3 |
| Ельники | 69 | 4–9 | 6.3 ± 0.4 | 23 | 4–10 | 5.5 ± 0.2 |
| Лиственные леса | 47 | 3–8 | 5.1 ± 0.3 | 53 | 3–10 | 5.3 ± 0.6 |
| Смешанные леса | 27 | 5–11 | 6.1 ± 0.3 | 29 | 2–8 | 5.8 ± 0.6 |
| Вырубки и мелколесье | 23 | 3–9 | 5.8 ± 1.2 | 39 | 2–10 | 5.4 ± 0.8 |
| Сельскохозяйственные угодья | 13 | 5–8 | 6.0 ± 1.0 | 26 | 4–7 | 4.5 ± 0.5 |
ЭМБРИОНАЛЬНАЯ И РАННЯЯ ПОСТЭМБРИОНАЛЬНАЯ СМЕРТНОСТЬ
В группе зимовавших полевок резорбция эмбрионов обнаружена у 5 из 93 (5.4%) исследованных беременных самок. Рассасывалось 6 зародышей из 539, т.е. средняя доля резорбции составила 1.1%. У прибылых самок эмбриональная смертность была вдвое выше. Она была отмечена у 17 из 137 самок (12.4%). Всего резорбировало 20 зародышей из 794, или 2.6%. Близкие значения приводят и другие отечественные авторы (Попов, 1960; Тупикова, Коновалова, 1971; и др.). В то же время у рыжих полевок Англии гибель эмбрионов достигает огромных размеров, что, в частности, было обнаружено у 49–69% исследованных самок (Coutis, Rowlands, 1969). Возможно, в этом и заключается причина констатируемой ими низкой плодовитости полевок английских популяций (средний размер выводка 4.0).
О размере “детской” смертности могут дать представление находки гнезд с выводками детенышей разного возраста. В нашем распоряжении имеются данные о 26 выводках: 17 со слепыми (в среднем 5.4 в выводке) и 9 со зрячими, свободно передвигающимися детенышами (4.9). Исходя из средней численности помета при рождении, равного 5.9 (с учетом эмбриональной смертности), постэмбриональная (в подсосный период) гибель может составить 16.9%. При этом в первую половину гнездовой жизни гибнет 8.5% детенышей, а во вторую – 9.25% оставшихся. Однако в действительности смертность в период нахождения детенышей в гнезде будет больше, так как в расчетах не учтена гибель выводков целиком.
Расчет ежемесячной гибели полевок в бесснежный период был проведен на основе средних за ряд лет данных о численности зверьков разного возраста в последовательные месяцы (табл. 7). При этом, не имея возможности оценить смертность зверьков разных месяцев рождения (для этого нужно определять возраст с точностью до полумесяца), мы проследили ее для трех возрастных групп – зимовавших, прибылых раннего (до середины июня) и позднего (со второй половины июня) рождения.
Таблица 7.
Ежемесячная смертность (% численности в предыдущем месяце) в популяции рыжей полевки
| Показатель | Апрель | Май | Июнь | Июль | Август | Сентябрь | Октябрь | Ноябрь |
|---|---|---|---|---|---|---|---|---|
| Общая численность | 0.48 | 0.40 | 0.93 | 1.4 | 2.3 | 3.7 | 2.9 | 4.7 |
| Численность зимовавших | 0.48 | 0.39 | 0.30 | 0.10 | 0.11 | 0.12 | 0.01 | 0 |
| Смертность за месяц | 30.4 | 15.9 | 32.5 | 17.7 | 56.9 | 71.0 | 67.1 | 100 |
| Численность прибылых ранних выводков | – | 0.03 | 0.22 | 0.50 | 0.84 | 1.24 | 0.33 | 0 |
| Смертность за месяц | – | – | – | – | 12.9 | 62.6 | 85.3 | 100 |
| Численность прибылых поздних выводков | – | – | – | 0.03 | 0.24 | 1.20 | 0.42 | 0.1 |
| Смертность за месяц | – | – | – | – | – | – | – | 69.4 |
В приведенных данных обращает на себя внимание значительная регрессия численности зимовавших полевок, особенно сильная начиная с августа. Мы связываем это с увеличением их гибели к концу лета. Наибольшего значения она достигает в сентябре (81%), к ноябрю зимовавшие полевки, как и размножавшиеся сеголетки, окончательно исчезают из уловов. Следует отметить и сравнительно высокую смертность зимовавших полевок весной (в апреле, например, она составляет 30.4%).
Прибылые зверьки ранних выводков также гибнут в основном осенью и до зимы практически не доживают. Высокая смертность этих полевок в переходные (ранневесенний и осенний) периоды связана с сезонной перестройкой физиологии (прежде всего терморегуляции). К тому же весной и осенью экологические условия для них наиболее неблагоприятны (неустойчивая погода, заморозки, холодные затяжные дожди, половодье и т.п.). Зимой же условия жизни стабилизируются, полевки живут оседло под надежной защитой снежного покрова и смертность их резко снижается.
ЗАКЛЮЧЕНИЕ
Отмеченные выше экологические особенности размножения рыжей полевки, проявляющиеся в специфических условиях северной периферии ареала, такие как высокая лабильность, рельефность и разнообразие адаптивных реакций репродуктивной сферы, их трансформация и смена по годам и сезонам в связи с изменением плотности населения зверьков, а также выработка широких проявляющихся как на организменном, так и популяционном уровне адаптивных комплексов, отличающихся динамичностью, высокой скоростью и глубиной компенсаторной перестройки, – все это вплотную приближает нас к разработке общей концепции периферических популяций мегаареальных политипических видов животных. Анализ закономерных географических отличий, характеризующих популяционную специфику рыжих лесных полевок, обитающих в разных частях видового ареала, подтвердил положение о том, что в экологическом центре (оптимуме) видового ареала плотность популяций не только выше, но и устойчивее, тогда как у северных границ распространения она изменяется в широком диапазоне и с большей амплитудой. Соответственно, более выражены и гораздо рельефнее проявляются в условиях северной периферии ареала и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить видам и их популяциям необходимое преодоление экстремальных условий экологического пессимума и в конечном счете содействовать как их широкой экспансии за границы ареала, так и успешному закреплению их в новых районах.
Работа проводилась в соответствии с Государственным заданием федерального исследовательского центра “Карельский научный центр Российской Академии наук”.
Список литературы
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 246 с.
Кошкина Т.В. Взаимоотношения близких видов мелких грызунов и регуляция их численности // Фауна и экология грызунов. М.: Изд-во МОИП, 1967. Вып. 8. С. 5–27.
Наумов Н.П. Очерки сравнительной экологии мышевидных грызунов. М.; Л.: Изд-во АН СССР, 1948. 203 с.
Попов В.А. Млекопитающие Волжско-Камского края. Казань: Госиздат, 1960. 466 с.
Тупикова Н.В., Коновалова Э.А. Размножение и смертность рыжих полевок в южно-таежных лесах Вятско-Камского междуречья // Фауна и экология грызунов. М.: Изд-во МГУ, 1971. Вып. 10. С. 145–171.
Шварц С.С. Биология размножения и возрастная структура популяций широко распространенных видов полевок на Крайнем Севере // Материалы по фауне Приобского Севера и ее использованию. Тюмень: Изд-во УрО АН СССР, 1959. Вып. 1. С. 239–254.
Шварц С.С. Пути приспособления наземных позвоночных животных к условиям существования в Субарктике. Т. 1. Млекопитающие (Труды Ин-та биологии Урал. фил. АН СССР). Свердловск: Изд-во УрО АН СССР, 1963. Вып. 33. 133 с.
Шварц С.С., Ищенко В.Г., Овчинникова Н.А., Оленев В.Г., Покровский А.В., Пястолова О.А. Чередование поколений и продолжительность жизни грызунов // Журн. общ. биологии. 1964. Т. 25. № 6. С. 417–433.
Buja1sкa G. Reproduction stabilizing elements in an island population of lethrionomys glareolus (Schreber, 1780) // Acta Theriol. 1970. V. 15. P. 24–31.
Coutis R.R., Rowlands I.W. The reproductive cycle of the Skomer vole (Clethrionomys glareolus skomerensis) // J. Zool. 1969. V. 158. № 1. P. 76–84.
Kaikusalo A. Population turnover and wintering of the bank vole, Clethrionomys glareolus (Schreb.), in southern and central Finland // Ann. Zool. Fenn. 1972. V. 9. № 4. P. 219–224.
Petrusewicz K., Bujalska G., Andrzejewski R., G1iwicz J. Productivity processes in an island population of Clethrionomys glareolus // Ann. Zool. Fenn. 1971. V. 8. № 1. P. 112–121.
Zejda J. Differential growth of three cohorts of the bank vole, Cleihrionomys glareolus Schreb. // Zook Listy. 1971. V. 20. № 3. P. 229–245.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая