

Известия Русского географического общества, 2020, T. 152, № 1, стр. 15-30
Пространственная структура растительного покрова болот северо-запада Архангельской области (на примере кряжа Ветреный Пояс)
В. А. Смагин a, *, В. К. Антипин b, c, **, М. А. Бойчук c, ***
a Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
b Институт биологии Карельского научного центра РАН
Петрозаводск, Россия
c Национальный парк “Водлозерский”
Республика Карелия, , Россия
* E-mail: smagin@binran.ru
** E-mail: antipin@krc.karelia.ru
*** E-mail: boychuk@krc.karelia.ru
Поступила в редакцию 13.12.2019
После доработки 20.01.2020
Принята к публикации 24.01.2020
Аннотация
Излагаются результаты проведенного в 2017 и 2019 г. изучения болот северо-западной части Архангельской области, расположенных на северном и южном склонах кряжа Ветреный Пояс и на Прибеломорской низменности. Приведена характеристика болот на выделенных формах рельефа модельной территории. Выявлены изменения в пространственной структуре растительного покрова болот. Установлено, что кряж Ветреный Пояс – южная граница распространения болотных массивов дистрофного южноприбеломорского типа. Он же служит и северной границей аапа болот, широко встречающихся начиная с южных склонов кряжа, причем они имеют переходные черты от карельских к северо-восточно-европейским типам. Болота долин центральной части кряжа отличает региональная специфика и при крупномасштабном болотном районировании центральный кряж должен рассматриваться в ранге отдельного болотного района. Кряж Ветреный Пояс – один из самых ярких примеров воздействия орографического фактора на болота, их типологический состав и географическое распределение.
ВВЕДЕНИЕ
Болота северо-запада Архангельской области изучены неравномерно. Ряд районов, как, например, кряж Ветреный Пояс, в этом отношении остается почти не исследованным. Наша цель – закрыть этот пробел, по крайней мере в отношении растительности и типологического разнообразия болот кряжа.
Болота – важнейшие элементы природных ландшафтов кряжа, заболоченность его территории 10–20%. Болота начали формироваться здесь в конце бореального периода голоцена – 8–8.3 тыс. лет назад [8]. Информация о болотах кряжа есть в двух публикациях [1, 8], причем сведения, содержащиеся в первой из них, сжаты и, как выясняется, не вполне достоверны. Более изучены примыкающие к кряжу с севера болота западной части Прибеломорской низменности [7] и болота южных склонов кряжа [2–4, 15]. Целью нашего исследования, проводившегося в 2017 и 2019 гг., было изучение болот и болотной растительности кряжа, выявление воздействия кряжа на географическое распространение типов болот, растительных сообществ и видов растений.
Ветреный пояс – возвышенность с абсолютными высотами 200–347 м и шириной 8–50 км [13], вытянутая с северо-запада на юго-восток на 250 км параллельно Прибеломорской низменности в пределах Республики Карелия и Архангельской области. “Хребет” или центральная часть кряжа представляет собой цепочку гряд. Самые высокие из них – это г. Карбасникова (347 м), Оловгора (344 м), Шуйгора (337 м), Большая Левгора (336 м), Енгишгора (331 м) и Шапочка (320 м).
Северные склоны кряжа крутые и местами обрывистые, резко переходящие в сильно заболоченную Прибеломорскую низменность. Склоны прорезаны руслами коротких бурных рек Малошуйка, Нюхча, Кожа, относящихся к бассейну Белого моря.
Южные склоны пологие, плавно переходящие в равнинные заболоченные ландшафты бассейнов рек Выг, Онега и Илекса. Относительные превышения кряжа над равнинами — 60–80 м в южной части и 100–120 м в северной [5]. В геологическом отношении кряж представляет собой реликт палеопротерозойского рифта, заполненного лавовым материалом в сочетании с вулканогенно-осадочными породами [12, 14]. В протерозойское время здесь происходили тектонические подвижки, сопровождаемые сбросовыми движениями и разломами кристаллического фундамента. Следы наиболее молодых тектонических движений имеются на северо-восточном склоне гряды в виде незначительного (от 2 до 10 м) вертикального перемещения отдельных блоков [9]. По ботанико-географическому делению кряж входит в полосу северотаежных ельников, среди которых преобладают редкостойные воронично-черничные и лишайниково-моховые каменистые типы леса [6, 16].
МЕТОДИКА ИССЛЕДОВАНИЙ
Модельный участок для исследований был выбран на основе анализа современных космических снимков (Bing,\ Landsat 7, Spot 5) и дешифрирования болотной растительности [17]. Он охватывает пространство от побережья Белого моря на севере до северной части национального парка “Водлозерский” (широта оз. Пелозеро) на юге (рис. 1).
Рис. 1.
Район исследований болот кряжа Ветреный Пояс. Условные обозначения: 1–5 –абсолютные высоты: 1 – от 0 до 100 м; 2 – 100–150 м; 3 – 150–200 м; 4 – 200–250 м; 5 – 250–300 м; 6 – линия профиля (см. рис. 2); 7 – границы национального парка “Водлозерский”; 8 – районы полевых исследований; 9 – населенные пункты. Fig. 1. Vetreny Poyas Range mire study area. Symbols: 1–5 – altitude: 1 – 0 − 100 m; 2 – 100–150 m; 3 – 150–200 m; 4 – 200–250 m; 5 – 250–300 m. 6 – profile line (see fig. 2); 7 – Vodlozersky National Park boundaries; 8 –field studies areas; 9 – settlements.
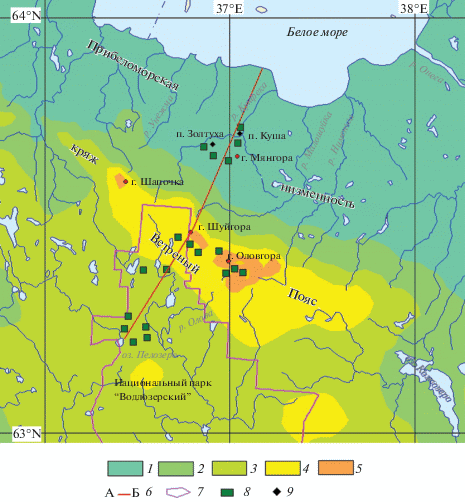
Западная граница модельного участка проходит вблизи горы Шапочка, восточная – близ горы Оловгора. На основании данных топографических карт составлен гипсометрический профиль протяженностью 84 км с абсолютными высотами от 0 м (Белое море) до 327 м (г. Шуйгора) (рис. 2).
Рис. 2.
Гипсометрический профиль модельной территории. Условные обозначения. I–VII – болотные массивы и их системы: I – дистрофные грядово-мочажинные и грядово-озерковые южноприбеломорские; II – мезотрофные сосново-травяно-сфагновые; III – мезотрофные травяно-сфагновые; IV – мезоолиготрофные осоково-пушицево-сфагновые; V – олиготрофные сфагновые грядово-мочажинные и олиготрофные пушицево-сфагновые; VI–VII – олиготрофные сфагновые грядово-мочажинно-озерковые и мезоолиготрофные грядово-мочажинные аапа. Fig. 2. Hypsometric profile of the model territory. Symbols. I–VII – mire complexes and their systems: I – dystrophic ridge-hollow and ridge-pond of SouthWhite Sea area; II – mesotrophic pine-grass-Sphagnum; III – mesotrophic grass-Sphagnum; IV – mesooligotrophic sedge-cotton grass-Sphagnum; V – oligotrophic Sphagnum ridge-hollow and oligotrophic cotton grass-Sphagnum; VI–VII – oligotrophic Sphagnum ridge-hollow-pond and mesooligotrophic ridge-hollow aapa.
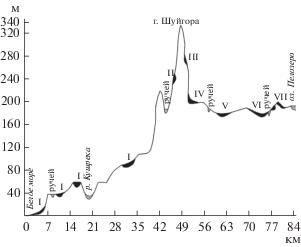
Профиль пересекает террасированную Прибеломорскую низменность (от 0 до 100 м), северные подножье и склоны кряжа (100−200 м), центральную часть кряжа (от 200 до 337 м – г. Шуйгора) и южные склоны кряжа (ниже 200 до 187 м – оз. Пелозеро).
Полевые исследования проводились на модельных болотах выделенных орографических форм кряжа и сопредельной с ним Прибеломорской низменности. Использовались общепринятые в болотоведении геоботанические методы. Описания растительности делались на площадках 10 × 10 м на открытых болотных участках и 20 × 20 м – на лесных. Учитывались формы микрорельефа (гряды, кочки, мочажины, сфагновые ковры и др.) и их растительный покров. В растительных сообществах весь видовой состав сосудистых растений, мхов и лишайников, их проективное покрытие оценивалось в процентах. Выполнено более 100 геоботанических описаний, собран гербарий видов растений и лишайников.
Названия видов сосудистых растений приводятся по [10], мхов – по [19].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Болотные массивы восточной части Прибеломорской низменности
Изучение южноприбеломорских болот проводилось нами к северу от станции и пос. Куша в 2019 г. (см. рис. 1). Все находящиеся здесь массивы слились в огромную болотную систему, занимающую все междуречье рек Куша и Унежма. Территория междуречья представляет собой равнину протяженностью с севера на юг на 8–10 км, приподнятую на 1–60 м над уровнем Белого моря.
Сформированные здесь болота представляют собой эталоны болотных массивов южноприбеломорских дистрофных болот [7, 18]. Почти вся их площадь занята грядово-мочажинными и грядово-озерковыми участками с вересково-водяниково-морошково-лишайниково-сфагновыми грядами, где покрытие лишайников иногда превышает покрытие Sphagnum fuscum (рис. 3).
Рис. 3.
Фрагмент пространственной структуры растительного покрова южноприбеломорского дистрофного болотного массива. В центре – озерко, сформировавшееся на месте деградированной сфагновой мочажины. Fig. 3. Fragment of the spatial plant cover structure of the South White Sea area dystrophic mire complex. A pond on the place of degraded Sphagnum hollow.

Как примесь к сфагнуму бурому часто отмечен S. capillifolium. Кустарничек кассандра (Chamaedaphne calyculata) встречается очень редко и единичными экземплярами. Сфагновые мочажины, в большинстве, заняты шейхцериево- и пушицево-сфагновыми сообществами, реже в травяном ярусе доминируют осока топяная (Carex limosa) и очеретник белый (Rhynchospora alba). Наиболее часто в моховом ярусе преобладают Sphagnum majus и S. lindbergii, чуть реже S. balticum. Изредка и, как правило, с небольшим покрытием, встречаются Sphagnum cuspidatum, S. tenellum, S. rubellum. На окаймляющих гряды узких коврах моховой ярус состоит из Sphagnum papillosum, на коврах местами в травяном ярусе доминирует пухонос дернистый (Trichophorum cespitosum), постоянно встречающийся и на грядах. В мочажинах с деградированным сфагновым покровом в травяном ярусе обычны пухонос дернистый и очеретник. Высокий и крутой склон болота, находящегося к северу от пос. Куша и обращенный к долине р. Кушрека, покрыт сосново-кустарничково-зеленомошными сообществами. Древостой из сосны высотой 8–15 м имеет сомкнутость крон 0.3, в кустарничковом ярусе (покрытие 70%) доминируют багульник и голубика, моховой ярус образует Pleurozium schreberi.
Болота северных подножий и склонов кряжа
Южноприбеломорские дистрофные болота располагаются и на поднимающихся к югу террасированных равнинах северного подножья кряжа, доходя вплоть до северных склонов центрального “хребта” Ветреного Пояса, образуя на ряде террас обширные системы. Нами они исследованы вблизи пос. Золотуха и вокруг возвышенности Мянгора (111 м). Крупные массивы, слившиеся в системы, относятся к одному типу с болотами Прибеломорской низменности. Тем не менее, есть между ними и отличия. Во-первых, уменьшается доля лишайников в моховом ярусе на грядах. Их суммарное покрытие не превышает 20–30%, а часто меньше 5%. На грядах, помимо Sphagnum capillifolium, к доминирующему в моховом ярусе S. fuscum добавляются S. angustifolium и S. rubellum. На грядах по-прежнему отсутствует или очень редко встречается кассандра, однако, в занимающих дренированные склоны сосново-кустарничково-сфагновых сообществах этот вид на равных с голубикой содоминирует с багульником. В травяном ярусе мочажин заметно чаще доминирует очеретник белый, а в моховом ярусе, наряду со Sphagnum majus и S. lindbergii, эту роль выполняет S. cuspidatum. Сказывается и положение болот на террасированных равнинах. Болотные системы тоже становятся террасированными и наряду с дистрофными включают олиготрофные и мезоолиготрофные массивы, располагающиеся на более низких террасах. Пространственная структура дистрофных массивов южноприбеломорского типа остается неизменной. За коротким и крутым склоном с сосново-багульниково-зеленомошно-сфагновыми сообществами следует уплощенный склон с сосново-вересково-морошково-лишайниково-сфагновыми сообществами с низким и разреженным древостоем с заметным кочковато-западинным микрорельефом, где в межкочьях много пухоноса дернистого. Основная часть – ровное платообразное пространство, занятое грядово-мочажинными и грядово-мочажинно-озерковыми участками с крупными и глубокими озерками, и, часто, не менее крупными черными мочажинами с деградированным сфагновым покровом. Для массивов, занимающих в системе нижние террасы, часто характерна вогнутость по центральной оси. Продольный край болота также представляет короткий и резкий подъем с сообществами болотных сосняков. На некоторых массивах на верхней пологой части краевого (бокового) склона вместо низкорослого вересково-сфагнового сосняка располагаются пушицево-морошково-сфагновые (Sphagnum angustifolium) сообщества с обильной голубикой. Далее следуют грядово-мочажинные участки, за которыми и располагается понижение с проточной топью, занятой либо коврово-мочажинными, либо грядово-коврово-мочажинными участками. Впрочем, гряды фрагментированы, имеют вид островков, занимающих 15 и менее процентов площади. Гряды – вересково-морошково-сфагновые (Sphagnum fuscum), сфагновые ковры покрыты пушицево-сфагновыми сообществами (Sphagnum papillosum), мочажины – шейхцериево-сфагновыми сообществами (Sphagnum majus, S. lindbergii). Эти массивы имеют наклонную поверхность, соответствующую уклону занимаемой ими террасы. В приподнятой части располагаются грядово-мочажинные участки с относительно дренированными нетопкими мочажинами, занятыми пушицево-сфагновыми сообществами Eriophorum vaginatum-Sphagnum papillosum (S. balticum). Гряды вересково-морошково-сфагновые (Sphagnum fuscum) с лишайниками, имеющими общее покрытие менее 5%. В нижней подсклоновой части болотного массива располагаются вышеописанные ковровые и коврово-мочажинные участки. Часто в этой части по центральной оси массива протекает ручей с открытым или погребенным руслом, индицируемым либо шейхцериево-, либо очеретниково-сфагновым (Sphagnum cuspidatum и S. lindbergii) сообществом. Проточная топь по центральной оси массива часто имеет выраженное русло ручья. Пушицево-морошково-сфагновые сообщества на умеренно дренированных слабонаклонных участках сближают болота террасированных равнин северного подножья кряжа с долинными массивами возвышенной части кряжа, находящимися у подножья гряд его центрального “хребта”.
В долинах и котловинах между грядами центрального “хребта” кряжа преобладают вогнутые ложбинные олиготрофные болота, на большей части площади пушицево-сфагновые (рис. 4).
Рис. 4.
Фрагмент пространственной структуры растительного покрова олиготрофного пушицево-сфагнового болотного массива, расположенного вблизи г. Шуйгора. Fig. 4. Fragment of the spatial plant cover structure of the oligotrophic cotton grass Sphagnum mire complex located near Mount Shuigora.

Форма болотных массивов обусловлена рельефом долин. Они вытянуты в меридиональном направлении на расстояние до нескольких километров. В классическом варианте их поверхность имеет в поперечнике форму желоба, вогнутого к центру и плавно поднимающегося к краям. Однако далее, по направлению к краям, наблюдаются вариации, зависящие от крутизны окружающих долину склонов. Модельные участки для изучения болот осевой части кряжа были выбраны вблизи вершин Шуйгора и Оловгора.
Наибольшую площадь (не менее 30–40%) массивов занимают участки с пушицево-сфагновыми сообществами, располагающимися в центральной части массивов, причем здесь одинаково часто встречаются сообщества ассоциаций Eriophoro vaginati-Sphagnetum baltici и Eriophoro vaginati-Sphagnetum angustifoli (см. рис. 4). Вторая ассоциация включает упоминавшиеся пушицево-морошково-сфагновые сообщества с обильной голубикой, ранее нами не описанные и заслуживающие отнесения в отдельный синтаксон ранга субассоциации или ассоциации. Сообщества Eriophoro vaginati-Sphagnetum baltici покрывают большую часть центральной пониженной части “желоба”, чередуясь с сообществами Scheuchzerietum palustris, Caricetum limosae, Trichophoro cespitosi-Sphagnetum baltici. Причем наряду со Sphagnum balticum, и чаще его, в моховом ярусе доминируют S. lindbergii и S. majus. Сообщества с пухоносом обычны по краю ручьев. Пушицево-морошково-сфагновые сообщества располагаются выше на слабо покатых частях “желоба”. Они же занимают всю площадь массивов, находящихся на покатых террасах со слабонаклонной поверхностью. О небольшой мощности торфяной залежи на таких массивах свидетельствуют валуны, выступающие на поверхность в их центре. Верхнюю часть склонов “желобов”, в зависимости от наклона, высоты прилегающей возвышенности и минерального богатства стекающих с нее вод, покрывают разные сообщества. Часто это те же пушицево-морошково-сфагновые ассоциации с вкраплениями кочек диаметром несколько метров с кустарничково-сфагновыми (Betula nana, Vaccinium uliginosum, Sphagnum fuscum, S. angustifolium) сообществами. Обращает на себя внимание очень скромная роль Sphagnum divinum (бывший S. magellanicum), доминирующего на таких местообитаниях в южной половине таежной зоны, и “замена”, в сравнении c болотами к югу от кряжа, кассандры на голубику. На склонах, получающих более богатое питание, располагаются сообщества мезоолиготрофной и мезотрофной растительности: ассоциаций Sphagno fallacis-Caricetum rostratae, Sphagno fallacis-Caricetum lasiocarpae, Molinio caerulei – Sphagnetum papillosi. В составе сообществ последней ассоциации молинии содоминируют осока волосистоплодная (Carex lasiocarpa) и пухонос дернистый. Эта ассоциация, главным образом, свойственна грядам аапа болот карельского типа (рис. 5). И болотные участки этого типа встречаются на тех же склонах. На грядах произрастают характерные для аапа болот седмичник (Trientalis europaea), можжевельник сибирский (Juniperus sibirica), мочажины занимают сообщества ассоциации Menyantho-Caricetum limosae с обильными хвощом речным (Equisetum fluviatile) и пузырчаткой промежуточной (Utricularia intermedia). Склоны болот у подножья высоких гряд, например, рядом с возвышенностью Шуйгора, имеющие заметный наклон, занимают можжевелово-молиниево-пушицево-сфагновые сообщества (Sphagnum angustifolium), где обильно произрастают пушица влагалищная, молиния (Molinia caerulea), пухонос дернистый, морошка (Rubus chamaemorus), голубика, вереск (Calluna vulgaris), береза карликовая (Betula nana).
Рис. 5.
Фрагмент пространственной структуры растительного покрова грядово-мочажинно-озеркового травяного и травяно-сфагнового болотного массива карельского аапа типа. На переднем плане – сфагновая гряда с молинией голубой (Molinia caerulea) и осокой волосистоплодной (Carex lasiocarpa). В озерке – вахта трехлистная (Menyanthes trifoliata). Fig. 5. Fragment of the spatial plant cover structure of a ridge-hollow-pond-grass mire complex and a grass-Sphagnum mire complex of Karelian aapa type. A Sphagnum ridge with Molinia caerulea and Carex lasiocarpa is in the foreground; Menyanthes trifoliata is in the pond.

Древесный ярус из сосны высотой 8–15 м сомкнутых крон не образует, при этом выражен ярус подлеска из можжевельника сибирского, создающего своеобразный аспект этим сообществам, простирающимся по склону на сотни метров. Болота понижений центрального “хребта” кряжа находятся на разных гипсометрических уровнях, часто занимая террасированную поверхность. Массивы, находящиеся на разных террасах, сливаясь, образуют каскадные болотные системы, чаще простые, но иногда и сложные, включающие массивы разного типа. Примером сложной, но небольшого размера, может служить система, находящаяся восточнее Шуйгоры. На верхней террасе располагается массив со слабонаклонной поверхностью, занятый описанными выше сообществами с можжевельником, молинией, пушицей и Sphagnum angustifolium. За ним следует более наклоненный участок протяженностью 300–400 м, на котором выражен грядово-мочажинный микрорельеф с растительностью аапа болота, где на грядах располагаются мезоолиготрофные сообщества Pinus sylvestris – Juniperus sibirica – Molinia caerulea – Calluna vulgaris – Rubus chamaemorus – Sphagnum fuscum. Мезотрофные виды, помимо доминирующей в травяном ярусе молинии, представлены немногочисленной пушицей узколистной (Eriophorum angustifolium). Мочажины залиты водой глубиной 0.1–0.2 м. В них располагаются сообщества ассоциации Menyantho-Caricetum limosae без сомкнутого мохового яруса. При этом обращает внимание несоответствие эколого-фитоценотических групп видов травяного и мохового ярусов. Мхи представлены видами олиготрофных и мезоолиготрофных болот – Sphagnum majus и S. compactum. Далее вниз по пологому склону располагается можжевельниково-сфагновое сообщество с разреженным древесным ярусом из сосны. Это сообщество, в свою очередь, сменяется пушицево-морошково-сфагновым сообществом, покрывающим нижний, чуть наклоненный, наибольший по размерам болотный массив системы. Другим примером каскадной болотной системы может послужить система, находящаяся вблизи возвышенности Оловгора. Верхний ее массив на значительной части площади занят грядово-мочажинными участками, ориентированными по уклону террасы. В обширной центральной части располагаются коврово-мочажинные участки, где низкие ковры со Sphagnum papillosum в виде островков окружены мочажинами с S. lindbergii. У этого массива есть склоновый участок с пушицево-морошково-сфагновыми сообществами, но он ориентирован не вверх, а вниз в сторону реки Олова. Грядово-мочажинные участки “обтекают” центр массива и переходят на массив, находящийся на ниже расположенной террасе, где сменяются пушицево-сфагновыми сообществами транзитной топи. Гряды здесь без сосны и лишайников, с моховым покровом из Sphagnum fuscum, с обильным вереском, водяникой и морошкой. Мочажины шейхцериево-сфагновые, на нижерасположенном массиве – пушицево-сфагновые. Наиболее длинные пространственные ряды растительности наблюдаются на склоновых участках, направленных к глубоким понижениям рельефа, занятым обширными обводненными топями. Здесь сверху вниз сменяются участки, занятые можжевельниково-пушицево-сфагновыми, пушицево-морошково-сфагновыми сообществами, на участки обводненных склонов с пухоносово-сфагновыми, пушицево-сфагновыми и осоково-сфагновыми сообществами. У уплощающихся подножий склона появляются грядово-мочажинные (топяные) участки, которые ниже в глубоких понижениях рельефа переходят в топь с коврово-мочажинным микрорельефом с низкими сфагновыми коврами, имеющими форму гряд. В этих топях формируются истоки рек. Склоновые участки спускаются к этой топи с разных сторон, отличаясь только по углу наклона, протяженности и числу звеньев пространственного ряда растительности. Здесь в мочажинах и топях неоднократно, в том числе, в роли доминанта отмечены приатлантические виды сфагновых мхов: Sphagnum tenellum, S. compactum. И на грядах, и в мочажинах в травяном ярусе часто доминирует пухонос дернистый.
В верхней половине склоновой части болота в нескольких местах отмечены разных размеров разрывы торфяной залежи, залитые водой или с открытой поверхностью торфа, где произрастает лишь осока вздутая (Carex rostrata). Они явно образовались из-за разрыва замерзшего торфа мощным потоком стекающих по склону талых вод. В нижних частях болот есть примеры, свидетельствующие в пользу растительного происхождения гряд, когда на ровном, слабонаклонном участке с пушицево-сфагновым сообществом пушица образует заметные полосы, перпендикулярные наклону поверхности болота.
На склонах кряжа вблизи Шуйгоры в средней его части обнаружено склоновое ключевое болото, питаемое напорными грунтовыми водами, но небогатого минерального состава. Об этом свидетельствует видовой состав сообществ, не включающий ни одного вида флоры ключевых местообитаний. Болото на несколько сотен метров вытянуто сверху вниз, при этом вместе с высотой снижается и его трофический уровень – от мезоевтрофного до олиготрофного. На находящихся выше по склону мезоевтрофных участках древостой (6Б4Е) высотой 5–7 м очень разрежен (сомкнутость крон 0.1). Травяной ярус образует хвощ речной, а моховой покров на равных слагают сфагновые и гипновые мхи: Sphagnum angustifolium, S. fallax, Rhizomnium pseudopunctatum, Warnstorfia exannulata. Средняя часть болота покрыта описанными выше сообществами с можжевельником. В их составе хвощ речной и молиния встречаются в небольшом количестве, а к нижнему краю массива исчезают и они: набор видов сводится к произрастающим на олиготрофных болотах, а моховой покров состоит исключительно из Sphagnum angustifolium. Выходы ключей отмечены во многих местах на склоновых участках болот как в виде родников с открытой водой, так и в виде затянутых сфагновым мхом блюдцеобразных топей диаметром до 20 м. Виды флоры ключевых болот богатого питания не обнаружены. В моховом покрове доминируют Sphagnum fallax, S. angustifolium, S. riparium, S. lindbergii. В травяном ярусе на первый план выходят “обычные” виды мезотрофных болот, но редко выполняющие роль доминантов: пушица узколистная, осока заливная (Carex paupercula). Ручьи, берущие начало в ключах, стекая вниз, пересекают склоновые, в том числе, грядово-мочажинные участки, имея четко выраженное русло. Характерной особенностью болот района является нахождение грядово-мочажинных участков на склонах, направленных вниз не от выпуклого центра массива к краю, а на снисходящих вниз участках склоновых болот.
Болота южных склонов кряжа Ветреный Пояс, в основных чертах, типичны для сильно заболоченных регионов северной тайги, но положение на склонах возвышенности, перехватывающей атмосферные осадки, террасированный рельеф и сложная геологическая история делают их разнообразными по режиму водно-минерального питания и растительному покрову. Нами они изучались на территории вокруг оз. Пелозеро [17].
По занимаемой площади преобладают олиготрофные сфагновые грядово-мочажинные болота (рис. 6).
Рис. 6.
Фрагмент пространственной структуры растительного покрова олиготрофного сфагнового грядово-мочажинного болотного массива. На переднем плане – кочка с морошково-пушицево-сфагновым (Sphagnum fuscum) сообществом. Она находится в пушицево-сфагновой мочажине (S. balticum). Окружают мочажину кустарничково-морошково-сфагновые гряды. Fig. 6. Fragment of the spatial plant cover structure of the oligotrophic Sphagnum ridge-hollow mire complex. A hummock with a cloudberry-cotton grass-Sphagnum (Sphagnum fuscum) community is in the foreground; placed in a cotton grass-Sphagnum (S. balticum) hollow. The hollow is surrounded by dwarf-shrub-cloudberry-Sphagnum ridges.

По совокупности признаков они отнесены нами к северо-восточноевропейскому географическому подтипу [3, 15, 18]. Для них характерна окраинная сосново-кустарничково-сфагновая полоса, сменяемая на нижней части склона кочковато-ковровыми участками с пушицево-сфагновыми (Sphagnum angustifolium) коврами и кустарничково-сфагновыми (S. fuscum), большей частью с сосной, кочками. Далее по направлению к центру массива следуют грядово-мочажинные и грядово-мочажинно-озерковые участки. Спецификой района вокруг оз. Пелозеро, затрудняющей рассмотрение пространственной структуры растительности грядово-мочажинных болот, является сильная его заболоченность. Болотные массивы образуют обширные сложные системы. Олиготрофные сфагновые грядово-мочажинные, мезотрофные травяно-сфагновые и аапа болота объединяются в системы, где различить ранг граничащих болотных выделов очень сложно – болотные ли это участки или отдельные массивы. По характеру растительности, главным образом растительности гряд, массивы южных склонов кряжа отличаются и от южноприбеломорских болот, и от болот подножья центрального “хребта” кряжа. От обоих вариантов их отличает отсутствие в большинстве сообществ вереска, встреченного нами лишь несколько раз и в очень небольшом количестве. Кроме того, здесь наблюдается почти стопроцентная встречаемость кассандры с высокими показателями проективного покрытия. От южноприбеломорских болот рассматриваемые болота отличает отсутствие или незначительное количество лишайников в моховом покрове. Ярус болотной сосны отличительным признаком служить не может, так как в половине описанных сообществ отсутствует, а в другой половине вполне выражен. Объединяющими болота Прибеломорской низменности, осевой части кряжа и его южных склонов признаками служат: постоянство и обилие голубики, карликовой березы, морошки, водяники (Empetrum nigrum), постоянство клюквы мелкоплодной (Oxycoccus microcarpus), что характерно для сообществ олиготрофных гряд в северной тайге. Главным отличием в растительности мочажин является редкость Sphagnum lindbergii, отмеченного нами в роли доминанта лишь один раз. Реже встречаются очеретник белый и пухонос дернистый, но по этому признаку отличия не столь резкие.
Ряд олиготрофных болотных массивов не достиг грядово-мочажинной стадии. Пространственный ряд растительности на них укорочен и завершается кочковато-ковровыми участками с преобладанием пушицево-сфагновых сообществ ковров. Они располагаются по всему центру массивов и в верхней части склонов.
На одной из приозерных наклонных террас отмечено олиготрофное болото с заметной вогнутостью к центру, по составу и пространственному размещению растительных сообществ похожее на олиготрофные болотные массивы, распространенные в долинах осевой части кряжа. Здесь также от края массива наблюдается направленный вниз склон, участки которого занимают пухоносово-сфагновые сообщества с покрытием пухоноса дернистого до 30%. В отличие от “желобообразных” массивов центрального “хребта”, на большей части площади сравнительно дренированных, здесь склон переходит в обширную обводненную топь, занимающую основную площадь массива. Она похожa на описанную выше топь глубоких понижений между гребнями “хребта”, куда направлен сток с нескольких склоновых болот. Для этой топи характерен коврово-мочажинно-озерковый микрорельеф, где островки-ковры диаметром 0.5–1.5 м покрыты пухоносово-сфагновыми сообществами с моховым ярусом из Sphagnum balticum, S. papillosum при участии S. compactum. Здесь же в роли доминанта один раз встретился Sphagnum tenellum. В мочажинах и озерках значительную площадь занимают очеретниково-сфагновые и очеретниково-печеночниковые сообщества, на других болотах этого района мало заметные. Здесь же располагаются сообщества ассоциаций Caricetum limosae, Scheuchzerietum palustris. Долинные олиготрофные массивы вогнутой формы являются “визитной карточкой” кряжа Ветреный Пояс: они встречаются от террас северного подножья до южных склонов кряжа, преобладая в его возвышенной центральной части.
Встречающиеся на южных склонах кряжа мезотрофные болота можно подразделить на осоково-сфагновые, занимающие наклонные долины, часто соседние с теми, где располагаются упомянутые выше вогнутые олиготрофные болота, и на структурированные грядово-мочажинные. Между этими разновидностями болот наблюдаются переходные формы, например, склоновые мезотрофные болота, где ровная ковровая поверхность местами разделена на сектора мочажинами-протоками. Осоково-сфагновые болота покрыты однообразными сообществами ассоциации Sphagno-fallacis-Caricetum rostratae. На участках со слабой проточностью болотных вод доминируют осока вздутая и пушица влагалищная, что индицирует процесс перехода массива на олиготрофную стадию развития. Мезотрофные грядово-мочажинные болота аапа типа встречаются здесь часто и занимают большие площади. Однако они, входя в одну сложную систему с олиготрофными грядово-мочажинными, плавно переходят друг в друга, что не позволяет понять, являются ли они самостоятельными массивами сложной болотной системы или обширными участками единого болотного массива. По растительности аапа болота южных склонов кряжа, в большинстве своем, следует отнести к карельскому типу. Встречены и аапа, имеющие признаки онежско-печорского типа, где моховой покров гряд образует Sphagnum divinum (magellanicum), травяной ярус состоит из осок волосистоплодной и вздутой и вахты. Основная же часть аапа болот соответствует признакам карельского типа, где в травяном ярусе гряд доминирует или значимо присутствует молиния, постоянно встречается седмичник. На грядах моховой ярус в большинстве сообществ образует Sphagnum papillosum, реже – S. fuscum. Однако признаки карельского типа здесь заметно утрачиваются, что проявляется не только в редукции диагностической группы видов до 2–3, но и в очень редкой встречаемости такого характерного для них вида как вереск. В мочажинах располагаются очеретниково-вахтовые, шейхцериево-вахтовые и топяно-осоково-вахтовые сообщества со сфагновым покровом и без него.
Аапа болота южных склонов кряжа различаются не только географическими типами, но и по трофности: наряду с мезотрофными встречаются и мезоевтрофные, перемежаемые участками (или массивами) евтрофных болот ключевого питания. Они в большей степени соответствуют, по растительности, болотам аапа карельского типа. Моховой ярус гряд, наряду с Sphagnum papillosum, образуют Sphagnum warnstorfii, S. centrale и внесенный в Красную книгу Архангельской области Sphagnum subfulvum [12]. Помимо молинии, седмичника, можжевельника сибирского и обоих видов пухоноса, на грядах отмечены плаунок (Selaginella selaginoides) и осока свинцово-зеленая (Carex livida). Залитые слоем воды мочажины заняты сообществами ассоциации Menyantho-Caricetum limosae без сплошного мохового яруса с обильной пузырчаткой промежуточной. На одном из мезоевтрофных болот аапа и на грядах, и в мочажинах обильно произрастает тростник (Phragmites australis), придающий им вид тростникового болота. В мочажинах этих аапа отмечены мхи Loeskypnum badium (вид Красной книги Архангельской области [12]), Sphagnum platyphyllum, Limprichtia scorpioides, местами образующие моховой ярус.
На болота евтрофного типа здесь приходится наименьший процент площади. Тем не менее, они имеются и разнообразны по растительности и типу минерального питания. Они делятся на болота лимногенного, поверхностного и напорного грунтового питания [20]. К первой группе относятся приречные и приозерные болота, питаемые речными и разливающимися весной озерными водами. Они имеют небольшие размеры и покрыты сообществами высокотравья, осоковыми, хвощовыми, вейниковыми и тростниковыми, принадлежащими к ассоциациям Comaro-Caricetum rostratae, Comaro-Caricetum lasiocarpae, Comaro-Equisetetum fluviatilis, Comaro-Phragmitetum australis, соседствующими с сообществами лесного болота ассоциации Thyselii palustris-Betuletum pubescentis. В их составе обильна ива лапландская (Salix lapponum). Болота поверхностного питания располагаются на высоких склонах речных долин и входят в болотные системы как болотные участки либо массивы. Описанные нами такие болота представляют продолжение слабонаклонных мезотрофных болот. По-видимому, помимо стекающих вниз по склону болотных и дождевых вод, в питании склонового болота участвуют и грунтовые воды. Они выклиниваются из-под склона долины. На большей части площади это лесные евтрофные болота с древесным ярусом высотой 8–12 м, сомкнутостью крон 0.3–0.4; преобладающий состав древостоя 8Б2С+Е. Травяной ярус образуют вейник сероватый (Calamagrostis canescens), хвощ речной, осока волосистоплодная, вахта и сабельник (Comarum palustre). О том, что болото подпитывают и грунтовые воды, свидетельствуют виды сосудистых растений ключевых болот: змеевик большой (Bistorta major), подмаренник топяной (Galium uliginosum). Моховой ярус образуют Sphagnum squarrosum, S. centrale, S. warnstorfii. В местах интенсивного стока располагаются открытые осоковые участки с сообществами асс. Caricetum diandrae. В болотной системе, включающей мезоевтрофные аапа, между массивами этого типа встречен участок ключевого питания – крупный ключевой бугор, где отмечены щавель кислый (Rumex acetosa), змеевик большой, грушанка круглолистная (Pyrola rotundifolia); тем не менее, доминируют здесь обычные виды евтрофных болот (осока волосистоплодная, сабельник, вахта), а в моховом ярусе – Sphagnum angustifolium с примесью S. warnstorfii. Участок с выклиниванием напорных грунтовых вод обнаружен и в центре одного из олиготрофных массивов, что вносит большой вклад в его биоразнообразие и разнообразие микроландшафтной структуры. Диаметр участка – несколько сотен метров; его занимает сообщество лесного мезоевтрофного болота, в составе которого отмечены редкие виды сфагнов Sphagnum aongstroemii, S. inundatum [17], а на отходящем от него склоне находится грядово-озерковый комплекс, характерный для болот аапа типа.
ЗАКЛЮЧЕНИЕ
В северо-западной части Архангельской области, пересекаемой кряжем Ветреный Пояс, на относительно коротком расстоянии в 84 км ярко проявляется широтная дифференциация типов болотных массивов и растительных сообществ. Происходит смена в пространственной структуре болотной растительности за счет резких изменений в рельефе. Простирающаяся на пути океанических воздушных масс возвышенность перехватывает значительную часть переносимых ими осадков, способствуя развитию болот.
В пределах исследованной нами восточной части Прибеломорской низменности также как и западной, доминируют болотные массивы дистрофного южноприбеломорского типа. Они занимают здесь до 80% территории, формируя болотный ландшафт. На террасированных равнинах северного подножья кряжа Ветреный Пояс все крупные болотные системы простого строения состоят из болотных массивов того же типа. Изменения в составе растительных сообществ на этом пространственном диапазоне минимальны: несколько уменьшается доля лишайников в моховом покрове гряд, чаще встречается кассандра. Она же становится обильной в составе облесенных болотных участков на дренированных краях массивов. Появляются террасированные болотные системы, включающие вогнутые олиготрофные массивы с транзитной топью в центральной части.
Большинство болотных массивов возвышенной центральной части кряжа относится к олиготрофным долинным, вогнутым пушицево-сфагновым болотам, образующим протяженные системы, включающие обширные склоновые олиготрофные болота. Реже встречаются олиготрофные грядово-мочажинные болотные массивы, на грядах которых роль лишайников минимальна и в моховом покрове безраздельно господствует Sphagnum fuscum. Кассандра здесь встречается редко и в небольшом количестве. На небольшой площади встречены аапа болота, относящиеся к карельскому и онежско-печорскому типам. К специфике возвышенной части кряжа следует отнести и склоновые ключевые болота, питаемые небогатыми грунтовыми водами.
Типологическое разнообразие болотных массивов и произрастающих на них растительных сообществ заметно возрастает на южных склонах кряжа. Преобладают по площади олиготрофные грядово-мочажинные болота, в большей степени соответствующие северо-восточноевропейскому типу. На них редок вереск и, наоборот, постоянно встречается и обильна кассандра. Становятся более редкими Sphagnum capillifolium (на грядах), S. lindbergii, S. tenellum (в мочажинах). Наряду с олиготрофными, большую площадь занимают и минеротрофные грядово-мочажинные болота аапа типа, представленные разными географическими типами и трофическими вариантами. Можно предположить, что по южному склону кряжа проходит северная граница распространения северо-восточноевропейских сфагновых грядово-мочажинных олиготрофных и аапа болот.
На значительно меньшей площади, но во многих местах, встречаются евтрофные болотные массивы, включая питаемые напорными грунтовыми водами. Благодаря этому видовое и фитоценотическое богатство болот южных склонов кряжа заметно выше, чем в его осевой части и на северном подножье.
Кряж Ветреный Пояс – один из самых ярких примеров воздействия орографического фактора на болота, их типологический состав и географическое распределение. По кряжу проходит ряд границ зональных типов болот, синтаксонов и видов болотной растительности.
Таким образом, кряж Ветреный Пояс служит южной границей распространения омбротрофных болот прибеломорского типа, северной границей распространения олиготрофных грядово-мочажинных и минеротрофных аапа болот. Центральную, наиболее возвышенную часть кряжа следует считать отдельным регионом при районировании болот, со свойственным ей типом олиготрофных долинных болот вогнутой, жолобообразной формы, включающих склоновые участки.
Работа выполнена при финансовой поддержке гранта РФФИ 17-04-01749, а также в рамках государственных заданий Ботанического института им. В.Л. Комарова РАН (АААА-А19-119030690058-2), Института биологии Карельского НЦ РАН (АААА-А19-119062590056-0), Национального парка “Водлозерский” (№ 051-00036-19-01).
Список литературы
Абрамова Т.Г., Кирюшкин В.Н. Районирование болот Архангельской области // Северо-Запад европейской части СССР. Л.: Изд-во ЛГУ, 1969. С. 81–113.
Антипин В.К., Елина Г.А., Токарев П.Н., Бразовская Т.И. Болотные экосистемы национального природного парка “Водлозерский”: прошлое, настоящее, будущее // Бот. журн. 1996. Т. 81. № 1. С. 21–37.
Антипин В.К., Бойчук М.А., Бразовская Т.И., Талбонен Е.Л. Растительный покров болот национального парка “Водлозерский” // Национальный парк “Водлозерский”: природное разнообразие и культурное наследие. Петрозаводск: КНЦ РАН, 2001. С. 135–144.
Антипин В.К., Бойчук М.А. Сфагновые сообщества с Molinia caerulea (Poaceae) на онежско-печорских аапа болотах // Бот. журн. 2004. Т. 89. № 2. С. 244–251.
Бискэ Г.С. Четвертичные отложения и геоморфология Карелии. Петрозаводск: Гоc. изд-во Карельской АССР, 1959. 307 с.
Геоботаническое районирование Нечерноземья европейской части РСФСР. Л.: Наука, 1989. 64 с.
Елина Г.А. Типы болот Прибеломорской низменности // Болота Карелии и пути их освоения. Петрозаводск, 1971. С. 51–79.
Елина Г.А., Юрковская Т.К. Болотные экосистемы низкогорий северной тайги // Болотные экосистемы Европейского Севера. Петрозаводск: Карельский филиал АН СССР, 1988. С. 5–24.
Корсаков А.К., Лукашенко С.В., Межеловский А.Д. Этапы магматизма в истории формирования Ветреного Пояса (Балтийский щит) // Актуальные проблемы геологии докембрия, геофизики и геоэкологии: материалы XXIII молодежной научной школы-конф., посв. памяти К.О. Кратца. Петрозаводск: КНЦ РАН, 2012. С. 15–16.
Кравченко А. В. Конспект флоры Карелии. Петрозаводск: КНЦ РАН, 2007. 403 с.
Красная книга Архангельской области. Архангельск: Ком. по экологии Арханг. обл., 2008. 351 с.
Куликова В.В., Бычкова Я.В., Куликов В.С., Терновой А.Н. Новые данные по минералогии коматиитовых базальтов палеопротерозойского рифта Ветреный Пояс // Геодинамика, магматизм, седиментогенез и минерагения Северо-Запада России. Петрозаводск: КНЦ РАН, 2007. С. 214–219.
Куликов В.С., Куликова В.В. Ветреный Пояс // Карелия: энциклопедия. Т. 2. Петрозаводск: ИД “ПетроПресс”, 2009. С. 117–118.
Куликов В.С., Бычкова Я.В., Куликова В.В. Ветреный Пояс: тектоно- и петротип палеопротерозоя юго-восточной Фенноскандии // Геология Карелии от архея до наших дней: материалы Всерос. конф., посв. 50-летию Института геологии КНЦ РАН. Петрозаводск: КНЦ РАН, 2011. С. 91–103.
Кутенков С.А. Болотные экосистемы // Природа и историко-культурное наследие Кожозерья. Архангельск: УрО РАН, 2006. С. 39–50.
Природа и историко-культурное наследие Кожозерья. Архангельск: УрО РАН, 2006. 310 с.
Смагин В.А., Антипин В.К., Бойчук М.А. Болота южных склонов кряжа Ветреный Пояс (Восточная Фенноскандия) // Бот. журн. 2019. Т. 104. № 4. С. 602–616.
Юрковская Т.К. География и картография растительности болот Европейской России и сопредельных территорий. СПб., 1992. 256 с.
Ignatov M.S., Afonina O. M. Checklist of mosses of East Europe and North Asia // Arctoa. 2006. V. 15. P. 1–130.
Mazing V. Mire typology of the Estonian SSR // Some aspects of the botanical research in the Estonian SSR. Tartu, 1975. P. 97–108.
Дополнительные материалы отсутствуют.
Инструменты
Известия Русского географического общества