

Известия РАН. Серия биологическая, 2021, № 6, стр. 660-672
Анализ адаптационных возможностей репродуктивной системы рыжей полевки Myodes glareolus, обитающей на территориях природных геохимических аномалий
В. П. Мамина 1, *, Е. А. Малкова 1
1 Институт экологии растений и животных, Уральское отделение РАН
620144 Екатеринбург, ул. 8 марта, 202, Россия
* E-mail: mamina@ipae.uran.ru
Поступила в редакцию 09.08.2019
После доработки 04.03.2020
Принята к публикации 16.07.2020
Аннотация
Проведен морфофункциональный анализ семенников и яичников у рыжей полевки (Myodes glareolus Shreb.), обитающей на территориях природных геохимических аномалий. На аномальных участках у животных при более низком уровне численности, по сравнению с фоновым участком, наблюдается усиление функциональной активности семенников и яичников, увеличение потенциальной и фактической плодовитости, снижение доимплантационных потерь. Адаптационные возможности репродуктивной системы самцов связаны с паракринными факторами, обеспечивающими поддержание сперматогенеза на оптимальном для оплодотворения уровне. У самок атрезия фолликулов, происходящая на ранних стадиях их развития позволяет сократить затраты энергии на их рост. Более энергоэкономный путь отбора и созревания фолликулов способствует увеличению числа овулирующих ооцитов и репродуктивного потенциала самок.
Проблема воздействия химических факторов среды на состояние организма и популяцию в целом до сих пор привлекает пристальное внимание исследователей. Это связано с тем, что избыток определенных химических элементов вызывает изменения в эндокринной, иммунной и репродуктивной системах (Авцын и др., 1991; Агаджанян, Скальный, 2001; Гашев, 2003). Данные о влиянии геохимического фактора естественной природы на репродуктивную систему мелких млекопитающих незначительны и фрагментарны. Районы, для которых характерно избыточное поступление ряда химических элементов в пищевую цепь, в основном из подстилающей горной породы относят к естественным положительным геохимическим аномалиям (Вернадский, 1980; Башкин, Касимов, 2004). Естественные геохимические аномалии (ЕГА) отличаются от антропогенных продолжительностью существования. Животные, обитающие на природных положительных геохимических аномалиях, в течение многих поколений подвергаются воздействию избыточного содержания определенных химических элементов. Данная проблема актуальна для Уральского региона, характеризующегося широким распространением естественных геохимических аномалий. Выживание популяции непосредственно связано с воспроизводством населения. В большинстве случаев в качестве показателя состояния репродуктивной функции мелких млекопитающих используется интенсивность размножения: величина помета, потенциальная и фактическая плодовитость, эмбриональные потери. Показатели, характеризующие репродуктивную функцию животных, по нарастанию эффектов располагаются следующим образом: величина помета – плодовитость – способность к зачатию – фолликулогенез – функциональный сперматогенез (Принципы сравнительной оценки …, 1984). Изучение фолликулогенеза и сперматогенеза у мелких млекопитающих позволяет выявить адаптационные возможности репродуктивной системы животных к длительному воздействию комплекса тяжелых металлов (ТМ), и роль самок и самцов в формировании эмбриональных потерь. В природных популяциях наиболее часто наблюдается гибель эмбрионов до имплантации или в течение имплантации (от 10 до 30%), и значительно реже – после имплантации (0–2%) (Башенина, 1977; Артемьев, Окулова, 1981). Варьирование соотношения до и – постимплантационной гибели имеет различную природу: ранние летали могут возникать за счет снижения оплодотворяющей способности сперматозоидов и нарушения овариальной функции, поздние – результат мутагенного эффекта на сперматогенные клетки (доминантно-летальные мутации).
Следует отметить, что репродуктивная функция у рыжей полевки связана с плотностью популяции (Ивантер, 1975, 2005; Bujalska, 1975, 1983, 1990; Bujalska et al., 1994). Механизм авторегуляции численности популяции обусловлен физиологическими факторами стресса, а именно гипофизо-адреналовым механизмом регуляции репродуктивной функции (Christian, Davis, 1964; Christian et al., 1965; Christian, 1980, Роговин, Мошкин, 2007). Активизация гипоталамо-гипофизарно-надпочечниковой системы при достижении порогового уровня плотности популяции может вызывать у животных угнетение репродуктивной системы и торможение скорости полового созревания (Науменко и др., 1983; Novikov, Moshkin, 1998; Novikov et al., 2012). Степень проявления механизма авторегуляции численности зависит от условий обитания популяции и положения в ареале (Hansson, Henttonen, 1985; Ivanter, Osipova, 2000; Ивантер, 2005). У рыжей полевки в оптимуме ее видового ареала при высокой численности популяции, превышающей пороговые значения, включаются механизмы плотностно-зависимой регуляции (Жигальский, Бернштейн, 1986, 1989; Жигальский, 1994). В коренных южно-таежных темнохвойных лесах Среднего Урала (Висимский государственный природный биосферный заповедник, который может служить фоновой территорией в геохимическом отношении) у рыжей полевки установлен трехлетний популяционный цикл с чередованием трех фаз: депрессия, рост, пик (Жигальский, Кшнясев, 2000; Кшнясев, Давыдова, 2005). На данной территории при пике численности популяции у рыжей полевки наблюдаемая активация гипофиз-адреналовой системы сопровождается торможением полового созревания и угнетением репродуктивной функции (Кшнясев, Давыдова, 2005; Михеева и др., 2006; Жигальский, Мамина, 2015; Мамина, 2016).
Сравнительный анализ показателей, характеризующих репродуктивную систему рыжей полевки на геохимически аномальных участках и фоновой территории, отличающихся типом подстилающих горных пород, позволит выявить особенности размножения животных на данных участках.
Цель работы – провести анализ морфофункционального состояния яичников, семенников и сперматозоидов, оценить плодовитость и уровень эмбриональных потерь у рыжей полевки, обитающей на территориях геохимических аномалий для выявления адаптационных возможностей репродуктивной системы к длительному воздействию избыточного содержания кобальта, никеля и хрома.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили на трех территориях, которые по схеме лесорастительного районирования Урала находятся в пределах южно-таежного округа Средне-Уральской горно-лесной области (Лесорастительные…, 1973). Основные породы древесной растительности – ель, пихта, сосна, береза, осина, а также неморальные реликты – липа и ильм. Основные кустарники – малина, шиповник, из ягод – черника, брусника и другие. На основании результатов оценки содержания химических элементов в почве выделены аномальные территории: участок в окрестностях п. Уралец (участок 1) и д. Анатольская (участок 2) Горнозаводского района Свердловской области. На аномальных участках основными подстилающими породами являются ультраосновные породы, которые характеризуются высокими концентрациями Ni, Co, Cr, максимальные концентрации которых превышают среднеуральские фоновые значения в 66.7, 20 и 20 раз соответственно. Исследования проводили на участках 1 и 2 (57°40′ с.ш. 59°40′ в.д.; 57°40′ с.ш. 60°07′ в.д. соответственно) с максимальной концентрацией ТМ (центр аномалии). (Михеева и др., 2003; Байтимирова, Михеева, 2005). Площадь участков 1 и 2 составляла 44 и 43 км2 соответственно. Третий участок – Висимский государственный природный биосферный заповедник (57°21′ с.ш. 59°48′ в.д., площадью в 36 км2), основными подстилающими породами которого являются габбро, диориты, гранитоиды, не содержащие в максимальных концентрациях химические элементы. Данный участок использовали в качестве фоновой территории. Изучение циклического характера динамики популяции рыжей полевки в Висимском государственном природном биосферном заповеднике проводится сотрудниками нашего Института с 1995 г. Полученные данные свидетельствуют о наличии трехлетнего популяционного цикла у рыжей полевки с чередованием трех фаз: депрессия, рост, пик. В период интенсивного летнего размножения отсутствие или незначительное число половозрелых сеголеток является характерной особенностью фазы ”пик”. Одним из маркеров фазы “пик” может быть и переход популяции на следующий год в фазу “депрессия” (Жигальский, Кшнясев, 2000; Кшнясев, Давыдова, 2005).
Объектом исследования служила европейская рыжая полевка (Myodes glareolus Schreber, 1780) – доминирующий вид среди грызунов. Отловы рыжей полевки проводили с помощью давилок с крючком и живоловок (на стандартную приманку) в период интенсивного размножения (с конца июня до середины августа); на фоновой территории – с 2001 по 2006 гг.; на аномальном участке 1 – с 2001 по 2004 гг.; на участке 2 – с 2004 по 2006 гг. Учеты относительной численности рыжей полевки на каждом участке проводили на стандартных ловушко-линиях, состоящих из 50 давилок с расстоянием между ними 5 м. Определяли индекс уловистости по объединенному улову за 4 сут. Также на каждом участке устанавливали площадки живоловок 50–100 шт. с целью дополнительного отлова животных. Линия давилок и площадки живоловок на каждом участке были удалены друг от друга не менее чем на 200 м. По массе и размеру тела, степени развития тимуса, состоянию генеративной системы (у самок – влагалище, размер матки, наличие плацентарных пятен, эмбрионов, желтых тел, у самцов – масса семенников, их тургор, величина семенных пузырьков, наличие сперматозоидов) животных классифицировали на три функционально-возрастные группы: перезимовавшие особи, размножающиеся и не размножающиеся сеголетки. Возраст животных определяли по особенностям строения второго верхнего коренного зуба (Кошкина, 1955; Тупикова и др., 1970). Для гистологического анализа были использованы 110 самцов и 61 самка. Семенники животных фиксировали в 10%-ном формалине, парафиновые срезы семенника толщиной 5–7 мк окрашивали гематоксилином–эозином. Гистологические препараты семенников подвергались морфологическому и морфометрическому анализу. Индекс сперматогенеза рассчитывали по формуле: Is = ∑a/N, где а – количество слоев, выделенных в каждом канальце (1 слой – сперматогонии, 2 слой – сперматоциты, 3 – сперматиды, 4 – сперматозоиды); N – количество просчитанных канальцев (Ухов, Астраханцев, 1983). С помощью программного продукта Siams PHOTOLAB определяли диаметр семенных канальцев, размер ядер сперматозоидов и клеток эндокринного отдела семенника. От каждого самца на поперечном (срединном) срезе семенника подсчитывали число семенных канальцев с дегенерацией и без дегенерации в 50 семенных канальцах. Оценку морфофункционального состояния сперматозоидов проводили на эпидидимальных мазках, окрашенных азур–эозином. (Молнар, 1969). С помощью лабораторного счетчика просчитывают 200 сперматозоидов, учитывая живые (неокрашенные), мертвые (окрашенные) сперматозоиды и сперматозоиды с аномалией хвоста (Методические рекомендации …, 1978).
Морфологический и морфометрический анализ яичников проведен на серийных срезах, изготовленных по стандартной методике (Волкова, Елецкий, 1982; Бессалова, 2006) с использованием программного продукта Siams Photolab. Фактическую (число плодов в матке) и потенциальную (число желтых тел в яичниках) плодовитость исследовали только у беременных самок (Тупикова, 1964). Для оценки интенсивности процесса фолликулогенеза в яичниках беременных самок рыжей полевки определяли количество и размеры однослойных, вторичных (компактные, полостные), третичных фолликулов. Проведена оценка частоты встречаемости атретических фолликулов и уровня эмбриональных потерь у беременных самок.
Для статистической обработки количественных данных использовали программное обеспечение Statistica 5.5. Гипотезу нормальности распределения значений в выборках проверяли с помощью теста Колмогорова–Смирнова, после чего применяли дисперсионный анализ, t-критерий Стьюдента или непараметрические критерий Краскела–Уоллиса, тест Манна–Уитни. В рамках дисперсионного анализа после изучения действия факторов на все группы, для дальнейших попарных сравнений использовали post hoc тест: LSD-тест.
Для анализа натуральных частот использовали критерий χ2 (в таблице 2 × 2). Для статистических тестов был принят 5%-ный уровень значимости.
Таблица 1.
Половозрастная структура популяции рыжей полевки в районах естественных геохимических аномалий (участок 1 и 2) и на фоновом участке
| Год/район | Перезимовавшие, % | Сеголетки, % | ||||
|---|---|---|---|---|---|---|
| неполовозрелые | половозрелые | |||||
| самки | самцы | самки | самцы | самки | самцы | |
| 2004-участок 2 | 22 | – | 33 | 45 | – | – |
| 2006-участок 2 | – | – | – | 75 | – | 25 |
| 2001-участок 1 | 18 | 15 | 44 | 23 | – | – |
| 2003-участок 1 | – | – | 50 | 50 | – | – |
| 2004-участок 1 | 9 | 21 | 26 | 35 | – | 9 |
| 2001-фон | 14 | 10 | 43 | 33 | – | – |
| 2003-фон | 6 | 12 | 29 | 18 | 12 | 23 |
| 2004-фон | 14 | – | 38 | 33 | 10 | 5 |
| 2006-фон | 11 | 8 | 30 | 40 | 11 | – |
Таблица 2.
Морфологические показатели семенников и сперматозоидов у рыжей полевки с фоновой и аномальных территорий (М ± m; p < 0/05)
| Признак | Фаза популяционного цикла | |||||
|---|---|---|---|---|---|---|
| Рост | пик | |||||
| фон n = 35 | участок 1 n = 17 | участок 2 n = 15 | фон n =24 | участок 1 n = 17 | участок 2 n = 15 | |
| Диаметр семенных канальцев, мкм | 176.4 ± 25.6 | 178.8 ± 18.2 | 175.6 ± 16.8 | 151.0 ± 23.4* | 179.2 ± 23.8** | 179.6 ± 25.3** |
| Число канальцев с дегенерацией сперматогенных клеток, % | 5.1 ± 0.81 | 5.5 ± 0.9 | 5.1 ± 0.8 | 10.1 ± 1.3* | 5.4 ± 0.8** | 5.6 ± 0.6** |
| Индекс сперматогенеза | 4.0 ± 0.4 | 3.85 ± 0.5 | 3.7 ± 0.6 | 2.75 ± 0.13* | 3.85 ± 0.5** | 3.9 ± 0.4** |
| Площадь ядер клеток Лейдига, мкм2 | 35.2 ± 4.2 | 34.9 ± 2.8 | 36.1 ± 2.5 | 30.1 ± 3.1* | 35.7 ± 4.3** | 36.0 ± 3.8** |
| Площадь ядер клеток Сертоли, мкм2 | 57.8 ± 4.8 | 58.1 ± 5.6 | 56.3 ± 6.0 | 51.8 ± 6.7* | 57.0 ± 5.6** | 57,3 ± 6.0** |
| Площадь ядер головок сперматозоидов, мкм2 | 18.3 ± 2.7 | 17.4 ± 2.9 | 19.1 ± 2.8 | 13.8 ± 3.2* | 17.2 ± 2.5** | 18.0 ± 2.2** |
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Численность рыжей полевки на фоновой территории колеблется от сотых долей до 40 особей на 100 ловушко-суток, на аномальных участках – до 12.5 особей на 100 ловушко-суток (рис. 1). Относительное обилие рыжей полевки, обитающей на аномальном участке 1 (У1) ниже, чем на фоновой территории: в 2001 г. в 3.2 раза; в 2003 г – в 1.7 раза; в 2004 г – в 1.6 (рис. 1).
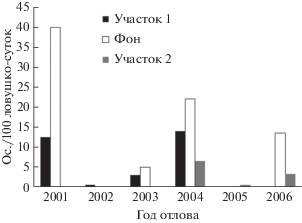
В отловах 2001 г. на исследуемых территориях сеголетки не размножались, в 2004 г. на фоновом участке половозрелые сеголетки составили 15%, на аномальном участке 1–9% (табл. 1).
Таким образом, на основании представленных данных 2001, 2004 гг. отнесены к фазе “пик”, 2002, 2005 гг. – к фазе “депрессия”, 2003, 2006 гг. – к фазе “рост”.
Морфологический анализ семенников и сперматозоидов. По морфологическим показателям семенников и сперматозоидов половозрелые сеголетки не отличаются от перезимовавших особей, поэтому данные по этим параметрам были объединены. На диаметр семенных канальцев статистически значимо влияет фаза цикла (F = 7.44; p < 0.001), геохимические условия (F = 11.23; p < 0.001) и взаимодействие этих факторов (F2, 11 = 10.65; p < 0.001). При пике численности у животных с фоновой территории диаметр семенных канальцев значимо меньше, чем при росте численности (p < 0.001; LSD-тест), на аномальных территориях – значимо больше по сравнению с фоновым участком (p < 0.001; LSD-тест; табл. 2). На фоновом и аномальных участках при росте численности популяции практически у всех животных диаметр семенных канальцев соответствует активному сперматогенезу.
Число семенных канальцев с дегенерацией сперматогенных клеток статистически значимо выше у животных с фоновой территории при “пике” численности по сравнению с фазой “рост” (χ2 = = 16.35; p < 0.001), на аномальных участках 1 и 2 значимо ниже по сравнению с фоновым (χ2 = 18.30; p < 0.001; χ2 = 15.70; p < 0.001 соответственно (табл. 2).
На индекс сперматогенеза, отражающего активность сперматогенеза, статистически значимо влияет фаза цикла (F = 13.75; p < 0.001;), геохимические условия (F = 12.60; p < 0.001) и взаимодействие этих факторов (F = 28.50; p < 0.001). У животных с фонового участка на пике численности индекс сперматогенеза значимо ниже по сравнению с ростом численности (p < 0.001; LSD-тест), на аномальных участках 1, 2 значимо выше по сравнению с фоновым участком (p < 0.001; LSD-тест) (табл. 2).
Площадь ядер клеток Лейдига и Сертоли, статистически значимо меньше у животных с фоновой территории на пике численности по сравнению с ростом численности (H = 16.38; n = 59; p < 0.001; H = 20.66; n = 59; p < 0.001 соответственно). У животных с аномальных территорий при пике численности площадь ядер клеток Лейдига и Сертоли значимо больше по сравнению с фоновым участком (H = 23.84; n = 56; p < 0.001 и H = 22.89; n = 56; p < 0.001 соответственно (тест Краскела–Уоллиса) и не изменяется при росте численности популяции (H = 2.75; n = 67; p = 0.2521; H = 1.62; n = 67; p = = 0.445 соответственно (табл. 2).
На размер ядер головки сперматозоидов влияет фаза численности (F = 7.33; p = 0.001), геохимические условия (F = 12.7; p = 0.001) и их взаимодействие (F = 5.42; p = 0.006). Размер ядер головки сперматозоидов у животных с фоновой территории при пике численности статистически значимо меньше по сравнению с ростом численности (p < 0.001, LSD тест). Однако у животных с аномальных участков 1 и 2 при пике численности размер ядер головок сперматозоидов значимо выше, чем у особей с фоновой территории (p < 0.001, LSD тест (табл. 2).
Количество живых сперматозоидов у животных с фоновой территории при пике численности статистически значимо ниже по сравнению с ростом численности (χ2 = 180.7; p < 0.001). На аномальных участках 1 и 2 при пике численности количество живых сперматозоидов значимо выше по сравнению с фоновым участком (χ2 = 34.75; p < 0.001; χ2 = = 76.06; p < 0.001 соответственно (рис. 2).
Рис. 2.
Частота встречаемости жизнеспособных сперматозоидов и с аномалией хвоста у рыжей полевки с фонового и аномальных участков 1, 2 на фазе роста и пика численности популяции. * – значимые различия между фазами “рост” и “пик” на фоновой территории, # # – значимые различия между фоновым участком и аномальными участками 1, 2 на фазе “пик”.
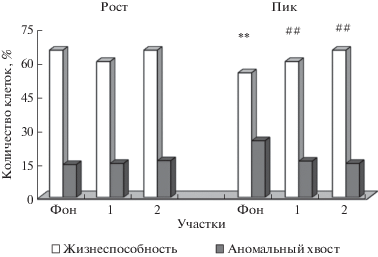
Число сперматозоидов с аномалией хвоста у животных с фоновой территории при пике численности статистически значимо выше по сравнению с ростом численности (χ2 = 47.58; p < 0.001). У животных, обитающих на аномальных участках 1 и 2, при пике численности количество сперматозоидов с патологией хвоста значимо ниже по сравнению с фоновым участком (χ2 = 28.82; p=0.001; χ2 = 21.01; p < 0.001 соответственно (рис. 2, 3). Сперматозоиды с аномальной головкой встречаются в единичных случаях (менее 1%).
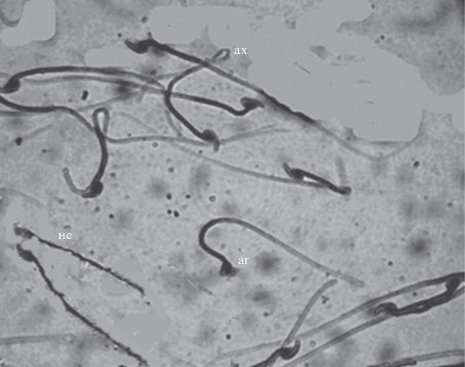
Рис. 3.
Сперматозоиды рыжей полевки: нс – нормальный сперматозоид, аг – сперматозоид с аномальной головкой, ах – сперматозоид с аномальным хвостом (в виде петли).
Морфологический анализ яичников. По морфологическим показателям яичников половозрелые сеголетки не отличаются от перезимовавших особей, поэтому данные по этим параметрам были объединены.
На фоновом участке в фазе “рост” численности количество растущих фолликулов и размер компактных фолликулов больше (t-критерий Стьюдента = 11.28, df = 15, p < 0.01; t = 2.61, df = 76, p = 0.01 соответственно (рис. 4а, 4в) по сравнению с пиком, также увеличивается число вторичных (t = 2.83, df = 15, p = 0.01; (рис. 4б) и третичных (t = 2.6, df = 15, p = 0.02 (рис. 4г) фолликулов.
Рис. 4.
Показатели интенсивности фолликулогенеза в яичнике рыжей полевки, обитающей в районах естественных геохимических аномалий (участок 1, 2) и на фоновом участке при разной численности популяции. (а) – * значимые различия на участке “фон” между фазами “рост”–“пик” (t = 11.28, df = 15, p < 0.01); *значимые различия между участками 1, 2 и фоновой территорией на фазе “пик” (t = 4.33, df = 16, p < 0.01; t = 5.24, df = 16, p < 0.01 соответственно); (б) – * значимое различие на участке “фон” между фазами “рост”–“пик” (t = 2.83, df = 15, p = 0.01); (в) – * значимое различие на участке “фон” между фазами “рост”–“пик” (t = 2.61, df = 76, p = 0.01); (г) – * значимое различие на участке “фон” между фазами “рост”–“пик” (t = 2.6, df = 15, p = 0.02).
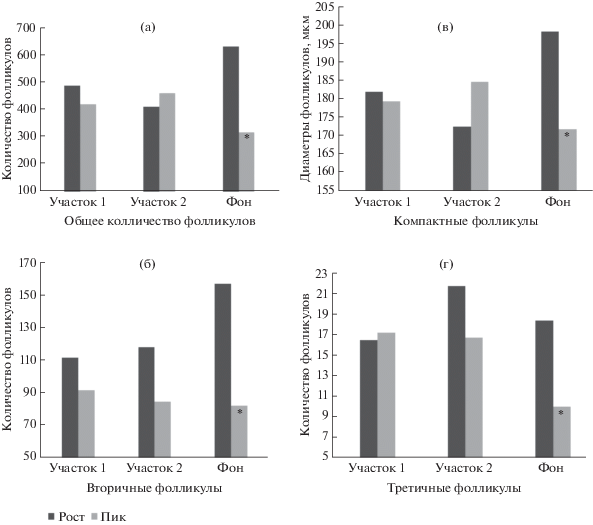
На аномальных участках 1 и 2 при пике численности число растущих фолликулов больше (t = 4.33, df = 16, p < 0.01; t = 5.24, df = 16, p < 0.01 соответственно (рис. 4а) по сравнению с фоновой территорией.
Количество вторичных фолликулов, подвергающихся атрезии, на пике численности выше у самок с участков 1 (t = 2.26, df = 16, p = 0.04) и 2 (t = 3.36, df = 12, p = 0.01 (рис. 5, 6) по сравнению с фоновой территорией. У животных с фонового участка при пике численности атрезия фолликулов преимущественно происходит на стадии третичных фолликулов, т.е. на поздних этапах их развития (рис. 5, 6).
Рис. 5.
Количество атретических фолликулов в яичнике рыжей полевки при высокой численности популяции (“пик”); * значимые различия между участком “фон” и участками 1, 2 (t = 2.26, df = 16, p = 0.04; t = = 3.36, df = 12, p = 0.01 соответственно).
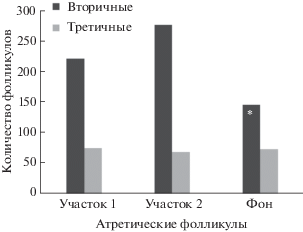
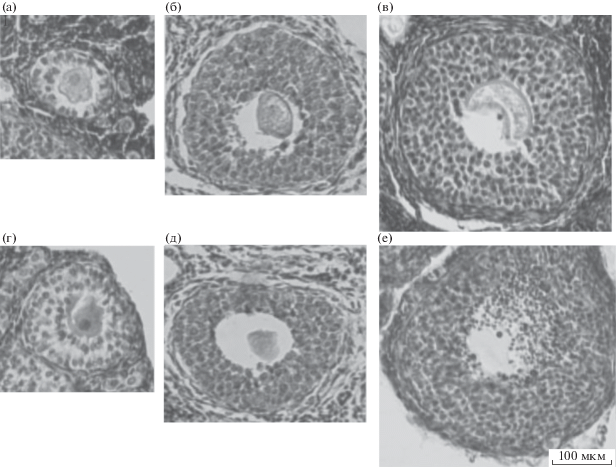
Рис. 6.
Растущие фолликулы с признаками атрезии в яичнике рыжей полевки. Лизис ооцита: (a), (в). Сморщенная блестящая оболочка: (б), (г), (д). Гибель клеток фолликулярного эпителия с образованием фрагментов ядер: (е).
Уровень потенциальной, фактической плодовитости и доимплантационной смертности у рыжей полевки на фоновом участке не зависит от фазы численности популяции и возраста животных (сеголетки, перезимовавшие), резорбция эмбрионов единична, поэтому для дальнейшего анализа данные по этим параметрам были объединены.
Значимых различий по потенциальной и фактической плодовитости, уровню доимплантационной смертности в зависимости от фазы численности и возраста (сеголетки, перезимовавшие) не обнаружено, поэтому данные были объединены. Фактическая плодовитость значимо выше на аномальных участках 1 и 2 (U = 90.5, Z = 2.45, p < 0.05; U = 36.0, Z = 2.0, p < 0.05 соответственно табл. 3), потенциальная плодовитость (число желтых тел беременности в яичниках беременных самок) – на участке 2 по сравнению с фоновой территорией (U = 19.50, Z = = –2.92, p < 0.05).
Таблица 3.
Плодовитость и доимплантационная смертность у рыжей полевки с фоновой и аномальных территорий (M ± m; p < 0.05)
| Признак | Фон n = 19 | Участок 1 n = 18 | Участок 2 n = 8 |
|---|---|---|---|
| Потенциальная плодовитость | 5.4 ± 0.22 | 6.1 ± 0.24 | 6.9 ± 0.30* |
| Фактическая плодовитость | 4.8 ± 0.23** | 5.7 ± 0.22 | 6.1 ± 0.52 |
| Доимплантационная смертность, % | 9.20 | 5.12 | 3.78 |
| Доля самок с доимплатанционными потерями, % | 31.58 | 27.78 | 25.0 |
Примечание. n – число животных; среднее арифметическое (М) ± ошибка среднего арифметического (m); * значимые различия между участком “фон” и участком 2 (U = 19.50, Z = –2.92, p < 0.05); ** значимые различия между участком “фон” и участками 1 и 2 (U = 90.5, Z = 2.45, p < 0.05; U = 36.0, Z = 2.0, p < 0.05).
Уровень доимплантационной гибели у самок на участках 1 и 2 ниже по сравнению с фоновой территорией, но различия статистически не значимы (табл. 3). Резорбция эмбрионов не обнаружена у животных с участка 1, а на участке 2 – единичные случаи.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Изучаемые районы естественных геохимических аномалий, как было представлено ранее, отнесены к природным биогеохимическим провинциям с избыточным содержанием кобальта, никеля и хрома, поэтому существование вида на данных территориях можно отнести к пессимальным условиям обитания. Ряд авторов отмечает, что в оптимальных условиях плотность населения вида не только выше, но и устойчивее, тогда как в пессимальных – она колеблется в большом диапазоне (Наумов, 1948; Майер, 1968; Европейская рыжая полевка …, 1981). Подобное явление обнаружено в популяции кустарниковой полевки, населяющей биогеохимическую молибденовую провинцию Анкавана (Сюзюмова, Карагезян, 1986). Результаты наших исследований показали, что, несмотря на низкий уровень относительной численности рыжей полевки на аномальных участках динамика многолетних колебаний численности популяций подобна динамике на фоновой территории. В литературе описаны случаи, когда циклика в популяциях сохраняется и при низком обилии населения, которого, вероятно, вполне достаточно для “включения” внутрипопуляционных механизмов, тормозящих размножение (Ивантер, 2005). В условиях Висимского заповедника пороговая плотность включения зависимых от плотности эффектов у рыжей полевки весной на фазе пика составляет 5–20 особей на 100 ловушко-суток (Кшнясев, Давыдова, 2005).
Снижение герминативной и эндокринной активности семенника у рыжей полевки на фоновой территории при пике численности обусловлено активацией гипофизарно-надпочечниковой системы, которая вызывает усиление продукции кортикостероидов (Михеева и др., 2006). На аномальных участках уровень плотности, вероятно, недостаточен для развития негативных эффектов адренокортикальной активности на репродуктивную систему по сравнению с фоновой территорией. Эффект адреналокортикальной активности на репродуктивную систему возможен лишь при превышении стрессором порогового уровня воздействия (Роговин, Мошкин, 2007).
Существуют две петли регуляции секреции тестостерона (Connell, Connell, 1977). В рамках большой петли кортикостероиды ингибируют выработку гонадотропных гормонов и тем самым угнетают андрогенную функцию клеток Лейдига. В рамках короткой петли (внутрисеменниковой) угнетение синтеза тестостерона может происходить за счет эстрадиола β-17, вырабатываемого клетками Сертоли, когда происходит снижение ингибина и как следствие – возрастание фолликулостимулирующего гормона (ФСГ). Показано, что у сезонно размножающихся грызунов в период регрессии семенников происходит уменьшение объема цитоплазмы и размера ядер у клеток Лейдига и Сертоли, при активации сперматогенеза – повышение функциональной активности клеток эндокринного отдела семенника (Райцина, 1985; Шевлюк и др., 1999). Наблюдаемое нами увеличение дегенеративных процессов в сперматогенном эпителии обусловлено снижением активности эндокринного отдела семенника и увеличением ФСГ, который служит маркером дегенеративных изменений в половых клетках (Rich, de Kretser, 1977). К наиболее часто встречающимся нарушениям в сперматогенном эпителии относятся десквамация и дезорганизация сперматогенного пласта.
Дегенерация сперматогенных клеток не всегда приводит к их гибели, возможно образование аберрантных клеток, из которых впоследствии происходит формирование атипичных или не жизнеспособных сперматозоидов. Значимое снижение числа живых сперматозоидов и увеличение количества сперматозоидов с патологией хвоста (хвост в виде петли и двойной хвост), обеспечивающего подвижность клеток, у самцов на фоновой территории при пике численности приводит к снижению оплодотворяющей способности сперматозоидов. Уменьшение размера ядер головок сперматозоидов свидетельствует о пониженном содержании ДНК, которое отражается на фертильности животных (Bouters et al., 1967; Sailer et al., 1996).
Увеличение герминативной активности семенника у животных с аномальных участков при максимальной численности связано с усилением андрогенной активности семенника. Известно, что в регуляции пролиферации и дифференциации клеток сперматогенного эпителия главная роль принадлежит тестостерону, синтезируемого клетками Лейдига. Рецепторы тестостерона обнаружены на клетках сперматогенного эпителия – сперматогониях, сперматоцитах и ранних сперматидах, присоединение гормона к рецепторам усиливает ядерный обмен (Sun et al., 1989, 1990; McLachlan et al., 1994; O’Donnell et al., 1994). Через ряд последовательных событий гормон осуществляет контроль биосинтеза белка и модулирует целый ряд внутриклеточных процессов, в том числе и клеточную пролиферацию. Следует отметить, что тестостерон регулирует функциональную активность клеток Сертоли, осуществляющих контроль над пролиферативной активностью сперматогониальных клеток. Усиление функциональной активности клеток Сертоли способствует поддержанию высокого внутрисеменникового уровня тестостерона. Кроме того, возрастание уровня ингибина, вырабатываемого клетками Сертоли, и блокирующего выработку ФСГ, уменьшает дегенеративные процессы в сперматогенном эпителии, что отражается на морфологическом состоянии сперматозоидов.
Увеличение индекса сперматогенеза, интегрального показателя генеративной функции семенника, на аномальных участках при максимальной численности популяции обусловлено паракринными факторами (Huleihel, Lunenfeld, 2004; Walker, Cheng, 2005).
У самок на всех исследуемых территориях количество однослойных фолликулов не зависит от уровня численности. Это связано с тем, что рекрутирование примордиальных фолликулов в основном контролируется паракринной взаимосвязью между ооцитом, фолликулярным эпителием, соседними текальными и интерстициальными клетками. Они находятся в микросреде, исключающей контакт с большинством других клеток и кровеносными сосудами, имеют ограниченный доступ к эндокринной системе. Экспрессия рецепторов ФСГ отмечается в фолликулярном эпителии первичных фолликулов уже после инициации их роста (Петров и др., 2017). Отсутствие межгодовых изменений размеров полостных фолликулов объяснятся подавлением их развития во время беременности внутрияичниковыми факторами роста: эпидермального (ЭФР) и трансформирующего (ТФР-а) через снижения рецепторной чувствительности клеток гранулезы фолликулов к ФСГ. Кроме того, рост полостных фолликулов подавляется во время беременности ингибинами, которые синтезируются плацентой (Gougeon, 1996). Уменьшение числа фолликулов и размера компактных фолликулов у животных с фоновой территории при максимальной численности популяции обусловлено активацией гипофизарно-надпочечниковой системы, которая вызывает угнетение синтеза гонадотропинов (Christian at al., 1963). Показано значение центральных механизмов в активации гипофизарно-надпочечниковой системы. Усиление нейросекреции вазопрессина в портальный кровоток гипофиза полевок в условиях переуплотненной популяции оказывает стимулирующее влияние на адренокортикотропную функцию гипофиза и приводит к угнетению репродуктивной функции полевок (Владимирова и др., 2006).
Процесс отбора овариальных фолликулов, их рост, созревание и овуляция регулируются не только гонадотропинами и половыми стероидами, но и сложными внутриорганными механизмами. Известно, что во время каждого овариального цикла из примордиальных фолликулов формируется пул растущих фолликулов, большинство из которых подвергается атрезии на разных стадиях развития (McGee, Hsueh, 2000). По современным представлениям ключевой процесс, лежащий в основе атрезии незрелых фолликулов это постнатальный апоптоз ооцитов. Регуляция апоптоза в ооцитах подразделяется на 2 основные категории: процесс, происходящий при естественном развитии яичников, и процесс, происходящий в ответ на патофизиологическое повреждение (Tilly et al., 2004). Митохондрии выполняют центральную роль в апоптозе, при этом наблюдается увеличение проницаемости митохондриальной мембраны. Баланс между про- и анти-апоптозных членов семейства Bcl-2 регулирует выход про-апоптозных веществ из митохондрий, ведущих к его запуску. Клеточный стресс (радиация, тяжелые металлы и др.) оказывает влияние на количество bcl-2 белков, определяя тем самым необходимость запуска апоптоза (Bröker et al., 2005). При этом более ранний отбор растущих фолликулов, позволяет сократить затраты энергии, связанные с их дальнейшим ростом, повысить качество и число овулирующих ооцитов. Наблюдаемое нами увеличение потенциальной плодовитости на аномальных участках, вероятно, связано с отбором растущих фолликулов.
Одной из возможных причин повышения потенциальной и фактической плодовитости у самок рыжей полевки на естественной геохимической аномалии является интенсификация функциональной активности щитовидной железы (Михеева, 2006). Тиреотропный гормон (ТТГ) и трийодтиронин (Т3), имеющие в яичниках рецепторы, оказывают стимулирующее действие на функциональную активность гранулезных клеток, секрецию прогестерона и эстрадиола желтым телом; влияют на способность ооцитов к оплодотворению и жизнеспособность эмбрионов Wakim et al., 1995; Oravec, Hlavacka, 2000).
Кроме того, согласно современным представлениям молекулярной биологии экспрессия BMP 15(CDF9B) гена важна для регуляции числа овулирующих ооцитов. У гетерозиготных животных по гену BMP15 происходит увеличение количества овулирующих ооцитов (Pasquale et al., 2004). Возможно, в популяциях рыжей полевки, обитающих на территориях природных геохимических аномалий, имеет место отбор гетерозиготных особей по данному признаку, обладающих повышенной плодовитостью.
Полученные результаты свидетельствуют о том, что половозрелые самки и самцы, обитающие в условиях естественных геохимических аномалий при высокой численности способны поддерживать и высокую репродуктивную активность,
Таким образом, наблюдаемые нами особенности функционирования репродуктивной системы самцов и самок рыжей полевки, обитающих на территориях естественных геохимических аномалий, являются результатом действия целого комплекса внутриорганных механизмов, которые можно считать проявлением неспецифической адаптивной реакции, не сопровождающейся патологическими изменениями (рис. 7). На популяционном уровне это проявляется общим снижением относительного обилия и “сглаживанием” плотностно-зависимых механизмов. Отмечаемые флуктуации численности, совпадающие с динамикой циклической популяции на фоновом участке, характеризуются поддержанием постоянства репродуктивного потенциала размножающихся особей.
Рис. 7.
Схема, отражающая состояние репродуктивной функции рыжей полевки, обитающей в условиях
природной геохимической аномалии. Стрелка  – усиление,
– усиление,  – подавление.
– подавление.
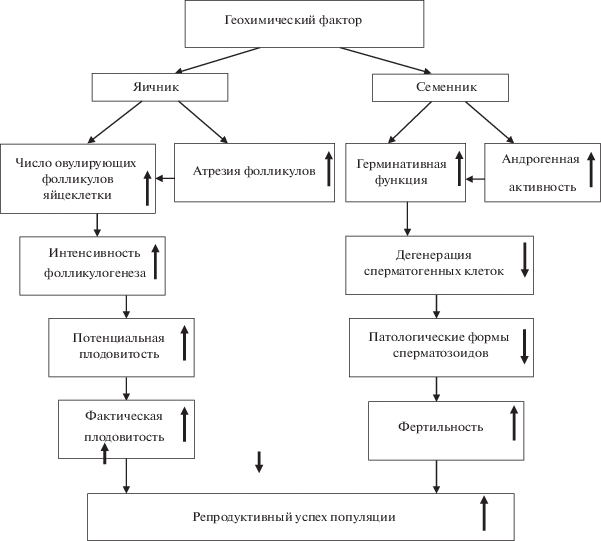
Подобная стратегия обеспечивает устойчивое функционирование популяций рыжей полевки, обитающей в условиях естественной геохимической аномалии с избыточным содержанием кобальта, никеля и хрома в течение многих поколений.
Список литературы
Авцын А.П., Жаворонков А.А., Риш М.А., Строчкова Л.С. Микроэлементозы человека: этиология, классификация, органопатология. М.: Медицина. 1991. 496 с.
Агаджанян Н.А., Скальный А.В. Химические элементы в среде обитания и экологический портрет человека. М.: Изд-во КМК. 2001. 83 с.
Артемьев Ю.Т., Окулова С.М. Методика полевого изучения эмбриональной смертности до имплантации у грызунов // Микроэволюция. 1981. С. 64–74.
Башенина Н.В. Пути адаптации мышевидных грызунов. М.: Наука. 1977. 353 с.
Башкин В.Н., Касимов Н.С. Биогеохимия. М.: Научный мир. 2004. 648 с.
Байтимирова Е.А., Михеева Е.В. Тяжелые металлы в печени животных, обитающих на территории естественной геохимической аномалии // Естествознание и гуманизм: сб. науч. работ. Томск, 2005. Т. 2. № 4. С. 36.
Бессалова Е.Ю. Оригинальная классификация фолликулов, адаптированная для экспериментального изучения яичников крупных млекопитающих // Сб. тр. Крым. гос. мед. ун-та им. С.И. Георгиевского. 2006. Т. 142. Ч. 1. С. 4–5.
Вернадский В.И. Проблемы биогеохимии. Тр. биогеохим. лаб. Ин-та геохимии и аналит. химии. М.: Наука. 1980. Т. 16. 226 с.
Владимирова Е.Г., Черниговская Е.В., Данилова О.А. Гипоталамо-гипофизарная нейросекреторная система красной полевки (Cletrionomys rutilus) в ходе популяционного цикла // Журн. Эвол. Биох. Физиол. 2006. Т. 32. Вып. 3. С. 308–317.
Волкова О.В., Елецкий Ю.К. Основы гистологии. М.: Медицина. 1982. 304 с.
Гашев Ю.П. Загрязнение окружающей среды и экологическая обусловленность патологии человека. Аналит. обзор. НЦКиЭМ СО РАН. Сер. Экология. Новосибирск, 2003. Вып. 68. 137 с.
Европейская рыжая полевка / Отв.ред. Башенина Н.В. М.: Наука. 1981. 351 с.
Жигальский О.А., Бернштейн А.Д. Популяционные факторы регуляции размножения рыжей полевки (Clethrionomys glareolus Schreb.) // Докл. АН СССР. 1986. Т. 291. № 1. С. 250–252.
Жигальский О.А., Бернштейн А.Д. Анализ факторов, определяющих численность и структуру населения рыжей полевки // Докл. АН СССР. 1989. Т. 305. № 6. С. 1509–1511.
Жигальский О.А. Зональные и биотопические особенности влияния эндо- и экзогенных факторов на население рыжей полевки (Clethrionomus glareolus Schreber, 1780). Экология. 1994. № 3. С. 50–60.
Жигальский О.А., Кшнясев И.А. Популяционные циклы европейской рыжей полевки в оптимуме ареала // Экология. 2000. № 5. С. 383–390.
Жигальский О.А., Мамина В.П. Плотностно-зависимые механизмы регуляции полового созревания самцов лесных полевок // Экология. 2015. № 6. С. 473–475.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука. 1975. 246 с.
Ивантер Э.В. Популяционные факторы динамики численности рыжей полевки (Cletryonomys glareolus) на северном пределе ареала // Тр. карельского науч. центра РАН. 2005. № 7. С. 48–63.
Кошкина Т.В. Метод определения возраста рыжих полевок и опыт его применения. Зоол. журн. 1955. Т. 39. Вып. 3. С. 631–639.
Кшнясев И.А., Давыдова Ю.А. Динамика плотности и структуры популяций лесных полевок в южной тайге. Вест. Нижегородского унив-та. Н. Новгород, 2005. Вып. 1. № 9. С. 113–123.
Лесорастительные условия и типы лесов Свердловской области Практ. руководство / Под ред. Колесникова Б.П. Свердловск: УНЦ АН СССР. 1973. 176 с.
Майер Э. Зоологический вид и эволюция. М.: Мир. 1968. 597 с.
Мамина В.П. Механизмы регуляции сперматогенеза у лесных полевок при разных фазах численности популяции Известия РАН. Сер. биологическая. 2016. № 3. С. 328–334.
Методические рекомендации “Методы экспериментального исследования по установлению порогов действия промышленных ядов на генеративную функцию с целью гигиенического нормирования” М., 1978.
Михеева Е.В., Жигальский О.А., Мамина В.П. Тяжелые металлы в системе почва – растение – животное в районе естественной геохимической аномалии // Экология. 2003. № 4. С. 318–320.
Михеева Е.В., Жигальский О.А., Мамина В.П., Байтимирова Е.А. Адаптация рыжей полевки к условиям природной биогеохимической провинции с избыточным содержанием никеля, кобальта и хрома // Журн. общ. биологии. 2006. Т. 67. № 3. С. 212–221.
Михеева Е.В. Морфофункциональные особенности надпочечника и щитовидной железы рыжей полевки на территории природной биогеохимической провинции: автореф. дис. канд. биол. наук Екатеринбург: ИЭРиЖ РАН, 2006. 26 с.
Молнар Е. Общая сперматология. Изд-во АН Венгрии. Будапешт. 1969. 294 с.
Науменко Е.В., Осадчук А.В., Серова Л.И., Шишкина Г.Т. Генетико-физиологические механизмы регуляции функций семенников. Изд-во “Наука” Сибирское отд. Новосибирск, 1983. 203 с.
Наумов Н.П. Очерки сравнительной экологии мышевидных грызунов М.: Л. Изд-во АН СССР, 1948. 203 с.
Петров И.А., Дмитриева М.Л., Тихоновская О.А., Петрова М.С., Логвинов С.И. Тканевые и молекулярные основы фолликулогенеза. Механизмы раннего фолликулярного роста // Проблемы репродукции. 2017. Т. 23. № 5. С. 33–41
Принципы сравнительной оценки радиационного и химического факторов / Под ред. Рамзаева П.В. М.: Энергоатомиздат. 1984. 232 с.
Райцина С.С. Сперматогенез и структурные основы его регуляции. М.: Наука, 1985. 206 с.
Роговин К.А., Мошкин М.П. Авторегуляция численности в популяциях млекопитающих и стресс (Штрихи к давно написанной картине) // Журн. общ. биологии. 2007. Т. 68. № 4. С. 244–267.
Сюзюмова Л.М., Карагезян А.Р. Особенности экологии кустарниковой полевки и лесной мыши в Анкаване // Техногенные элементы и животный организм. Свердловск, 1986. С. 14–22.
Тупикова Н.В. Изучение размножения и возрастного состава популяции мелких млекопитающих // Мет. изучения прир. очагов болезней человека. М.: Медицина, 1964. С. 154–192.
Тупикова Н.В., Сидорова Г.А., Коновалова Э.А. Опр. возраста лесных полевок. Материалы к познанию фауны и флоры СССР. 1970. Вып. 75, № 60. С. 160–167.
Ухов Ю.И., Астраханцев А.Ф. Морфометрические методы в оценке функционального состояния семенников. // Арх. анат. 1983. Т. 84. Вып. 3. С. 66–69.
Шевлюк Н.Н., Руди В.Н., Стадников А.А. Биология размножения наземных грызунов из семейства беличьих. Екатеринбург: Изд-во УрО РАН, 1999. 145 с.
Bouters R., Esnault C., Ortavant R., Salisbury G.W. Comparison of DNA revealed by Feulgen and by ultraviolet light in rabbit spermatozoa after storage in the male efferent ducts // Nature. 1967. V. 213. P. 181–182.
Bujalska G. Reproduction and mortality of bank voles and the changes in the size of an island population. Acta Theriol. 1975. V. 20. P. 41–56.
Bujalska G. Reproduction. Ecology of the Bank Vole // Acta Theriol. 1983. V. 28. P. 148–161.
Bujalska G. Social system of the bank vole, Clethrionomys glareolus. in: Social systems and population cycles in voles (Eds. R.H. Tamarin, R.S. Ostfeld, S.R. Pugh, G. Bujalska). Birkhauser Verlag: BaseL. 1990. P. 155–167.
Bujalska G., Gerlinskaya L., Grika L., Evsikov V., Moshkin M. Adrenocortical variability in life history of bank voles // Pol. Ecol. Studies. 1994. V. 7. P. 229–243.
Bröker L. E., Frank A.E. Kruyt and giuseppe giaccone. Cell death independent of caspases: a review // Clin. Can. Res. 2005. V. 11. Issue 9. P. 3155–3162.
Christian J.J., Mayer W., Gelder R. Endocrine adaptive mechanisms and the physiologic regulation of population growth // J. Physiol. Mammal. 1963. V. 1. P. 189–353.
Christian J.J., Davis D.E. Endocrines, behavior and population // Science. 1964. V. 146. P. 1550–1560.
Christian J.J., Lloyd J.A., Davis D.E. The role of endocrine in the self-regulation of mammalian populations in the self-regulation of mammalian populations // Rec. Progr. Hormone Res. N.Y.L. 1965. V. 21. P. 501.
Christian, J.J. Endocrine factors in population regulation, in biosocial mechanisms of population regulation. New Haven: Yale Univ. Press, 1980. P. 55–115.
Connell C.J., Connell G.M. The interstitial tissue of the testis // The testis / Eds Johnson A.D., Gomes W.R., Vandemark N.L. N.Y.: Acad. Press, 1977. V. 4. P. 333–361.
Gougeon A. Regulation of ovarian follicular development in primates: facts and hypotheses // Endocrin Rev. 1996. № 17. P. 121–155.
Hansson L., Henttonen H. Regional differences in cyclicity and reproduction in Clethrionomys species: are they related? // Ann. Zool. Fenn. 1985. V. 22. № 3. P. 277–288.
Huleihel M., Lunenfeld E. Regulation of spermatogenesis by paracrine/autocrine testicular factors // Asian J. Androl. 2004. V. 6. P. 259–268.
Ivanter E.V., Osipova O.V. Population dynamics of the bank volt in the eastern part of its distribution range // Pol. J. Ecol. 2000. V. 48. P. 179–195.
McLachlan R.I., Wreford N.G., Meachem S.J., De Kretser D.M., Robertson D.M. Effects of testosterone on spermatogenic cell populations in the adult rat // Biol. Reprod. 1994. V. 51. P. 945–955.
McGee E., Hsueh A. Initial and cyclic recruitment of ovarian follicles // Endocrin Rev. 2000. № 21. P. 200–214.
Novikov E.A., Panov V.V., Moshkin M.P. Density-dependent regulation in populations of northern red-backed voles (Myodes rutilus) in optimal and suboptimal habitats of Southwest Siberia // Biol. Bull. Rev. 2012. V. 2. № 5. P. 431–438.
Novikov E., Moshkin M. Sexual maturation, adrenocortical function and population density of red-backed vole, Clethrionomys rutilus (Pall.) // Mammalia. 1998. V. 62. P. 529–540.
O’Donnell L., McLachlan R.I., Wreford N.G., Robertson D.M. Testosterone promotes the conversion of round spermatids between stages VII and VIII of the rat spermatogenic cycle // Endocrinology. 1994. V. 135. P. 2608–2614.
Oravec S., Hlavacka S. Disorders of thyroid function and fertility disorders // Ceska Gynecol. 2000. V. 65. № 1. P. 53–57.
Pasquale E., Beck-Peccoz P., Persani L. Hypergonadotropic ovarian failure associated with an inherited mutation of human bone morphogenetic protein-15 (BMP15) gene // Amer. J. Hum. Gen. 2004. V. 75. № 1. P. 106–111.
Rich K.A., de Kretser D.M. Effect of differing degress of destruction of the rat seminiferous epithelium on levels of FSH and androgen binding protein // Endocrinology. 1977. V. 101. P. 959–974
Sailer B.L., Jost L.K., Evenson D.P. Bull sperm head morphometry related to abnormal chromatin structure and fertility // Cytometry 1996. V. 24. P.167–173.
Sun Y.T., Irby, D.C., Robertson D.M., de Kretser D.M. The effects of exogenously administered testosterone on spermatogenesis in intact and hypophysectomised rats // Endocrinology. 1989. V. 125. P. 2141–2149.
Sun Y.T., Wreford N.G., Robertson D.M., de Kretser D.M. Quantitative cytological studies of spermatogenesis in intact and hypophysectomized rats: identification of androgen-dependent stages // Endocrinology. 1990. V. 127. P. 1215–1223.
Tilly J.L, Pru J.K, Rueda B.R. Role apoptosis in development, functioning and infringement of function ovary // The Ovary. 2004. V. 19. P. 321–345.
Wakim A.N., Polizotto S.L., Burholt D.R. Influense of thyroxine on human granulosa cell steroidogenesis in vitro // J. Assist. Reprod. Genet. 1995. V. 12. № 4. P. 274–277.
Walker W.H., Cheng J. FSH and testosterone signaling in Sertoli cells // Reproduction. 2005. V. 130. P. 15–28.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая