

Известия РАН. Серия биологическая, 2021, № 6, стр. 602-614
Генетическая изменчивость и потенциальный ареал Darevskia rostombekowi в Закавказье
Ф. А. Осипов 1, *, А. А. Вергун 2, 3, М. С. Аракелян 4, Р. К. Петросян 4, Н. Н. Дергунова 1, Л. А. Неймарк 1, В. Г. Петросян 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский пр., 33, Россия
2 Институт биологии гена РАН
119334 Москва, ул. Вавилова 34/5, Россия
3 Московский педагогический государственный университет
119991 Москва, Малая Пироговская ул. 1/1, Россия
4 Биологический факультет Ереванского государственного университета
0025 Ереван, ул. А. Манкуяна, 1, Армения
* E-mail: osipov_feodor@mail.ru
Поступила в редакцию 26.08.2019
После доработки 11.11.2019
Принята к публикации 11.11.2019
Аннотация
Представлены результаты анализа генетической изменчивости партеногенетического вида Darevskia rostombekowi (Darevsky, 1957) с использованием четырех микросателлит-содержащих локусов. На основе 118 записей с географическими координатами присутствия данного вида в Закавказье были созданы карты потенциального ареала. Анализ генетической структуры популяций показал, что, несмотря на установленную мультиклональность (7 клональных линий в 4 популяциях), D. rostombekowi образован в результате единичного акта гибридизации между близкими двуполыми видами. Прогнозируемое распределение D. rostombekowi с помощью созданного потенциального ареала выявило новые небольшие местообитания, где присутствие вида ранее не сообщалось. Результаты этого исследования и отсутствие множественных актов гибридизации этих форм могут указывать на регрессию численности. Следовательно, оценка статуса сохранения партеногенетического вида представляется оправданной.
За последние 50 лет получены результаты цитогенетического, молекулярно-генетического анализов однополых и двуполых комплексов видов рода Darevskia (Даревский, 1967; Uzzell, Darevsky, 1975; Мартиросян и др., 2002; Малышева и др., 2006; Осипов и др., 2016; Ryskov et al., 2017). Эти результаты указывают на концепцию гибридогенного видообразования у партеногенетических видов скальных ящериц, состоящую в формировании новых однополых гибридных видов, несущих в себе геномы, унаследованные от двух близкородственных родительских видов (Uzzell, Darevsky, 1975; Ryskov et al., 2017). Проведенный комплексный анализ показал, что партеногенетический вид D. rostombekowi образован в результате естественной гибридизации между близкими двуполыми видами D. portschinskii (отцовский вид) и D. raddei raddei (материнский вид) (Uzzell, Darevsky, 1975). Несмотря на определенные успехи исследования D. rostombekowi (MacCulloch et al., 1997; Мартиросян и др., 2002; Осипов и др., 2016; Ryskov et al., 2017), вопросы о границах его ареала и видоспецифических параметров абиотических факторов среды, определяющих область распространения изолированных популяций, остаются открытыми. Анализ аллозимных локусов особей различных популяций D. rostombekowi, за исключением изолированной популяции Цовак (Tsovak), не выявил изменчивости, что предположительно указывало на моноклональную генетическую структуру (MacCulloch et al., 1997).
Существуют фрагментарные данные по характеру распределения отдельных популяций на территории Армении. Известно, что ящерица D. rostombekowi (один из семи партеногенетически размножающихся видов скальных ящериц рода Darevskia) занимает сравнительно небольшой по площади ареал, состоящий из нескольких разных по величине изолированных популяций в пределах Северной Армении, прилежащих районах Северо-Западного Азербайджана, и оторванной от основного ареала, маленькой высокогорной реликтовой (~12 тыс. лет) популяции на юго-восточном побережье оз. Севан (MacCulloch et al., 1997; Мартиросян и др., 2002; Arakelyan et al., 2011, Petrosyan et al., 2020a). Границы ареала вида в целом, а также отдельных популяций в литературе отсутствуют.
Цель исследования: 1) определить генетическую структуру популяций, обитающих в разных экологических условиях на территории Армении; 2) построить карту пространственного распределения вида D. rostombekowi на основе данных находок; 3) выявить биоклиматические, топографические и ландшафтные переменные, определяющие потенциальный ареал вида.
МАТЕРИАЛЫ И МЕТОДЫ
Молекулярно-генетический анализ
Для исследования были отобраны образцы ДНК партеногенетического вида D. rostombekowi (n = 42) из четырех популяций на территории Армении (табл. 1). Ящерицы были отловлены в период с 1997 по 2006 гг. в их естественных местах обитания. Все полевые исследования проводили до момента включения вида D. rostombekowi в Красную книгу Армении как вид, “находящийся в опасном состоянии” в соответствии с критериями МСОП (Endangered, EN B1ab) (Agasyan et al., 2010). В работе анализ проводили на основе трех ранее описанных микросателлит-содержащих локусов Du215, Du281, Du323 (Ryskov et al., 2017) и нового Du47G. Данные локусы были установлены для исследования других партеногенетических видов данного рода: D. unisexualis (Badaeva et al., 2008), D. dahli (Vergun et al., 2014).
Таблица 1.
Список исследованных образцов ДНК
| Название популяции | Количество особей | Географические координаты |
|---|---|---|
| Гош | 4 | 40°42′20.3′′ N 45°00′57.7′′ E |
| Цовак | 8 | 40°10′45.0′′ N 45°37′22.7′′ E |
| Папанино | 21 | 40°42′27.7′′ N 44°45′43.8′′ E |
| Спитак | 9 | 40°48′50.0′′ N 44°16′48.7′′ E |
Постановку локус-специфической полимеразной цепной реакции проводили с использованием ранее подобранных пар праймеров и условий амплификации (Korchagin et al., 2007; Vergun et al., 2014).
Методика выделения ДНК, анализ микросателлит-содержащих локусов Du215, Du281, Du323, Du47G представлены в работах (Korchagin et al., 2007; Осипов и др., 2016; Ryskov et al., 2017). Полученные в результате секвенирования последовательности были депонированы в GenBank (KM573728–KM573762; HM014002–HM014003; KR559279KR559316; MK622868-MK622872).
Анализ и сопоставление полученных последовательностей проводили в программе MEGA v.7.0.21. Число аллелей, аллельное богатство как мера количества аллелей с поправкой на размер выборки и ожидаемая гетерозиготность как мера разнообразия генов были рассчитаны по каждому локусу и популяции с использованием Fstat v.2.9.3.2. GenePop v.4.2 и online версии POPTREEW. AMOVA-анализ проводили с использованием пакета Poppr (Kamvar et al., 2014). Реализацию множественных выравниваний последовательностей микросателлитных аллелей трех видов производили с помощью алгоритма MUSCLE (Edgar RC. et al., 2004). Эти матрицы были использованы для строительства сети методом ближайшего соседа с помощью MEGA v.7.0.21 (Tamura et al., 2013). Составление генотипов производили путем сравнения и анализа комбинаций аллельных вариантов в популяциях с помощью программы LabConverter V1. (Омельченко и др., 2016).
Создание карты потенциального ареала вида
Сбор полевых данных. Материалом для работы послужили данные, собранные соавторами во время полевых исследований в 1997–2019 гг., и коллекционные материалы кафедры зоологии Ереванского государственного университета (ЕРГУ), Зоологического музея МГУ им. М.В. Ломоносова (ZMMU), музея Зоологического института РАН (ZISP) и Канадского музея Royal Ontario Museum (ROM). Изучены потенциальные местообитания на маршрутах общей протяженностью 3000 км в 2018 г. и 1800 км в 2019 г. Учетные работы проводили в утренние солнечные безветренные часы. Для каждого локалитета географические координаты и высоты были определены с помощью GPS-навигатора Garmin Montana 680t (Garmin Corp., Olathe, KS, США) с погрешностью ±3.5 м. Ящерицы с каждого участка были отловлены и определены в полевых условиях с помощью определителя (Darevsky, 1967; Arakelyan et al., 2011).
Векторные слои мест находок. Для создания векторной базы данных (ВБД) в среде ArcGIS Desktop 10.4.1 использовали полевые, музейные и литературные данные (Даревский, 1967; Uzzell, Darevsky, 1975; MacCulloch et al., 1997; Мартиросян и др., 2002; Arakelyan et al., 2011; Ryskov et al., 2017).
Растровые слои для построения карт ареала. Был проведен обзор литературы для выбора важнейших переменных, определяющих распространение скальных ящериц (Darevsky, 1967; Uzzell, Darevsky, 1975; Kaliontzopoulou et al., 2008; Tarkhnishvili et al., 2010; Arakelyan et al., 2011; Doronin, 2015; Freitas et al., 2016, Petrosyan et al., 2019, 2020b). Выбранные экологические переменные характеризуют климат, топографию и земельный покров/землепользование. Пространственные климатические переменные были взяты из набора данных WorldClim 2.0 (http://worldclim.org/version2) c разрешением 30 s, включающие 21 биоклиматических переменных. В работе анализ проводили для трех различных разрешений (3, 9 и 30 arc second, т.е. ~90, ~270 и ~900 м соответственно) растровых слоев рельефа, климатических параметров и ландшафта. Все эти слои были подготовлены с помощью соответствующего программного модуля Arc GIS Desktop 10.4.1 с использованием кубической функции аппроксимации. В результате этих преобразований все растровые слои были представлены с разрешением ~90, ~270 и ~900 м соответственно.
Для проверки пространственной автокорреляции точек находок из ВБД использовали двухшаговую процедуру, описанную в работах (ESRI, 2017; Aiello-Lammens et al., 2015; Petrosyan et al., 2020b).
Потенциальные модели распределения D. rostombekowi были определены с помощью метода максимальной энтропии с использованием MaxEnt 3.4.1 (Phillips et al., 2006) из окружения EcoSpat (De Pous et al., 2011; Di Cola et al., 2017). Для построения модели распределения вида (SDM) использовали 10 запусков MaxEnt со случайным выбором тестовых и обучающих выборок. Во всех вариантах запуска MaxEnt 80% записей находок использовали как обучающие выборки, а 20% записей – как тестовые выборки. Мы использовали индекс Бойса (Bind) для оценки пригодности моделей с помощью пакета EcoSpat (Di Cola et al., 2017). Средний показатель индекса Бойса определяли усреднением индексов 10 моделей для каждого разрешения в отдельности. Важность каждой предикторной переменной определяли, используя таблицу вкладов MaxEnt (Phillips et al., 2006). Важными переменными считались те, которые вносят существенный вклад в модель, т.е. имеют высокие значения перестановочной важности (PI) или высокие значения процента вклада (PC) (Phillips et al., 2006).
Сходство экологических ниш. Сравнительный анализ экологических ниш популяций D. rostombekowi, полученных для различных наборов данных проводили с использованием общей концепции, представленной в работах (Petitpierre et al., 2012; Di Cola et al., 2017). Оценку перекрытия ниш, полученных на основе разных наборов точек присутствия, проводили с использованием индекса Schoener’s D (Di Cola et al., 2017).
Сравнительный анализ центроидов (средних значений переменных) ниш, определяющих пространственное расположение популяций D. rostombekowi, проводили с использованием процедуры общей линейной модели (GLM). В этой модели был использован однофакторный дисперсионный анализ (ANOVA). Если ANOVA показывал существенное различие центроидов, то с помощью методов множественного сравнения Post hoc Тьюки HSD определяли, какие именно центроиды отличались друг от друга.
Статистический анализ и построение диаграмм были выполнены в среде RStudio 1.1.463.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Генетическая характеристика популяций. Анализ четырех микросателлит-содержаших локусов Du215, Du281, Du323 и Du47G выявил, что все исследованные особи D. rostombekowi являются гетерозиготными по данным локусам. В исследованных популяциях было выявлено от 2 до 5 аллелей в зависимости от локуса. Аллельные варианты каждого локуса представлены в табл. 2.
Таблица 2.
Структура аллельных вариантов микросателлитных локусов Du215, Du281, Du323 и Du47G партеногенетического вида D. rostombekowi
| Аллель | Длина (п.о.) | Последовательность микросателлитного кластера | Фиксированные нуклеотидные замены на флангах* | Gene Bank ac. № |
|---|---|---|---|---|
| Du215 (rost)1 | 256 | 5' (GAT)(GATA)10(GCAA)11 3' | – | GU972528 |
| Du215 (rost)2 | 252 | 5' (GAT)(GATA)9(GCAA)11 3' | – | GU972529 |
| Du215 (rost)3 | 250 | 5' (GAT)(GATA)9 (GCAA)10 3' | TT (–98/99) | GU972530 |
| Du215 (rost)4 | 248 | 5' (GAT)(GATA)8 (GCAA)11 3' | – | GU972531 |
| Du215 (rost)5 | 227 | 5' (GAT)(GATA)4(GAT)(GATA)7(GCAA)2 3' | – | GU972532 |
| Du281 (rost)1 | 265 | 5' (GATA)2(GAGAT)(GATA)11(GAT)(GATA)12(TA)(GATA) 3' | T (–84), A (–19), T (+15), A (+25) | HM114225 |
| Du281 (rost)2 | 261 | 5' (GATA)2(GAGAT)(GATA)10(GAT)(GATA)12(TA)(GATA) 3' | T (–84), A (–19), T (+15), A (+25) | HM070256 |
| Du281 (rost)3 | 253 | 5' (GATA)2(GAGAT)(GATA)11(GAT)(GATA)9 (TA)(GATA) 3' | T (–84), A (–19), T (+15), A (+25) | HM070257 |
| Du281 (rost)4 | 191 | 5' (GATA)10 3' | C (–84), G (–19), C (+15), G (+25) | HM070258 |
| Du323 (rost)1 | 195 | 5' (AC)6…(GATA)6(GAT)(GATA)2…(GA)4 3' | C (–23), T (+39) | HM013995 |
| Du323 (rost)2 | 184 | 5' (AC)4…(GATA) (GGT)(GATA)3(GAT)(GATA)…(GA)4 3' | A (–23), C (+39) | HM013996 |
| Du47G (rost)1 | 180 | 5' (GATA)8(GACA)4(GATA)2(GAT)(GATA)2 3' | A(+7), A(+21), G(+52) | MK622868 |
| Du47G (rost)2 | 168 | 5' (GATA)9(GACA)(GATA)(GAT)(GATA)2 3' | T(+7), T(+21), C(+52) | MK622869 |
| Du47G (rost)3 | 168 | 5' (GATA)8(GACA)(GATA)2(GAT)(GATA)2 3' | T(+7), T(+21), C(+52) | MK622870 |
| Du47G (rost)4 | 164 | 5' (GATA)8(GACA)(GATA)(GAT)(GATA)2 3' | T(+7), T(+21), C(+52) | MK622871 |
| Du47G (rost)5 | 152 | 5'(GATA)2(GACA)(GATA)4(GAT)(GATA)2 3' | A(+7), A(+21), G(+52) | MK622872 |
Структурные различия микросателлитных локусов Du215, Du281 и D323 ранее были описаны (Осипов и др., 2016; Ryskov et al., 2017). Анализ локуса Du47G позволил выявить 5 аллельных вариантов, которые различались структурой микросателлитного кластера, для которого характерны GATA- и GACA-повторы, а так же несовершенные звенья GAT. Кроме того аллельные варианты различались по фиксированным точечным заменам на флангах, по которым их можно разделить на две группы: 1) A (+7), A (+21), G (+52) T (+58), характерная для аллелей Du47G(rost)1 и Du47G(rost)5; 2) T (+7), T (+21), C (+52) T (+58), характерная для аллелей Du47G(rost)2-Du47G(rost)4 (табл. 2).
Количество генотипов было установлено по анализу сочетания аллельных вариантов каждой особи (табл. 3, рис. 1). Анализ аллельных комбинаций по 4 локусам выявил 7 генотипов, неравномерно распространенных в исследуемых популяциях. Особи с одинаковыми генотипами формировали клональные линии. Мажорный клон (генотип 1), был установлен у 24 особей (57% от всех исследованных особей) и представлен в трех популяциях – Гош, Папанино (окрестности г. Дилижан), Спитак. Клон с генотипом 2 встречался у 7 особей (17%). Клон с генотипами 3 (5 особей – 12%) и 4 (3 особи – 7%) были обнаружены только в изолированной севанской популяции Цовак. Остальные генотипы (5, 6 и 7) были представлены только одной особью (2%) и встречались в популяциях Гош, Папанино, Спитак.
Таблица 3.
Генотипическое разнообразие в популяциях D. rostombekowi по четырем микросателлитным локусам
| Генотип | Комбинация аллельных вариантов | Популяции | Количество особей с данным генотипом | |||
|---|---|---|---|---|---|---|
| Гош | Цовак | Папанино | Спитак | |||
| 1 | Du215(2 + 5) + Du281(1 + 4) + Du323(1 + 2) + Du47G(4 + 5) | 2 | 0 | 14 | 8 | 24 |
| 2 | Du215(2 + 5) + Du281(2 + 4) + Du323(1 + 2) + Du47G(4 + 5) | 0 | 0 | 6 | 1 | 7 |
| 3 | Du215(4 + 5) + Du281(3 + 4) + Du323(1 + 2) + Du47G(2 + 1) | 0 | 5 | 0 | 0 | 5 |
| 4 | Du215(4 + 5) + Du281(3 + 4) + Du323(1 + 2) + Du47G(4 + 5) | 0 | 3 | 0 | 0 | 3 |
| 5 | Du215(2 + 5) + Du281(2 + 4) + Du323(1 + 2) + Du47G(3 + 5) | 0 | 0 | 1 | 0 | 1 |
| 6 | Du215(3 + 5) + Du281(2 + 4) + Du323(1 + 2) + Du47G(4 + 5) | 1 | 0 | 0 | 0 | 1 |
| 7 | Du215(1 + 5) + Du281(1 + 4) + Du323(1 + 2) + Du47G(4 + 5) | 1 | 0 | 0 | 0 | 1 |
| Общее количество особей | 4 | 8 | 21 | 9 | 42 | |
| Генотипическое разнообразие | 3 | 2 | 3 | 2 | ||
Таблица 4.
Показатели генетического разнообразия в популяциях D. rostombekowi по четырем микросателлитным локусам
| Локус | Популяция | N | Rs | HE |
|---|---|---|---|---|
| Du215 | Гош | 4 | 4.000 | 0.750 |
| Цовак | 2 | 2.000 | 0.533 | |
| Папанино | 2 | 1.997 | 0.512 | |
| Спитак | 2 | 2.000 | 0.529 | |
| ∑ | 5 | 2.738 | 0.581 | |
| Du281 | Гош | 3 | 3.000 | 0.679 |
| Цовак | 2 | 2.000 | 0.533 | |
| Папанино | 3 | 2.773 | 0.626 | |
| Спитак | 3 | 2.443 | 0.582 | |
| ∑ | 4 | 3.127 | 0.605 | |
| Du323 | Гош | 2 | 2.000 | 0.571 |
| Цовак | 2 | 2.000 | 0.533 | |
| Папанино | 2 | 1.997 | 0.512 | |
| Спитак | 2 | 2.000 | 0.529 | |
| ∑ | 2 | 1.995 | 0.536 | |
| Du47G | Гош | 2 | 2.000 | 0.571 |
| Цовак | 4 | 3.774 | 0.783 | |
| Папанино | 3 | 2.186 | 0.535 | |
| Спитак | 2 | 2.000 | 0.529 | |
| ∑ | 5 | 2.883 | 0.604 | |
| Все локусы | 16 | 2.685 | 0.581 |
Таблица 5.
Наборы предикторных переменных, определяющих модели распределения D. rostombekowi с указанием индексов пригодности AUC и Бойса для карт потенциальных ареалов, полученных с помощью MaxEnt и EcoSpat. Представлены только переменные, которые внесли вклад больше 5% либо по PC или по PI
| Переменная* | Полный набор точек находок | Сокращенный набор точек находок | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Разрешение | ||||||||||||
| 90 м | 270 м | 900 м | 90 м | 270 м | 900 м | |||||||
| PC | PI | PC | PI | PC | PI | PC | PI | PC | PI | PC | PI | |
| MeanTempDrQ | 20.1 | 31.5 | 19.6 | 35.4 | 20.3 | 31.6 | 29 | 66.8 | 30 | 63.7 | 27.6 | 60 |
| HighWay | 17.6 | 1.2 | 12.9 | 0.5 | 9.7 | 1.4 | 6.2 | 0.9 | 4.6 | 0 | 4.6 | 0.1 |
| Sradmean | 15.7 | 17.9 | 1.2 | 6.4 | 4.3 | 9.4 | 16.2 | 5.5 | 0.7 | 6.1 | 3 | 6.8 |
| PrecipCoefVar | 11.2 | 17.3 | 19.3 | 21.7 | 21.2 | 23.4 | 19.6 | 13.5 | 21.4 | 12.1 | 22.3 | 16.8 |
| IsoTerm | 9.8 | 3.3 | 19.3 | 9 | 21.7 | 16.1 | 8.2 | 4.8 | 18.2 | 3.5 | 18.6 | 5.8 |
| PrecWarmQ | 7.9 | 3.5 | 15.8 | 6.2 | 14.3 | 7.7 | 9.8 | 2.6 | 14.3 | 3.4 | 15.2 | 3.5 |
| StdDivTemp | 4.2 | 18.3 | 3.7 | 18.6 | 1.7 | 5.0 | 4.9 | 4.9 | 5.2 | 9.5 | 4.2 | 5.8 |
| Elevation | 4.7 | 5.7 | 1.5 | 1.6 | 1 | 0.8 | 0.2 | 1 | 0.2 | 1 | 0.4 | 0.4 |
| Индекс AUC | 0.979 ± 0.011 | 0.98 ± 0.01 | 0.978 ± 0.007 | 0.979 ± 0.009 | 0.982 ± 0.007 | 0.985 ± 0.007 | ||||||
| Индекс Бойса | 0.958 ± 0.004 | 0.926 ± 0.006 | 0.916 ± 0.003 | 0.814 ± 0.009 | 0.813 ± 0.02 | 0.939 ± 0.003 | ||||||
Примечание. * MeanTempDrQ – средняя температура сухой четверти года (°С), HighWay – расстояние до дорог (м), Sradmean – средняя солнечная радиация (КДж m-2 день-1), PrecipCoefVar – коэффициент вариации осадков (%), IsoTerm – изотермичность (%), PrecWarmQ – сумма осадков в самой теплой четверти года (mm), StdDivTemp – стандартное отклонение температур (°С), Elevation – высота над уровнем моря (м).
Рис. 1.
Схематическое изображение семи генотипов, образованных аллельными комбинациями микросателлитных локусов Du215, Du281, Du323 и Du47G у 42 особей D. rostombekowi. Специфичные для родительских видов маркеры SNV показаны в квадратах. Вариабельные микросателлитные кластеры показаны для каждого аллеля.
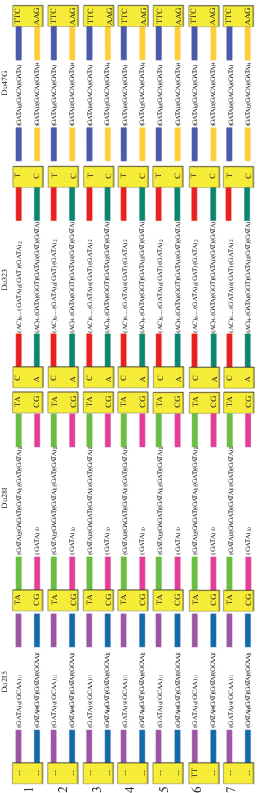
Генотипы 1–7 отличаются друг от друга последовательностью микросателлитного кластера. Редкие генотипы, обнаруженные у единичных особей в популяциях, вероятно являются мутационными производными основного (представленного большим числом особей и распространненого в 3 из 4 исследованных популяциях) генотипа. Генотипы 3 и 4 представлены только в изолированной популяции Цовак и являются самыми отдаленными от основного генотипа. Анализ пространственно-частотных распределений наиболее широко распространенных генотипов 1 и 2 и популяционно-специфических генотипов 3 и 4 выявил зависимость между частотами клонов и географическим разделением между тремя северными популяциями и севанской популяцией Цовак.
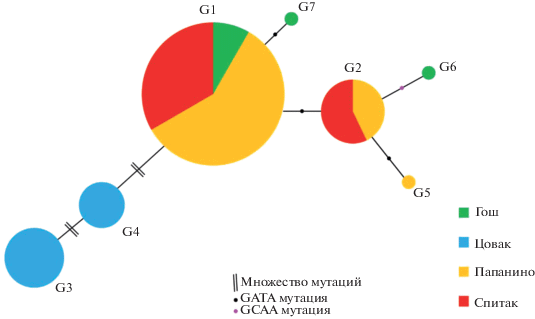
Принимая генотип 1(G1) за исходный, согласно модели Паркера (Parker et al., 1989), мы можем считать, что остальные обнаруженные генотипы могли быть образованы в результате микросателлитных мутаций уже сформировавшегося основного клона. Это событие отражено на схеме (рис. 2), которая построена на основе сравнительного анализа сочетаний аллельных вариантов по каждому локусу с учетом распространения генотипа и частоты его встречаемости в популяциях. Как видно их схемы, генотипы G2, G7 отличаются от генотипа G1 одной GATA- мутацией в микросателлитном кластере по одному из 4 локусов (G1 отличается от G7 одной мутацией в локусе Du215, G1 от G2 – одной мутацией в локусе Du281). Такая же картина наблюдается между генотипами G2 и G5. Генотип G6 хоть и представлен в популяции Гош, однако имеет сходство с генотипом G2 (представленным в популяциях Спитак и Папанино) за исключением делеции одного GCAA звена в локусе Du215. Несмотря на то, что генотипы G3 и G4 не были обнаружены в других популяциях, кроме Цовак; они связаны с основным генотипом G1 общими аллельными вариантами, однако сильно “удалены” от него множественными изменениями внутри микросателлитного кластера.
Рис. 2.
Схематическое изображение, отражающее взаимоотношения между генотипами 1–7(G1–G7) в исследованных популяциях D. rostombekowi. Размер кругов соответствует представленности генотипа в популяции.
В табл. 4 приведены данные расчетов индексов генетического разнообразия в популяциях партеновида по четырем микросателлитным локусам. Уровень ожидаемой гетерозиготности (HE ) варьируется от 0.533 до 0.783 (в среднем, 0.581), а число аллелей (N) от 2 до 4. Показатель степени аллельного богатства (Rs) варьируется от 1.995 до 4.000 (в среднем, 2.685). Самый высокий показатель данного значения установлен в популяции Гош по локусам Du215, Du281 и Du323.
Важнейшие предикторные переменные среды и потенциальный ареал партеногенетического вида. Мы собрали 118 записей с географическими координатами присутствия D. rostombekowi на территории Армении и Азербайджана (49 литературных данных и 69 собственных полевых записей). После удаления кластерных точек остались 56 записей, включенных в сокращенный набор для дальнейшего анализа. Эти записи в дальнейшем (полные: n = 118, сокращенные n = 56) использовали для построения моделей распределения видов и сравнения экологических ниш для трех разрешений ~90, ~270 и ~900 м соответственно.
Мы получили “высокие” значения индекса Бойса (Bind ± SD) и AUC (±SD) для всех моделей распределения вида. Вклады переменных для различных SDM моделей представлены в табл. 5 . Из таблицы следует, что индекс AUC (Phillips et al., 2006) имеет достаточно высокие значения и варьирует от 0.978 (±0.007) до 0.985 (±0.007), которые слабо различаются друг от друга. Последнее означает, что вне зависимости от типа использования данных (сокращенных, полных) и пространственного разрешения слоев, индекс AUC пригодности модели меняется слабо (табл. 5 ). В отличие от AUC, индекс Бойса чувствителен и позволяет выделять наилучшую модель. В нашем случае наиболее высокая точность модели распределения вида наблюдается только для одного случая, т.е. когда используются полные данные с пространственным разрешением ~90 м. В этом случае индекс Бойса составляет 0.958 ± 0.004. Наихудшая модель по индексу Бойса 0.813 (±0.02) построена при использовании сокращенных данных с пространственным разрешением ~270 м. Карты распределения вида для этих случаев представлены в рис. 3.
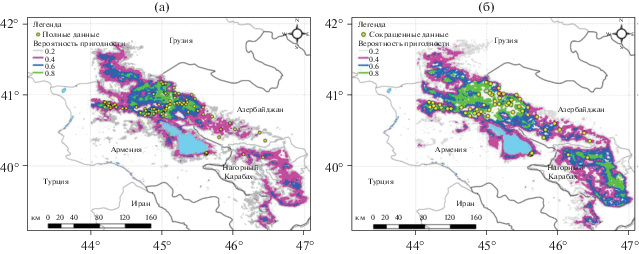
Рис. 3.
Карты распределения D. rostombekowi с использованием разных наборов данных и разрешений. (а) Полный набор точек находок и пространственное разрешение 90 м, (б) сокращенный набор данных и пространственное разрешение 270 м.
Из табл. 5 следует, что из 8 предикторных переменных только пять (MeanTempDrQ, Sradmean, PrecipCoefVar, IsoTerm, PrecWarmQ) общие для всех 60 моделей (10 репликации × 2 типа набора данных × 3 разрешения). Важность переменной StdDivTemp выделяется почти для всех моделей за исключением моделей, полученных для сокращенных данных для разрешения ~90 м. Важность переменной HighWay, выделяется для всех моделей с разным разрешением при использовании полного набора данных точек находок, а также для моделей с разрешением ~90 м при использовании сокращенных данных. Единственная переменная, которая выделяется только для одного сочетания набора данных (полных) и разрешения (~90 м), является высота над уровнем моря (Elevation).
Визуальный качественный анализ наилучшей (рис. 3а) и наихудшей (рис. 3б) карт показывает, что они, в целом, совпадают в общих чертах по границам ареала, но существует большое различие в деталях. Наилучшая карта (рис. 3а) указывает, что наиболее пригодные местообитания (с вероятностью >0.6) в основном расположены на севере Армении, включая все известные местообитания популяций (Севан, Спитак, Гош, Дилижан, Иджеван-Ноемберян), а также новые небольшие местообитания западной части оз. Севан и в восточной части Нагорного-Карабаха, северной части Армении на границе с Грузией. Наихудшая карта показывает обширные пригодные местообитания на западной части оз. Севана, обширную зону на восточной части Нагорного Карабаха, в северной Армении на границе с Грузией и участки в южной части Грузии.
Индексы сходства экологических ниш популяций партеногенетического вида. Результаты анализа перекрывания ниш для разных наборов данных представлены на рис. 4. Показано, что индекс сходства экологических ниш для разных разрешений изменяется в диапазоне от 86 до 90%, в зависимости от разрешения используемых слоев. Доля общих местообитаний, т.е. перекрытие ниш варьируется от 97.8 до 98.8%. Тест сходства экологических ниш, полученных на основе разных наборов данных, не отклоняется для всех разрешений (P < 0.02).
Рис. 4.
Экологические ниши D. rostombekowi (а), (б), (в), представленные с использованием двух осей главных компонент, и сдвиги центроидов ниш (г), (д), (е) по высоте, полученных на основе сокращенных точек находок относительно полных. Затемненные области показывают реализованные ниши для разных наборов точек и разрешений, пунктирная и сплошная линии на рис. (а), (б), (в) – 90, 100% площади района исследования.
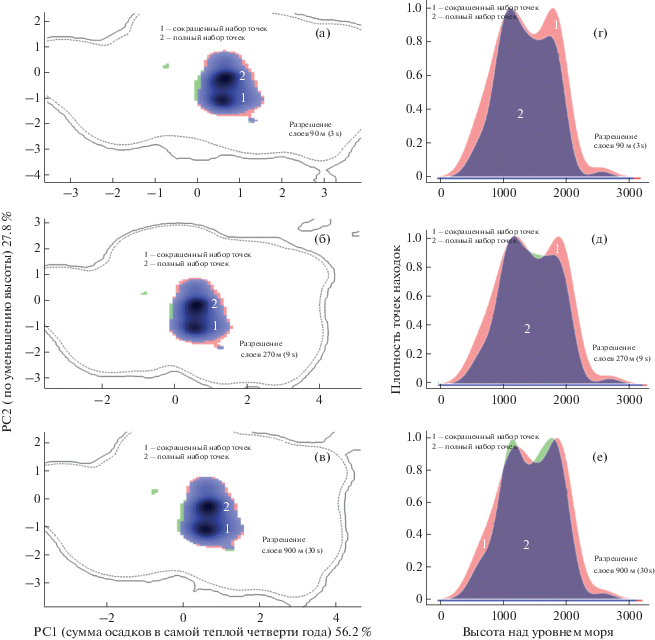
Несмотря на высокое сходство экологических ниш при использовании разных наборов данных, тем не менее, центроид по высоте смещается вверх для сокращенных данных (рис. 4г, 4д, 4е). Это означает, что разреживание с целью удаления кластеризованных точек приводит к значительному уменьшению количества точек в местообитаниях северных популяций (Спитак, Гош, Папанино, Иджеван-Ноемберян) и оставляет без изменения количества точек находок в южной части оз. Севан (Цовак). Это в итоге приводит к увеличению центроида высоты и увеличению пригодности обитания на высоте более 2000 м.
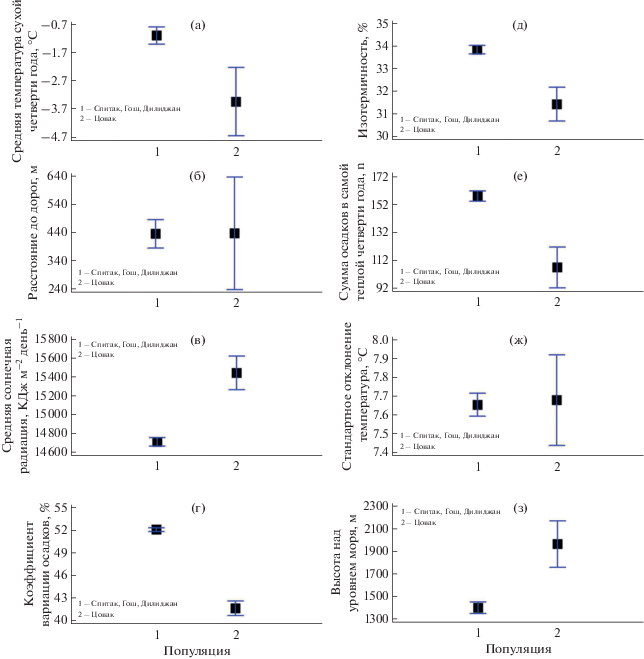
Сравнительный анализ центроидов ниш популяций партеногенетического вида. Для проверки статистической значимости различия центроидов ниш по абиотическим факторам среды разных популяций мы проводили их сравнительный анализ. Результаты сравнения средних значений предикторных переменных (центроидов ниш) объединенных северных популяций и изолированной популяции Цовак представлены на рис. 5. Из этого рисунка следует, что экологические условия популяции Цовак статистически значимо различаются от серверных популяций по средней температуре и суммарными осадками сухой четверти года, средней солнечной радиацией, коэффициентом вариации осадков, изотермичностью и высотой над уровнем моря. Южная популяция различается от северных не только по расстоянию до дорог, но и по стандартному отклонению температуры.
Рис. 5.
Сравнение средних значений предикторных переменных (центроидов) серверных популяций и изолированной популяции Цовак. На графике представлены средние значения с 95% доверительными интервалам Тьюки с использованием ANOVA GLM: (а) F = 6.9; P = 0.01; (б) F = 0.001; P = 0.99; (в) F = 30.95, P $ \ll $ 0.001; (г) F = 212.8, P $ \ll $ 0.001; (д) F = 18.98, P $ \ll $ 0.001; (е) F = 22.36, P $ \ll $ 0.001; (ж) F = 0.02, P = 0.89; (з) F = 13.9, P = 0.0003.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сравнительный анализ единичных нуклеотидных мутаций вне микросателлитного кластера (SNV) партеновида D. rostombekowi с родительскими видами (D. r. raddei, D. portschinskii) (Осипов и др., 2016, Ryskov et al., 2017) позволяет утверждать, что партеновид мог образоваться в результате единичного акта гибридизации между близкими бисексуальными особями родительских видов, поскольку исследованные особи партеновида не различались по SNV во всех четырех микросателлитных локусах (рис. 1).
Анализ распределений наиболее широко распространенных генотипов 1 и 2 и популяционно-специфических генотипов 3 и 4 выявил зависимость между частотами клонов и географическим разделением между тремя северными популяциями и севанской популяцией Цовак. Остается нерешенным вопрос, какой из 7 установленных клонов D. rostombekowi образовался первым. Предполагая, что у партеновида, образованного в результате одного гибридизационного события, имеется распространенный и чаще всего встречающийся “мажорный” клон с несколькими “минорными” клонами (Parker et al., 1989), то, вероятно, генотип G1 мог быть исходным. Остальные же генотипы являются мутациями G1. Мутации de novo во всех четырех исследованных локусах обусловлены делецией или вставкой микросателлитного повтора (рис. 1, табл. 2). Заметные различия генотипов G3 и G4 от основного генотипа G1 и обнаруженных в популяциях Гош, Папанино и Спитак генотипов G2, G5, G6, G7 могут быть связаны со значительными различиями в экологических условиях их местообитаний (рис. 5). Вероятно, другие (переходные клональные линии) между генотипами 1–3 и 4 могли быть утеряны в результате сокращения и/или разрыва ареала D. rostombekowi.
В работе показано, что SDM является важным инструментом, который можно использовать для выявления экологических требований популяций D. rostombekowi к среде и выбора наиболее важных переменных, определяющих их пространственное распределение. Дополнительный анализ с помощью метода ординации позволяет проводить сравнительный анализ экологических ниш различных популяций и определить меру их пересечения, сходства и выявить значимые сдвиги ниш при определении потенциальных ареалов на основе разных наборов данных точек находок и разрешений слоев предикторных переменных. Эти два подхода, а также генетическая структура популяций, позволяют выявить их особенности и оценить роль экологических факторов в формировании северных и южной популяций данного вида.
Важнейшие переменные, определяющие модель размещения партеногенетического вида. Как показали SDM средняя температура и суммарные осадки в сухой и теплой четверти года, солнечная радиация, коэффициент вариации осадков, изотермичность и высота над уровнем моря являются важными факторами среды, которые оказывают влияние на формирование пространственного распределения ящериц на территории Армении, Нагорного Карабаха и Азербайджана (рис. 3, табл. 5 ). Из этого следует, что набор переменных, определяющих пригодные местообитания, в значительной степени связаны с термическими условиями и влажностью в сухой и теплый период года соответственно, а также годовыми коэффициентами вариации этих переменных. Причем изотермичность должна быть в пределах 28–34%, а коэффициент влажности осадков 41–52% (рис. 4). Важно отметить, что изотермичность, отражающая достаточно высокую стабильность температуры и низкие ее колебания в местообитаниях, ранее не была отмечена для других видов ящериц, как важнейшая предикторная переменная (Kaliontzopoulou et al., 2008; De Pous et al., 2011). Узкий диапазон изменения этих переменных означает достаточно высокую чувствительность вида к изменению климата. Установленные переменные объясняют экологические особенности, связанные с распределением D. rostombekowi и дают представление о факторах, ограничивающих его распространение. Другая важная переменная – солнечная радиация, которая обеспечивает прогреваемость грунта в местообитаниях и меняется в узком диапазоне 4.1–4.2 кВт ⋅ ч/м2. Высота над уровнем моря является важной переменной только для одной – наилучшей модели, которая была получена с использованием слоев с разрешением 90 м (3 s) (табл. 5 ).
Потенциальный ареал партеногенетического вида. Точки находок и потенциальный ареал D. rostombekowi показывают, что вид распространен достаточно широко в северно-восточной части Армении, в северо-западной части Азербайджана и на территории Нагорного Карабаха (рис. 3а). Большинство мест, где ранее наблюдался D. rostombekowi, хорошо поддаются предсказанию с помощью SDM (Darevsky, 1967; Uzzell, 1975; Мартиросян и др., 2003; Arakelyan et al., 2011). Фактически, модель потенциального ареала D. rostombekowi предсказала более широкий диапазон распространения за пределами уже известных подходящих мест обитания популяций Спитак, Гош, Папанино, Иджеван, Ноемберян, Тавуш, а также южной популяции на берегу оз. Севан, где уже был зарегистрирован вид. Кроме этого, модель выделяет новые небольшие потенциальные местообитания западной части оз. Севан и восточной части Нагорного-Карабаха, северной части Армении на границе с Грузией. В целом, не высокое требование вида к растительности по результатам моделирования MaxEnt, отражено в потенциальной модели распределения, включающей в себя районы, которые расположены в лесной зоне (64%), горных лугах (12%), горных степей (14%) и антропогенно-трансформированных местообитаниях 9% на высотах характерных для горных лесов (1394 ± 37 м) и горных лугов и степей (1957 ± 146 м). Северные популяции в основном расположены в лесной зоне, изолированная популяция Севан − в горных лугах и степях.
Данное исследование систематизировало знания о генетической изменчивости и распределении партеногенетического вида D. rostombekowi в нативной части ареала с использованием четырех микросателлит-содержащих локусов Du215, Du281, Du323, Du47G и 118 записей с географическими координатами присутствия вида на территории Армении, Азербайджана и Нагорного Карабаха, охватывающих большую часть его ареала. В ходе исследования мы дополняли имеющиеся литературные данные собственными полевыми наблюдениями и построили потенциальный ареал вида (ПАВ) на основе анализа климатических, топографических и ландшафтных переменных среды. Результаты генетического структуры (ГС) популяций показали, что, несмотря на наличие семи клональных линий, партеновид характеризуется одной монофилетической формой, т.е. образован в результате единичного акта гибридизации между близкими бисексуальными особями родительских видов. Анализ ПАВ и ГС указал на различие изолированной популяции юго-востока оз. Севан и северных популяций в Армении. Эти различия выражаются как в сдвиге экологических ниш этих популяций относительно друг от друга, так и в различии установленных генотипов. Прогнозируемое в настоящее время распределение D. rostombekowi с помощью ПАВ выявило новые небольшие местообитания западной части оз. Севан и восточной части Нагорного Карабаха, северной части Армении на границе с Грузией, где присутствие вида ранее не сообщалось в литературе. Результаты этого исследования и отсутствие полифилетических форм могут указывать на его регрессию. Следовательно, оценка статуса сохранения вида представляется оправданной. Результаты ценны для содействия будущим исследованиям и могут использоваться для ориентации природоохранных организаций, с целью сохранения наиболее важных местообитаний изучаемого вида.
Благодарности. Авторы благодарны международной организации ERSI за предоставление бесплатной лицензии для использования ArcGIS Desktop 10.4.1 по линии Conservation Grant (Esri Sales Order number 3128913; Esri Delivery number – 81833751, Sold-To/End-user customer number – 535452).
Финансирование. Сбор, анализ и подготовка материалов для статьи стали возможными благодаря финансовой поддержке грантов РФФИ (18-34-00361) и (17-00-00427). Молекулярно-генетические исследования выполнены при финансовой поддержке РНФ (19-14-00083).
Список литературы
Даревский И.С. Скальные ящерицы Кавказа: систематика, экология, филогения полиморфных ящериц подрода Аrсhаеolасеrtа. Л.: Наука, 1967. 214 с.
Малышева Д.Н., Даревский И.С., Токарская О.Н., Петросян В.Г., Мартиросян И.А., Рысков А.П. Изучение генетической изменчивости у однополых и двуполых видов ящериц Юго-Восточной Азии рода Leiolepis // Генетика. 2006. Т. 42. № 5. С. 581–586.
Мартиросян И.А., Рысков А.П., Петросян В.Г., Аракелян М.С., Асланян А.В., Даниелян Ф.Д., Даревский И.С., Токарская О.Н. Изменчивость мини- и микросателлитных маркеров ДНК в популяциях партеногенетической скальной ящерицы Darevskia rostombekovi // Генетика. 2002. Т. 38. № 6. С. 828–835.
Омельченко А.В., Вергун А.А., Омельченко Е.В., Лушникова А.А. Свидетельство о государственной регистрации программы для ЭВМ № 2016610517 Лабораторный конвертер № 1 – LabConverterV1, дата регистрации 13.01.16 (https://patentinform.ru/programs/reg-2016610517.html, проверено 26.08.2019).
Осипов Ф.А., Вергун А.А., Гирнык А.Е., Кутузова Н.М., Рысков А.П. Молекулярно-генетическая характеристика аллельных вариантов микросателлитных локусов Du281, Du215 и Du323 у партеногенетических ящериц Darevskia rostombekovi (сем. Lacertidae) // Мол. генетика, микробиология и вирусология. 2016. Т. 34. № 2. С. 58–61.
Agasyan A.L., Kalashyan M.Yu. (eds.). The Red Book of Animals of the Republic of Armenia. Yerevan. Zangak publ. 2010. 368 p.
Aiello-Lammens M.E., Boria R.A., Radosavljevic A., Vilela B., Anderson R.P. SpThin: an R package for spatial thinning of species occurrence records for use in ecological niche models // Ecography. 2015. № 38. P. 541–545.
Arakelyan M.S., Danielyan F.D., Corti C., Sindaco R., Leviton A.E. Herpetofauna of Armenia and Nagorno-Karabakh. SSAR Publications. 2011. Salt Lake City. 154 p.
Badaeva T.N., Malysheva D.N., Korchagin V.I., Ryskov A.P. Genetic variation and de novo mutations in the parthenogenetic Caucasian rock lizard Darevskia unisexualis // PLoS ONE. 2008. V. 3. P. 2730.https://doi.org/10.1371/journal.pone.0002730PMID: 18648496
De Pous P., Mora E., Metallinou M., Escoriza D., Comas M., Donaire D., Pleguezuelos J.M., Carranza S. Elusive but widespread? The potential distribution and genetic variation of Hyalosaurus koellikeri (Günther. 1873) in the Maghreb // Amphib-Reptil. 2011. № 32. P. 385–397.
Doronin I.V. Distribution data of rock lizards from the Darevskia (praticola) complex (Sauria: Lacertidae) // Curr. Studies Herpet. 2015. V. 15. № 1/2. P. 3–38.
Di Cola V., Broennimann O., Petitpierre B., Breiner F.T., D’Amen M., Randin C., Engler R., Pottier J., Pio D., Dubuis L., Pellissier R.G., Mateo W., Hordijk N., Salamin N., Guisan A. Ecospat: an R package to support spatial analyses and modeling of species niches and distributions // Ecography. 2017. V. 40. P. 1–14.
Edgar R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. // Nucl. Acids Res. 2004. V. 32. P. 1792–1797.https://doi.org/10.1093/nar/gkh340PMID:15034147
ESRI (Environmental Systems Research Institute) (2017): Arc GIS Desktop 10.4.1 – ESRI. Redlands. California.
Freitas S., Rocha S., Campos J., Ahmadzadeh F., Corti C., Sillero N., Ilgaz Ç., Kumlutaş Y., Arakelyan M., Harris D.J., Carretero M.A. Parthenogenesis through the Ice Ages: a biogeographic analysis of Caucasian rock lizards (genus Darevskia) // Mol. Phylogen. Evol. 2016. V. 102: 117–127.
Kaliontzopoulou A., Brito J., Carretero M.A., Larbes S., Harris D.J. Modelling the partially unknown distribution of wall lizards Podarcis in North Africa: ecological affinities. potential areas of occurrence and methodological constraint // Can. J. Zoology. 2008. V. 86. P. 992–1101.
Kamvar Z.N., Tabima J.F., GruÈnwald N.J. Poppr: an R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction // Peer. J. 2014. V. 2. P. 281. https://doi.org/10.7717/peerj.281
Korchagin V.I., Badaeva T.N., Tokarskaya O.N., Martirosyan I.A., Darevsky I.S., Ryskov A.P. Molecular characterization of allelic variants of (GATA)n microsatellite loci in parthenogenetic lizards Darevskia unisexualis (Lacertidae) // Gene. 2007. V. 292. P. 126–133.
MacCulloch R.D., Murphy R.W., Kupriyanova L.A., Darevsky I.S. The Caucasian rock lizard Lacerta rostombekovi: a monoclonal parthenogenetic vertebrate // Biochem. Syst. Ecol. 1997. V. 25. P. 33–37.
Parker E.D., Walker J.M., Paulissen M.A. Clonal diversity in Cnemidophorus: ecological and morphological consequences. In: Dawley RM, Bogart JP, editors. Evolution and Ecology of Unisexual vertebrates. Albany: New York State Museum Bulletin 466. 1989. P. 72–76.
Petitpierre B., Kueffer C., Broennimann O., Randin C., Daehler C., Guisan A. Climatic niche shifts are rare among terrestrial plant invaders // Science. 2012. V. 335. P. 1344–1348.
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Nazarenko E., Omelchenko A., Danielyan F., Arakelyan M. Analysis of geographical distribution of the parthenogenetic rock lizard Darevskia armeniaca and its parental species (D. mixta, D. valentini) based on ecological modeling // Salamandra. 2019. V. 55. P. 173–190.
Petrosyan V.G., Osipov F.A., Bobrov V.V., Dergunova N.N., Kropachev I.I., Danielyan F.D., Arakelyan M.S. New records and geographic distribution of the sympatric zones of unisexual and bisexual rock lizards of the genus Darevskia in Armenia and adjacent territories // Biodiversity Data J. 2020a. V.8. P. e56030. https://doi.org/10.3897/BDJ.8.e56030
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Omelchenko A., Varshavskiy A., Danielyan F., Arakelyan M. Species Distribution Models and Niche Partitioning among Unisexual Darevskia dahli and Its Parental Bisexual (D. portschinskii, D. mixta) Rock Lizards in the Caucasus // Mathematics. 2020b. V. 8(8). https://doi.org/10.3390/math8081329
Phillips S.J., Anderson R.P., Schapire R.E. Maximum entropy modeling of species geographic distributions // Ecol. Model. 2006. V. 190. P. 231–259.
Ryskov A.P., Osipov F.A., Omelchenko A.V., Semyenova S.K., Girnyk A.E., Korchagin V.I., Vergun A.A. Murphy R.W. The origin of multiple clones in the parthenogenetic lizard species Darevskia rostombekowi // PLoS ONE. 2017. V. 12(9). e0185161. https://doi.org/10.1371/journal.pone.0185161
Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. MEGA6: Molecular evolutionary genetics analysis version 6.0 // Mol. Biol. Evol. 2013. V. 30. P. 2725–2729.
Tarkhnishvili D., Gavashelishvili A., Avaliani A., Murtskhvaladze M., Mumladze L. Unisexual rock lizard might be out competing its bisexual progenitors in the Caucasus. // Biol. J. Linn. Soc. 2010. V. 101. P. 447–460.
Uzzell T., Darevsky I.S. Biochemical evidence for the hybrid origin of the parthenogenetic species of Lacerta saxicola complex (Sauria. Lacertidae) with a discussion of some ecological and evolutionary implications // Copeia. 1975. V. 2. P. 204–222.
Vergun A.A., Martirosyan I.A., Semyenova S.K., Omelchenko A.V., Petrosyan VG., Lazebny O.E. Clonal diversity and clone formation in the parthenogenetic Caucasian rock lizard Darevskia dahli // PLoS ONE. 2014. V. 9(3): e91674. https://doi.org/10.1371/journal.pone.0091674PMID: 24618670
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая