

Известия Русского географического общества, 2020, T. 152, № 3, стр. 3-16
Концепция подводного ландшафта
К. М. Петров *
Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: k.petrov@spbu.ru
Поступила в редакцию 27.06.2020
После доработки 19.09.2020
Принята к публикации 20.09.2020
Аннотация
Комплексное изучение береговой зоны моря – одна из актуальных задач современной географии. Подводное ландшафтоведение в России опирается на теоретические положения отечественной географической школы В.В. Докучаева–Л.С. Берга. Развитию морских ландшафтных исследований в XX в. способствовало появление аквалангов и технических средств дистанционного зондирования. В статье обсуждаются общие положения концепции подводного ландшафта, дается его определение, рассматриваются особенности морфологического строения и основные ландшафтообразующие факторы. Ландшафт береговой зоны моря определяется как относительно однородный участок подводного берегового склона, обладающий однотипным геологическим строением, рельефом, грунтами, гидроклиматом (температурой, соленостью, течениями, волновыми процессами) и однотипными группировками гидробионтов. Он характеризуется определенным сочетанием внутриландшафтных природных комплексов (фаций, угодий), представляющих собой особые биотопы донных биоценозов. Ландшафты береговой зоны моря охватывают прибрежную сушу, подверженную воздействию прибоя (супралитораль), окраину суши, заливаемую во время прилива (литораль) и подводный береговой склон до глубины, где воздействие волновых процессов затухает и освещенность недостаточна для формирования сообществ макрофитов (сублитораль). Подводный ландшафт рассматривается как основная исходная единица ландшафтного районирования морских мелководий. Программы работ, разработанные с учетом названных положений, обеспечат полноту и сравнимость исследований, выполненных разными авторами. Концепция подводного ландшафта иллюстрируется примером результатов изучения Притаманского ландшафта в северо-восточной части Чeрного моря.
Береговая зона и шельф – это часть географической оболочки, богатая и разнообразная природа которой формируется в результате активного взаимодействия гидрометеорологического, геолого-геоморфологического и биогидроценотического звеньев физико-географического процесса [13]. Развитие ландшафтоведения береговой зоны и шельфа в России опирается на теоретические положения отечественной географической школы В.В. Докучаева–Л.С. Берга. Крупным событием начала 60-х гг. XX в. стало создание сотрудниками Зоологического института АН СССР под руководством Е.Ф. Гурьяновой и Г.У. Линдберга карт подводных ландшафтов Южного Сахалина и южных Курильских островов [4, 9]. Применение методов аэрофотосъемки дна морских мелководий и комплекса морских и подводных работ для дешифрирования аэрофотоизображений морского дна открыли новые возможности в изучении береговой зоны моря [2, 11]. Несмотря на важность аэрофотометодов, их применение в СССР было ограничено из-за режима секретности как аэросъемки, так и распространения еe материалов. Дальнейшему развитию морских ландшафтных исследований способствовало появление и широкое использование аквалангов [1, 6–8]. В начале XXI в. большие перспективы изучения береговой зоны и шельфа Мирового океана открылись благодаря современным методам дистанционного зондирования (космические изображения высокого разрешения, аэросъемки с беспилотных летательных аппаратов – дронов).
Развитие ландшафтных исследований в береговой зоне и на шельфе требует четкого определения понятия подводный ландшафт, его морфологической структуры и ландшафтообразующих факторов, а также рассмотрения подводного ландшафта как основной исходной единицы ландшафтного районирования морских мелководий. Программы работ, построенные с учетом названных требований, могут обеспечить полноту и сравнимость результатов исследований, выполненных разными авторами. Ландшафты береговой зоны моря (подводного берегового склона) обладают сложной природой, но вместе с этим наиболее доступны для изучения и картографирования.
При написании статьи использованы материалы экспедиций 1960-х гг., проводимых Лабораторией аэрометодов АН СССР (ныне Всероссийский научно-исследовательский институт аэрокосмических методов, ВНИИКАМ). Научный руководитель экспедиции В.В. Шарков, начальник геоморфологического отряда З.И. Гурьева, начальник гидробиологического отряда К.М. Петров. Морские исследования включали эхолотное и геолокационное профилирование, а также взятие образцов грунта с помощью дночерпателя и драги. Подводные исследования проводились автором, они включали описание облика донных природных комплексов, подводное фотографирование, отбор образцов бентоса и горных пород.
ПОДВОДНЫЙ ЛАНДШАФТ, ЕГО ПРИРОДНЫЕ ОСОБЕННОСТИ И СТРУКТУРА
Подводный ландшафт береговой зоны моря (ПЛБЗ) – относительно однородный участок береговой зоны, качественно отличающийся от других участков своей структурой, т.е. с однотипным геологическим строением, рельефом, грунтами, гидроклиматом (температурой, соленостью, течениями, волновыми процессами) и с однотипными группировками гидробионтов. ПЛБЗ – наименьшая исходная таксономическая единица ландшафтного районирования, он не пересекается границами таксонов более высокого ранга, относится к одной природной зоне, лежит в пределах верхнего пояса неритического яруса, ограничен одной региональной морфоструктурой [14]. Каждый подводный ландшафт характеризуется своеобразным сочетанием внутриландшафтных природных комплексов (фаций, угодий), представляющих собой биотопы донных биоценозов. ПЛБЗ охватывают прибрежную сушу, подверженную воздействию прибоя – супралитораль, заливаемую во время прилива – литораль и морское дно до глубины, где воздействие волновых процессов затухает и освещенность недостаточна для формирования сообществ макрофитов – сублитораль (верхний пояс неритического яруса). Глубже ПЛБЗ сменяются ландшафтами элиторальной зоны (среднего пояса неритического яруса).
Благодаря положению ландшафтов береговой зоны на рубеже суши и моря их формирование происходит при активном взаимодействии компонентов основных геосфер: атмосферы, гидросферы, литосферы, биосферы. К природным факторам, обусловливающим богатство и разнообразие ландшафтов морских мелководий, относятся:
– связь гидрологических условий с метеорологическим режимом атмосферы, выраженный сезонный ритм природных процессов;
– подвижность вод, контролирующая процессы абразии, литодинамики и аккумуляции осадков, а также способствующая хорошей аэрации, притоку питательных веществ и разносу зачатков организмов;
– проникновение солнечной радиации, поддерживающей фотосинтез фитопланктона и фитобентоса;
– разгрузка речного стока, вызывающая сильную изменчивость солености морских вод, их обогащение биогенными и органическими веществами; твердый сток принимает основное участие в питании вдольберегового потока наносов;
– большое видовое разнообразие и богатство жизненных форм, способствующие высокой плотности заселения всевозможных экологических ниш на поверхности моря, в толще воды, на поверхности дна и в грунте; высокая биологическая продуктивность;
– влияние плейстоценовой регрессии, с которой связаны реликтовые формы рельефа и фации донных отложений, разорванные ареалы организмов и влияние голоценовой трансгрессии, с которой связана молодость подводных ландшафтов.
Отметим основные свойства подводных ландшафтов:
– ландшафт морского дна обособляется на участке земной коры, имеющем в общем одинаковое геологическое строение; как правило, он связан с развитием одной региональной морфоструктуры;
– каждому ландшафту свойственен определенный набор литологических разностей современных донных отложений или выходов коренных пород, контролирующих характер скульптурных микро- и мезоформ подводного рельефа;
– подводная освещенность, температура и волновые процессы изменяются с глубиной, что обусловливает вертикальное подразделение береговой зоны;
– разнообразие форм рельефа, грунтов, гидрологических обстановок определяют пестроту биoтопов и соответственно разнообразие донных биоценозов.
Все это служит основанием для выделения системы морфологических единиц внутриландшафтной дифференциации и применения ландшафтно-биономического метода исследования структуры бентоса.
Донные природные комплексы (ДПК), связанные с характерными формами рельефа, грунтами и группировками гидробионтов, как правило, обладают своеобразным внешним видом, поэтому в ландшафтоведении они получили название морфологических единиц внутриландшафтного подразделения. Характеристика морфологической структуры ПЛБЗ моря включает описание единиц горизонтального и вертикального подразделений.
Основными единицами горизонтального подразделения ПЛБЗ являются фация и угодье.
Подводная фация – наименьший элементарный ДПК. Она представляет конкретный биотоп, связанный с одной формой микрорельефа или одним элементом мезорельефа (вершина, склон, подножье банки), и расположена в определенном интервале глубин. Фация сложена одной литологической разностью современных осадков или приурочена к однородному по вещественному составу выходу горных пород и занята одним биоценозом. Комплекс фаций образует подводное угодье.
Подводное угодье – это ДПК, связанный с определенной мезоформой рельефа, обладающий хорошо выраженными границами. Дифференцированное развитие локальных структур определяет формирование угодий двух типов. На структурах, испытывающих поднятие, происходит размыв дна, и формируются угодья абразионно-скульптурного типа: вдольбереговой пояс скал, подводные банки и рифы вдали от берега. У структур, испытывающих опускание, происходит седиментационное выравнивание дна и формирование угодий аккумулятивного типа. На участках активной литодинамики подводные угодья представлены береговыми валами, подводными косами и т.п. В спокойной гидродинамической обстановке на участках устойчивого осадконакопления происходит формирование угодий монотонных песчано-илистых равнин.
Важным экологическим фактором названных угодий выступают свойства грунта. Хорошо известны сообщества гидробионтов-обрастателей, связанных с каменистыми грунтами, и сообщества организмов, закапывающихся в рыхлый грунт, в облике которых находят отражение приспособительные признаки, обусловленные образом жизни на разных типах грунта.
Вертикальная дифференциация морских мелководий отражает высоту воздействия прибоя, ритмику приливо-отливных явлений, ослабление волнения и угасание подводной освещенности с глубиной. Под влиянием названных факторов находятся процессы рельефообразования, осадконакопления, а также вся совокупность экологических условий, определяющих распределение донных биоценозов. Батиметрический профиль служит основой сопряженного ряда ДПК. Закономерной смене с глубиной подвержены и фации, и угодья, и целые ландшафты.
Главными единицами подразделения ПЛБЗ по глубине являются вертикальные зоны. Предлагается выделять в верхнем поясе неритического яруса три вертикальные зоны: супралитораль, литораль (псевдолитораль), сублитораль [10]. Глубже простирается элиторальная зона, относящаяся к среднему поясу неритического яруса.
Следующую единицу, на которую зоны делятся по вертикали, предлагается именовать этажом. В супралиторали они отражают высоту воздействия прибоя; в литорали – ритмы прилива и отлива, в сублиторали – ослабление воздействия волн на дно и угасание подводной освещенности. Иногда внутри этажей выделяются ступени, отличающиеся по составу донных биоценозов. Величина интервалов вертикальных подразделений меняется с глубиной от сантиметров до метров; в целом система единиц вертикального подразделения ПЛБЗ напоминает пружину, сжатую в начале и растянутую на конце.
Представление о морфологических единицах занимает особое место в учении о подводных ландшафтах. Именно они служат непосредственным объектом морских и подводных исследований и картографирования. В результате анализа закономерных пространственных сочетаний морфологических единиц выделяются и сами ландшафты.
РАЙОН ИССЛЕДОВАНИЯ И МЕТОДЫ КАРТОГРАФИРОВАНИЯ ПОДВОДНЫХ ЛАНДШАФТОВ
Район исследования занимает подводный склон Таманского полуострова на северо-востоке Чeрного моря: на западе он ограничен Керченским проливом, на востоке – Анапской пересыпью. Это материковая отмель шириной до 40–60 км, которая выделяется в качестве Притаманского подводного ландшафта [12]. Морское дно сложено отложениями миоцен-плиоценового возраста, состоящими из рыхлой толщи глин, песков с прослоями и включениями плотносцементированных пород мергеля, бурого железняка, известняка. Подводная абразионно-аккумулятивная равнина осложнена несколькими брахиантиклинальными складками. Многочисленные банки, рифы, гряды и т.п. приурочены к куполам антиклиналей и крыльям антиклинальных складок. Там, где дно сложено легко разрушающимися породами, возникают пространства абразионного выравнивания [3].
Покров современных морских осадков в Притаманском подводном ландшафте обладает незначительной мощностью, местами он прерывается. Ограниченность выноса пролювиального терригенного материала с суши объясняется отсутствием на Таманском полуострове развитой гидрографической сети. Характерная особенность Притаманского ландшафта – широкое распространение обширных выровненных пространств, покрытых маломощным слоем ракуши. У мыса Железный рог в результате абразии надрудных песчаных слоев на подводном береговом склоне формируются аккумулятивные формы рельефа.
В Чeрном море, где приливы практически не выражены, выделяется псеводлитораль, где земноводный режим поддерживается омыванием кромки берега волнами. Мелководье Притаманского ландшафта на глубине до 20–25 м относится к сублиторальной зоне. На глубинах более 25 м формируется ландшафт илисто-ракушечной равнины, относящийся к элиторальной зоне.
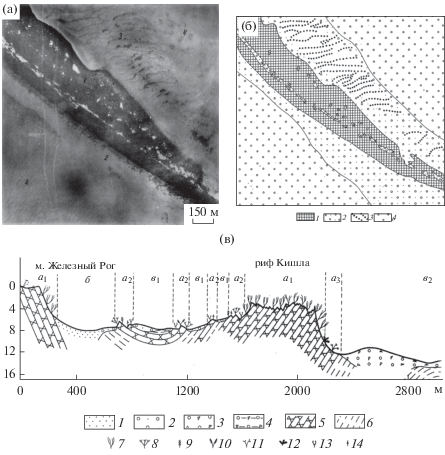
Основным методом изучения и картографирования Притаманского ландшафта служило дешифрирование аэрофотоснимков (АФС) морских мелководий, опирающееся на комплекс морских, геофизических и подводных исследований [2, 11]. Притаманский ландшафт характеризуется значительной шириной мелководья, доступного для аэрофотографирования. Изображение морского дна было получено для большей части абразионно-аккумулятивной равнины, на удалении до 6–7 км от берега. Разнообразие рисунков на АФС морского дна связано с угодьями скал и камней, и их сочетаниями с угодьями полей ракуши. Например, рассмотрим дешифровочные признаки подводного рифа Кишла, образованного пластами бурого железняка. Здесь водоросли, обрастающие скалы, придают изображению рифа на АФС темный тон, позволяющий распознать его очертания на фоне светлого тона аккумулятивной равнины (рис. 1а). С помощью простейшего преобразования изображение может быть переведено в графическую форму и представлено в виде карты (рис. 1б). Более полное представление о расположении подводных угодий, отображенных на АФС и карте, о подводной растительности и грунтах дает ландшафтный профиль (рис. 1в).
Рис. 1.
Притаманский подводный ландшафт, акватория рифа Кишла: а) аэрофотоснимок: 1 – подводный риф, образованный мощными пластами бурых железняков рудного горизонта, обросших бурыми водорослями Treptacantha (Cystoseira) barbata и др., 2 – выровненные пространства, покрытые песчано-ракушечными наносами, 3 – положительные песчано-ракушечные формы волновой аккумуляции, 4 – поля ракуши; б) ландшафтная карта: 1 – угодье скал и камней, биоценоз Treptacantha (Cystoseira) barbata, 2 – выровненные пространства, покрытые песчано-ракушечными наносами, 3 – положительные песчано-ракушечные формы волновой аккумуляции, 4 – поля ракуши; в) ландшафтный профиль: 1–6 – грунты: 1 − песок, 2 − детритный песок с примесью ракуши, 3 − ракуша (танатоценоз створок мидий, устриц, гребешка и др.), 4 − заиленная ракуша, 5 − камни и скалы (устойчивые к разрушению пласты бурого железняка), 6 − глины в коренном залегании; 7–14 – доминанты растительных сообществ: 7 − Treptacantha (Cystoseira) barbata, 8 − Nereia filiformis, 9 − Chondria capillaris, 10 − Dasya baillouviana, 11 − Ceramium virgatum, 12 − Phyllophora crispa, 13 – Dictyota (Dilophus) fasciola, 14 – Laurencia obtusa; а–в – подводные угодья: а – скалы и камни, б – песчаные равнины, в – поля ракуши.
Fig. 1. The Taman underwater landscape, water area of Kishla reef: a) aerial picture: 1 – underwater reef formed by thick layers of brown ironstone ore horizon overgrown with brown algae Treptacantha (Cystoseira) barbata, etc., 2 – leveled spaces covered with sand-shell sediments, 3 – positive sand-shell forms of wave accumulation, 4 – shell fields; б) map of landscape: 1 – landform of rocks and stones, biocenosis of Treptacantha (Cystoseira) barbata, 2 – leveled spaces covered with sand-shell deposits, 3 – positive sand-shell forms of wave accumulation, 4 – shell fields; в) profile of landscape: 1–6 – deposits: 1 – sand, 2 – detritus sand with an admixture of shells, 3 – shells (thanatocoenose of the valves of mussels, oysters, scallops, etc.), 4 – silted shells, 5 – stones and rocks (resistant to abrasion layers of brown iron ore), 6 – clay in situ; 7–14 – dominants of phytocoenoses: 7 – Treptacantha (Cystoseira) barbata, 8 – Nereia filiformis, 9 – Chondria capillaris, 10 – Dasya baillouviana, 11 – Ceramium virgatum, 12 – Phyllophora crispa, 13 – Dictyota (Dilophus) fasciola, 14 – Laurencia obtusa; a–с – underwater landforms: а – stones and rocks, б – sandy plains, в – shell fields.
ПРИТАМАНСКИЙ ПОДВОДНЫЙ ЛАНДШАФТ, СЕВЕРО-ВОСТОК ЧЁРНОГО МОРЯ
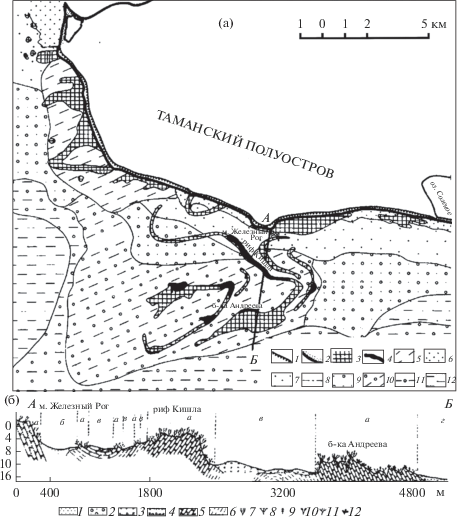
Характерные черты морфологической структуры Притаманского подводного ландшафта отражены на составленной в результате дешифрирования АФС ландшафтной карте (рис. 2а). Взаимное отношение подводных угодий, показанных на карте, отображено на ландшафтном профиле (рис. 2б). Приведем описание основных типов подводных угодий: 1) скалы и камни, 2) выходы коренных глин, 3) поля ракуши, 4) песчаные равнины.
Рис. 2.
Биономическая структура Притаманского ландшафта: а) ландшафтная карта, линия АБ – место ландшафтного профиля. Условные обозначения: 1 − абразионные берега с активным клифом, 2 − аккумулятивные берега с песчаным пляжем; 3–12 – формы рельефа, определяющие подводные угодья: 3–5 – абразионно-скульптурные формы (3 − скалы и камни, сочетающиеся с полями ракуши, 4 − четко выраженные банки и рифы, 5 − обнажения коренных глин), 6–8 – песчаные равнины сублиторали (6 − верхний этаж, среднезернистые пески с примесью ракуши, 7 − средний этаж, слегка заиленные тонкозернистые пески, 8 − нижний этаж, заиленные тонкозернистые пески с примесью ракуши), 9–11 – поля ракуши сублиторали (9 – верхний этаж, детритные пески с примесью ракуши, 10 − средний этаж, ракуша, 11 − нижний этаж, заиленная ракуша), 12 − илисто-ракушечные равнины элиторали; б) ландшафтный профиль: 1–6 – грунты: 1 − песок, 2 − детритный песок с примесью ракуши, 3 − ракуша (танатоценоз створок мидий, устриц, гребешка и др.), 4 − заиленная ракуша, 5 − камни и скалы (устойчивые к разрушению пласты мергеля, известняка, бурого железняка), 6 − глины в коренном залегании; 7–12 – доминанты растительных сообществ: 7 − Treptacantha (Cystoseira) barbata, 8 − Nereia filiformis, 9 − Chondria capillaris, 10 − Dasya baillouviana, 11 − Ceramium virgatum, 12 − Phyllophora crispa; а–г – подводные угодья: а – скалы и камни, б – песчаные равнины, в – поля ракуши, г – заиленная ракуша.
Fig. 2. Bionomic structure of the Taman underwater landscape: а) map of landscape, line AB – location of the landscape profile. Symbols: 1 – abrasive coasts with an active cliff, 2 – accumulative coasts with a sandy beach; 3–12 – the forms of relief that define underwater landforms: 3–5 – the abrasive-sculptural forms (3 – rocks and stones combined with shell fields, 4 – clearly defined banks and reefs, 5 – outcrops of clay in situ); 6–8 – the sandy plains in sublitoral (6 – the upper floor, medium-grained sands with an admixture of shell, 7 – the middle floor, slightly silty fine-grained sand, 8 – the lower floor, silty fine-grained sands with an admixture of shell), 9–11 – the shell fields in sublitoral (9 – the upper floor, detritus sand with an admixture of shells, 10 – the middle floor, shells, 11 – the lower floor, silted shells), 12 – the silty-shell plain in elitoral; б) profile of landscape: 1–6 – deposits: 1 – sand, 2 – detritus sand with an admixture of shells, 3 – shells (thanatocoenosis of mussels, oysters, scallops, etc.), 4 – silted shell, 5 – stones and rocks (resistant to abrasion layers of marl, limestone, brown iron ore), 6 – clay in situ; 7–12 – dominants of phytocoenoses: 7 – Treptacantha (Cystoseira) barbata, 8 – Nereia filiformis, 9 – Chondria capillaris, 10 – Dasya baillouviana, 11 – Ceramium virgatum, 12 – Phyllophora crispa; а–г – underwater landforms: а– rocks and stones, б – sandy plains, в – shell fields, г – silted shell.
Угодья скал и камней – абразионно-скульптурные формы рельефа, связанные с обнажением пластов крепко сцементированных пород и отторгнутыми от них обломками. Они формируются на месте локальных структур, испытывающих поднятие. Отпрепарированные пласты горных пород образуют банки и гряды, изгибающиеся в зависимости от простирания пластов, что определяет своеобразие рисунка угодий данного типа на ландшафтной карте (рис. 2а, условные знаки 3 и 4). Мощные рифы длиной до 20 км образуются на месте пластов бурого железняка.
Угодья скал и камней представляют особый биотоп, занятый донными биоценозами гидробионтов-обрастателей. В фитобентосе господствуют заросли бурой водоросли цистозиры (Cystoseira). Зообентос образован разнообразными беспозвоночными животными, обрастающими скалы. С угодьями скал и камней связана группировка нектобентоса – рыб каменистых грунтов.
Угодья скал и камней занимают сублиторальную зону на абразионно-аккумулятивной равнине Притаманского ландшафта на глубинах до 20–25 м. Ослабление освещенности и уменьшение воздействия волн обусловливает вертикальную дифференциацию сублиторали, занимаемой угодьем скал и камней, на три этажа.
Верхний этаж от нуля глубин до 3–5 м характеризуется сообществом многолетней бурой водоросли Cystoseira flaccida. Под покровом водорослей на камнях в массе селится, образуя щетки, моллюск Mytilaster lineatus.
Средний этаж на глубине от 3–5 до 10–15 м занят пышными зарослями многолетней бурой водоросли Treptacantha (Cystoseira) barbata, определяющей характерные черты типичного для Черного моря биоценоза цистозиры. Биоценозы цистозиры состоят из средиземноморских вселенцев, первым из которых выступает сама цистозира. На глубине до 6–7 м распространены сообщества Treptacantha (Cystoseira) barbata–Cladostephus spongiosus f. verticillatus–Ellisolandia elongata (=Corallina mediterranea) с хорошо развитой синузией эпифитов (рис. 3). На глубине от 6–7 до 10–11 м распространены сообщества Treptacantha (Cystoseira) barbata–Phyllophora crispa–Gelidium spinosum с обедненной синузией эпифитов.
Рис. 3.
Фитоценоз Treptacantha (Cystoseira) barbata + Cystoseir flaccida–Cladostephus spongiosus f. verticillatus–Ellisolandia elongata (Corallina mediterranea) – синузия эпифитов, вершина банки Аксенова, глубина 7 м (подводная фотография).
Fig. 3. Phytocoenosis Treptacantha (Cystoseira) barbata + Cystoseir flaccida–Cladostephus spongiosus f. verticillatus–Ellisolandia elongata (Corallina mediterranea) – synusia of epiphytes, the top of Aksenov Bank, depth 7 m (underwater photography).

Биоценоз цистозиры содержит все характерные жизненные формы зообентоса угодья скал и камней. Особая группировка связана с кронами зарослей цистозиры, это микрооброст (диатомовые одноклеточные водоросли, мелкие ракообразные, мелкие полихеты, личинки крупных ракообразных, моллюсков и полихет), а также эпифитные макрофиты и беспозвоночные (брюхоногие моллюски, полихеты, двустворчатые моллюски, изоподы, амфиподы и декаподы). Микрооброст служит фундаментом, на котором зиждется пищевая цепь всех видов животных цистозирового сообщества; его основную часть составляют диатомовые водоросли, обладающие высокой продуктивностью [5]. Формирование сессильного зообентоса происходит в процессе конкурентной борьбы за свободные каменистые поверхности. На глубинах, доступных сомкнутым зарослям водорослей, зообентос плохо развит: здесь он занимает преимущественно нижние поверхности нависающих глыб. По мере увеличения глубины плотность зарослей водорослей уменьшается и доминирующую роль на поверхности камней начинают играть сообщества обрастателей. Характер фитобентоса средней сублиторали заметно изменяется при уменьшении приподнятости скульптурных форм над дном, сложенным рыхлыми осадками. В этом случае на поверхности каменистого субстрата образуется тонкий слой наилка, препятствующего расселению цистозиры. Подводная растительность претерпевает существенные изменения: на смену зарослям цистозиры приходят сообщества нитевидных бурых и красных водорослей с доминированием Chondria capillaris, Ceramium virgatum, Nereia filiformis, Dasya baillouviana и корковой водоросли Zanardinia typus.
Нижний этаж сублиторали на глубине от 10–15 до 20–25 м занят сообществом многолетней красной водоросли Phyllophora crispa. На подводном склоне Таманского полуострова сообщества филлофоры, в силу неблагоприятного рельефа для ее расселения, уступают место сообществам нитевидных красных водорослей Dasya baillouviana, Chondria capillaris, Carradoriella elongata. Здесь фитобентос играет подчиненную роль, а фауна обрастателей занимает господствующее положение.
Угодья выходов коренных глин – скульптурные поверхности, выработанные в толще коренных глин. В Притаманском ландшафте эти угодья связаны с толщей миоцен-плиоценового возраста. Они распространены на глубине 10–15 м, образуя значительные по площади обнажения, лишенные покрова рыхлых наносов (см. рис. 2а, условный знак 5).
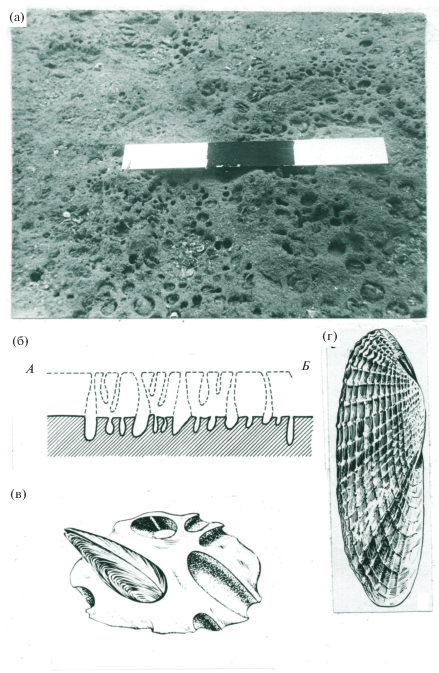
С глинами связан биоценоз моллюсков-сверлильщиков – фолад (Pholas dactylus). Поверхности выходов глин источены до такой степени, что напоминают пчелиные соты (рис. 4): на 1 м2 дна приходится более 5000 отверстий, а плотность живых моллюсков достигает 2600 экз./м2. Входные отверстия нор имеют разный диаметр – от 1 до 10 мм.
Рис. 4.
Биоценоз моллюска-сверлильщика (Pholas dactylus): (а) ноздреватая от ходов моллюсков поверхность коренных глин, глубина 11 м (подводная фотография); (б) схема, иллюстрирующая процесс внедрения моллюсков в породу и ее разрушение (линия АБ – исходная поверхность); (в) моллюск-сверлильщик фолас в куске породы; (г) створка Pholas dactylus, хорошо видны острые сверлящие скульптурные ребра раковины.
Fig. 4. Biocoenosis of the drill shell (Pholas dactylus): (а) the porous surface of the clay in situ worn down by molluscs, depth 11 m (underwater photography); (b) scheme illustrating the process of introducing shells into the rock and its destruction (line AB is the original surface); (c) Pholas dactylus in the piece of rock; г) Pholas dactylus, clearly visible sharp sculpted edges of the shell.
Угодья полей ракуши – абразионно-аккумулятивная равнина, покрытая ракушей. В Притаманском ландшафте это угодье приурочено к глубинам 10–15 м и связано с накоплением аллохтонных раковин моллюсков преимущественно мидий и устриц, обрастающих подводные скалы и камни (см. рис. 2а, условные знаки 9–11). Раковины моллюсков, обрастающих скалы, после отмирания осыпаются и разносятся волнами, образуя танатоценоз – поля ракуши на выровненных пространствах дна (рис. 5).
Рис. 5.
Угодье полей ракуши: (а) поверхность поля ракуши на глубине около 10 м, виден характерный состав танатоценоза (подводная фотография); (б) образец грунта, поднятый драгой с глубины 10 м; видны створки мидий и устриц, а также галька и щебень коренных пород.
Fig. 5. Landform of the shell fields: (а) the surface of the shell field at a depth of about 10 m, a typical composition of thanatocoenosis is visible (underwater photography); (b) deposit’s sample raised by the dredge from a depth of 10 m; mussels and oysters, as well as pebbles and gravel bedrock are visible.

Как уже отмечалось, ракуша представляет танатоценоз моллюсков, обрастающих скалы и камни. Дополнительным источником раковин служат местные биоценозы. Господствующую роль в последних играют двустворчатые моллюски (Gouldia minima, Divaricella divaricata, Pecten ponticus, Tapes sp., Venus gallina и др.), а также брюхоногие моллюски (Nassa, Cerithium, Gibbula, Rapana, Calyptrea). В состав зообентоса входят также крабы Porcellana, Portunus, раки-отшельники (Diogenes). Поля ракуши служат стацией представителя древнейших хордовых – ланцетника Branchiostoma (Amphioxus) lanceolata.
Угодья песчаных равнин в Притаманском ландшафте образуются в результате аккумуляции продуктов абразии толщи песков надрудного горизонта (см. рис. 2а, условные знаки 6–8). Их внешний облик характеризуется однообразным строением, лишенным резких морфологических отличий. Затухание с глубиной воздействия волновых процессов на дно обусловливает обогащение песчаных осадков алевролитовой и пелитовой фракциями, а также автохтонным ракушечным материалом. Поступление аллохтонной ракуши происходит вблизи выходов скальных пород, на которых обитают сессильные моллюски. С угодьями песчаных равнин связан биотоп, занятый донными биоценозами псамофильных гидробионтов, обитающими на поверхности дна и закапывающимися в грунт, а также группировкой нектобентоса – рыбами песчаных грунтов. Фауна угодий песчаных равнин образована преимущественно широко распространенными и наиболее эврибионтными средиземноморскими видами.
Ослабление воздействия волн на дно обусловливает вертикальную дифференциацию сублиторали угодья песчаных равнин на три этажа.
Верхний этаж простирается до глубины 3–5 м. Характерно формирование подводных береговых валов. По дну бегают, подбирая детрит, многочисленные раки-отшельники Diogenes pugilator; в песок закапываются двустворчатые моллюски-фильтраторы Donax julianae.
Средний этаж – до глубины 10–15 м. Дно ровное с микроформами рельефа волнового происхождения, песчаные отложения мелкозернистые, слегка заиленные. На поверхности дна по-прежнему многочисленны раки отшельники, в толще песка обильны полихета-пескожил (Arenicola marina) и двустворчатые моллюски-фильтраторы (Venus gallina, Spisula subtroncata, Divaricella divaricata).
Нижний этаж протягивается до глубины 20–25 м, где воздействие волн на дно постепенно затухает. Дно ровное сложено заиленным алевритовым песком со значительной примесью автохтонной ракуши. Бентос образован сообществами двустворчатых моллюсков-фильтраторов (Venus gallina, Meretrix rudis), возрастает роль пелофильных гидробионтов – полихет (Nereis longissimi, Nephthys hombergi).
Содержащаяся в описаниях ДПК характеристика бентоса, полученная в 1960-х гг., в настоящее время, когда Притаманский ландшафт вошел в зону активного гидротехнического строительства, может служить критерием для оценки состояния донных биоценозов и экологической обстановки в береговой зоне в целом.
ВЫВОДЫ
1. Методологической основой изучения и картографирования подводных ландшафтов служат теоретические положения географической школы В.В. Докучаева–Л.С. Берга.
2. Развитие подводного ландшафтоведения связано как с внедрением новой техники подводных исследований и дистанционного зондирования морских мелководий, так и с дальнейшей разработкой теории ландшафтов береговой зоны моря.
3. Биоцентрический подход к описанию донных природных комплексов позволяет на их основе осуществлять мониторинг экологического состояния прибрежных акваторий.
Автор выражает искреннюю благодарность старшему научному сотруднику Лаборатории экологии прибрежных донных сообществ Института океанологии им. П.П. Ширшова РАН, кандидату биологических наук У.В. Симаковой за актуализацию видовых названий водорослей.
Список литературы
Беспалова Л.А. Экологическая диагностика и оценка устойчивости ландшафтной структуры Азовского моря. Ростов-на-Дону: Изд-во Ростовского ун-та ООО “ЦВВР”, 2006. 271 с.
Гурьева З.И., Петров К.М., Рамм Н.С., Шарков В.В. Геолого-геоморфологическое изучение морских мелководий и берегов по материалам аэрофотосъемки. Методическое руководство. Л.: Наука, 1968. 371 с.
Гурьева З.И., Петров К.М., Шарков В.В. Аэрофотометоды геолого-геоморфологического исследования внутреннего шельфа и берегов морей. Атлас аннотированных аэрофотоснимков. Л.: Недра, 1976. 227 с.
Гурьянова Е.Ф. Теоретические основы составления карт подводных ландшафтов // Вопросы биостратиграфии континентальных толщ. Тр. Третьей сессии Всесоюзного палеонтологического о-ва. М.: Госгеолтехиздат, 1959. С. 52–61.
Маккавеева Е.Б. Биоценоз Cystoseira barbata // Тр. Севастопольской биологической станции. Севастополь: АН СССР, 1959. Т. 12. С. 168–191.
Мануйлов В.А. Подводные ландшафты залива Петра Великого. Владивосток: Изд-во ДВГУ, 1990. 148 с.
Митина Н.Н., Чуприна Е.В. Подводные ландшафты Черного и Азовского морей: структура, гидроэкология, охрана. М.: Типография Россельхозакадемии, 2012. 320 с.
Пенно М.В. Развитие научных представлений о морских ландшафтах // Экополитика и геоэкодинамика регионов. 2014. Т. 10. № 1. С. 166–172.
Петров К.М. Рецензия на Атлас океанографических основ рыбопоисковой карты Южного Сахалина и южных Курильских островов. Т. I. Карты распределения кормовых и поисковых организмов. Л., 1955. Т. II. Карты подводных ландшафтов. Л., 1956. Отв. ред. издания Е.Н. Павловский // Изв. ВГО. 1958. № 5. С. 480–481.
Петров К.М. Вертикальное распределение подводной растительности Черного и Каспийского морей // Океанология. 1967. Т. 7. Вып. 2. С. 170–178.
Петров К.М. Подводные ландшафты: теория, методы исследования. Л.: Наука, 1989. 128 с.
Петров К.М. Ландшафтно-биономический подход к картографированию и районированию бентали морских мелководий // Изв. РАН. Сер. геогр. 2003. № 1. С. 33–44.
Петров К.М. Развитие комплексных исследований подводных ландшафтов // Изв. РАН. Сер. геогр. 2014. № 1. С. 100–108.
Петров К.М. Принципы биономического районирования береговой зоны и шельфа Мирового океана // Океанология. 2020. Т. 60. № 3. С. 381–392.
Дополнительные материалы отсутствуют.
Инструменты
Известия Русского географического общества