

Известия РАН. Серия физическая, 2022, T. 86, № 2, стр. 276-281
Рекуррентный анализ потенциала Р300 на одиночных временных рядах ЭЭГ
1 Автономная некоммерческая организация высшего образования
“Университет Иннополис”
Иннополис, Россия
2 Федеральное государственное автономное образовательное учреждение высшего образования
“Балтийский федеральный университет имени Иммануила Канта”
Калининград, Россия
* E-mail: e.pitsik@innopolis.ru
Поступила в редакцию 01.10.2021
После доработки 11.10.2021
Принята к публикации 22.10.2021
- EDN: XEEGIY
- DOI: 10.31857/S0367676522020247
Аннотация
Рекуррентный анализ временных рядов использован для детектирования потенциала P300 по электроэнцефалографических данным. Было показано, что возникновение P300 как результата реакции на стимул связано с повышением сложности электрической активности мозга. Показано, что меры рекуррентного анализа обладают достаточной чувствительностью для того, чтобы детектировать эти изменения даже на отдельных коротких временных рядах.
ВВЕДЕНИЕ
Связанный с событием потенциал мозга (ССП) – это естественная электрофизиологическая реакция мозга на внешнюю стимуляцию или изменения окружающей среды, которая проявляется в виде характерной формы волны электрической активности коры головного мозга, регистрируемой с помощью электроэнцефалографии (ЭЭГ) [1]. Как правило, он наиболее выражен на ЭЭГ сенсорах, покрывающий височный, префронтальный и соматосенсорный отделы коры головного мозга и связан с кратковременной низкочастотной модуляцией динамики локальных нейронных популяций [2].
С одной стороны, ССП отражает свойства нейрональной обработки и рабочей памяти и, следовательно, является одним из широко применяемых методов в фундаментальных исследованиях для проверки функционирования мозга в различных экспериментальных условиях [3, 4]. С другой стороны, будучи неотъемлемым атрибутом реакции мозга на внешнее воздействие, ССП широко используется в качестве источника обратной связи в интерфейсах мозг-компьютер для реабилитации и общения с пациентами, чьи двигательные функции ограничены или полностью утрачены [5–8].
Одиночные сигналы ЭЭГ содержат несколько одновременно измеренных процессов нейронной активности [9]. Традиционно применяется усреднение по достаточному количеству испытаний, чтобы отделить связанные с задачей компоненты ССП от случайных колебания напряжения. Идентификация ССП по небольшому количеству электрофизиологических данных является сложной задачей, в которой инструменты нелинейного анализа данных могут оказаться полезными.
В последнее время вызывает большой интерес применение различных форм рекуррентного анализа к изучению нейрофизиологических данных [10–12]. В ранних работах Марвана и др. авторы продемонстрировали эффективность количественного рекуррентного анализа временных рядов при различении компонентов ССП [13‒15]. С этой целью они использовали меру ламинарности, основанную на оценке вертикальных линий рекуррентных карт, для количественной оценки компонент P300 и N400. В настоящей работе применяется более продвинутый количественный показатель сложности сигнала, основанный на распределении времени возврата, а именно энтропия времени возврата, RTE [16], для решения проблемы обнаружения компоненты P300. Данная мера сложности отлично подходит для анализа изменения характеристик нестационарных процессов [16], переходов от регулярной динамики к хаосу в модельных детерминированных системах [17], а также хорошо зарекомендовала себя при решении задачи по идентификации паттерна моторной активности по сигналам ЭЭГ [18].
Детектирование потенциала P300 проводится в ходе эксперимента тренировке сенсомоторной интеграции при повторяющейся стимуляции испытуемых короткими звуковыми сигналами. На основе анализа многоканальной ЭЭГ, покрывающей всю поверхность коры головного мозга, показано, что локальное увеличение меры сложности RTE в физиологически значимых датчиках ЭЭГ указывает на наличие компоненты P300. Мы ожидаем, что из-за свойств сигнала, к которым чувствительна мера RTE, этот количественный показатель имеет перспективы для идентификации ССП по отдельным отрезкам сигналов ЭЭГ.
МЕТОДЫ
Дизайн эксперимента
Массив данных ЭЭГ записывался в ходе экспериментальной сессии по тренировке сенсомоторной интеграции, во время которой испытуемые были проинструктированы совершать движения руками после звуковых сигналов [19, 20]. При этом короткий звуковой сигнал (250 мс) являлся командой для движения левой рукой, длинный звуковой сигнал (500 мс) – правой рукой. Во время эксперимента испытуемые располагались в удобном кресле, расположив руки на столе перед ними в расслабленном положении, чтобы избежать не связанной с заданием мышечной активности. Каждый испытуемый совершил по 30 движений каждой рукой, в сумме N = 60 движений. Длительность экспериментальной сессии составляла около 10 мин для каждого испытуемого.
Всего в экспериментах приняло участие 13 добровольцев в возрасте 25.5 ± 5.3 лет. Каждый участник подписал информированное согласие на проведение эксперимента. Дизайн экспериментального исследования был одобрен комиссией по этике АНО ВО “Университет Иннополис” и проведен в соответствии с Хельсинской Декларацией.
Сигналы ЭЭГ записывались с помощью электроэнцефалографа-регистратора “Энцефалан ЭЭГР-19/26” (Медиком МТД, Таганрог) с частотой дискретизации 250 Гц. При записи использовалась расширенная расстановка “10–20” из 32 сенсоров. К записанным сигналам был применен режекторный фильтр с частотой отсечки 50 Гц. Также сигналы ЭЭГ были очищены от глазодвигательных и кардио-артефактов с помощью анализа независимых компонент и отфильтрованы в диапазоне 1–40 Гц фильтром Баттерворта 5‑го порядка.
После предварительной обработки, для каждого испытуемого был получен набор из 30 отрезков сигналов ЭЭГ (триалов) продолжительностью 1.5 с, включающие 500 мс до и 1000 мс после короткого звукового сигнала. После визуальной инспекции, из финальной выборки были исключены триалы с артефактами, которые не удалось удалить на этапе предобработки (по 10 триалов для каждого испытуемого).
Рекуррентный анализ
Меры рекуррентного анализа позволяют произвести оценку сложности временных рядов на основе количественного анализа рекуррентных состояний. Пользуясь фундаментальным свойством динамических систем повторять свои состояния во времени, или свойством рекуррентности, мы можем определить рекуррентную матрицу:
(1)
${{R}_{{i,j}}} = \Theta \left( {\epsilon - \left| {\left| {{{x}_{i}} - {{x}_{j}}} \right|} \right|} \right),\,\,\,\,i,\,\,\,\,j = 1 \ldots N,$Визуализацией бинарной матрицы $R$ является рекуррентная диаграмма, состоящая из “черных” (рекуррентных) и “белых” (нерекуррентных) точек. Пример рекуррентной диаграммы, посчитанной для двух временных интервалов ЭЭГ, представлен на рис. 1. Как “черные”, так и “белые” точки образуют структуры, количественный анализ которых предоставляет возможность оценить характеристики сложности сигнала. В данной работе были применены меры детерминизма (DET), ламинарности (LAM) и энтропии рекуррентного времени.
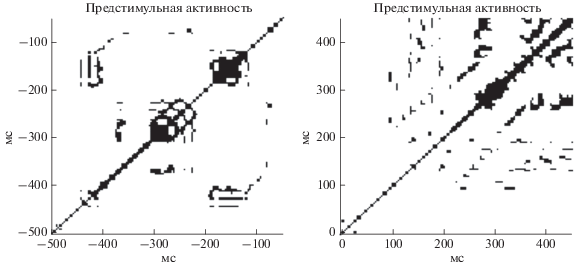
Рис. 1.
Пример рекуррентных диаграмм, посчитанных для двух сегментов временного ряда ЭЭГ: предстимульной активности (500 мс до сигнала) и постстимульной активности (500 мс после сигнала).
Мера детерминизма является оценкой “черных” диагональных линий:
(2)
${\text{DET}} = \frac{{\sum\limits_{l = {{l}_{{min}}}}^N {lP\left( l \right)} }}{{\sum\limits_{l = 1}^N {lP\left( l \right)} }},$“Черные” вертикальные линии оцениваются мерой LAM и характеризуют временной интервал, в течение которого состояние системы не изменялось либо изменялось очень медленно:
(3)
${\text{LAM}} = \frac{{\sum\limits_{\nu = {{\nu }_{{min}}}}^N {\nu P\left( \nu \right)} }}{{\sum\limits_{\nu = 1}^N {\nu P\left( \nu \right)} }}.$Помимо этого, в контексте рекуррентного анализа, важной характеристикой процесса является рекуррентное время или временной интервал, необходимый для возвращения фазовой траектории в окрестность прежде посещенного состояния. Оценить рекуррентное время на РД можно с помощью вертикальных (горизонтальных) “белых” линий:
(4)
${\text{RTE}} = - \frac{1}{{{\text{ln}}{{T}_{{max}}}}}\sum\limits_{{{t}_{w}} = 1}^{{{T}_{{max}}}} {p\left( {{{t}_{w}}} \right)lnp\left( {{{t}_{w}}} \right)} \in \left[ {0,1} \right],$Рекуррентный анализ временных рядов ЭЭГ производился в плавающем окне шириной 50 точек (200 мс) и шагом 2 точки (8 мс). Все меры были скорректированы на уровень предстимульной активности.
Статистический анализ
Значимость представленных результатов была подтверждена с помощью кластерных статистических тестов на основе случайных перестановок (permutation test). Для статистического анализа на сенсорном уровне был применен пространственно-временной кластерный статистический тест [21].
РЕЗУЛЬТАТЫ
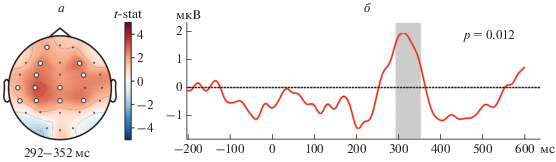
На рис. 2 представлен результат пространственно-временного кластерного теста полученного набора данных (${{t}_{{critical}}}$ = 3.05, ${{p}_{{pairwise}}}$ = 0.005). Из рис. 1а видно, что область значимых изменений сосредоточена в районе левой моторной и височной области с переходом в правую моторную область. При этом кластерный тест выделил значимый временной интервал 292–352 мс, что соответствует локализации ССП P300 (см. рис 1б).
Рис. 2.
Пространственно-временной кластерный тест данных ЭЭГ, соответствующих постстимульной активности, в группе испытуемых: t-статистика по ЭЭГ-сенсорам. Белыми кружками выделены ЭЭГ-сенсоры, вошедшие в значимый кластер (а). Временной ряд ЭЭГ, усредненный по значимому кластеру. Серым выделен интервал времени, в которых отклонения активности от нулевого уровня являются наиболее значимыми (б).
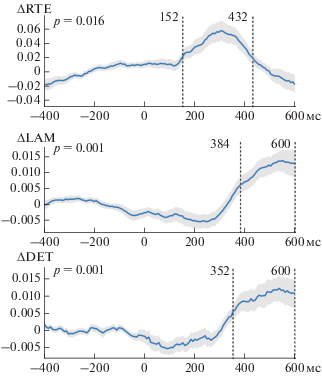
На следующем этапе был произведен рекуррентный анализ временных рядов ЭЭГ в группе испытуемых. На рис. 3 изображены полученные временные зависимости мер ΔDET, ΔLAM и ΔRTE, усредненные по группе сенсоров, входящих в значимый кластер. Отметим, что у временной зависимости ΔRTE присутствует пик в интервале 152–452 мс, что охватывает интервал локализации потенциала P300, в то время как меры ΔLAM и ΔDET имеют пик значимости после 384 и 352 мс, соответственно. Ранее, рост меры ΔDET был связан с возникновением связанной с событием десинхронизации мю-ритма при совершении человеком движений руками [18]. Однако видно, что меры на основе “черных” точек не обладают достаточной чувствительностью для анализа СПП.
Рис. 3.
Временные ряды мер рекуррентного анализа, усредненные по группе людей. Временные ряды представлены со стандартным отклонением (серая полупрозрачная область). Пунктирными линиями выделены временные интервалы с наиболее значимыми отклонениями значений мер от нулевого уровня.
Напротив, мера ΔRTE на основе “белых” линий указывает на связь потенциала P300 с локализованным всплеском сложности сигнала ЭЭГ в соответствующем интервале времени. Увеличение числа длинных “белых” линий, указывающее на увеличение рекуррентных времен, свидетельствует о переходе сигнала ЭЭГ в более сложное, реже повторяющееся состояние.
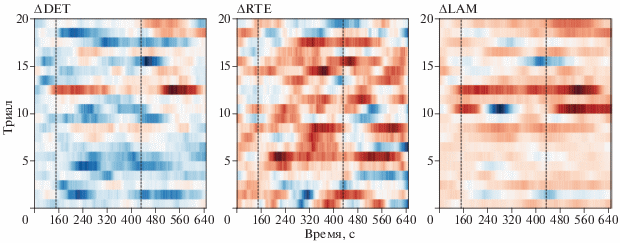
Присутствие статистически значимого интервала времени на мере ΔRTE указывает на воспроизводимость данного эффекта в группе испытуемых. На рис. 4 проиллюстрировано детектирование ССП P300 на отдельных триалах ЭЭГ при усреднении по выявленному на предыдущем этапе значимому кластеру. В то время как меры ΔLAM и ΔDET не показывают значимого результата при детектировании ССП по отдельным триалам, мера ΔRTE имеет выраженный пик, для каждого триала умещающийся в ранее выделенный диапазон 152–452 мс. Данный результат указывает на то, что мера RTE способна успешно детектировать даже тонкие характеристики электрической активности мозга, связанной с ССП P300, возникающие на отдельных триалах ЭЭГ.
Рис. 4.
Рекуррентный анализ одиночных триалов ЭЭГ, соответствующих потенциалу P300. Триады усреднены по значимому кластеру из рис. 2а. Пунктирными линиями обозначен интервал времени 152–432 мс, соответствующий области значительного отклонения меры ΔRTE от нулевого уровня.
ЗАКЛЮЧЕНИЕ
Предложен подход к детектированию потенциалов P300 на основе рекуррентного анализа сложности сигналов ЭЭГ, записанных во время тренировки сенсомоторной интеграции в группе молодых здоровых испытуемых. Была выделена группа сенсоров с наиболее выраженным потенциалом P300, локализующаяся в сенсомоторной области со сдвигом в левую височную долю. Было показано, что мера рекуррентного анализа на основе рекуррентного времени, или “белых” вертикальных (или горизонтальных) линий, обладает высокой чувствительностью к изменениям характеристик сложности сигналов ЭЭГ, связанных с восприятием стимула. В частности, возникновение потенциала P300 связано с увеличением количества “белых” вертикальных линий, что характеризует переход электрической активности мозга из регулярного в более сложное, хаотическое состояние. Помимо этого, мера RTE продемонстрировала высокую чувствительность к данным изменениям даже на одиночных триалах ЭЭГ, что является особенно важным свойством в контексте разработки систем детектирования и классификации активности мозга на основе ССП.
Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований и ГФЕН (проект № 19-52-55001), а также Советом по грантам Президента РФ (проект № НШ-2594.2020.2 для государственной поддержки ведущих научных школ Российской Федерации)
Список литературы
Rugg M.D., Curran T. // Trends Cogn. Sci. 2007. V. 11. No. 6. P. 251.
Luck S.J., Kappenman E.S. The Oxford handbook of event-related potential components. Oxford Univ. Press, 2011. 627 p.
Herrmann C.S., Knight R.T. // Neurosci. Biobehav. Rev. 2001. V. 25. No. 6. P. 465.
Kotchoubey B. // Neurosci. Biobehav. Rev. 2006. V. 30. No. 1. P. 42.
Ганин И.П., Шишкин C.Л., Кочетова А.Г., Каплан А.Я. // Физиология человека. 2012. Т. 38. № 2. С. 5.
Ganin I.P., Shishkin S.L., Kaplan A.Y. // PloS One. 2013. V. 8. No. 10. Art. No. e77755.
Ганин И.П., Каплан А.Я. // ЖВНД им. И.П. Павлова. 2014. Т. 64. № 1. С. 32.
Каплан А.Я. // Физиология человека. 2016. Т. 42. №. 1. С. 118.
Jung T.P., Makeig S., Westerfield M. et al. // Hum. Brain Mapp. 2001. V. 14. No. 3. P. 166.
Храмов А.Е., Фролов Н.С., Максименко В.А. и др. // УФН. 2021. Т. 191. С. 614.
Frolov N.S., Maksimenko V.M., Hramov A.E. // Chaos. 2020. V. 30. Art. No. 121108.
Maksimenko V.A., Frolov N.S., Hramov A.E. et al. // Front. Behav. Neurosci. 2019. V. 13. P. 220
Marwan N., Meinke A. // IJBC. 2004. V. 14. No. 2. P. 761.
Schinkel S., Marwan N., Kurths J. // Cogn. Neurodynamics. 2007. V. 1. No. 4. P. 317.
Marwan N., Groth A., Kurths J. // Chaos Complex. Lett. 2007. V. 2. P. 301.
Webber C.L., Marwan N. Recurrence quantification analysis. Theory and best practices. Springer, 2015. 421 p.
Kraemer K.H., Donner R.V., Heitzig J., Marwan N. // Chaos J. Nonlinear Sci. 2018. V. 28. No. 8. Art. No. 085720.
Pitsik E., Frolov N., Hauke Kraemer K. et al. // Chaos. J. Nonlinear Sci. 2020. V. 30. No. 2. Art. No. 023111.
Frolov N.S., Pitsik E.N., Maksimenko V.A. et al. // PloS One. 2020. V. 15. No. 9. Art. No. e023394.
Pavlov A.N., Pitsik E.N., Guyo G.A. et al. // EPJ Plus. 2021. V. 136. Art. No. 408.
Maris E., Oostenveld R. // J. Neurosci. Meth. 2007. V. 164. No. 1. P. 177.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия физическая