

Известия РАН. Серия биологическая, 2021, № 4, стр. 408-422
Мышечная система Gyrocotyle urna (Plathelminthes, Gyrocotylida)
И. В. Ястребова 1, *, М. В. Ястребов 1
1 Ярославский государственный университет им. П.Г. Демидова
150057 Ярославль, пр. Матросова 9, Россия
* E-mail: mvy@uniyar.ac.ru
Поступила в редакцию 30.05.2019
После доработки 30.07.2019
Принята к публикации 29.08.2019
Аннотация
Описана архитектоника мускулатуры тела, прикрепительных органов и половых протоков Gyrocotyle urna. Показано, что боковые складки тела и лопасти розетки характеризуются наименьшим количеством и разнообразием мускулатуры. В покровах положение каждого шипа регулируется тремя группами мышц. Бóльшая часть половых протоков окружена мышечным плексусом. Присосковидный орган характеризуется чрезвычайной концентрацией мускулатуры. От присосок трематод его отличает обратная пропорциональность в степени развития радиальных мышц, с одной стороны, и прочих групп мышц – с другой. Обсуждается функциональная нагрузка различных групп мышц, механизмы фиксации и локомоции гельминта.
Паразитические плоские черви, обитающие в пищеварительных трактах позвоночных животных, с экологической точки зрения, являются реофильными формами, которые вынуждены противостоять изгоняющему потоку веществ. Для сохранения локализации они, как правило, используют активную фиксацию, т.е. сочетание процессов прикрепления и локомоции (Ошмарин, 1960). Доля каждого процесса существенно отличается в разных систематических группах. Отличия проявляются, в частности, в особенностях внешнего строения и архитектонике мускулатуры гельминтов. Изучение архитектоники мускулатуры – один из путей к познанию способов освоения среды паразитическими червями и закономерностей их морфологической эволюции. В этом плане существенный интерес представляет небогатый видами класс Gyrocotylida Bychowsky, 1937. Его представители, обитающие в спиральном клапане хрящевых рыб, имеют единообразный, легко узнаваемый габитус. Мускулистое тело со своеобразными прикрепительными органами на полюсах и фестончатыми складками на боковых сторонах предполагает высокое своеобразие пространственного распределения мышечных элементов в теле гельминтов. Однако относительно полное иллюстрированное описание мускулатуры гирокотилид в настоящее время отсутствует, и его выполнение является целью настоящей работы.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом послужили половозрелые представители вида Gyrocotyle urna Grube et Wagener, 1852 из спирального клапана химеры европейской (Chimaera monstrosa L., 1758), добытой в Баренцевом море. Морфология гельминтов изучена под световым микроскопом МИКМЕД-1 (ОАО “ЛОМО”, Россия) по полным сериям фронтальных, сагиттальных и поперечных парафиновых срезов (по две серии в каждой проекции) толщиной 7 мкм, изготовленных по стандартной методике и окрашенных методом Маллори.
Промеры выполнены с помощью градуированной окулярной линейки. В размерах присосковидного органа указаны последовательно длина вдоль оси органа, ширина по ширине тела и толщина в дорсовентральном направлении. При измерении диаметров мышечных волокон первый диаметр (вертикальный) перпендикулярен базальной пластинке, вдоль которой идет волокно, второй (горизонтальный) параллелен ей. Если первый диаметр больше второго, форма поперечного сечения волокна именуется овальной, если наоборот – поперечно-овальной, при равенстве диаметров – округлой. Для групп мышц, которые не удается расположить строго перпендикулярно плоскости среза, указана толщина, измеренная в одной плоскости. В мышечных слоях расстояние между соседними мышцами приводится в относительных величинах, а именно, в их горизонтальных диаметрах. По вертикальным диаметрам оценивается толщина слоя. Под расстоянием между диагональными мышцами подразумевается расстояние между теми из них, которые параллельны друг другу. Под углом пересечения мышц в этом слое понимается угол, ориентированный к концам тела. Рисунки выполнены с помощью рисовального аппарата РА-4 (завод “Прогресс”, СССР).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
У изученных экземпляров тело вытянутое, уплощенное дорсо-вентрально, длиной 11–18 мм при максимальной ширине 5–7 мм. На переднем конце тела находится присосковидный орган с субтерминальным устьем, которое окружено маленькой предротовой губой кольцевой формы. Сторона тела, к которой обращено устье присосковидного органа и на которой открывается отверстие матки, в дальнейшем изложении называется брюшной. Данная сторона тела более плоская, нежели спинная. Боковые края тела позади присосковидного органа образуют многочисленные крупные складки с общей дорсо-вентральной ориентацией, сходящие на нет возле заднего конца тела. На слегка суженном заднем конце тела расположен прикрепительный орган – розетка. Его центр представляет собой довольно обширное углубление в заднем “торце” тела. От краев углубления отходят 5–7 радиально направленных лопастей, каждая из которых делится на лопасти второго порядка, а те, в свою очередь, – на тонкие, длинные и многочисленные лопасти третьего порядка. Упомянутое углубление продолжается сужающимся каналом – воронкой. Воронка открывается наружу дорсальной порой с утолщенными краями, расположенной в основании розетки.
Покровы типичны для паразитических плоских червей и включают наружную пластинку тегумента, базальную пластинку, цитоны, а также три последовательно расположенных слоя мускулатуры – кольцевой, продольный и диагональный (рис. 1в). Покровы вооружены, и элементы вооружения образуют две топографически различные группы. Относительно немногочисленные “фарингеальные” шипы сконцентрированы вокруг и немного позади отверстия присосковидного органа и редко расположены на боковых сторонах тела до середины длины данного органа. Упорядоченность в их расположении не обнаружена. Соматические шипы располагаются диагональными рядами в задней части тела. На дорсальном и дорсо-латеральных участках покровов вооружение распространяется от проксимальной части матки до лопастей розетки, но не на сами лопасти. На вентральном и вентро-латеральных участках вооруженная зона короче и начинается немного впереди от уровня отверстия воронки. На названных участках частота и регулярность расположения шипов увеличиваются в направлении спереди назад, а также в направлении от плоскости симметрии тела к его боковым сторонам. Наиболее густо вооружение покрывает основание розетки. Все шипы вытянутые, округлые в поперечном сечении, расширяющиеся от скругленного основания, а затем плавно сужающиеся к вершине (рис. 1а). При максимальном диаметре 17.3–27.9 мкм их длина варьирует в пределах 74.9–163.2 мкм, и лишь боковые фарингеальные шипы несколько короче (не более 93 мкм). На срезах шипы выглядят слоистыми, словно состоят из разных веществ. Значительная часть каждого шипа находится в инвагинации базальной пластинки, а вершина – в наружной пластинке тегумента. С каждым шипом связано много мышечных волокон, описанных ниже.
Рис. 1.
Мускулатура G. urna: (а) – мышцы шипа на сагиттальном срезе тела; (б) – фрагмент семенного пузырька на фронтальном срезе тела; (в) – терминальная часть влагалища на сагиттальном срезе тела. 1 – наружная пластинка тегумента, 2 – базальная пластинка, 3 – отверстие влагалища, 4 – цитоны тегумента, 5 – кольцевые, 6 – продольные, 7 – диагональные мышцы покровов, 8 – окончания дорсо-вентральных мышц, 9 – шип, 10 – продольные мышцы шипа, 11 – сфинктер шипа, 12 – эпителий стенки семенного пузырька, 13 – мышечный плексус.

На спинной и брюшной сторонах тела максимальный диаметр кольцевых мышц покровов (1.4–3.8 × 1.4–3.9 мкм) наблюдается на уровне присосковидного органа. От его заднего края до основания розетки диаметр равен 1–2.4 × 1–1.9 мкм, а на боковых сторонах тела (за исключением складок) составляет 1–1.4 мкм. Самые тонкие кольцевые мышцы (не более 1 мкм) находятся в лопастях розетки и боковых складках тела. В районе отверстия присосковидного органа кольцевые мышцы поперечно-овальные в сечении, позади отверстия на спинной и брюшной сторонах передней половины тела – овальные, на прочих участках покровов – округлые.
Слой продольных мышц, подобно кольцевому, на боковых сторонах тела тоньше, чем на спинной и брюшной (табл. 1). При этом на всех сторонах тела наблюдаются сходные изменения его параметров в направлении спереди назад. Максимальная толщина наблюдается на уровне присосковидного органа, особенно на брюшной стороне тела. Позади органа до уровня семяприемника включительно продольный слой становится тоньше в несколько раз, уже без различий между спинной и брюшной сторонами. Позади семяприемника он вновь несколько утолщается. В боковых складках тела и лопастях розетки диаметр продольных мышц не более 1 мкм. В поперечном сечении они овальные или округлые на всех участках покровов. Расстояние между соседними мышцами в кольцевом и продольном слоях не превышает их горизонтального диаметра. В покровах полостей розетки и воронки порядок обсуждаемых слоев обратный. Здесь диаметр овальных в сечении кольцевых мышц составляет 1–4.8 × 1–2.9 мкм. Продольный слой в покровах полости розетки тоньше кольцевого (1–2.9 × 2.4–4.8 мкм), а в воронке, напротив, толще (2.9–6.7 × 1.4–5.8 мкм).
Таблица 1.
Диаметр продольных мышц покровов G. urna (мкм)
| Участок тела | Сторона | ||
|---|---|---|---|
| спинная | брюшная | боковая | |
| На уровне присосковидного органа | 1.4–4.8 × 1–2.4 | 2.9–6.7 × 1.9–3.9 | 1–1.9 × 1–1.9 |
| От присосковидного органа до матки | 1.4–2.4 × 1–1.4 | 1.4–2.4 × 1–1.4 | 1–1.4 × 1 |
| На уровне матки и семяприемника | 0.5–1.4 × 0.5–1.4 | 0.5–1.4 × 0.5–1.4 | 0.5–1.2 |
| От семяприемника до основания розетки | 1–1.9 × 1–1.4 | 1–1.9 × 1–1.4 | 0.5–1.2 |
| Основание розетки | 0.5–2.9 × 0.5–1.9 | 0.5–2.9 × 0.5–1.9 | 0.5–1.4 × 0.5–1.4 |
Диагональная мускулатура располагается сразу под цитонами тегумента. Она не обнаружена в покровах полостей воронки и розетки и в лопастях третьего порядка розетки. Угол пересечения диагональных мышц тупой. В каждой ячейке табл. 2 последовательно представлены толщина отдельных мышечных волокон, толщина диагонального слоя и расстояние между соседними мышечными волокнами на соответствующем участке покровов. Максимальная толщина диагональных мышц не меняется от переднего конца тела до заднего края семяприемника и на брюшной стороне тела немного превосходит таковую на спинной стороне. Позади семяприемника толщина убывает на обеих сторонах тела. Самые тонкие мышечные элементы (1–1.9 мкм) находятся в боковых складках тела и в лопастях розетки. Оценка толщины не отдельных мышц, а слоя в целом показывает иную закономерность: разница в толщине на спинной и брюшной сторонах тела просматривается лишь на уровне присосковидного органа. Позади органа слой становится более рыхлым, и значение данного показателя резко нарастает, уменьшаясь вновь только в основании розетки. На боковых сторонах тела толщина слоя на уровне присосковидного органа составляет 6.7–7.7 мкм, а позади него уменьшается с 19.2 мкм в проксимальных частях боковых складок до 4.9 мкм в их дистальных частях. В лопастях розетки происходит аналогичное уменьшение с 12 до 3.7 мкм.
Таблица 2.
Параметры диагональной мускулатуры покровов G. urna (мкм)
| Участок тела | Сторона | |
|---|---|---|
| спинная | брюшная | |
| На уровне присосковидного органа | 2.9–4.8
5.8–9.6 6.7–14.4 |
2.9–5.8
14.4–19.2 4.8–14.4 |
| От присосковидного органа до семяприемника | 1.4–4.8
24.6–49.2 4.8–11.5 |
1.4–5.8
24.6–49.2 4.8–14.4 |
| От семяприемника до основания розетки | 1.4–2.9
24.6–49.2 4.8–9.6 |
1.4–4.8
24.6–49.2 3.8–11.5 |
| Основание розетки | 1–2.9 6.7–16.3 1.9–5.8 |
1–2.9
6.7–16.3 1.9–6.7 |
Периферической группой паренхимных мышц являются наружные продольные, расположенные в виде слоя под диагональными мышцами. Их передние концы крепятся к покровам в задней половине полости присосковидного органа, а также вокруг органа, задние концы – к покровам наружной (обращенной в полость кишечника хозяина) стороны лопастей первого и второго порядка розетки (рис. 2–5). На уровне присосковидного органа и в основании розетки форма сечения мышц поперечно-овальная при одинаковой толщине на всех сторонах тела (табл. 3). Позади данного органа форма сечения мышц овальная, реже округлая, а толщина резко возрастает на спинной и особенно брюшной сторонах тела, уменьшаясь вновь лишь в основании розетки. На боковых сторонах тела обсуждаемые мышцы самые тонкие, без закономерных изменений толщины в направлении спереди назад. В большей части тела они располагаются на расстоянии меньше собственного диаметра друг от друга. На участке от заднего края семяприемника до розетки расстояние составляет один диаметр, а в розетке и дистальных участках боковых складок тела – четыре диаметра.
Рис. 2.
Мускулатура G. urna на срединном сагиттальном срезе тела. 1 – диагональные, 2 – наружные продольные паренхимные, 3 – кольцевые паренхимные, 4 – дорсо-вентральные, 5 – внутренние продольные паренхимные мышцы, 6 – присосковидный орган, 7 – матка, 8 – проток матки, 9 – семенной пузырек, 10 – семяизвергательный канал, 11 – полость воронки, 12 – радиальные мышцы розетки, 13 – хордальные мышцы воронки, 14 – скорлуповые железы, 15 – желточник, 16 – семяприемник, 17 – выделительный канал, 18 – фрагмент нервной комиссуры.
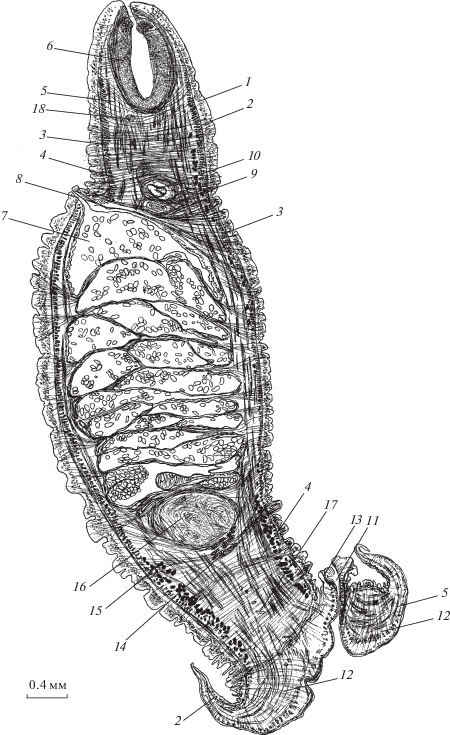
Рис. 3.
Мускулатура G. urna (поперечный срез тела на уровне середины матки). 1 – наружная пластинка тегумента, 2 – матка, 3 – яйца, 4 – желточники, 5 – влагалище, 6 – выделительный канал, 7 – нервный ствол, 8 – диагональные, 9 – наружные продольные паренхимные, 10 – кольцевые паренхимные, 11 – внутренние продольные паренхимные, 12 – дорсо-вентральные мышцы, 13 – мышечный плексус вокруг матки и влагалища.
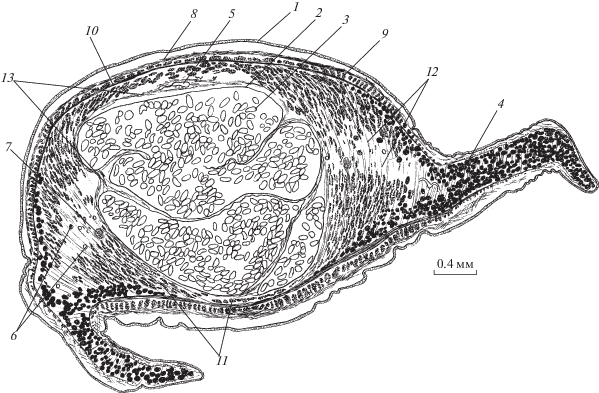
Рис. 4.
Мускулатура G. urna (поперечный срез тела позади семяприемника). 1 – наружная пластинка тегумента, 2 – диагональные, 3 – наружные продольные паренхимные, 4 – кольцевые паренхимные, 5 – дорсо-вентральные, 6 – внутренние продольные паренхимные мышцы, 7 – шип, 8 – продольные мышцы шипа, 9 – нервный ствол, 10 – выделительный канал.
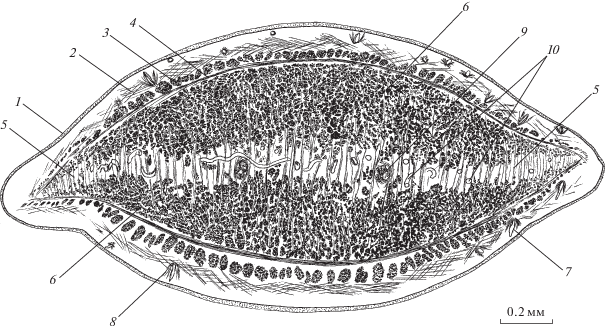
Рис. 5.
Мускулатура G. urna (поперечный срез тела на уровне розетки). 1 – наружная пластинка тегумента, 2 – диагональные, 3 – наружные продольные паренхимные, 4 – кольцевые паренхимные, 5 – внутренние продольные паренхимные мышцы, 6 – продольные, 7 – кольцевые мышцы покровов в полости розетки, 8 – радиальные, 9 –“поперечные” мышцы розетки, 10 – выделительный канал, 11 – лопасти розетки.

Таблица 3.
Диаметр наружных продольных паренхимных мышц G. urna (мкм)
| Участок тела | Сторона | ||
|---|---|---|---|
| спинная | брюшная | боковая | |
| На уровне присосковидного органа | 2.9–3.4 × 2.9–14.4 | 2.9–3.4 × 2.9–14.4 | 2.9–3.4 × 2.9–14.4 |
| От присосковидного органа до матки | 17.2–24.6 × 9.8–17.2 | 29.5–44.3 × 12.3–24.6 | 2.5–12.3 × 2.5–7.4 |
| На уровне матки и семяприемника |
17.2–24.6 × 9.8–17.2 | 17.2–49.2 × 9.8–24.6 | 2.5–7.4 × 2.5–7.4 |
| От семяприемника до основания розетки | 17.2–49.2 × 12.3–36.9 | 27.1–56.6 × 12.3–49.2 | 4.9–17.2 × 4.9–12.3 |
| Основание розетки | 9.6–19.7 × 9.6–24.6 | 9.6–19.7 × 9.6–24.6 | 9.6–19.7 × 9.6–24.6 |
Слой кольцевых паренхимных мышц наблюдается практически по всей длине тела, кроме лопастей третьего порядка розетки (рис. 2–5). На уровне передней половины присосковидного органа он расположен под диагональными мышцами, позади названного участка – под наружными продольными паренхимными мышцами, в воронке и в полости розетки – под кольцевыми мышцами покровов. Диаметр мышц составляет 4.8–14.4 × 4.8–7.7 мкм в устье присосковидного органа, 29.5–39.4 × 17.2–22.1 мкм на уровне передней и задней третей органа и 9.8–17.2 × 14.8–29.5 мкм на уровне его широкой средней трети. Между присосковидным органом и дистальной частью матки диаметр кольцевых паренхимных мышц значительно больше (24.0–72.0 × 9.6–38.4 мкм). На всех названных участках он одинаков на спинной и брюшной сторонах тела. На уровне матки и семяприемника диаметр мышц уменьшается, причем на спинной стороне тела сильнее, чем на брюшной (17.2–36.9 × 9.8–17.2 и 36.9–49.2 × × 24.6–36.9 мкм соответственно). Позади семяприемника до основания розетки включительно их диаметр вновь возрастает, достигая 49.2–61.5 × × 19.7–49.2 мкм. Максимальная толщина слоя (49.2–73.8 × 4.9–7.4 мкм) зарегистрирована на границе полостей розетки и воронки. В боковых складках тела обсуждаемые мышцы намного тоньше, чем в его центральной части (2.5–9.8 × × 2.5–12.3 мкм), а в лопастях розетки их диаметр минимален (1.2–3.7 × 1.2–3.7 мкм). В этих двух частях тела и на уровне средней трети присосковидного органа форма сечения мышц поперечно-овальная, редко округлая, на прочих участках покровов – в основном овальная. Расстояние между соседними мышцами не превышает их диаметра и лишь в лопастях розетки может достигать трех диаметров.
Внутренние продольные паренхимные мышцы расположены под кольцевыми паренхимными и прослеживаются от переднего конца тела до лопастей розетки включительно (рис 2–5). На уровне присосковидного органа данные мышцы образуют почти правильный однорядный слой. Над и под органом они имеют диаметр 1.9–9.6 × × 4.8–14.4 мкм и расположены вплотную друг к другу. По бокам от органа диаметр составляет 1.9–30.7 × 1.9–11.5 мкм, а расстояние между ними постепенно нарастает в латеральных направлениях от одного до пяти собственных диаметров. На участке от присосковидного органа до снования розетки внутренние продольные паренхимные мышцы образуют две мощные зоны на спинной и брюшной сторонах тела. В этих зонах их диаметр варьирует в пределах 4.9–93.9 × 3.7–39.4 мкм. На боковых сторонах тела (но не в боковых складках, где их нет), описываемые мышцы располагаются однорядным слоем. Здесь их толщина растет в направлении спереди назад: 2.5–19.7 × 2.5–12.3 мкм на участке от присосковидного органа до протока матки и 4.9–36.9 × 2.5–12.3 мкм на уровне матки и позади нее. Внутренние продольные паренхимные мышцы обнаружены не только на периферии тела, но и в его центральной части возле влагалища и семяизвергательного канала, между семенниками и редко рядом с семяприемником и яичником. В основании розетки (рис. 5) толща тела буквально заполнена этой группой мышц. Здесь мышечные элементы диаметром 7.4–61.5 × 4.9–19.7 мкм распределены равномерно и отстоят друг от друга на расстояние, не превышающее максимальный диаметр. От основания розетки данные мышцы заходят в лопасти, располагаясь слоем в центральной части каждой из них. В направлении спереди назад диаметр мышц плавно убывает и составляет в самых тонких лопастях 4.9–12.3 × 4.9–12.3 мкм при расстоянии между соседними мышцами до трех максимальных диаметров.
G. urna имеет хорошо развитую дорсо-вентральную мускулатуру. Она распространена от средней части присосковидного органа до места разделения розетки на лопасти. По бокам от присосковидного органа концы дорсо-вентральных мышц имеют наклон к плоскости симметрии тела, отчего мышцы обретают форму дуг, огибающих орган справа и слева. Концы мышц, расположенных сразу позади присосковидного органа, наклонены вперед (рис. 6–7). Здесь мышцы толще таковых на участке от присосковидного органа до матки (табл. 4). На уровне матки и семяприемника их толщина максимальна. Мышцы, расположенные между петлями матки, имеют крупные терминальные конусы (рис. 2). Кроме того, в узком пространстве между маткой и семяприемником проходит дорсо-вентральный мышечный пучок толщиной до 32 мкм. Непосредственно за семяприемником расположен еще более мощный пучок с толщиной неразветвленной части до 98.4 мкм. Оба его конца с большими терминальными конусами заметно смещены вперед. Позади данного пучка дорсо-вентральные мышцы вновь становятся тоньше, и в основании розетки наблюдается их наименьшая толщина. В боковых складках тела они в основном тоньше, чем в его центральной части, и расстояние между ними меньше. Здесь закономерности изменений толщины мышц и расстояния между ними сходны с таковыми в центральной части тела, но не идентичны им. Позади семяприемника толщина мышц в боковых складках уменьшается не столь заметно и выравнивается с толщиной в центральной части тела. Дополнительно отметим, что некоторые из дорсо-вентральных мышц крепятся к стенкам влагалища и семяизвергательного канала рядом с половыми отверстиями.
Рис. 6.
Мускулатура переднего конца тела G. urna на срединном сагиттальном срезе. 1 – наружная пластинка тегумента, 2 – кольцевые, 3 – продольные, 4 – диагональные мышцы покровов, 5 – наружные продольные паренхимные, 6 – кольцевые паренхимные, 7 – дорсо-вентральные, 8 – внутренние продольные паренхимные мышцы, 9 – наружные продольные мышцы, 10 – сфинктер, 11 – наружные кольцевые мышцы, 12 – внутренние продольные мышцы, 13 – внутренние кольцевые мышцы, 14 – диагональные мышцы, 15 – радиальные мышцы присосковидного органа.
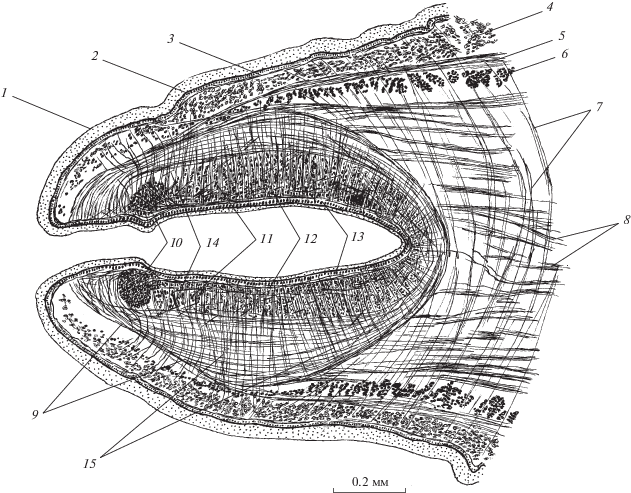
Рис. 7.
Мускулатура G. urna (поперечный срез тела на уровне середины длины присосковидного органа). 1 – наружная пластинка тегумента, 2 – базальная пластинка, 3 – диагональные мышцы покровов, 4 – кольцевые паренхимные, 5 – внутренние продольные паренхимные, 6 – дорсо-вентральные мышцы, 7 – радиальные, 8 – наружные продольные, 9 – диагональные, 10 – наружные кольцевые, 11 – внутренние продольные, 12 – внутренние кольцевые мышцы присосковидного органа, 13 – нервный ствол, 14 – выделительный канал.
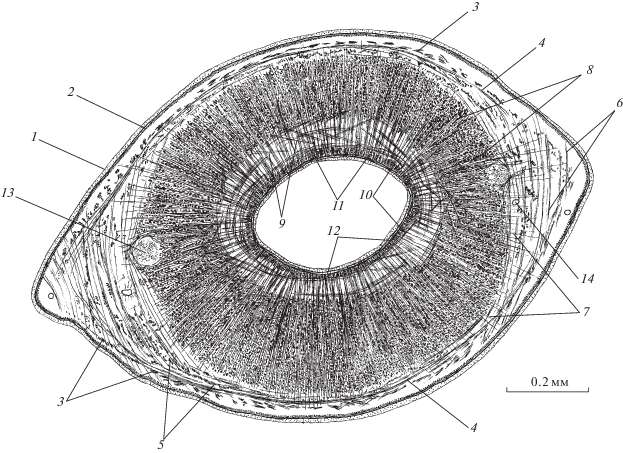
Таблица 4.
Толщина дорсо-вентральных мышц G. urna и (в скобках) расстояние между ними (мкм)
| Уровень | Участок тела | |
|---|---|---|
| центральная часть | боковые складки | |
| Присосковидный орган | 2.9–12.3 (2.9–12.3) |
1.9–2.9 (1.9–7.4) |
| От присосковидного органа до матки | 2.5–9.6 (28.8–33.6) |
1.9–4.8 (2.9–9.6) |
| Матка и семяприемник | 2.5–14.4 (50–390) |
2.5–7.4 (7.4–24.6) |
| От семяприемника до основания розетки | 2.5–6.2 (2.5–24.6) |
1.2–6.2 (2.5–9.8) |
| Основание розетки | 1–2.9 (6.7–24.6) |
1.2–4.9 (2.5–9.2) |
К паренхимной мускулатуре относятся также группы мышц, обнаруженные в розетке и воронке. Каждая лопасть первого и второго порядка розетки содержит многочисленные “поперечные” мышечные волокна толщиной 1 мкм или меньше, которые соединяют покровы на ее наружной и внутренней сторонах (рис. 5). Часть волокон собрана в пучки толщиной до 4.8 мкм. Помимо этого, в розетке до ее разделения на лопасти и в воронке имеются радиальные мышцы толщиной 1–3.8 мкм. Наконец, в воронке обнаружены немногочисленные хордальные мышцы (рис. 2). Они соединяют покровы в полости воронки на ее границе с розеткой и покровы апикальной части воронки. На их наружных концах хорошо выражены терминальные конусы. Толщина неразветвленных участков 4.8–9.6 мкм.
Интересной особенностью архитектоники паренхимной мускулатуры является наличие вокруг влагалища (за исключением терминальной части) и матки общего для обоих органов плексуса из мышечных волокон толщиной 1–4.8 мкм (рис. 1б, 3). На срезах в любой проекции плексус напоминает рыболовную сеть, ячеи которой не одинаковы по форме и размеру. Его толщина достигает 19.2 мкм. Аналогичные плексусы окружают проксимальную половину семенного пузырька и семяприемник.
К мускулатуре специального назначения мы относим мускулатуру шипов и половых протоков. Продольная мускулатура каждого шипа образована многочисленными тонкими (менее 1 мкм) волокнами, отдельными или сгруппированными в пучки толщиной 1.9–2.9 мкм (рис. 1а). Внутренние концы этих волокон крепятся ко дну инвагинации базальной пластинки, в которой находится шип, а наружные – к базальной пластинке по периметру устья инвагинации. Столь же тонкие и многочисленные кольцевые мышцы шипа формируют в устье инвагинации сфинктер диаметром 11.5–18.2 × 1.9–5.8 мкм. Сфинктер вытянут вдоль шипа. Глубже сфинктера, примерно до середины глубины инвагинации кольцевая мускулатура представлена однорядным слоем волокон на расстоянии 1–4 собственных диаметра друг от друга. Подчеркнем, что кольцевые и продольные мышцы шипа лежат в плоскостях, перпендикулярных тем, в которых расположены одноименные мышцы покровов. С каждым шипом связаны окончания дорсо-вентральных мышц. Одни из них крепятся по периметру устья инвагинации базальной пластинки снаружи от продольных мышц шипа, другие – ко дну инвагинации.
Мускулатура разных участков половых протоков дифференцирована в разной степени. В стенке дистальной половины семенного пузырька обнаружены кольцевой и диагональный мышечные слои, а в стенке семяизвергательного канала последовательно расположены продольный, кольцевой и диагональный. Та же последовательность мышечных слоев наблюдается в дистальной части влагалища (рис. 1в). Диаметр продольных мышц составляет в обоих протоках 1–2.9 × 1–3.8 мкм. В стенке влагалища они расположены вплотную друг к другу, а в семяизвергательном канале расстояние между ними 1–2 собственных диаметра. Кольцевые мышцы диаметром 1–2.4 × 1–1.9 мкм разнесены на расстояние 1–3 диаметра. Показатели диагональной мускулатуры такие же, как у одноименных мышц покровов в основании розетки. Продольные мышцы влагалища заходят вглубь протока до уровня переднего края семенного пузырька. В стенке матки имеются тонкие (1 мкм и меньше), тесно расположенные кольцевые мышцы. В стенке ее короткого выводного протока, помимо кольцевой, обнаружена диагональная мускулатура. При этом кольцевые мышцы формируют хорошо различимый сфинктер толщиной 6.7–12.5 мкм.
Особым участком тела G. urna является присосковидный орган. В отличие от типичных присосок, мы называем его органом лишь условно ввиду отсутствия собственной оболочки (рис. 6–7). Он представляет собой углубление покровов на переднем конце тела, с которым топографически связано большое число мышечных элементов, относящихся как к мускулатуре покровов, так и паренхимной мускулатуре. Однако для удобства изложения мы описываем его как дискретное образование с условной границей по внешней поверхности слоя наружных продольных мышц. Группы мышц данного органа названы так же, как группы в присосках трематод (Ястребов и др., 2008) в силу сходства архитектоники.
Орган довольно крупный (995–1274 × 837–995 × × 762–930 мкм), слегка сжатый дорсо-вентрально, с почти цилиндрической полостью диаметром 298–381 × 251–298 мкм. В полости органа под покровами расположен слой внутренней кольцевой мускулатуры, а глубже него – слой внутренней продольной. Оба слоя фактически относятся к мускулатуре покровов и состоят из тесно расположенных, овальных в сечении мышечных волокон диаметром 2.9–5.8 × 1.4–3.8 и 5.8–11.5 × 1.9–3.8 мкм соответственно. Наружные кольцевые мышцы диаметром 1.4–29.5 × 2.5–7.4 мкм находятся непосредственно под внутренними продольными и образуют “многоэтажный” слой толщиной 91–123 мкм. Они располагаются как вплотную друг к другу, так и реже. Максимальные расстояния (12.3–17.2 мкм) обусловлены размером клеток паренхимы. Данная группа мышц формирует крупный (118.1–147.9 × 68.9–103.3 мкм) сфинктер из плотно “упакованных” мышечных волокон диаметром меньше 1 мкм. Внутренние продольные мышцы дают множество ответвлений в сфинктер, который пронизан ими по всему объему. Ответвления не выходят за пределы сфинктера и, возможно, крепятся к его элементам, создавая на данном участке мышечный ретикулум. Наружные продольные мышцы являются (по крайней мере, частично) производными внутренних продольных паренхимных мышц. Они образуют мощный слой толщиной в средней трети органа 172–204 мкм, состоящий из округлых в сечении мышечных волокон диаметром 1–3.7 мкм. Часть из них почти касается друг друга, часть разделена расстоянием до двух собственных максимальных диаметров. В передней трети органа толщина слоя снижается до 73.8–98.4 мкм, в задней трети – до 49.2–61.5 мкм. Передние концы наружных продольных мышц имеют терминальные конусы. Зона их крепления к покровам находится в устье органа впереди от сфинктера. На боковых сторонах органа зона протяженней, чем на спинной и брюшной, и подходит ближе к переднему концу тела. Диагональные мышцы толщиной 1–4.9 мкм расположены более рыхло, чем на иных участках покровов, и довольно глубоко заходят в наружный кольцевой слой. Радиальные мышцы выходят за пределы органа, соединяя покровы в его полости с покровами тела вокруг него. На наружных концах имеются высокие, но не широкие терминальные конусы. Толщина неразветвленных участков 1.9–5.8 мкм, расстояние между ними превосходит большую толщину в 2–3 раза.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Мускулатура G. urna мощная и разнообразная. Предыдущие исследования показывают, что в покровах снаружи от цитонов тегумента последовательно расположены кольцевой и продольный слои, а под цитонами – кольцевой, продольный и вновь кольцевой (Lyons, 1969). Целесообразно скорректировать представления цитированного автора в двух пунктах. Во-первых, мышечный слой, расположенный сразу под цитонами тегумента, является диагональным. Предположительно, он был принят за кольцевой потому, что диагональные мышцы, пересекающиеся под тупым углом, на срезах нередко похожи на кольцевые, и для определения их хода требуется сравнение срезов в разных проекциях. Во-вторых, мы относим к мускулатуре покровов лишь кольцевой, продольный и диагональный слои, а прочие – к паренхимной мускулатуре.
По вопросу о нижней границе покровов плоских червей нет общепринятого мнения. Если считать границей цитоны тегумента, то диагональную мускулатуру обсуждаемого вида следует отнести к паренхимной. Между тем “трехслойная” схема мускулатуры покровов хотя и не тотальна, но обычна в разных группах плоских червей (Иванов, 1952; Быховский, 1957; Гинецинская, 1968; Иоффе, 1981; Дубинина, 1982; Smyth, Halton, 1983; Rieger et al., 1991; Mair et al., 1998; Halton, Maule, 2004; Tyler, Hooge, 2004). При этом диагональные мышцы могут располагаться не только над, но и под цитонами тегумента, что отмечается, например, у трематод отряда Cyclocoelida Odening, 1961 (Ястребов, 1998; Ястребова и др., 2008).
Независимо от топографии диагональных мышц, мы считаем их наличие своего рода маркером монозоичности плоских червей. Монозоичные формы используют гидравлическую локомоцию, в ходе которой диагональные мышцы скручивают тело относительно его продольной оси. У полизоичных цестод, напротив, в покровах обычно регистрируются лишь кольцевые и продольные мышцы (Lumsden, Byram, 1967; Braten, 1968; Тимофеев, Куперман, 1968; Rees, 1969; Andersen, 1975; Протасова, 1977; Делямуре и др., 1985; Ward et al., 1986; Valkounova, 1987). Причина состоит, на наш взгляд, в частичном переходе полизоичных форм от гидравлической локомоции к волновой, чему способствует сильно вытянутая стробила. При волновой локомоции особое значение обретают продольные мышцы, изгибающие тело. У цестод (в дополнение к покровам) они обильно развиваются в паренхиме (Фрезе, 1965; Дубинина, 1966; Спасская, 1966; Мовсесян, 1977). Диагональные мышцы покровов характерны лишь для монозоичных видов отряда Caryophillaeidea Deneden in Carus, 1863 (Протасова и др., 1990). В полизоичных группах их наличие можно ожидать в области сколекса, шейки и, возможно, первых члеников стробилы.
Крупный размер шипов G. urna имеет два морфологических следствия. Во-первых, они располагается в глубоких инвагинациях базальной пластинки, что, вероятно, предохраняет от выламывания. Во-вторых, каждый шип “обслуживается” тремя группами мышц. Продольные мышцы шипа являются его протракторами. Помимо них в протракции участвуют дорсо-вентральные мышцы, окончания которых крепятся вокруг устья инвагинации, где расположен шип. Ретракция производится также дорсо-вентральными мышцами, но крепящимися уже к основанию шипа. Кольцевая мускулатура шипа формирует сфинктер. Возможно, его сокращение замыкает устье инвагинации, если шип втянут в нее полностью, а также фиксирует положение шипа на определенном уровне протракции или ретракции. Таким образом, движение шипов регулируется довольно тонко.
Вооружение G. urna имеет конвергентное сходство с вооружением адорального диска трематод семейства Echinostomatidae Dietz, 1909. Их объединяют крупный размер, заглубление в покровы и наличие собственных продольных мышц (Fujino et al., 1994). Причину сходства мы видим в высокой механической нагрузке на сравнительно немногочисленные шипы. У представителей вида Patagifer bilobus Rud., 1819 к основаниям угловых шипов диска подходят, как у G. urna, дорсо-вентральные мышцы, осуществляющие их ретракцию. Отличие шипа трематод состоит в том, что он не погружен в инвагинацию базальной пластинки, а целиком заключен в образованную пластинкой капсулу (Ястребов, Ястребова, 2014).
В региональной дифференцировке мускулатуры G. urna обращает на себя внимание инвертированное положение кольцевого и продольного слоев в покровах полостей розетки и воронки. Сходная картина обнаружена на некоторых участках тела трематод, а именно в хвостовой камере и хвостовом придатке Lecithochirium sp. (Lecithochiriidae Skrjabin et Guschanskaja, 1954), а также в вентральной части переднего сегмента тела и в органе Брандеса Ichthyocotylurus platycephalus (Creplin, 1825) (Strigeidae Railliet, 1919) (Ястребова, Ястребов, 1999; Ястребова и др., 2004). В названных случаях инверсия мышечных слоев отмечается на специализированных и филогенетически относительно молодых участках тела гельминтов. Возможно, ее причина кроется в происхождении данных участков и будет понята при изучении времени их закладки в онтогенезе и механизмов развития.
Пространственное распределение мышечных элементов показывает отсутствие диагональной мускулатуры в покровах полостей воронки и розетки и в лопастях третьего порядка розетки. В лопастях розетки нет также кольцевых паренхимных и наружных продольных паренхимных мышц, а в боковых складках тела – внутренних продольных паренхимных мышц. Обе группы продольных паренхимных и кольцевые паренхимные мышцы в розетке наиболее разрежены. Кроме того, в розетке и в боковых складках тела все мышцы покровов, дорсо-вентральные, продольные и кольцевые паренхимные мышцы имеют наименьшую толщину.
Таким образом, боковые складки тела и лопасти розетки характеризуются минимальным количеством мускулатуры и минимальным разнообразием групп мышц. Мускулатура сосредоточена главным образом в массивной “основной” части тела и способна изменять его форму любым необходимым способом при фиксации и передвижении. Тупой угол пересечения диагональных мышц свидетельствует, что они не столько скручивают тело, сколько служат синергистами кольцевой мускулатуры. Большой размер и, соответственно, вес тела обусловливает наличие мощной дорсо-вентральной мускулатуры, а также развитие в паренхиме продольных и в меньшей степени кольцевых мышц, дополняющих одноименные мышцы покровов.
Сокращение паренхимных мышц механически воздействует на полые репродуктивные органы и перемещает по ним гаметы и яйца. Особая роль в этом принадлежит мышечному плексусу вокруг половых протоков. Та его часть, что окружает мешковидное расширение матки, необходима также для выведения яиц во внешнюю среду. Наличие расширения позволяет предположить, что они выводятся не перманентно, а периодически, довольно большими порциями, хотя причины этого нам не ясны. В период накопления яиц узкий выводной проток матки замкнут сфинктером, отмеченным также другими авторами (Poddubnaya et al., 2009). При накоплении и выведении яиц сфинктер и плексус, по-видимому, работают в противофазе. Похожие матки с мешковидным расширением дистального участка и порционным выведением яиц отмечены у трематод (Ошмарин, 1975). Их сходство с маткой обсуждаемого вида является конвергентным.
Изучение срезов розетки G. urna показывает, что ее строение иное, чем описано на живых и фиксированных особях. Есть мнение, что развитие розетки начинается на постларвальной стадии с чашевидной прикрепительной структуры, захватывающей одну ворсинку кишечника хозяина. В ходе роста и развития червя число таких структур увеличивается и может превышать десяток (Halvorsen, Williams, 1968). Возможно, развитие розетки начинается именно так, но не учтено ее деление на лопасти у взрослых особей.
Архитектоника мускулатуры говорит о высокой подвижности всех частей розетки. В функционировании данного органа, по-видимому, используются два механизма прикрепления. Первый из них не требует мышечных усилий и обеспечивается комплексом желез, секреты которых способствуют как слипанию лопастей с тканями хозяина, так и его прекращению (Poddubnaja et al., 2008). Не исключено, что с субстратом слипаются и боковые складки тела гельминта. Учитывая конфигурацию розетки и складок, очевидно, что общая площадь контакта тканей паразита и хозяина весьма велика. Вторым механизмом фиксации служит присасывание. Размер полости розетки свидетельствует о значительном присасывательном эффекте. При фиксации увеличение полости производится сокращением радиальной мускулатуры розетки и заходящих в нее продольных паренхимных мышц.
В воронке радиальная и кольцевая мускулатура регулируют диаметр просвета, продольные мышцы – ее длину, а хордальные – то и другое. Очевидно, что быть самостоятельным органом прикрепления взрослых особей воронка не может из-за слишком мелкого размера. Розетка и воронка функционируют как единое целое, и последняя, по-видимому, способна создавать лишь дополнительный, не слишком большой присасывательный эффект. Мы считаем справедливым мнение, что отверстие воронки, закрытое во время присасывания, открывается для стравливания давления в полости розетки и открепления гельминта от субстрата (Halvorsen, Williams, 1968). Края дорсальной поры воронки заметно утолщены, но мускулатуры на данном участке не много. Возможно, закрывание поры осуществляется не только мышечными усилиями, но и гидравлически, нагнетанием жидкости в данный участок тела.
Относительный размер присосковидного органа, разнообразие, степень развития и архитектоника его мускулатуры показывают, что он является весьма подвижным прикрепительным образованием. Здесь наблюдается этап эволюции мышечной системы плоских червей, на котором отсутствие присосок в собственных оболочках не мешает эффективной фиксации.
Интересно сравнить мускулатуру присосковидного органа и прекрасно развитых брюшных присосок трематод. Сравнение с ротовыми присосками не столь целесообразно, так как они являются сквозными органами. Группы мышц, типичные для брюшных присосок трематод, помимо радиальных, включают наружные и внутренние кольцевые, а также наружные и внутренние продольные. Один или оба кольцевых слоя не всегда, но часто формируют в устье органа сфинктер. Более редкими являются хордальные мышцы, обычно проходящие наискось от устья присоски к ее оболочке позади устья. И совсем редко в брюшных присосках наблюдается диагональная мускулатура (Ястребов, Ястребова, 2014). Легко заметить, что в присосковидном органе имеются все перечисленные группы. Передние концы наружных продольных мышц по топографии соответствуют хордальным мышцам присосок и являются их функциональными аналогами, раскрывая отверстие органа.
При всем сходстве групп мышц, присосковидный орган сильно отличается от брюшных присосок трематод. Отличие состоит в обратной пропорциональности развития радиальных мышц с одной стороны и прочих групп мышц – с другой. В брюшных присосках трематод слои кольцевых, продольных и диагональных мышц составляют периферическую мускулатуру. Наружные и внутренние слои разделены толщей стенки присоски, где обычно нет никаких мышц, кроме радиальных. Это же можно сказать и о ротовых присосках трематод, за исключением представителей отряда Paramphistomatida Skrjabin et Schulz, 1937, имеющих в этих органах срединные слои мускулатуры (Burdakova et al., 2015). В присосковидном органе радиальная мускулатура, напротив, теряется на фоне кольцевых и продольных мышц, заполняющих практически весь его условный объем.
Скорее всего, причина наблюдаемого различия – наличие у присосок трематод оболочки. Оболочка делает орган компактным, и сравнительно слабая периферическая мускулатура достаточна для изменения его формы. В случае с присосковидным органом кольцевые и продольные мышцы перемещают большой объем тканей на переднем конце тела гельминта, в том числе за условной границей органа, что требует от них большой мощности. Можно предположить, что функционирование присосковидного органа связано больше с обжиманием субстрата, чем с созданием присасывательного эффекта. Добавим, что концы дорсовентральных мышц, огибающих орган сзади, имеют заметный наклон вперед и при сокращении производят его протракцию. Ретракторами органа являются внутренние продольные паренхимные мышцы, крепящиеся к нему сзади.
Общий обзор архитектоники мускулатуры G. urna показывает, что сохранение гельминтом локализации достигается прикреплением и перемещением. Органы прикрепления расположены на концах тела, и на каждом конце дополнены вооружением. Топография вооружения говорит о том, что на заднем конце тела прикрепительная зона не ограничивается розеткой, а заходит вперед от нее до границы распространения соматических шипов. Локомоция, вероятно, сходна с таковой у всех плоских червей (моногеней, ряда трематод и турбеллярий), прикрепительные органы которых расположены на полюсах тела. Однако у G. urna ожидаемы два нюанса. Во-первых, возможно, что боковые складки тела участвуют в локомоции ундулированием. Подобное явление отмечено для плоских боковых полей тела при перемещении трематоды Fasciola hepatica L., 1758 (Fasciolidae Railliet, 1895) (Русак, Папина, 1970). Во-вторых, в теле G. urna четко выделяются два специализированных участка отлично различимых на рис. 2, которым, по всей вероятности, принадлежит главная роль в удлинении и укорочении тела. Их общими чертами являются отсутствие массивных частей половой системы и высокая концентрация паренхимной мускулатуры. Протяженность переднего участка – от присосковидного органа до переднего края матки, второго – от заднего края семяприемника до основания розетки.
Таким образом, на примере G. urna можно видеть, что мышечная система достигает высокой степени сложности даже на относительно ранних этапах эволюции плоских червей. Несомненно, некоторые качественные черты данной системы (наличие или отсутствие определенных групп мышц, последовательность мышечных слоев и др.) являются диагностическими или специфическими признаками таксонов различного ранга. Кроме того, сравнение принципов устройства мышечных систем в разных классах Plathelminthes полезно для выявления филогенетических связей внутри типа не меньше, чем сравнение покровов, половых систем или иных структур. В настоящее время материал по сравнительной анатомии мышечных систем фрагментарен, но его накопление и анализ имеет большие перспективы.
Список литературы
Быховский Б.Е. Моногенетические сосальщики, их система и филогения. М.–Л.: Изд-во АН СССР, 1957. 509 с.
Гинецинская Т.А. Трематоды, их жизненные циклы, биология и эволюция. Л.: Наука, 1968. 411 с.
Делямуре С.Л., Скрябин А.С., Сердюков А.М. Дифиллоботрииды – ленточные гельминты человека, млекопитающих и птиц / Основы цестодологии. Т. 11. Ред.: Судариков В.Е. М.: Наука, 1985. 200 с.
Дубинина М.Н. Ремнецы фауны СССР. М.–Л.: Наука, 1966. 261 с.
Дубинина М.Н. Паразитические черви класса Amphilinida. Л.: Наука, 1982. 143 с.
Иванов А.В. Строение Udonella caligorum Jonston, 1835 и положение Udonellidae в системе плоских червей // Тр. ЗИН АН СССР. Т. 14. М.–Л.: Изд-во АН СССР, 1952. С. 112–163.
Иоффе Б.И. Строение темноцефалид и морфологические изменения при переходе к паразитизму у плоских червей // Паразитология. 1981. Т. 15. № 3. С. 209–218.
Куперман Б.И. Функциональная морфология низших цестод. Л.: Наука, 1988. 167 с.
Мовсесян С.О. Цестоды фауны СССР и сопредельных территорий. Давэнеаты. М.: Наука, 1977. 272 с.
Ошмарин П.Г. К понятию “фиксация гельминтов” // Биологические ресурсы Дальнего Востока. М.: Изд-во АН СССР, 1960. С. 182–190.
Ошмарин П.Г. Экологическое значение расширения дистального участка матки некоторых трематод // Паразитология. 1975. Т. 9. № 5. С. 434–438.
Протасова Е.Н. Ботриоцефаляты – ленточные гельминты рыб / Основы цестодологии. Т. 8. Ред.: Рыжиков К.М. М.: Наука, 1977. 298 с.
Протасова Е.Н., Куперман Б.И., Ройтман В.А., Поддубная Л.Г. Кариофиллиды фауны СССР. М.: Наука, 1990. 238 с.
Русак Л.В., Папина Р.И. Двигательная активность Fasciola hepatica (Trematoda: Fasciolidae) in vitro // Зоол. журн. 1970. Т. 49. № 9. С. 1285–1289.
Спасская Л.П. Цестоды птиц СССР. Гименолепидиды. М.: Наука, 1966. 698 с.
Тимофеев В.А., Куперман Б.И. Ультратонкое строение кутикулы и субкутикулярного слоя процеркоида, плероцеркоида и взрослых особей Triaenophorus nodulosus (Pall.) // Паразитология. 1968. Т. 2. № 1. С. 42–49.
Фрезе В.И. Протеоцефаляты – ленточные гельминты рыб, амфибий и рептилий. М.: Наука, 1965. 538 с.
Ястребов М.В. Локомоторные аппараты некоторых Trematoda (Plathelminthes) с недиффернцированным телом // Зоол. журн. 1998. Т. 77. № 6. С. 627–638.
Ястребов М.В., Ястребова И.В. Мышечная система трематод (строение и возможные пути эволюции). М.: КМК, 2014. 343 с.
Ястребов М.В., Ястребова И.В., Филимонова Л.В. Об эволюционной пластичности мышечной системы гермафродитного поколения трематод (Plathelminthes, Trematoda) // Систематика и биология паразитов. Тр. Центра паразитологии ИПЭЭ РАН. Т. 45. М.: Наука, 2008. С. 184–213.
Ястребова И.В., Ястребов М.В. Мускулатура стенки тела Ichthyocotylurus platycephalus (Trematoda, Strigeidae) // Зоол. журн. 1999. Т. 78. № 11. С. 1267–1274.
Ястребова И.В., Ястребов М.В., Фрезе В.И. Мышечная система Lecithochirium sp. (Trematoda, Lecithochiriidae) // Успехи общей паразитологии. Тр. ИНПА РАН. Т. 44. М.: Наука, 2004. С. 433–450.
Ястребова И.В., Ястребов М.В., Филимонова Л.В. Мышечная система двух представителей отряда Cyclocoelida (Plathelminthes, Trematoda) // Систематика и биология паразитов. Тр. Центра паразитологии ИПЭЭ РАН. Т. 45. М.: Наука, 2008. С. 214–231.
Andersen K. The functional morphology of the scolex of Diphyllobothrium Cobbold (Cestoda, Pseudophyllidea). A scanning electron and light microscopical study on scolex of adult D. dendriticum (Nitzsch), D. latum (L.) and D. ditremum (Creplin) // Intern. J. for Parasitol. 1975. V. 5. P. 487–493.
Braten T. The fine structure of the tegument of Diphyllobothrium latum (L.). A comparison of the plerocercoid and adult stages // Z. Parasitenk. 1968. Bd. 30. S. 194–112.
Burdakova E.N., Yastrebova I.V., Yastrebov M.V. The Oral Sucker Muskles of Six Representatives of the Order Paramphistomatida (Plathelminthes, Trematoda) // Biology Bull. 2015. V. 42. P. 108–116.
Fujino T., Fried B., Hosier D. The expulsion of Echinostoma trivolvis (Trematoda) from ICR mice: extension/retraction mechanisms and ultrastructure of the collar spines // Parasitol. Res. 1994. V. 80. P. 581–587.
Halton D., Maule A. Flatworm nerve–muscle: structural and functional analysis // Can. J. Zool. 2004. V. P. 316–333.
Halvorsen O., Williams H. Gyrocotyle (Platyhelminthes) in Chimaera monstrosa from Oslo Fjord with emphasis on its mode of attachment and a regulation in the degree of infection // Nytt. Mag. Zool. 1968. V. 15. P. 130–142.
Lumsden R., Byram J. The ultrastructure of Cestode muscle // J. Parasitol. 1967. V. 53. P. 326–342.
Lyons K. The fine structure of the body wall of Gyrocotyle urna // Z. Parasitenk. 1969. Bd. 33. S. 95–109.
Mair G., Maule A., Shaw C., Halton D. Muscling in on parasitic flatworms // Parasitol. Today. 1998. V. 14. P. 73–76.
Poddubnaya L., Scholz T., Kuchta R., Levron C., Gibson D. Ultrastructure of the surface structures and secretory glands of the rosette attachment organ of Gyrocotyle urna (Cestoda: Gyrocotylidea) // Folia Parasitologica. 2008. V. 55. P. 207–218.
Poddubnaya L., Kuchta R., Levron C., Gibson D., Scholz T. The unique ultrastructure of the uterus of the Gyrocotylidea Poche, 1926 (Cestoda) and its phylogenetic implications // Syst. Parasitol. 2009. V. 74. P. 81–93.
Rees G. Cestodes from Bermuda fishes and an account of Acompsocephalum tortum (Linton, 1905) gen. nov. from the lizard fish Synodus intermedius (Agassiz) // Parasitology. 1969. V. 59. P. 519–548.
Rieger R., Salvenmoser W., Legniti A., Reindl S., Adam H., Simonsberger P., Tyler S. Organization and differentiation of the body-wall musculature in Macrostomum (Turbellaria, Macrostomidae) // Hidrobiologia. 1991. V. 227. P. 119–129.
Smyth J., Halton D. The physiology of trematodes. Cambridge: Cambr. Univ. press, 1983. 446 p.
Tyler S., Hooge M. Comparative morphology of the body wall in flatworms (Platyhelminthes) // Can. J. Zool. 2004. V. 82. P. 194–210.
Valkounova J. Comparative studies on the morphology, histology and histochemistry of Metacestodes (Hymenolepididae, Dilepididae and Dipylididae) // Folia parasitol. 1987. V. 34. P. 117–128.
Ward S., McKerr G., Allen G. Structure and ultrastructure of muscle systems within Grilotia erinaceus metacestodes (Cestoda: Trypanorhyncha) // Parasitology. 1986. V. 93. P. 587–597.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая