

Известия РАН. Серия физическая, 2021, T. 85, № 3, стр. 353-360
Диатомовый сдвиг в озерах Арктики: реакция на глобальное потепление или изменение спектральных характеристик излучения Солнца?
Е. А. Касаткина 1, 2, О. И. Шумилов 1, 2, *, Д. В. Макаров 1, Д. Б. Денисов 1
1 Федеральное государственное бюджетное учреждение науки
Институт проблем промышленной экологии Севера Кольского научного центра Российской академии наук
Апатиты, Россия
2 Федеральное государственное бюджетное научное учреждение
Полярный геофизический институт
Апатиты, Россия
* E-mail: oleg@aprec.ru
Поступила в редакцию 25.09.2020
После доработки 20.10.2020
Принята к публикации 27.11.2020
Аннотация
За последние десятилетия произошло значительное увеличение биомассы фитопланктона во многих озерах мира. В качестве наиболее вероятной причины обнаруженного эффекта в некоторых работах было предложено глобальное потепление. Нами проанализирован недавний рост общей численности диатомовых водорослей в озере Раббватнет (Северная Норвегия). Для анализа использованы палеолимнологические, дендрохронологические и метеорологические наборы региональных данных, а также информация о локальном загрязнении. Обсуждаются возможные причины наблюдаемого роста, включая глобальное потепление климата и вариации спектральных характеристик солнечной активности.
ВВЕДЕНИЕ
Диатомовые водоросли (диатомеи) являются важнейшими и наиболее распространенными представителями фитопланктонного сообщества и обитают практически повсеместно в морских и пресноводных экосистемах (рис. 1а). Эти одноклеточные организмы, размером от 2 до 500 мкм, поглощают углекислый газ и вырабатывают почти 25% кислорода нашей планеты посредством фотосинтеза, способствуя углеродному циклу Земли [1–3]. Препятствуя возрастанию атмосферного СО2, диатомеи участвуют в фотосинтетическом связывании углерода и переносе его на дно водоемов, чем обеспечивают поступление большого количества (до 40%) органического углерода и составляют, таким образом, основу пищевой цепочки водных экосистем [1, 2, 4].
Рис. 1.
Композитная микрофотография различных по размеру и форме представителей диатомовых водорослей: быстро тонущие рода Aulacoseira (цилиндрические) и термофильные, более плавучие рода Cyclotella (дискообразные) [2] – (а); модельное изображение наноструктуры диатомовой оболочки (фрустулы) [5] – (б).
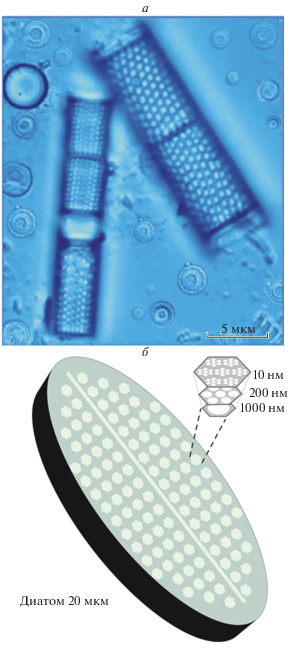
Диатомеи отличаются от других групп водорослей строением своей оболочки (фрустулы), состоящей из кремнезема (оксида кремния SiO2). Диатомовые фрустулы благодаря своей пористой иерархической наноструктуре, приспособленной для улавливания света (рис. 1б), находят все более широкое применение в светозахватывающих нанотехнологиях, например, в производстве элементов для солнечных батарей [5, 6]. Специфические свойства диатомовых водорослей, а именно их способность к биоминерализации (процессу синтеза неорганических твердых веществ в живых организмах) позволяют использовать их при создании полупроводников и сверхпрочных биокомпозитных материалов [7–9]. Интересно, что кроме создания свойственных собственно диатомовым комплексам соединений типа SiO2, эти микроводоросли могут использоваться для синтеза других важных химических соединений, таких как кристаллический карбонат кальция (CaCO3) [8] и металлы (платина (Pt) и кадмий (Сd)) [7]. Использование процесса биоминерализации диатомовых комплексов находит все более широкое применение в бионанотехнологиях по сравнению с обычными методами синтеза аналогичных материалов, которые требуют создания специальных режимов высоких температур и являются очень энергозатратными [7, 8]. Диатомовые водоросли являются также перспективным источником биологически активных веществ, полезных для человека (полиненасыщенные жирные кислоты, полисахариды, витамины и др.) [10]. Сверхчувствительность диатомовых водорослей к изменениям внешней среды позволяет использовать их для мониторинга качества воды в пресноводных водоемах с повышенной антропогенной нагрузкой [11], а также для очистки воды и утилизации отходов [7].
Диатомеи являются эффективным показателем изменчивости климата из-за высокой чувствительности к любым изменениям естественных факторов окружающей среды, таким как солнечная радиация, величина ледяного покрова, метеопараметры (температура воздуха, ветер, осадки) [2, 4, 12]. Поэтому изучение диатомовых комплексов в донных отложениях различных водоемов важно для палеонтологических, палеоклиматических и палеогеографических реконструкций на различных временных масштабах, начиная от голоцена (~10 тыс. лет) [13] и заканчивая эпохой эоцена (~50 млн лет) [14].
В то же время, взаимодействие между изменениями климата и пресноводными диатомовыми сообществами имеет сложную структуру, поскольку другие факторы, такие как морфометрические характеристики озера (географическое положение, глубина, площадь), а также плотность воды, наличие питательных ресурсов и степень стратификации, могут влиять на численность и динамику состава диатомовых водорослей [2, 4]. Кроме того, климат также может оказывать непрямое воздействие на диатомеи, изменяя сами факторы окружающей среды [4].
В последние десятилетия во многих озерах было зафиксировано увеличение биомассы фитопланктона [15–18]. В некоторых исследованиях в качестве вероятной причины наблюдаемого сдвига рассматривалось недавнее глобальное потепление [15, 17, 19]. Действительно, по данным измерений температура поверхности многих озер в последние десятилетия увеличивалась, как полагают, вследствие повышения температуры воздуха [20‒25]. Тем не менее, другие факторы и механизмы, в том числе не связанные с температурой, включая солнечную радиацию, также могут рассматриваться в качестве возможных причин наблюдаемого увеличения биомассы диатомовых водорослей [2, 4, 16, 17]. Кроме того, реакция диатомовых водорослей на изменение климата может значительно различаться в зависимости от географического положения и морфометрических характеристик озера с увеличением степени воздействия в высоких широтах [2, 12].
В данной работе представлены результаты диатомового анализа колонки донных отложений из арктического озера Раббватнет в северной Норвегии за период 1895–2012 гг. Для изучения возможных причин “диатомового сдвига” были использованы региональные палеоклиматические (дендрохронологические) и метеорологические данные, а также параметры загрязнения окружающей среды.
РАЙОН ИССЛЕДОВАНИЯ
Озеро Раббватнет (69.7° с.ш., 30.5° в.д.) – небольшое субарктическое озеро олиготрофного типа, расположенное на высоте 83 м на побережье Баренцева моря. Площадь озера составляет 0.4 км2, а максимальная глубина 10 м. Озеро находится на территории Норвегии, в приграничной зоне между Россией и Норвегией. Начиная с 1940-х годов озерная экосистема подвержена влиянию атмосферных выбросов диоксида серы (SO2) и тяжелых металлов (Ni, Cu, Co) с заводов предприятия “Печенганикель” (российское предприятие “Норильский Никель”), находящихся в городах Никеле (69.4° с.ш., 30.3° в.д.) и Заполярном (69.4° с.ш., 30.8° в.д.), приблизительно в 30 км от озера Раббватнет. Наиболее значительный рост содержания тяжелых металлов в озерных отложениях начиная с 1970–1980-х годов связан с началом переработки норильской руды с высоким (до 30%) содержанием серы [26]. Общий объем выбросов SO2 в течение этого периода составлял более 400 тыс. т. в год, в настоящее время, с началом переработки местной руды, выбросы сократились примерно до 100 тыс. т. [27].
В г. Никель средняя годовая температура воздуха составляет + 0.2°C. Январь является самым холодным, а июль – самым теплым месяцем со средними значениями температуры воздуха –10.7 и + 13.1°C, соответственно [27]. Зимой преобладающими направлениями ветра являются южное и юго-западное, а летом преобладают северные и северо-восточные ветры. Среднегодовая скорость ветра в г. Никель составляет 3.8 м ∙ с–1 [27]. Зимой территория находится в условиях полярной ночи с полным отсутствием солнечного света.
Для более детального изучения климата в регионе были использованы метеоданные норвежской станции Варде (70.4° с.ш., 31.1° в.д.) и собственная древесно-кольцевая хронология Кольского полуострова (Кузрека; 66.6° с.ш., 34.8° в.д.). Данная хронология является ближайшей к оз. Раббватнет древесно-кольцевой хронологией, охватывающей исследуемый период, включая последнее десятилетие.
МАТЕРИАЛЫ И МЕТОДЫ
Отбор проб донных отложений состоялся в 2013 г. с помощью пробоотборника открытого гравитационного типа с автоматически закрывающейся диафрагмой в самой глубокой (~10 м) части оз. Раббватнет. Колонка мощностью 44 см была разделена на слои по 1 см. Диатомовый анализ донных отложений проводился в ИППЭС КНЦ РАН по стандартной общепринятой методике [11]. Определение таксономической идентификации и подсчет количества диатомей проводились с помощью светового микроскопа “Motic BA 300" при увеличениях в 400–1000 раз. Для каждого слоя донных отложений определялись следующие показатели: общая численность диатомей N (млн экз./г), относительная численность каждого таксона (%), а также видовое разнообразие по индексу Шеннона–Уивера I (бит/экз.):
pi – удельная численность i-го таксона, pi = = ${{{{N}_{i}}} \mathord{\left/ {\vphantom {{{{N}_{i}}} {\sum {{{N}_{i}}} }}} \right. \kern-0em} {\sum {{{N}_{i}}} }}$, где Ni – численность i-го таксона. Номенклатура приведена в соответствии с международной альгологической базой данных [28].
Датировка колонки донных отложений и расчет средней скорости осадконакопления были выполнены в Институте геохимии и аналитической химии им. В.И. Вернадского РАН. Определение радиоактивности изотопов 137Cs и 210Pb в пробах донных отложений проводили с использованием методов гамма-спектрометрии. Расчетное значение абсолютной скорости седиментации для озера Раббватнет составило 0.65 мм/год, а приблизительный возраст колонки – более 680 лет.
Реконструкция величины водородного показателя pH проводилась в соответствии с методами, изложенными в [29] по следующей формуле:
где phi – индивидуальное значение для каждого таксона-индикатора, ki – величина численности индикаторного таксона.В работе использована древесно-кольцевая хронология (1592–2014 гг.), полученная по образцам сосны Pinus sylvestris L. в районе д. Кузрека (66.6° с.ш., 34.8° в.д.). Ширина колец измерялась с точностью до 0.01 мм с использованием сканера Epson Perfection V850 и соответствующего программного обеспечения TREMET [30]. При построении хронологии применялись современные методики дендроанализа (перекрестное датирование, стандартизация) с использованием программ COFECHA и ARSTAN [31, 32].
РЕЗУЛЬТАТЫ
При анализе диатомовых комплексов в колонке донных отложений оз. Раббватнет было выявлено 255 таксонов. Типичными представителями диатомовых являются Aulacoseira alpigena, Aulacoseira distans, Fragilariales и представители вида Cyclotella. Наиболее массовые виды родов Aulacoseira и Cyclotella составляют основу численности диатомовых в озере Раббватнет (рис. 1а). Доминирующими видами на всем временном интервале (~680 лет) были Aulacoseira alpigena некоторые представители рода Cyclotella, но общая численность N оставалась относительно низкой до ~1940 г. (рис. 2д). На рис. 2 представлены вариации N (рис. 2д) и относительной численности доминирующих таксонов (Aulacoseira alpigena и Cyclotella schumanni) (рис. 2г) оз. Раббватнет за период 1895–2012 гг. В верхних слоях, начиная с ~1940 г. доминировал таксон Aulacoseira alpigena (>50%) (рис. 2г). В этот период в диатомовом сообществе появляются некоторые новые виды. Основные виды рода Cyclotella были доминантными (>40%) до ~1950 г., но с тех пор их численность стала уменьшаться (рис. 2г). Общая численность диатомовых водорослей N характеризовалась постепенным ростом с 1920 г. (рис. 2д). Как видно из рис. 2д, помимо двух максимумов в 1965 и 1980 гг., наиболее значительное увеличение величины N наблюдалось с ~2002 г. до настоящего времени. Изменения в видовом разнообразии диатомового сообщества оз. Раббватнет происходили на протяжении всего исследуемого периода (рис. 2в).
Рис. 2.
Вариации по данным измерений донных отложений в оз. Раббватнет концентрации фосфора (мг ∙ г–1) – (а); водородного показателя pH – (б); индекса видового разнообразия (бит экз.–1) – (в); относительной численности представителей различных таксонов диатомовых (%): Aulacoseira alpigena (светло-серый) и Cyclotella schumanni (черный) – (г); общей численности диатомовых (млн экз./г).
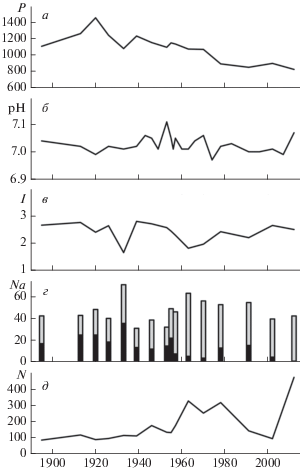
В первой половине ХХ в. наблюдается снижение видового разнообразия до ~1930 г., когда величина I достигает своего минимального значения (1.66 бит/экз.) (рис. 2в). Причем это снижение происходит одновременно с ростом относительной численности диатомовых комплексов Aulacoseira alpigena и Cyclotella schumanni (рис. 2в). Следующий минимум величины I зафиксирован в ~1960 г. на фоне роста N и относительной численности Aulacoseira alpigena, после чего начинается рост индекса Шеннона–Уивера (рис. 2в).
Азот и фосфор являются основными элементами минерального питания, которые могут существенным образом ограничить рост диатомовых водорослей. Видно, что с 1920 г. до настоящего времени наблюдалось постепенное снижение концентрации фосфора Р (рис. 2а), хотя в целом концентрации этих питательных веществ в оз. Раббватнет являются типичными для пресноводных арктических водоемов [27].
Значения pH, реконструированные по диатомовым водорослям, на протяжении сего временного интервала изменялись от 7 до 7.05, что, в целом, свидетельствует о нейтральном статусе воды в оз. Раббватнет (рис. 2б). Самые высокие значения рН были обнаружены в верхнем слое (0–0.5 см) колонки (рис. 2б). Несмотря на значительные атмосферные выбросы соединений серы предприятиями “Печенганикель”, во второй половине ХХ в., величина рН не уменьшалась и вариации водородного показателя не превышали естественный фон.
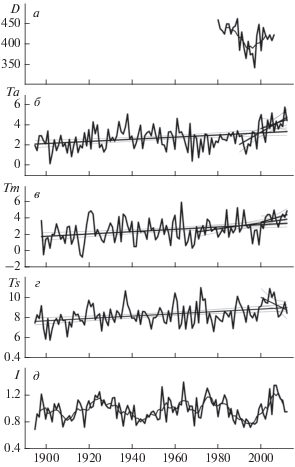
Ниже представлены кривые вариаций общего содержания озона (ОСО) в высоких широтах (>65° с.ш.) [33] (рис. 3а), среднегодовых изменений температуры за сентябрь–ноябрь (рис. 3б), май (рис. 3в) и июнь–август (рис. 3г) по данным норвежской метеостанции Варде, а также вариаций годичного прироста колец деревьев на Кольском п-ове (рис. 3д). За 1895–2012 гг. во все сезоны отмечены тенденции повышения температуры воздуха практически с одинаковой скоростью 0.011–0.012°С · год–1. Тем не менее, в последнем десятилетии скорости роста температуры воздуха, в зависимости от сезона, различаются. Наиболее интенсивное потепление за период 2000–2012 гг. наблюдается в осенний сезон (0.12°С · год–1), наименьшее – в мае (0.074°С · год–1) (рис. 3б, 3в). Летом зафиксирован даже отрицательный тренд, т.е. в последние десять лет в данном регионе происходило снижение температуры воздуха (рис. 3г). Следует отметить, что в осенний период более значительное повышение температуры воздуха (0.12°С · год–1), по сравнению с последним десятилетием, началось еще раньше (с 1990 г.) (рис. 3б). Величина коэффициента положительного тренда для мая 1970–2012 гг. составила 0.038°С · год–1, что почти в два раза меньше скорости роста температуры воздуха в этом месяце в последние десять лет (рис. 3в). На рис. 3д приведены вариации индекса прироста годичных колец деревьев на Кольском полуострове. Начиная с 1900 г. в вариациях годичного прироста наблюдаются максимумы: в 1925 г., 1955–1965 гг., в 1975 г. и в 2008 г. (рис. 3д). Причем величина максимума прироста в последнем десятилетии наибольшая (рис. 3д). Значительный минимум в приросте колец деревьев зафиксирован в 1990–2000 гг. и совпадает по времени с аналогичным минимум в общем содержании озона (до 25%) в высоких широтах (рис. 3а, 3д). Можно видеть, что годичный прирост колец деревьев имеет примерно ту же тенденцию, что и общая численность диатомей N (рис. 2д и 3д).
Рис. 3.
Вариации усредненной концентрации общего содержания озона в высоких широтах (>63° с.ш.) [33] – (а); среднегодовых значений температуры воздуха (ºС) по данным метеостанции Варде (70.4° с.ш., 31.1° в.д.) за сентябрь–октябрь (б), май – (в) и июнь–август – (г); изменения индексов годичного прироста сосны на Кольском п-ове (Кузрека; 66.6° с.ш., 34.8° в.д.) – (д). Тренды температурных изменений с 95%-ными доверительными интервалами изображены тонкими линиями.
ОБСУЖДЕНИЕ
Анализ донных отложений показал, что во второй половине XX века происходили существенные изменения состава и численности диатомовых комплексов озера Раббватнет. Наиболее значительный рост общего количества диатомовых отмечен в последнем десятилетии (рис. 2д). Изменения происходили также в структуре и видовом разнообразии диатомовых комплексов. Индекс видового разнообразия Шеннона–Уивера I испытывал вариации в середине исследуемого периода, тем не менее, его величина не претерпела существенных изменений за последние три десятилетия (рис. 2в). Подобный рост биомассы фитопланктона в различных озерных системах последние десятилетия был отмечен во многих исследованиях [15–18]. В ряде работ утверждается, что основной причиной наблюдаемого диатомового сдвига в пресноводных системах является глобальное потепление температуры воздуха и, как следствие, соответствующее повышение температуры воды [15, 17, 19]. Действительно, данные измерений свидетельствуют об увеличении поверхностной температуры во многих озерах в последние десятилетия, но при этом отмечается, что рост температуры воды в региональных водоемах значительно превышает рост температуры воздуха [20, 22, 23].
Гипотеза глобального потепления климата (“парниковый эффект”), являющаяся предметом оживленной дискуссии в последнее время, базируется на возрастании СО2 в атмосфере за счет, в основном, антропогенных источников. Вместе с тем, величина изменений солнечной радиации ∆I в прошлом столетии по данным о солнечных корональных полях составила ∆I = 1.65 ± 0.23 Вт/м2 [34], по другим оценкам (светимости Солнца в зависимости от площади, занимаемой солнечными пятнами и факелами) – ∆I = 2 Вт/м2 [35]. Приведенные оценки сопоставимы с величиной радиационного воздействия углекислого газа на климат за этот же период, которая, согласно данным МГЭИК, составляет 1.56 Вт/м2 [36]. Такие вариации солнечной радиации могут объяснить до 50% глобального потепления за период 1900–2000 гг. и до 35% роста температуры с 1980 г. по настоящее время [34, 35, 37]. С учетом непрямых воздействий (галактических космических лучей на облачный покров и ультрафиолетовой радиации на химический состав и озоновый слой) вклад солнечной вариабельности в изменения климата может даже возрасти [37–39]. Кроме того, увеличение концентрации СО2 в атмосфере за счет обменных процессов между атмосферой и океаном, может значительно превышать антропогенные выбросы, что и происходило в прошлом на протяжении сотен тысяч лет [40]. Диатомовые водоросли посредством фотосинтеза образуют до 40% органического углерода в океане и, скорее всего, играют решающую роль в процессах обмена и переноса углерода с поверхности в глубокие слои океана [1, 2, 4].
Обычно повышение температуры воды вызывает термофикацию и термическое расслоение водоемов, что, в свою очередь, приводит к появлению сравнительно небольших по размеру диатомовых водорослей, таких как Cyclotella [2]. Эти термофильные диатомеи дискообразной формы с более высоким значением отношения площади поверхности к объему и более низкими скоростями погружения могут сохранять плавучесть в верхних фотических слоях, обеспечивая таким образом доступ к солнечному свету в течение относительно длительного времени [2]. Типичные представители диатомовых комплексов из родов Cyclotella и Aulacoseira приведены на рис. 1а.
Также следует отметить, что другие факторы (pH, антропогенные загрязнения, питательные вещества, солнечная радиация) могут изменять характеристики сообщества диатомовых, В то время как температура остается неизменной [2, 4, 15, 16, 18, 23, 27]. Например, обнаружение факта весеннего цветения фитопланктона под ледяным покровом в арктических озерах, указывает на то, что низкая температура не препятствует росту фитопланктона, и солнечная радиация, по-видимому, играет не менее важную роль в его биопродуктивности, особенно в высоких широтах [2, 4, 41, 42].
Результаты нашего анализа указывают на то, что региональные инструментальные и дендрохронологические данные не демонстрируют значительного потепления в течение последних десяти лет (рис. 3б–3д). Кроме того, в оз. Раббватнет в последние десятилетия уменьшилось количество представителей рода Cyclotella, в то же время численность других, более крупных диатомовых водорослей, например, Aulacoseira alpigena увеличивалась (рис. 2г). Концентрации минеральных веществ в оз. Раббватнет, составляющих основу питания диатомовых комплексов (азот, фосфор, органический углерод) также не претерпели значительных изменений в последнее время и соответствовали средним показателям, характерным для высокоширотных пресноводных экосистем [27] (рис. 2а). Реконструированные вариации рН оз. Раббватнет показали, что в течение всего периода вода характеризовалась почти нейтральными значениями, а колебания величины водородного показателя были незначительными (рис. 2б).
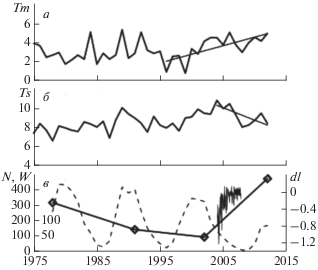
Для более детального анализа ниже приведены вариации температуры в мае Tm (рис. 4а) и летом Ts (рис. 4б) по данным метеостанции Варде, а также изменения общей численности диатомовых в оз. Раббватнет N (рис. 4в) за временной интервал 1975–2012 гг. Видно, что значительное увеличение температуры в мае наблюдалось с 1996 г. (1.8°С/10 лет; R2 = 0.52), т.е. значительно раньше начала увеличения общей численности диатомовых в 2002 г. (384 млн экз./10 лет) (рис. 4а, 4в). Причем до 2002 г. происходило снижение показателя N в оз. Раббватнет (рис. 4в). Отметим, что снижение N наблюдалось на фоне роста майских температур. Что касается летних температур, то в этом случае наблюдается даже отрицательный тренд с 2004 г. (2.6°С/10 лет; R2 = 0.51) (рис. 4б).
Рис. 4.
Вариации среднегодовых значений температуры воздуха (ºС) по данным метеостанции Варде (70.4° с.ш., 31.1° в.д.) за май Tm – (а) и июнь–август Ts – (б); (в) – вариации общей численности диатомовых в оз. Раббватнет N (млн экз./г.) – значения по оси Y слева, среднегодовых значений чисел Вольфа W (штриховая линия) – значения по оси Y справа, изменений интегрального потока ССИ в диапазоне 400–691 нм dI относительно величины 9.11.2007 г. (Вт · м2) – тонкая линия.
Хорошо известно, что условия освещенности (солнечная радиация, продолжительность дня, облачность и ледяной покров, прозрачность водяного столба) имеют очень важное значение для фотосинтеза и, следовательно, могли внести значительный вклад в зафиксированное в последнее время смещение в биомассе диатомовых водорослей в пресноводных экосистемах [2, 4, 16, 18, 23, 41, 42]. Причем, в условиях Арктики степень этого воздействия только возрастает: облачный слой над покрытыми льдом поверхностями может существенно увеличить дозу солнечного излучения на поверхности за счет многократного рассеяния [27]. Диатомовые комплексы активно поглощают солнечное излучение в процессе фотосинтеза, и большинство из них имеет пики поглощения в диапазоне длин волн около ~400 нм (синий свет) и ~680–750 нм (ближнее инфракрасное и инфракрасное излучение) [1, 6]. Об этом свидетельствует строение диатомовых фрустул, приспособленных для улавливания света, что позволяет их использовать в различных светозахватывающих нанотехнологиях [5, 6] (рис. 1б).
УФ-Б излучение (280–315 нм), которое в значительной степени поглощается атмосферным озоном, также влияет на биомассу фитопланктона, подавляя фотосинтез и повреждая дезоксирибонуклеиновую кислоту (ДНК) [33, 43, 44]. Выше было показано, что примерно в 1990–2000 гг. наблюдалось значительное снижение (до 25%) общего содержания озона (рис. 3а), которое сопровождалось снижением общей численности диатомей в оз. Раббватнет (рис. 2д). Известно, что уменьшение ОСО на 10% приводит к увеличению потока УФ-Б на 20%, что, в свою очередь, может привести к гибели некоторых микроорганизмов, в частности фитопланктона и зоопланктона [44]. В работе [45] было показано, что солнечная активность (СА), в основном УФ-излучение, в сочетании с антропогенными воздействиями может значительно повлиять на продуктивность зоопланктона в арктическом оз. Имандра. Мы полагаем, что снижение общей численности диатомовых комплексов в оз. Раббватнет, начавшееся в ~1990 г., было вызвано снижением ОСО и повреждающим действием УФ-Б-излучения.
Интересно, что два максимума в вариациях общей численности диатомовых (в 1965 и 1980 гг.) совпали с максимумами в вариациях индексов годичного прироста колец деревьев (рис. 2д и 3д). В работе [46] при сравнении вариаций солнечной активности и ширины колец деревьев Кольского п-ова, обнаружено, что спектральное солнечное излучение (ССИ) является одним из основных солнечных агентов, влияющих на рост деревьев за Полярным кругом. Недавние прямые измерения на борту спутника Solar Radiation and Climate Experiment (SORCE) показали, что в период с 2004 по 2007 г. на фазе спада 23-го 11-летнего солнечного цикла изменения величины потоков ССИ в видимом и инфракрасном диапазонах спектра были противоположны вариациям в УФ-диапазоне и интегральной солнечной радиации [47, 48]. То есть, согласно данным ИСЗ SORCE, наблюдаемые потоки ССИ в видимом и инфракрасном диапазонах увеличивались на фоне общего снижения уровня солнечной активности.
На рис. 4в приведены вариации чисел Вольфа (W), характеризующие изменения СА, и относительного изменения интегрального потока солнечной радиации спектральном диапазоне 400–691 нм dI данным SORCE [47]. Из рис. 4в видно, что на фоне спада СА в 23-м цикле наблюдается увеличение потока ССИ в видимом диапазоне длин волн, в котором находятся пики поглощения солнечной радиации диатомовыми комплексами [1, 6]. В результате модельных вычислений было показано [49], что наблюдаемое изменение потоков ССИ в данном спектральном диапазоне может являться значимым для состава и температуры атмосферы. В работе [46], при сопоставлении дендрохронологических данных с вариациями СА, сделано предположение, что изменение величины потоков ССИ в видимом и инфракрасном диапазонах могут быть ответственными за радиальный годичный прирост деревьев в высоких широтах во время вековых минимумов СА.
Для более детального изучения наблюдаемых эффектов предполагается провести дополнительные полевые исследования пресноводных экосистем в полярных широтах Евро-Арктического региона для создания палеоклиматических реконструкций при помощи дендрохронологического и диатомового анализа и прогноза глобальных изменений климата.
Таким образом, на основе проведенного анализа мы полагаем, что наблюдаемый в период 2000–2012 гг. значительный рост общей численности диатомовых водорослей в оз. Раббватнет и, по-видимому, в других арктических озерах, вызван зафиксированным ИСЗ SORCE увеличением величины ССИ в видимом и инфракрасном диапазоне длин волн.
ЗАКЛЮЧЕНИЕ
Результаты проведенного анализа позволили сделать вывод о том, что недавний рост общей численности диатомовых водорослей, наблюдаемый в озере Раббватнет, а также в других озерах, может быть обусловлен в значительной степени увеличением фотосинтетически активных потоков спектрального излучения в видимом и инфракрасном диапазонах, зарегистрированных измерениями SORCE. Региональная температура воздуха не являлась основной причиной недавно обнаруженного сдвига в общей численности диатомовых водорослей оз. Раббватнет и, по-видимому, в других арктических озерах. Однако нельзя полностью исключить влияние некоторых других факторов, включая изменение температуры воды в озере. Дальнейшие исследования, касающиеся диатомовых сообществ в арктических озерах, безусловно, необходимы, поскольку откроют новые аспекты их применения в палеоклиматологии и создании природоподобных бионанотехнологий. Учитывая то, что диатомовые водоросли являются источником полезных для человека структурных материалов и биологически активных веществ, полученные результаты важны также для создания биотехнологических производств, основанных на их промышленном культивировании.
Список литературы
Kirk J.T.O. Light and photosynthesis in aquatic ecosystems. Cambridge: Cambridge University Press, 2011. 649 p.
Ruhland K.M., Paterson A.M., Smol J.P. // J. Paleolimnol. 2015. V. 54. P. 1.
Benoiston A.-S., Ibarbalz F.M., Bittner L. et al. // Phil. Trans. R. Soc. B. 2017. V. 372. Art. No. 20160397.
Winder M., Sommer U. // Hydrobiol. 2012. V. 698. P. 5.
Jeffryes C., Campbell J., Li H. et al. // Energy Environ. Sci. 2011. V. 4. P. 3930.
Chen X., Wang C., Baker E., Sun C. // Sci. Rep. 2015. V. 5. Art. No. 11977.
Jamali A., Akbari F., Ghorakhlu M. et al. // Bioimpacts. 2012. V. 2. P. 83.
Ehrlich H., Motylenko M., Sundareshwar P.V. et al. // Adv. Funct. Mater. 2016. V. 26. P. 2503.
Памирский И.Э., Голохваст К.С. // УНЖ. 2012. № 5. С. 4765.
Нехорошев М.В., Рябушко В.И., Железнова С.Н., Геворгиз Р.Г. // РБЖ. 2016. Т. 15. С. 74.
Денисов Д.Б., Даувальтер В.А., Кашулин Н.А. // Тр. Кар. НЦ РАН. 2015. № 9. С. 10.
Adrian R., O’Reilly C.M., Zagarese H. et al. // Limnol. Oceanogr. 2009. V. 54. P. 2283.
Смирнова М.А., Казарина Г.Х., Матуль А.Г., Макс Л. // Океанол. 2015. Т. 55. С. 425; Smirnova M.A., Kazarina G.K., Matul A.G., Max L. // Oceanol. 2015. V. 55. P. 383.
Barron J.A., Stickley C.E., Burky D. // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015. V. 422. P. 85.
Larsen J., Jones V.J., Eide W. // J. Paleolimnol. 2006. V. 36. P. 175.
Deng J., Zhang W., Qin B. et al. // PLOS One. 2018. V. 13. Art. No. e0205260.
Lehnherr I., Louis V.L.S., Sharp M. et al. // Nat. Commun. 2018. V. 9. Art. No. 1290.
Anneville O., Chang C.-W., Dur G. et al. // Oikos. 2019. V. 128. P. 1666.
Elliot J.A. // Glob. Change Biol. 2010. V. 16. P. 864.
Schneider P., Hook S.J. // Geophys. Res. Lett. 2010. V. 37. Art. No. L22405.
Fink G., Schmid M., Wahl B. et al. // Water Resour. Res. 2014. V. 50. P. 2072.
O’Reilly C.M., Sharma S., Gray D.K. et al. // Geophys. Res. Lett. 2015. V. 42. Art. No. 10773.
Schmid M., Koster O. // Water Resour. Res. 2016. V. 52. Art. No. 8103.
Woolway R.I., Dokulil M.T., Marszelewski W. et al. // Clim. Change. 2017. V. 142. P. 505.
Ефремова Т.В., Пальшин Н.И., Белашев Б.З. // Водн. ресурсы. 2016. Т. 43. С. 228; Efremova T.V., Pal’shin N.I., Belashev B.Z. // Water Resour. 2016. V. 43. P. 402.
Даувальтер В.А., Кашулин Н.А., Денисов Д.Б. // Тр. Кар. НЦ РАН. 2015. № 9. С. 62.
Ylikorkko J., Christensen G.N., Kashulin N. et al. Environmental challenges in the joint border area of Norway, Finland and Russia. Rep. 41. Centre for Economic Development, Transport and the Environment for Lapland. Juvenes Print, 2011. 165 p.
Guiry M.D., Guiry G.M., Morrison L. et al. // Cryptogamie Algol. 2014. V. 35. P. 105.
Моисеенко Т.И., Разумовский Л.В. // ДАН. 2009. Т. 427. С. 132; Moiseenko T.I., Razumovsky L.V. // Dokl. Biol. Sci. 2009. V. 427. P. 325.
Канатьев А.Г., Шумилов О.И., Касаткина Е.А. // ПТЭ. 2014. № 2. С. 127; Kanatjev A.G., Shumilov O.I., Kasatkina E.A. // Instrum. Exp. Tech. 2014. V. 57. P. 214.
Cook E.R., Kairiukstis L. Methods of dendrochronology. Dordrecht: Kluwer Academic Publishing, 1990. 340 p.
Holmes R.L. // Tree-Ring Bull. 1983. V. 44. P. 69.
McKenzie R.L., Aucamp P.J., Bais A.F. et al. // Photochem. Photobiol. Sci. 2007. V. 6. P. 218.
Lockwood M., Stamper R. // Geophys. Res. Lett. 1999. V. 26. P. 2461.
Lean J.R., Beer J., Bradley R. // Geophys. Res. Lett. 1995. V. 22. P. 3195.
Solomon S. et al. IPCC, Climate change 2007: the physical science basis. Cambridge: Cambridge Univ. Press, 2007.
Scafetta N., West B.J. // Geophys. Res. Lett. 2006. V. 33. Art. No. L05708.
Svensmark H. // Astron. Geosci. 2007. V. 48. P. 18.
Распопов О.М., Шумилов О.И., Касаткина Е.А. // Биофизика. 1998. Т. 43. С. 902.
Petit J.R., Jouzel J., Raynaud D. et al. // Nature. 1999. V. 399. P. 429.
Sommer U., Lengfellner K. // Glob. Change Biol. 2008. V. 14. P. 1.
Vehmaa A., Salonen K. // Aquat. Ecol. 2009. V. 43. P. 693.
Smith R.C., Prezelin B.B., Baker K.S. et al. // Science. 1992. V. 255. P. 952.
Зуев В.В. Биоиндикация стратосферного озона. Новосибирск: СО РАН, 2006. 228 с.
Shumilov O.I., Kasatkina E.A., Kashulin N.A., Sandimirov S.S. // Caspian J. Env. Sci. 2005. V. 3. P. 139.
Kasatkina E.A., Shumilov O.I., Timonen M. // J. Atmos. Sol. Terr. Phys. 2019. V. 193. Art. No. 105075.
Harder J.W., Fontenla J.M., Pilewskie P. et al. // Geophys. Res. Lett. 2009. V. 36. Art. No. L07801.
Haigh J.D., Winning A.R., Toumi R., Harder J.W. // Nature. 2010. V. 467. P. 696.
Смышляев С.П., Галин В.Я., Блакитная П.А., Лемищенко А.К. // Изв. РАН. Физ. атм. и океана. 2016. Т. 52. С. 19; Smyshlyaev S.P., Galin V.Ya., Blakitnaya P.A., Lemishchenko A.K. // Izv. Atm. Ocean. Phys. 2016. V. 52. P. 16.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия физическая