

Известия РАН. Серия биологическая, 2021, № 6, стр. 651-659
Экология сибирской пеночки-теньковки Phylloscopus collybita tristis Blyth на северном пределе ареала
В. Н. Рыжановский *
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: ryzhanovskiy@yandex.ru
Поступила в редакцию 22.10.2019
После доработки 19.02.2020
Принята к публикации 04.03.2020
Аннотация
Сибирская пеночка-теньковка, будучи обитателем лиственных и хвойных лесов, в Западной Сибири расширила гнездовой ареал за счет тундровой зоны. Освоению Субарктики должны способствовать особенности экологии подвида, рассмотренные в настоящей работе.
В Российской Федерации гнездятся три подвида пеночки-теньковки: восточно-европейские (Phylloscopus collybita abietinus), сибирские (Ph. c. tristis) и кавказские Ph. c. caucasicus) (Коблик и др., 2006). Восточно-европейские теньковки населяют Скандинавию и часть Восточной Европы, сибирские обитают не только в пределах всей Сибири, но проникают в Европу. Граница между восточно-европейским и сибирским подвидами проходит западнее Урала, примерно от п-ова Канин до г. Оренбург). На пространстве от этой границы до восточного склона Урала встречаются птицы с песней восточно-европейского типа, сибирского типа и гибридной песней (Коровин, 2004; Минеев, Минеев, 2012; Марова и др., 2014; Андреев, 2016).
Сибирская теньковка является выраженным лесным видом, населяющим лиственные и хвойные леса с кустарниковым и травяным ярусами (Рогачева, Сыроечковский (ред.) 2003). Относительно недавно, в середине прошлого века, считалось, что теньковка осваивает лесотундру (Птушенко, 1954; Данилов, 1966). В настоящее время северная часть ареала сибирской теньковки заходит не только в лесотундру, где пойменных лесов достаточно, но и в тундры, в частности, кустарниковые и мохово-лишайниковые (субарктические) тундры п-ова Ямал (Мониторинг .., 1997; Рябицев, Примак, 2006). Несомненно, сибирская теньковка расширяет ареал за счет Субарктики. Возможно, этому помогает текущее потепление климата, но освоение высокоширотных районов невозможно без соответствующих экологических адаптаций (Данилов, 1966).
Цель работы – описание экологии сибирской теньковки в Приобской лесотундре и на п-ове Ямал на основе собственных исследований и публикаций коллег для обсуждения адаптаций, способствующих проникновению этой пеночки в Субарктику.
МАТЕРИАЛЫ И МЕТОДЫ
Статья основана на результатах полевых наблюдений автора и коллег по Институту экологии растений и животных УрО РАН на Полярном Урале, в Нижнем Приобье и на п-ове Ямал; на материалах, полученных при обработке птиц, пойманных паутинными сетями и ловушками, на результатах экспериментальных исследований, проведенных в окрестностях г. Лабытнанги (полевые стационары Харп и Октябрьский) в летнее время и в лаборатории Экологического стационара УрО РАН (г. Лабытнанги) и Института экологии растений и животных УрО РАН (г. Екатеринбург) в зимнее время. Полевыми наблюдениями охвачена территория от долины р. Собь (66°50′ с.ш., 66°30′ с.ш.) в среднем ее течении, окрестностей г. Лабытнанги (66°35′ с.ш., 66°40′ с.ш) Приобье до широты фактории Тамбей (71°28′ с.ш.) на Северном Ямале в период с 1971 по 1990 гг. (рис. 1).
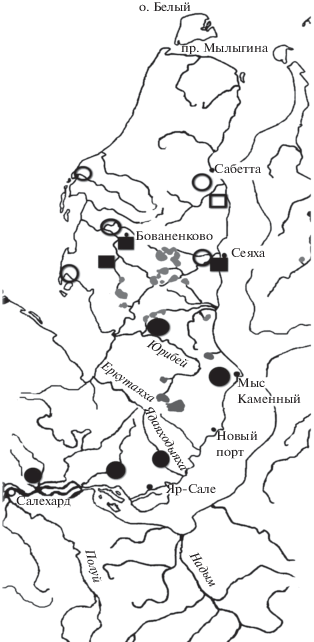
Рис. 1.
Карта встречаемости/отсутствия гнездящихся пар теньковок на п-ове Ямал в 1974–1976 гг. – кружки, в 1988–2006 гг. – квадраты. Зачернено присутствие птиц в районе наблюдений.
Учеты птиц проводились методом картирования гнездящихся пар на пробных площадках от 380 га (стационар Харп) до 22 га (стационар Октябрьский) в Нижнем Приобье, от 20 до 160 га в разных районах п-ова Ямал. В период учетов отыскивали гнезда, фиксировали сроки яйцекладки, вылупления, вылета, размер кладки, судьбу кладки и т.д.
Отловы птиц проводили в среднем течении р. Собь в 1976–1978 гг., в долине Нижней Оби (стационар Октябрьский) в 1978–1990 гг. и 2002–2015 гг. Птиц отлавливали лучками на гнездах, паутинными сетями в кустах и по берегам водоемов, в 1979–1988 гг. на берегу протока Выл-Посл стояла ловушка “рыбачинского” типа, обращенная входом на юг весной, и на север во второй половине лета. Основные годы отлова теньковок 1977–1982 гг., не регулярно в июле–августе птиц ловили в последующие годы, вплоть до 2018 г. Прижизненная обработка отловленных птиц предусматривала регистрацию веса, упитанности, пола, возраста, длины крыла. Состояние оперения описывали по известной методике (Носков, Рымкевич, 1977).
Трех слетков выкормили и содержали с 10-суточного возраста до окончания линьки при короткодневном сокращающемся фотопериоде (16С : 8Т в середине июля с последующим сокращением дня на 30 мин каждые 5 сут до 12С : 12Т в начале сентября); 4 слетка жили при естественном фотопериоде широты Полярного круга (сокращающийся от 24С : 0Т в середине июля до 16С : 8Т в начале сентября). Вместе со слетками были взяты и содержались при естественном дне до окончания линьки две самки. Два взрослых самца были пойманы в августе и жили в вольере до сентября.
Для изучения предбрачной линьки 2 взрослых самца и 2 молодые птицы были пойманы в конце июля 2011г., перевезены на Средний Урал, где жили до следующего лета при фотопериоде, сокращающемся от 17С : 8Т в начале августа до 12С : 12Т в конце сентября, при 11С : 13Т – с начала октября до конца ноября, при 14С : 10Т – с начала декабря до конца марта с последующим ростом естественной продолжительности дня за окнами помещения до 18С : 6Т в конце мая. В сентябре 2015 г. на Среднем Урале были пойманы 27 первогодков теньковки сибирского подвида, 4 из них передержаны в клетках до следующего лета. В начале октября светлую часть суток сократили до 11 ч (11С : 13Т). При таком режиме, предполагаемом фотопериоде зимовки в Северной Индии и Пакистане (Птушенко, 1954), теньковки жили до 15 февраля, затем день увеличили до 12С : 12Т, при котором птицы жили до начала естественного увеличения дня за окнами помещения в марте, после весеннего равноденствия.
При обработке материалов применяли общепринятые методы статистики программы Statistika v. 6.0 (StatSoft Ink.,1984–2003). Различия считали достоверными при Р ≤ 0.05. Среднепопуляционные даты начала, окончания и длительность линек определяли, используя формат линейного тренда программы Microsoft® Excel 2002.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Пространственное и биотопическое распространение. Распространение сибирской теньковки на Ямале (рис. 1) определяется наличием подходящих гнездовых стаций – зарослей ивняков и ерников. В районе пос. Бованенково в 1988–1990 гг. теньковки были весьма обычны (Мониторинг …, 1997), а в 2006 г. они были обычны в кустарниках окрестностей пос. Сеяха, в самом поселке (Рябицев, Примак, 2006) и в кустарниках бассейна р. Мордыяха (Слодкевич и др., 2007). Но ранее, в 1974 и 1975 гг., теньковок мы не встречали ни на восточном побережье, в устье р. Сеяха и поселке, ни в пойме междуречья Сеяха–Ясавейяха в 30 км от побережья, ни в верховьях р. Мордыяха, ни на западном побережье у мыса Моррасале. В 1974 и 1975 гг. теньковок не встречали в районе пос. Харасавэй и Тамбей, в 1988–1991 гг. эти пеночки не встречены в нижнем течении р. Венуйеуо, на границе субарктических и арктических тундр (Рябицев, 1993).
На широте пос. Мыс Каменный, в долине р. Нурмаяха и нижнем течении р. Юрибей теньковки были все годы (Данилов и др., 1984). В настоящее время южнее широты пос. Сеяха теньковки гнездятся по всей полосе тундр, от побережья Обской губы до берега Карского моря и Байдарацкой губы. Таким образом, за короткий период времени теньковки освоили северные субарктические (мохово-лишайниковые) тундры. На Южном Ямале, в Приобской лесотундре, на восточном склоне Полярного Урала теньковки встречены во всех точках, где работали мы или наши коллеги (Данилов и др., 1984; Головатин, 1999; Соколов, 2006; Головатин, Пасхальный, 2005, 2008).
В лесотундре теньковка является обитателем лесных участков. В тундровой зоне теньковки селятся в ивняковых зарослях, причем на севере субарктических тундр исключительно в поймах, а на юге тундровой зоны также и на водоразделах, по ручьям. На границе с лесотундрой они из тундр исчезают и встречаются только в пойменных лесах, на водоразделы выходят лишь отдельные пары (Головатин, Пасхальный, 2005).
Плотность гнездования. В лесотундре на широте г. Лабытнанги и на восточном склоне Полярного Урала теньковки малочисленны. В долине р. Собь в районе Красного камня плотность в 2002–2004 гг. составляла 3.0–3.2 пар/км2. На территории стационара Харп площадью 380 га в 1971–1984 гг. теньковки не гнездились, несмотря на наличие ивняков, ерников и лиственничных редколесий. В лесу склона коренного берега и в пойме стационара Октябрьский на учетной площадке 22.5 га эти пеночки гнездились в течение 5 лет из 8 с максимальной плотностью 21 пар/км2. В сезоны, когда теньковок не было на площадке, они гнездились за ее пределами. На учетных площадках в долине Хадытаяхи птицы гнездились также не регулярно. В пойменном лесу стационара Хадыта было максимально 15.4 пар/км2, на стационаре Ласточкин берег было максимально 14.3 пар/км2. Из 8 лет учетов теньковок регистрировали на последней площадке 5 лет (Рябицев, 1993).
Для кустарников поймы р. Юрибей Головатин и Пасхальный (2008) приводят следующие величины плотности: верховья 5.2 ± 1.4 пар/км2, верхнее течение 9.0 ± 0.9 пар/км2, среднее течение 3.9 ± 0.7 пар/км2, нижнее течение 1.9 ± 0.8 пар/км2. В тундрах плакора теньковки гнездились только в верховьях Юрибея 0.5 ± 0.2 пар/км2 и верхнем течении 1.2 ± 0.2 пар/км2. На западе Южного Ямала, в долине р. Еркутаяха, теньковки в 2003–2005 г. гнездились с плотностью 1–4 пар/км2 (Соколов, 2006). Учетная площадка стационара Хановэй в нижнем течении р. Нурмаяха (Данилов и др., 1984; Рябицев, 1993) была значительна по размерам (160 га), за 12 лет учетов теньковки регистрировались на ней ежегодно с плотностью 1.2–9.4 пар/км2. Таким образом, для всей подзоны кустарниковых (южных субарктических) тундр Ямала характерно гнездование теньковки с плотностью 1–10 пар/км2. В северных субарктических (мохово-лишайниковых) тундрах в среднем течении Ясавэйяхи в 1975 г. на учетной площадке 50 га в пойме и 56 га на плакоре птиц не было, как не учтены они на маршрутах в 1975 г. на мысе Моррасале (Данилов и др., 1984). В окрестности пос. Бованенково в 1988–1990 гг. теньковки гнездились в ивняках плакора с плотностью 0.8–10.0, в среднем 5.1 пар/км2 и в ивняках пойм с плотностью 5.7–13.8, в среднем 9.0 пар/км2 (Мониторинг … 1997).
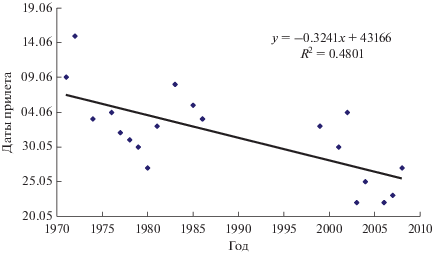
Весенняя миграция. Прилет в лесотундру начинается 22.05.2003–15.06.1972, средняя дата 31.05 (n = 20). Наблюдается достоверный тренд смещения начала прилета на более ранние даты (рис. 2) в связи с более теплыми веснами последних лет (Рыжановский, Гилев, 2020). Среднесуточная температура воздуха в день регистрации первого самца –0.6–7.8, в среднем 3.6°С (n = 19). Самки прилетали на 3–7 дней позднее первых самцов. Интенсивность пролета низкая. В долине Оби только в 1978 г. за первые две декады с начала миграции линией сетей пойманы 28 теньковок, в другие годы там же ловили не более 15 птиц. Открытых пространств в период миграции избегают. В ловушку с низкой растительностью и поляной перед входом птицы не залетали. Из 19 птиц, пойманных весной, тощие и маложирные птицы были в меньшинстве – 8 особей, упитанность остальных определена как средняя и больше средней. Летом в сети попадались птицы с небольшими запасами жира и без него.
Гнездование. Для восточно-европейского подвида весьма обычны два цикла гнездования (Лапшин, 1990). Рябицев (2014) отмечает, что даже на севере лесной зоны Сибири самка сибирского подвида может загнездиться повторно после докармливания птенцов первого выводка. В Приобской лесотундре и тундрах Ямала случаи второго гнездования не наблюдали. Гнездо представляет собой шарик из сухой травы, обычно неправильной формы с боковым входом, лежащий на земле, на кустах, на жестких стеблях прошлогодней травы, на деревьях. Лоток всегда выстлан перьями, число которых в 3 гнездах было от 119 до 125 (Данилов и др., 1984). В лесотундре из 27 найденных гнезд 5 были на вершине или сбоку кочек, 1 на пеньке, 1 на земле, 10 гнезд на елях на высоте от 1 до 11 м, 10 гнезд были на кустах или на сухих стеблях травы. В тундрах Среднего Ямала на земле, чаще сбоку кочки или на склоне оврага найдены 22 гнезда, на кустах ив найдены 43 гнезда. В заболоченном пойменном ивняке большинство найденных гнезд располагались над водой и иногда затапливались после повышения уровня воды.
Гнездо строит самка, самец поблизости активно поет. От находки гнезда на стадии каркаса до завершения перьевой выстилки проходило 2–4 сут. В эти дни наблюдается частое спаривание пары. Первое яйцо откладывалось чаще в ночь после завершения строительства, но были самки, гнезда которых пустовали 2, 3 и даже 4 дня. Период от начала прилета до начала откладывания яиц в лесотундре в три разных сезона продолжался 18, 22, 26 сут. Появление первых яиц в гнездах в Нижнем Приобье и на Южном Ямале отмечали: 20.06.1971 г., 16.06.1973 г., 1.07.1978 г. (поздневесенний год), 24.06.1979 г., 19.06.1980 г., 26.06.1981 г., в среднем 22.06. На Среднем Ямале в период между 1983–1992 гг. наиболее ранняя дата начала сезона яйцекладки – 19.06.1990 г, наиболее поздняя 29.06.1987 и 1992 гг., средняя дата 25.06. Сезон яйцекладки, как правило, короткий. На Среднем Ямале в 1987–1990 гг. в контрольных гнездах (от 4 до 8) откладывание яиц продолжалось 8–12 сут, в 1991 и 1992 гг. в связи с, предположительно, повторными кладками, сезон продолжался 20 и 21 сут (13 и 10 контрольных гнезд).
В лесотундре в 23 гнездах с полными кладками было 5–9 яиц, в среднем 5.8 ± 0.2 яиц, на Среднем Ямале в 61 кладке было 3–6 яиц, в среднем 5.4 ± ± 0.08 яиц. Сокращение величины кладки в тундровой зоне не достоверно, кладка из 9 яиц, возможно, была сдвоенной, но три гнезда в лесотундре содержали по 7 яиц.
Насиживает самка, продолжительность инкубации от первого яйца до начала вылупления составляет 16–19, в среднем 17.6 сут, от последнего яйца до вылупления 12–13, в среднем 12.5 сут (n = 8). Самки насиживали в дневные часы после откладки 3-х и более яиц, При откладывании первого–второго яйца самки в гнезде только ночуют, т.к. гнезда с начатыми кладками мы находили только в ночные часы.
Вылупление в контрольных гнездах наблюдали преимущественно во второй декаде июля, в гнездах птенцы сидели 12–14, в среднем 13 сут (n = 7). Птенцов кормили чаще всего самки, самцы с кормом встречались редко. В лесотундре вылет слетков из гнезд регистрировали, преимущественно, в третьей декаде июля – 16.7–27.7, в среднем 21.7 (n = 7). На Среднем Ямале вылет наблюдали 21.7–27.7 (n = 7), в среднем 23.7. Там же было 2 гнезда с повторными кладками, птенцы из которых вылетели 4 и 13 августа.
Эффективность размножения. В Нижнем Приобье и на Южном Ямале из 111 контрольных яиц теньковки вылупилось 75 птенцов (67.6%), из 63 птенцов, бывших под наблюдением с момента вылупления, гнезда покинули 40 слетков (63.4%). Общая успешность размножения по 23 гнездам 45.4%. На Среднем Ямале из 269 контрольных яиц, вылупилось 190 птенцов (70.6%); из этих птенцов гнезда покинули 143 слетка (75.3%). Общая успешность размножения по 49 гнездам – 53.1%. В лесотундре основная причина гибели яиц и птенцов – серая ворона, сорока, ласка и горностай, в тундровой зоне – песец и горностай. В поймах в дожди гнезда размокали или тонули в поднявшейся воде.
Территориальный консерватизм и филопатрия. На территории стационара Октябрьский в весенне-летне-осенний период окольцовано 48 взрослых теньковок. На следующий год сетями поймано 3 самца (6.25%). Из 90 молодых теньковок, пойманных и окольцованных на участке в послегнездовое время, на следующий год вернулись 2 птицы (2.22%). На стационар Ласточкин берег вернулся один самец из 5 (20.0%), на стационар Хановэй вернулись 3 самца из 8 (37.5%) и 2 самки из 27 (7.4%) (Рябицев, 1993). Из окольцованы в гнездах 45 слетков, на следующий год в районе рождения никто не пойман.
Послегнездовые кочевки. На стационаре Октябрьский пойманы три окольцованные в гнезде теньковки через 21, 25 и 29 сут после вылупления. В послегнездовой период теньковки в пойме Оби были малочисленны, только в 1980 г. поймано 49 птиц, в другие годы – от 2 до 13. Интенсивность отловов возрастала в последней декаде августа в связи с началом миграции. Из 63 окольцованных в августе птиц повторно, через день, поймана одна особь (1.6%). Среди 36 птиц, помеченных в сентябре, повторно не поймано ни одной. Участок отлова нельзя считать непригодным для вида, ежегодно здесь гнездились 1–3 пары птиц. Видимо, низкий процент повторных отловов отражает особенности послегнездового периода этого вида.
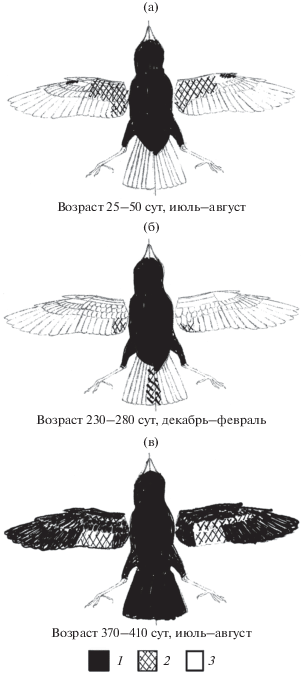
Постювенальная линька. В природе теньковки заменяют кроющие туловища в обычном для воробьиных птиц объеме, т.е. перья, выросшие в гнезде. На крыле у некоторых птиц заменялись большие верхние кроющие второстепенных маховых, средние верхние кроющие второстепенных маховых, чаще внутренние, заменялись у большинства птиц, малые верхние кроющие второстепенных маховых – также у некоторых птиц. У большинства птиц заменялись верхние и нижние кроющие пропатагиальной складки, верхние и нижние кроющие кисти, средние нижние кроющие второстепенных маховых и нижние кроющие третьестепенных маховых (рис. 3).
Рис. 3.
Полнота, возраст и сроки постювенальной (а), предбрачной (б) и послебрачной (в) линек в течение первого года жизни: 1 – линяет у всех осмотренных птиц, 2 – линяет у части или некоторых осмотренных птиц, 3 – не линяет; слева – верх тела, справа – низ тела.
Линька начинается рано, до полного отрастания рулевых и маховых перьев. Предшествует линьке дорастание юношеских перьев по периферии крупных птерилий. Начинается дорастание в возрасте 15–17 сут, продолжается до 30–35-суточного возраста. Три теньковки из не распавшегося выводка, пойманные повторно в возрасте 23 сут, имели дорастающие перья по всему телу, но линька не начиналась. В неволе при короткодневном фотопериоде (16С : 8Т) выкормлены 3 теньковки, при естественном дне – 4. Птицы первой группы начали линьку в 24–29, в среднем в 25.7 сут, птицы второй группы – в 24–25, в среднем в 24.2 сут. При коротком дне линька пеночек длилась 23–35, в среднем 27 сут, при естественном дне 36–40, в среднем 38 ± 0.9 сут. Несомненно, возраст начала линьки у этого вида контролируется эндогенно, но темпы ее зависят от внешних фотопериодических условий (Рыжановский, 2017).
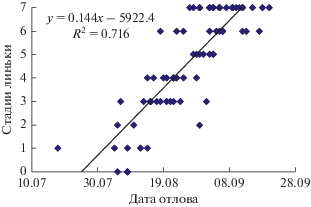
Начинающие линьку теньковки отлавливались в первой декаде августа, с 2.08 по 12.08, но в раннем 1977 г. одна птица поймана 19 июля. Три не начавшие линьку слетка пойманы 8 августа (рис. 4). При вылуплении птенцов в первой декаде июля, расчетный возраст начала линьки особи в природе был меньше 30 сут. Основной период линьки молодых северных теньковок – август. После 27.08 мы ловили птиц, закончивших линьку. По регрессии средняя многолетняя дата начала линьки – 31 июля, окончания – 11 сентября, средняя длительность – 42 сут. Самая поздняя дата отлова на стационаре Октябрьский заканчивающей линьку птицы – 18.09.1978 г. (фенологически поздний год), но основная масса птиц, заканчивающих линьку и в новом оперении, отлавливалась в первой половине сентября. Доля таких птиц среди теньковок, пойманных за вторую половину лета, значительна – 42.6%. Это дает основание считать, что молодые теньковки включаются в миграцию на последней стадии линьки или после ее окончания. Сезон линьки в 1977 г. длился 45–50 сут, в 1980 г – 35–40 сут.
Рис. 4.
Сроки отлова теньковок на разных этапах постювенальной линьки. Обозначения: ось ординат 0 – линька не началась, 7 – линька закончилась. Линия тренда соединяет средние даты начала и окончания линьки.
Послебрачная линька. Для части теньковок Нижнего Приобья характерна неполная линька маховых (рис. 3). Из пары, взятой с выводком, самка не сменила второстепенные маховые, малые верхние кроющие второстепенных маховых и часть нижних кроющих крыла, самец сменил все перья. Но 6 теньковок, пойманные на завершающих этапах линьки, заменяли все маховые перья. С гнездованием линька теньковок если и совмещается, то незначительно. Все 4 взрослые птицы, пойманные у гнезда с 10–12-суточными птенцами, были в старом наряде. В дальнейшем один самец был случайно выпущен, линька второго началась по достижении птенцами 19-суточного возраста, линька самок началась через 27 и 29 сут после вылупления птенцов.
Начинающих линьку взрослых пеночек отлавливали в период с 20.7 по 7.08, в старом наряде птицы попадали в сети до 31 июля (рис. 5), средняя дата начала линьки – 27.07. Среди птиц, первыми вступившими в линьку, преобладали самцы, среди поздно ее начавших – самки. Период вступления в линьку продолжается 25–30 сут. Две теньковки, заканчивающие смену наряда (на 11-й стадии), пойманы 19.08 и 20.08. Средняя дата окончания линьки – 27.08, средняя длительность линьки 32 сут, длительность сезона линьки – 50–60 сут. В вольере при естественном дне вместе с выводком содержали самца. Линьку он начал 4 августа и закончил через 39 сут, 11 сентября.
Рис. 5.
Сроки отлова теньковок на разных этапах послебрачной линьки. Обозначения: ось ординат 0 – линька не началась, 12 – линька закончилась. Линия тренда соединяет средние даты начала и окончания линьки.
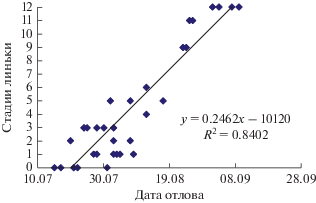
Предбрачная линька у теньковок из Нижнего Приобья в клетках началась в последней пятидневке декабря, через 3 недели после перевода с фотопериода 11С:13Т на 14С:10Т и закончилась в первых числах февраля. Птицы, пойманные на Среднем Урале, начали линьку в первых числах марта, через 2 недели после увеличения светлой фазы с 11С до 12С. Полнота линьки теньковок из Приобья и пойманных на Урале существенно не отличалась. У всех передержанных птиц заменились контурное оперение головы и туловища. У двух северных первогодков заменились 17, 18, 19-е маховые и центральные рулевые, у 2 птиц, пойманных на Среднем Урале, заменились 18-е маховые и центральные рулевые. У остальных птиц перелиняли только голова и туловище (рис. 3). Продолжалась линька 30–35 сут, до конца марта.
Осенняя миграция. Теньковки мигрируют ночью (Большаков, 1975). Наблюдения за клеточными птицами показали, что с начала сентября, находясь в возрасте старше 50 сут две пеночки ночью периодически, сидя на жердочке, трясли крыльями, 7 сентября одна летала по большой вольере. Миграционное ожирение началось через 1–3 сут после окончания линьки, в возрасте 55–58 сут. В Нижнем Приобье последняя молодая птица была поймана 11.09.1975 г., 6.09.1977 г., 21.9.1978 г., 9.09.1980 г.; взрослая – 10.09.1979 г. Общая продолжительность весенне-летне-осеннего сезона теньковок в лесотундре 112–118 сут, в северных субарктических тундрах менее 100 сут.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Данилов (1966) выделял следующие направления адаптаций птиц к условиям Субарктики и Арктики: внутривидовую дифференциацию с формированием субарктических подвидов и популяций; способность к отложению жира в немиграционный период; повышение пороговых значений чувствительности к освещенности; повышение резистентности к низким температурам среды, сокращение длительности основных сезонных явлений: прилета, гнездования, линьки, отлета.
Северных подвидов теньковки нет. Ареал сибирской теньковки имеет большие, по сравнению с другими подвидами размеры. Теньковки номинативного подвида (Ph. c. collybita) населяют юго-запад Европы и в Субарктику не проникают, восточно-европейские теньковки (Ph. c. abietinus) населяют Восточную Европу и север Скандинавии с ее достаточно мягким климатом. Сибирские теньковки освоили не только лесотундру Сибири и Восточной Европы, но на Ямале и Гыданском полуострове глубоко проникают в тундровую зону (Рябицев, 2014). На пространстве Европейской части России восточно-европейские и сибирские теньковки обитают совместно, образуют пары. Территория, где регистрируют сибирских теньковок и потомство смешанных пар, самцов с гибридной песней, растет к западу. Мальчевский и Пукинский (1983) не приводят случаев встреч сибирской теньковки или регистрации гибридных песен для Ленинградской области и прилегающей территории. Но в настоящее время район регистрации сибирского подвида и гибридов включает Новгородскую область (Архипов, 2018) и Карелию (Хохлова, 2017). Возможно, одновременно восточно-европейская теньковка продвигается на восток, т.к. гибридные песни слышали на Среднем и Южном Урале при доминировании сибирской теньковки (Коровин, 2004; Торгашов, 2004; Марова и др., 2014).
Расширение ареала сибирской теньковки в Западной Сибири за счет тундр Ямала и Гыданского полуострова может свидетельствовать о формировании тундровой популяции сибирских теньковок, не требующих для гнездования биотопов с лесной растительностью. В северных субарктических тундрах кустарниковый ярус имеет высоту 0.8–1 м, на границе с арктическими тундрами 0.6–0.8 м (Мониторинг …, 1997) при плотности пар, не меньшей чем в лесотундре и северной тайге. В умеренных широтах существуют значительные биотопические отличия между весничкой, также проникающей в северные субарктические тундры и теньковкой (Батова, Бурский, 2008), в тундрах отличий нет. Здесь птицы занимают общий биотоп – ивняки травяно-моховые пойм и водоразделов. Но если обитатель кустарников и куртинного подроста умеренных широт весничка незначительно изменила местообитания в тундрах, то теньковка почти радикально.
Небольшие запасы жира в гнездовое время, найденные у теньковок лесотундры, как у весничек и таловок (Рыжановский, 2004), можно рассматривать в качестве адаптации к нерегулярным летним возвратам холодов. Но полярный день, при необходимости, позволяет тундровым птицам кормиться круглосуточно, что они иногда делают. Возможно, летние запасы жира являются побочным следствием изобилия беспозвоночных в тундрах в середине лета (Ольшванг, 1992).
В лесотундре полярный день длится месяц, от занятия парами территорий до вылупления птенцов, в северных субарктических тундрах – три месяца. Почти весь период пребывания северных сибирских теньковок в тундрах не должен контролироваться фотопериодической реакцией, в отличие от теньковок умеренных широт, где наличествуют темная и светлая фазы суток. Минимальный возраст начала линьки у обоих европейских подвидов одинаковый: 32–35 сут, но максимальный возраст разный: 39–40 сут в Приладожье, 58–76 сут – в Германии, несмотря на более долгий день естественного фотопериода Приладожья, т.е. европейские подвиды отличаются реакцией на длину дня (Gwinner, 1969; Лапшин, 1990). Восточно-европейские теньковки при фотопериоде 70-й параллели (24С:0Т до конца июля) начнут линьку в первой половине августа, в возрасте не старше 40 сут. Западно-европейские теньковки Ph. c. collybita при фотопериоде 70-й параллели должны начать линьку в возрасте 60–70 сут, в конце августа, что слишком поздно, поэтому в Скандинавии гнездятся Ph. c. abietinus, более адаптированные к световому режиму приполярных широт.
Сведений об особенностях фотопериодического контроля годового цикла таежных сибирских теньковок у нас нет, адаптация лесотундровых и тундровых теньковок к световому режиму Субарктики, возможно, происходит в настоящее время. Весной прилетающие в лесотундру теньковки нуждаются в фотостимуляции гонад полярным днем, т.к. период от прилета до яйцекладки достаточно длинный (18–28, в среднем 22.0 сут; n = 3), однако не достоверно длиннее, чем у весничек (12–26, в среднем 20.9 ± 1.3; n = 11 сут), необходимость фотостимуляции которых выявлена в эксперименте (Рыжановский, 2001). Сроки начала постювенальной линьки теньковок определяются эндогенно, как продолжение роста-развития, что характерно для ряда северных видов и популяций и адаптивно в летних условиях Субарктики (Рыжановский, 2007). Послебрачная линька теньковок начинается поздно, после распадения выводков (конец июля – начало августа), в отличие от настоящих субарктов (Рыжановский, 1997), совмещающих линьку с кормлением птенцов (иногда с насиживанием)и тем самым сдвигающих ее на более ранние даты, на середину июля. Темпы линьки северных теньковок высокие. Постювенальная и послебрачная линьки птиц из Нижнего Приобья округленно длилась при естественном дне 30–40 сут, в Приладожье 45–60 сут, в Западной Европе – 60–70 сут (Рыжановский, 2017). Но даже при высоких темпах линек северные теньковки отлетают из тундр и лесотундры последними из славковых, так как миграционное состояние формируется по окончанию линьки. На Среднем Ямале в районе пос. Бованенково в 1989 г. теньковки встречались на маршрутах до 6.09. (Шутов, личное сообщение), в лесотундре отлет заканчивается еще позднее.
Несомненно, теньковки холодоустойчивы. Даже восточно-европейские теньковки на Куршскую косу прилетают раньше других пеночек, улетают позднее, находясь в наиболее жестких, для пеночек, ранневесенних и позднеосенних погодных условиях (Паевский, 2018). В Приобскую лесотундру и тундровую зону сибирские теньковки прилетают и отлетают при температурах, близких 0°С. Безморозный период на Среднем Ямале длится 65–70 сут (Шиятов, Мазепа, 1995) при длительности весенне-летне-осеннего сезона теньковок 100–110 сут, т.е. часть годового цикла тундровых птиц протекает при низких положительных температурах. Даже через Средний Урал теньковки летят во второй половине сентября – начале октября, при температуре воздуха годы несколько выше 0°С, также последними из славковых.
* * *
На пространстве Западной Сибири сибирская теньковка за последние десятилетия расширила гнездовой ареал за счет северных субарктических тундр. При этом изменились требования к биотопу, вместо многоярусной растительности птицам достаточно низких разреженных ивняков. У этих пеночек нет второго цикла размножения, редки повторные кладки, весенне-лнтне-осенний сезон растянут в связи с особенностями фотопериодической реакции и холодостойкостью.
Работа выполнена при финансовой поддержке Президиума УрО РАН (проект 18-9-4-22).
Список литературы
Андреев А.А. Пеночки Phylloscopus в Архангельске и его пригородной зоне // Рус. орнитол. журн. 2016. Т. 25. № 1262. С. 939–947.
Архипов В.Ю. Первая регистрация сибирской теньковки Phylloscopus collybita tristis в Новгородской области // Рус. орнитол. журн., 2018. Т. 27. № 1702. С. 5869–5872.
Батова О.И., Бурский О.В. Гнездовые местообитания пеночек (Phylloscopus) в центральной Сибири // Бюл. Моск. о-ва испытателей природы. Отд. биологии. 2008. Т. 113. №. 3. С. 18–28.
Большаков К.В. Некоторые особенности звуковой сигнализации мигрирующих ночью птиц // Сообщения Прибалт. комиссии по изучению миграций птиц. Тарту. 1975. № 9. С. 137–149.
Головатин М.Г. Птицы бассейна реки Войкар // Мат-лы к распространению птиц на Урале, в Приуралье, и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 1999. С. 75–81.
Головатин М.Г., Пасхальный С.П. Птицы Полярного Урала. Екатеринбург: Изд-во Уральского ун-та, 2005. 564 с.
Головатин. М.Г., Пасхальный С.П. Современное состояние орнитофауны долины р. Юрибей (Южный Ямал) и перспективы создания в бассейне реки природного парка. Научный вестник ЯНАО. Региональные аспекты исследований. Вып. 8(60) Салехард. 2008. C. 81–102.
Данилов Н.Н. Пути приспособлений наземных позвоночных животных к условиям существования в Субарктике. Т. 2. Птицы. Труды ин-та биологии УФАН СССР. Свердловск, 1966. 140 с.
Данилов Н.Н., Рыжановский В.Н., Рябицев В.К. Птицы Ямала. М.: Наука. 1984. 334 с.
Коблик Е.А., Редькин Я.А., Архипов В.А. Список птиц Российской Федерации. М.: КМК, 2006. 281 с.
Коровин В.А. О встречах восточно-европейской теньковки на Среднем Урале // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 2004. С. 105–106.
Лапшин Н.В. Пеночка-теньковка – Phylloscopus collybita (Vieill.) // Линька воробьиных птиц Северо-Запада СССР. Л.: Изд-во Ленинградского ун-та, 1990. С. 28–33.
Мальчевский А.С., Пукинский Ю.Б. Птицы Ленинградской области и сопредельных территорий. Л.: Изд-во Ленинградского ун-та, 1983. Т. 2. 504 с.
Марова И.М., Шипилина Д.А., Алексеев В.Н., Федоров В.В., Иваницкий В.В. Зона вторичного контакта восточно-европейской и сибирской теньковок (Phylloscopus collybita abietinus – Ph. c. tristis) на Южном Урале: свидетельство гибридизации // Труды Южно-Уральского гос. природ. заповедника. Уфа. 2014. С. 95–117.
Минеев Ю.Н., Минеев О.Ю. Птицы Большеземельской тундры и Югорского полуострова. СПб.: Наука, 2012. 383 с.
Мониторинг биоты полуострова Ямал в связи с развитием объектов добычи и транспорта газа / Под ред. Добринского Л.Н. Екатеринбург: Аэрокосмоэкология, 1997. 191 с.
Носков Г.А., Рымкевич Т.А. Методика изучения внутривидовой изменчивости линьки у птиц // Методика исследования продуктивности и структуры видов в пределах их ареалов. Вильнюс: Изд-во Москлас, 1977. Ч. 1. С. 37–48.
Ольшванг В.Н. Структура и динамика населения насекомых Южного Ямала. Екатеринбург: Наука, 1992. 103 с.
Паевский В.А. Сравнительно-экологический анализ трех симпатричных видов пеночек – веснички Phylloscopus trochilus, теньковки Ph collybita и трещотки Ph. sibilatrix // Рус. орнитол. журн., 2018. Т. 27. № 1608. С. 2244–2253.
Птушенко Е.С. Род пеночка // Птицы Советского Союза. М.: Сов. Наука, 1954. Т. 6. С. 153–161. С. 2869–2872.
Рогачева Э.В., Сыроечковский Е.Е. (ред.). Атлас гнездящихся птиц Европы европейского совета по учетам птиц / Под ред. Рогачевой Э.В., Сыроечковского Е.Е. М.: ИПЭЭ РАН, 2003. 338 с.
Рыжановский В.Н. Экология послегнездового периода жизни воробьиных птиц Субарктики. Екатеринбург: Изд-во Уральского ун-та, 1997. 283 с.
Рыжановский В.Н Гнездовой сезон как часть годового цикла воробьиных Субарктики // Гнездовая жизнь птиц. Пермь, 2001. С. 3–22.
Рыжановский В.Н. Масса тела и жировые резервы воробьиных птиц Нижнего Приобья // Рус. орнитол. журн., 2004. Т. 13. № 271. С. 799–812.
Рыжановский В.Н. Роль фотопериодических условий высоких широт в дифференциации популяций воробьиных птиц // Рус. орнитол. журн., 2007. Т. 16. № 380. С. 307–314.
Рыжановский В.Н. Подвидовые особенности линьки пеночки-теньковки (Phylloscopus collibita L.) из Европы и Западной Сибири // Экология, 2017. № 3 С. 216–222.
Рыжановский В.Н., Гилев А.В. Об иерархии факторов, определяющих сроки начала прилета воробьеобразных птиц (Passeriformes) в Приобскую лесотундру // Зоологический журнал 2020. № 2. С. 1–16.
Рябицев В.К. Территориальные отношения и динамика сообществ птиц в Субарктике. Екатеринбург: Наука, 1993. 296 с.
Рябицев В.К. Птицы Урала, Приуралья и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 2001. 605 с.
Рябицев В.К. Птицы Сибири. Москва; Екатеринбург: Кабинетный ученый. 2014. Т. 1. 438 с.
Рябицев В.К., Примак И.В. К фауне птиц Среднего Ямала // Материалы к распределению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 2006. С. 184–190.
Слодкевич В.Я., Пилипенко Д.В., Яковлев А.А. Материалы по орнитофауне долины реки Мордыяха // Мат-лы к распространению птиц на Урале, в Приуралье, и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 2007. С. 221–233.
Соколов В.А. Население птиц на юго-западном Ямале и его динамика. Дис. канд. биол. наук. Екатеринбург: Ин-т экологии растений и животных УрО РАН. 2006. 170 с.
Торгашов О.А. К распространению восточно-европейского и сибирского подвидов теньковки в горно-лесной зоне Южного Урала // Мат-лы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та, 2004. С. 182–183.
Хохлова, Т.Ю. Встреча гибридной теньковки Phylloscopus collybita abietinus х Ph. c. tristis в Себежском районе Карелии // Русский орнитол. журн., 2017.Т. 26. № 1519. С. 4552–4553.
Шиятов С.Г., Мазепа В.С. Климат / Природа Ямала. Екатеринбург: УИФ Наука. 1995. С. 37–51.
Gwinner E. Untersuchung zur Jahresperiodik von Laubsangern. Entwicklung der Gefieders, der Gewichter und der Zugunruhe bei Jungvogeln der Arten Phylloscopus: Ph. trochilus, Ph. sibilatrix und Ph. collybita // J. Ornithol. 1969. Bd. 110. № 1. S. 1–21.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая