

Известия Русского географического общества, 2021, T. 153, № 6, стр. 73-89
Реконструкция палеогеографических условий позднеледниковья-голоцена в Центральной Карелии (на основе комплексного анализа озерных отложений озера Южное Хаугилампи)
Т. С. Шелехова a, *, Н. Б. Лаврова a, **, Д. А. Субетто b, ***
a Институт геологии Карельского научного центра РАН
Петрозаводск, Россия
b Российский государственный педагогический университет им. А.И. Герцена
Санкт-Петербург, Россия
* E-mail: shelekh@krc.karelia.ru
** E-mail: lavrova@krc.karelia.ru
*** E-mail: subetto@mail.ru
Поступила в редакцию 21.05.2021
После доработки 30.05.2021
Принята к публикации 30.06.2021
Аннотация
Результаты диатомового, палинологического, радиоуглеродного анализов позволили установить, что накопление толщи донных отложений озера Южное Хаугилампи происходило на протяжении последних 13–12 тыс. календарных лет. В позднеледниковье в водоеме формировались терригенные осадки (пески, глины), а начиная с пребореала – органогенные (гиттия). Реконструкции динамики растительности охватывают временной интервал от позднего дриаса до настоящего времени. После отступания ледника последнего валдайского оледенения на территории исследования распространилась перигляциальная (Artemisia-Chenopodiaceae) растительность, тундровые и лесотундровые березовые сообщества. В пребореале тундровая и лесотундровая растительность сменилась березовыми редкостойными лесами. Позже, в бореальное время сосновые среднетаежные леса достигли максимального развития. В атлантическое время среднетаежные леса сменяются южнотаежными сосновыми и еловыми лесами с участием термофильных пород. Cреднетаежный облик леса начали приобретать в позднем голоцене. В позднеледниковье в водоеме диатомовая флора практически не развивалась. В осадках этого времени выявлены лишь единичные, возможно переотложенные створки морских диатомей, а также незначительное количество пионерных видов – обрастателей и донных. В пребореальном периоде в озере развивается довольно скудный по численности и разнообразию диатомовый комплекс, представленный пионерными формами, характерными для многих северных водоемов: Ellerbeckia arenaria, Pseudostaurosira brevistriata и др. Массовая вегетация диатомовых водорослей начинается с бореального периода. Прослежена динамика развития озера с позднего дриаса до суббореального времени.
ВВЕДЕНИЕ
Изучение динамики природной среды в позднем неоплейстоцене-голоцене (позднеледниковье-послеледниковье) необходимо для понимания современного ее состояния и прогноза развития экосистем в условиях глобальных климатических изменений. Донные отложения озер представляют собой детальную непрерывную летопись изменений условий палеосреды, дают возможность реконструировать эволюцию озер и природно-климатических изменений в позднем неоплейстоцене−голоцене [17].
Особый интерес к исследованной территории обусловлен тем, что она подвергалась неоднократным оледенениям, которые полностью изменяли существовавшие ландшафты. Расположение территории исследования между краевыми образованиями невской стадии и стадией сальпаусселькя I (ругозерской) позволяет предположить, что она освободилась от льда между 12 500–10 800 14С лет назад [24]. По данным [29, 34], формирование краевых гряд сальпаусселькя I завершилось в начале позднего дриаса, около ~10 700 14C л. н. Следует отметить, что когда ледник отступил в пределы Фенноскандинавского кристаллического щита, то он был слабо обогащен обломками устойчивых к эрозии кристаллических пород и, лишенный супрагляциального комплекса, быстро таял. Согласно полученным ранее данным, озера образовывались практически сразу после таяния ледника [8]. Известно, что время их зарождения было гетерохронным в зависимости от типа котловины, геоморфологического положения и климатических условий. С этих позиций определение возраста малых озер на обширной территории между конечными моренами невской стадии и стадии сальпаусселькя I представляют значительный интерес.
Центральная Карелия в палеогеографическом отношении остается малоизученной. В XX в. опубликованы спорово-пыльцевые диаграммы озерно-болотных отложений болот Мини-Тумба и Чудесное, где формирование озерных отложений в первом из них по палинологическим данным было отнесено к позднему дриасу. Во втором болоте были вскрыты лишь торфяные отложения, начало накопления которых датируется бореалом (8450 14С л. н.) [10]. Данные о диатомовой флоре для территории исследования отсутствуют.
Целью настоящих исследований стало изучение времени зарождения одного из малых озер Центральной Карелии и установление этапов его развития, особенностей осадконакопления и эволюции растительного покрова водосбора со времени дегляциации территории.
РАЙОН ИССЛЕДОВАНИЙ
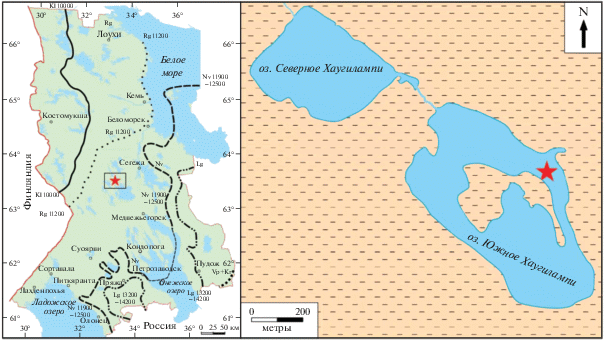
Озеро Южное Хаугилампи расположено в Медвежьегорском районе Республики Карелия в пределах восточной окраины Фенноскандинавского кристаллического щита (63°33.224′ с.ш.; 33°20.262′ в.д.; урез воды – 153 м над уровнем моря (рис. 1).
Рис. 1.
Местоположение оз. Южное Хаугилампи и изученного разреза. Звездочкой обозначено место отбора колонки донных отложений. Границы стадий дегляциации (14С лет назад – по [24]): Vp + Kr (вепсовско-крестецкой); Lg (лужской): 14 200–13 200; Nv (невской): 12 500–11 900; Rg (ругозерской – сальпаусселькя I): 11300–10800; Kl (калевальской – сальпаусселькя II): 10 500–10 200. Fig. 1. Location of the lake Yuzhnoye Haugilampi and the studied section. The asterisk marks the place where the bottom sediment core was taken. Boundaries of deglaciation stages (14С years ago – according [24]): Vp + Kr (Vepsovian-Krestets); Lg (Luga): 14 200–13 200; Nv (Neva): 12 500–11 900; Rg (Rugozerian – Salpausselkä I): 11 300–10 800; Kl (Kalevala – Salpausselkä II): 10 500–10 200.
МАТЕРИАЛЫ И МЕТОДЫ
Весной 2018 г. со льда была пробурена скважина и отобран керн донных осадков, мощностью 350 см, послуживший материалом для данной работы. Глубина воды в месте отбора колонки 250 см. Сверху вниз были вскрыты следующие осадки:
250–350 см – гомогенный черный сапропель с редкими растительными остатками;
350–410 см – гомогенный желеобразный черно-оливковый сапропель с растительными остатками;
410–450 см – черный обводненный слегка разжиженный сапропель;
410–510 см – черный гомогенный сапропель с редкими волосовидными растительными остатками;
510–530 см – черный гомогенный сапропель;
530–538 см – слой коричневато-черного гомогенного сапропеля;
538–564 см – диатомовый? сапропель зеленовато-оливкового цвета (внутри слоя 564–554 см сапропель зеленовато-серого цвета с большим количеством органики);
564–574 см – сапропель черного цвета;
574–578 см – прослой песка с органикой серовато-черного цвета;
578–583 см – песок мелко-тонкозернистый отмытый, с включениями гравия до 1 см;
583–600 см – отмытая гравийная смесь с включениями мелкой гальки до 3–4 см в диаметре.
Образцы на микропалеонтологические анализы отобраны послойно, каждые 5–10 см. Обработка проб для спорово-пыльцевого анализа осуществлялась по общепринятым методикам [16]. При идентификации пыльцевых зерен привлекали справочники-определители [2, 12, 13, 31 и др.], а также эталонную коллекцию пыльцы и спор Института геологии КарНЦ РАН. Спорово-пыльцевая диаграмма построена с использованием компьютерных программ TILIA-2 [25] и TILIA GRAPH [26]. Процентное содержание пыльцы и спор рассчитывались следующим образом. Общая суммы пыльцы древесных и кустарниковых пород (ΣAP), кустарничковой и травянистой растительности (ΣNAP) была принята за 100% (ΣAP + ΣNAP = 100%). Содержание водной (Q) и споровой (F) растительности вычислялось от ΣAP + ΣNAP + ΣQ и ΣAP + ΣNAP + ΣF. На основании изменения состава и соотношений компонентов спорово-пыльцевых спектров (СПС) и применения кластерного анализа (CONISS) выделены палинологические зоны (ПЗ). Полученные результаты сопоставлены с данными радиоуглеродного датирования. Основой периодизации разреза донных отложений послужили схема Блитта–Сернандера, модифицированная Н.А. Хотинским [19] и уточненная для Карелии Г.А. Елиной [10]. Параллельно с палинологическим исследованием в образцах проведено видовое определение водорослей Pediastrum по [27].
Техническая обработка проб и приготовление постоянных препаратов на диатомовый анализ выполнялись по общепринятой методике [7, 9]; при идентификации флоры использовалась различная справочная литература [1, 3–6, 11, 15, 28, 32, 35]. На радиоуглеродное датирование были отобраны шесть образцов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты радиоуглеродного анализа представлены в табл. 1.
Таблица 1.
Радиоуглеродный и калиброванный возраст донных отложений оз. Южное Хаугилампи Table 1. Radiocarbon and calibrated age of bottom sediments of the lake Yuzhnoye Haugilampi
| № п/п | IGAN AMS Лаб. номер | Глубина отбора от поверхности воды, см | Тип отложений для датирования | Радиоуглеродный возраст, лет назад (1σ) | Калиброванный возраст*, лет назад 68,3% (1σ) |
|---|---|---|---|---|---|
| 1 | 6712 | 574–578 | Растительные остатки | 10 660 ± 40 | 12 597–12 681 |
| 2 | 6711 | 570–574 | Растительные остатки | 10 850 ± 35 | 12 706–12 747 |
| 3 | 6710 | 533–537 | Растительные остатки | 10 085 ± 35 | 11 415–11 423 |
| 4 | 6709 | 485–490 | Растительные остатки | 8900 ± 30 | 9937–9997 |
| 5 | 6708 | 380–385 | Растительные остатки | 6710 ± 25 | 7567–7597 |
| 6 | 6707 | 280–285 | Растительные остатки | 5520 ± 20 | 6289–6314 |
Примечание. * RADIOCARBON CALIBRATION PROGRAM CALIB REV7.1.0 Copyright 1986-2018. M. Stuiver and P.J. Reimer. References for calibration datasets: Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Bronk Ramsey C., Buck C.E., Cheng H., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P., Haflidason H., Hajdas I., Hatt G.C., Heaton T.J., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., Manning S.W., Niu M., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Turney C.S.M., van der Plicht J. IntCal13 and MARINE13 radiocarbon age calibration curves 0–50 000 years calBP Radiocarbon 55(4). DOI: 10.2458/azu_js_rc.55.16947
В результате спорово-пыльцевого анализа в донных осадках озера Южное Хаугилампи выделено шесть ПЗ (рис. 2).
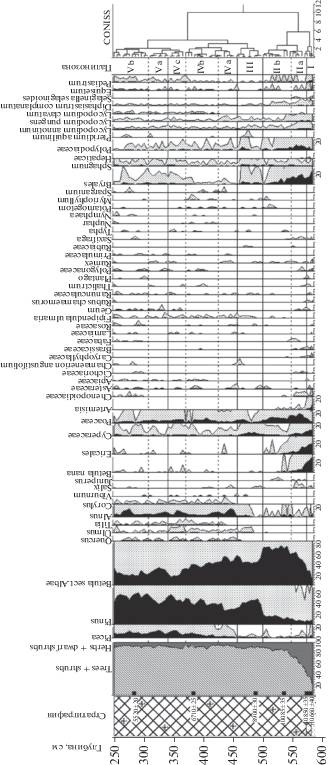
Первый образец, отобранный в крупнозернистых песках (глубина 600–582 см), оказался немым. Вероятно, это обусловлено тем, что пыльца и споры аккумулируются в более тонкой фракции, имеющей такую же скорость седиментации. Скорость осаждения песка несравнимо выше, чем пыльцевых зерен.
ПЗ I (глубина 582–575 см) (песок мелко- и тонкозернистый отмытый, с включениями гравия до 1 см) характеризуется наиболее высоким содержанием пыльцы травянистых растений и кустарничков (60–70%), преимущественно Betula nana L. (27%) и Eri-cales (15–20%), Artemisia (1–10%), вклад которых снижается к верхней границе палинозоны. Количество Cyperaceae, напротив, увеличивается до 10%. Доля пыльцы Be-tula sect. Albae (главным образом, Betula czerepanovii N.I.Orlova) возрастает от 10 до 25%, а доля Pinus колеблется в пределах 5–7%. Вклад пыльцы Picea составляет 3%; у верхней границе зоны эта пыльца отсутствует. Из травянистых следует отметить пыльцу Caryophуllaceae, Brassicaceae, Polygonaceae, Rumex, Saxifraga, Rubus chamaemorus L. В группе споровых преобладают Bryales, Lycopodiaceae, Polypodiaceae. Идентифицированы споры Hepaticae, Selaginella selaginoides (L.) Link. Описанные спорово-пыльцевые спектры отражают наиболее холодный период за все время накопления осадков. Согласно полученным данным, можно предположить, что СПС ПЗ I формировались на заключительном этапе позднего дриаса. Даты 10 660 ± 40 14С л. н./12 597–12 681 кал. л. н. (глубина 574–578 см), 10 850 ± 35 14С л. н. или 12 706–12 747 кал. л. н. (глубина 570–574 см) не соотносятся с данными палинологического анализа и, на наш взгляд, являются несколько завышенными (в сторону бóльшего возраста), что, возможно, связано с переотложением более древней органики.
На окружающей территории наиболее широко были распространены ерниково-зеленомошные и кустарничковые тундровые палеосообщества (Betula nana, Salix, Ericales, Rubus chamaemorus, Selaginella selaginoides, Lycopodium pungens Alderw, Bryales). Роль пионерных полынно-маревых ценозов была уже менее значительной. Возможно, березы (Betula czerepanovii, в меньшей степени Betula pubescens (Ehrh.) создавали островные лесотундровые сообщества, ограниченные благоприятными, защищенными от ветра местообитаниями. Учитывая высокую летучесть пыльцы сосны, можно предположить, что практически вся она была дальнезаносной. В озере были распространены водоросли Pediastrum (P. integrum var. integrum, P. kawraiskyi, P. privum), характерные для холодных олиготрофных водоемов. Самое высокое количество спор плаунов за всю историю накопления отложений, присутствие Нepaticae и низкая насыщенность осадков пыльцой указывает на то, что растительный покров был несомкнутым. Это послужило причиной денудации поверхностных отложений на водосборе и поступления в водоем обломочного минерального материала.
ПЗ II (глубина 575–500 см) отличается увеличением количества пыльцы древесной растительности, главным образом, Betula sect. Albae (Betula pubescens), которая достигает своего максимума у верхней границы палинозоны. Между тем, некоторые особенности СПС позволяют выделить в рамках палинозоны II две подзоны. Подзона ПЗ II а (575–547 см) характеризуется ростом количества пыльцы Betula sect. Albae и снижением количества пыльцы Betula nana, Ericales, Cyperaceae, спор Bryales. Зафиксирована пыльца Salix, Alnus. В подзоне ПЗ II b (547–501 см) отмечается стабильно высокое содержание пыльцы Betula sect. Albae (65–70%), увеличение содержания пыльцы Pinus, дальнейшее снижение вклада пыльцы и спор вышеперечисленных видов и групп (Betula nana, Ericales, Bryales). В обеих подзонах присутствует пыльца Juniperus. В спектрах подзоны ПЗ II b отмечены пыльцевые зерна Typha sp., Potamogeton, Sparganium, Myriophyllum alterniflorum DC, а также колонии водорослей Pediastrum. Следует отметить, что начиная с глубины 450 см и в вышележащих отложениях встречаются устьица сосны, что указывает на то, что эта порода произрастала на изучаемой территории. Полученные палинологические данные и сравнение их с региональными спектрами свидетельствуют о том, что СПС палинозоны II формировались в пребореальное время. Это подтверждается и радиоуглеродной датой 10 085 ± 35 14С л. н./11 415–11 423 кал. л. н. (глубина 533–537 см) (см. табл. 1).
Увеличение содержания пыльцы древесных растений было обусловлено значительным улучшением климатических условий. Это благоприятствовало распространению, в первую очередь, березы, как пионерной породы. На изучаемой территории доминировали березовые лесотундровые сообщества, которые позже сменились березовыми и сосново-березовыми редкостойными лесами северотаежного облика. О редкостойности древесных палеосообществ свидетельствует присутствие в СПС пыльцы разнообразных плаунов и можжевельника. В то время мелководья водоема начинают зарастать водно-болотными видами. Среди водорослей Pediastrum появились более теплолюбивые виды Pediastrum boryanum var. boryanum и P. duplex var. duplex. Распространение водорослей, прибрежно-водных растений и закрепление субстратов растительностью, препятствующей сносу минерального материала в палеоводоем, а также активное почвообразование привели к снижению интенсивности денудационных процессов на водосборе, уменьшению твердого стока в озеро и, как следствие, к смене условий накопления отложений от минеральных осадков к органогенным (илы и гиттии).
Нижняя граница ПЗ Ш (глубина 500–458 см) маркируется значительным увеличением количества пыльцы Pinus (50%) и снижением содержания пыльцы Betula sect. A-lbae (42–45%). Появляется в СПС пыльца Ulmus, увеличивается процентное содержание зерен Alnus (в т. ч. Alnus glutinosa). Среди спор преобладают Bryales (2–4%), Polypodiaceae (1–3%), Lycopodiaceae (~2%). Идентифицированы пыльца Juniperus, споры Pteridium aquilinum. В группе травянистых растений доминирует пыльца Cyperaceae, увеличивается частота встречаемости пыльцы Asteraceae, Filipendula ulmaria, Ranunculaceae. Помимо пыльцевых зерен Typha, Potamogeton и Sparganium появляется пыльца Nymphaea. Верхняя граница палинозоны проведена выше максимума содержания пыльцы Pinus. Согласно данным спорово-пыльцевого анализа палиноспектры рассматриваемой ПЗ формировались в бореальный период, что согласуется с датировкой 8900 ± 30 14С л. н./9937–9997 кал. л. н. (глубина 485–490 см). Для этого периода характерно потепление климата на фоне дефицита влажности. Уменьшение влажности наряду с гляциоизостатическим поднятием Фенноскандинавского щита вызвало понижение уровня грунтовых вод [22], что способствовало широкому распространению сосны. В напочвенном покрове сосновых лесов появляется орляк обыкновенный – представитель борового ценогенетического комплекса. Наряду с сосновыми лесами продолжали существовать березово-сосновые и, в наиболее влажных местообитаниях, крупнотравные березовые сообщества. Леса постепенно приобретали среднетаежный облик, о чем косвенно свидетельствует уменьшение роли плаунов. Во время максимального для бореального периода потепления [18] граница ареала распространения вяза (Ulmus) приблизились к исследуемой территории, что нашло отражение и в диаграмме. В озерной экосистеме усиливаются продукционные процессы, что способствовало активному накоплению автохтонного органического вещества в донных отложениях и постепенному заилению котловины озера. Расширялась зона хорошо прогреваемых мелководий палеоводоема, которые продолжали зарастать водными и водно-болотными видами.
ПЗ IV (458–340 см) характеризуется невысоким, но стабильным содержанием пыльцы термофильных и широколиственных пород (Quercus, Tilia, Ulmus, Corylus, Al-nus glutinosa (L.) Gaertn). Данные спорово-пыльцевого анализа, а также радиоуглеродная дата 6710 ± 25 14С л. н./7567–7597 кал. л. н. (глубина 380–385 см) позволяют отнести время формирования этой палинозоны к атлантическому периоду. В СПС наблюдается тенденция увеличения вклада пыльцы Picea и Pinus и уменьшения содержания пыльцы Betula sect. Albae. Изменения в соотношениях этих основных компонентов СПС позволяют выделить несколько подзон. Чередование пиков пыльцы сосны и березы, возможно, обусловлено следующими причинами: 1) неоднократные изменения влажности климата в атлантическое время; 2) пожары, после которых на гарях формировались постпирогенные березняки.
Подзона ПЗ IV a (глубина 458–425 см) характеризуется уменьшением количества пыльцы Pinus, тогда как вклад пыльцы Betula sect. Albae, Picea, Alnus увеличивается. В палиноспектрах, помимо пыльцы Ulmus, появляется Quercus, Corylus, Alnus glutinosa, у верхней границы палинозоны – Tilia. Скорее всего, это связано с изменением климата в сторону потепления и увеличения влажности, что характерно для атлантического периода. Следует отметить, что перенос пыльцы липы и дуба по воздуху происходит в пределах лишь нескольких десятков километров [30]. Вероятно, термофильные породы уже произрастали на исследуемой территории.
В спорово-пыльцевых спектрах подзоны ПЗ IV b (глубина 425–368 см) отмечается постепенное увеличение содержания пыльцы Pinus за счет пыльцы Betula sect. Albae. Доля пыльцы Picea в спектрах возрастает до 7–9%, что свидетельствует о существенном участии ели в растительном покрове. Вклад пыльцы термофильных пород незначителен, однако их кривые на диаграмме непрерывны. Cреди травянистых лидирует пыльца Poaceae. В СПС палинозоны возрастает вклад пыльцы водных и прибрежно-водных растений. Согласно полученным данным, сосна вновь возвращается на прежние позиции, вытесняя березовые сообщества. Уровень палеоводоема понижается. Мелководья его хорошо прогреваются, зарастают водными и прибрежно-водными растениями, о чем свидетельствует повышение их доли в СПС. На территории исследования были распространены южнотаежные елово-сосновые, сосново-еловые, сосново-березовые леса, в составе которых принимали участие термофильные древесные породы. Наиболее влажные местообитания занимали еловые сообщества с ольхой черной, вязом и лещиной.
В СПС подзоны ПЗ IV c (глубина 368–340 см) зафиксировано наиболее высокое участие пыльцы термофильных пород. Вероятно, импульсом для их распространения послужил максимум атлантического потепления [18]. Кроме этого, отмечаются пик содержания пыльцы Pinus (60%), увеличение вклада пыльцы Piceа (до 15%) и падение доли пыльцы Betula sect. Albae до 18%.
Потепление и увеличение влажности климата создало условия для расселения термофильных пород. Широкое распространение имели елово-сосновые, сосново-еловые и сосново-березовые леса с ольхой и теплолюбивыми древесными породами и кустарниками (вяз, липа, дуб и лещина). Крупнотравные березовые с ольхой серой и елово-черноольховые леса с примесью вяза и лещины произрастали в депрессиях рельефа с проточным увлажнением и богатыми почвами.
ПЗ V (глубина 340–305 см) У нижней границы палинозоны зафиксировано снижение количества пыльцы термофильных пород, доля которых впоследствии вновь возрастает, но не достигает значений, зарегистрированных в предыдущей палинозоне. Среди основных компонентов спектров отмечается следующая тенденция – увеличение количества пыльцы Betula sect. Albae до 35%, Picea (15%) и уменьшение доли пыльцы Pinus до 43%. Известно, что количество пыльцы сосны и березы в спектрах выше по отношению к участию этих пород в составе древостоя, а ели и широколиственных пород, напротив, ниже [10, 14 и др.].
В следующей ПЗ VI (глубина 305–250 cм) пыльца Pinus вновь преобладает (58%), на втором месте – пыльца Betula sect. Albae (20–28%), на третьем – пыльца Picea (5–10%). Надо отметить довольно существенный вклад пыльцы Alnus (5–16%).
Принимая во внимание снижение вклада пыльцы термофильных пород и увеличение в палиноспектрах количества пыльцевых зерен Picea, мы предполагаем, что палиноспектры двух последних палинозон были сформированы в позднем голоцене. На исследуемой территории на увлажненных местообитаниях, наряду с елово-сосновыми лесами, произрастали монодоминантные ельники, а на сухих песчаных и каменистых субстратах – сосняки. Участие широколиственных пород в лесах сократилось.
Методом диатомового анализа было исследовано 30 образцов в интервале глубин 600–275 см. В донных отложениях выявлено 122 вида и разновидности диатомовой флоры из 37 родов. В результате исследования по составу полученных комплексов выделено пять диатомовых зон (DZ), соответствующих различным этапам развития водоема и палеоклиматическим периодам позднеледниковья и голоцена. Результаты исследований представлены на диаграмме (рис. 3).
Рис. 3.
Диатомовая диаграмма донных отложений оз. Южное Хаугилампи. Fig. 3. Diatom diagram of bottom sediments from the lake Yuzhnoye Haugilampi.
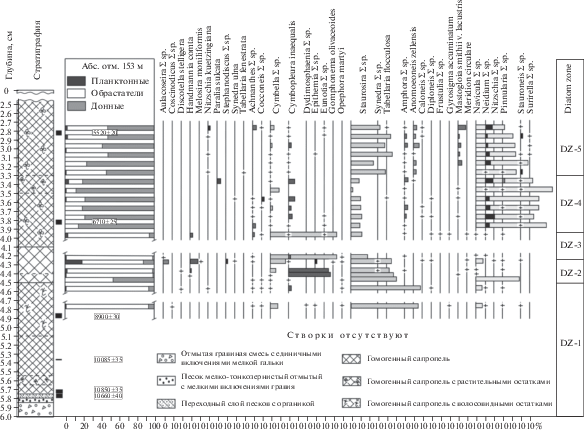
В отложениях на глубине 600–470 cм ископаемая диатомовая флора практически отсутствует. С глубины 555–550 см выявлены Nitzschia sp.; на глубине 540–538 см – единичная морская створка Pinnularia quadratarea. В образце с глубины 525–520 см определено всего 43 створки, представленные видами Staurosira venter, St. pinnata, Fragilaria construens, Achnanthes linearis, Ach. minutissima, Discotella stelligera, Cyclotella kuetzingiana, Navicula radiosa и др. Большинство из них корродированы. С глубины 490–485 см в осадках также выявлены единичные сильно корродированные створки видов Ellerbeckia arenaria, Pseudostaurosira brevistriata, Grammatophora oceanica, Coscinodiscus sp., Mastogloia sp., Navicula pupula var. pupula и десмидиевая водоросль Ephidatia muellery. Все обнаруженные виды, кроме мелкого обломка Coscinodiscus sp. и Grammatophora oceanica – пресноводные. Можно предположить, что отсутствие диатомовой флоры в нижних слоях разреза связано с высокими значениями рН (более 8.5), при которых диатомеи почти не могли развиваться. Однако более достоверным, с нашей точки зрения, является предположение, что из-за холодных условий позднеледниковья в водоем поступало недостаточно кислорода и растворенной кремнекислоты, необходимой для активной вегетации диатомовых водорослей.
Наиболее полноценный диатомовый комплекс установлен в осадках, представленных гомогенными сапропелями черного цвета с редкими волосовидными растительными остатками с глубины 480 см.
DZ 1 (490–450 см) выделена на основе доминирования видов обрастаний рода Staurosira, достигающих почти 80% от общего состава флоры; остальные 20% представлены формами рода Cymbella sp. (причем большинство из них корродированно и разъедено, что вызывало трудности при идентификации). Из определенных форм следует отметить пресноводные Cymbella laevis, C. ventricosa; возможно в обломках представлены и морские формы этого рода. Среди донных видов рода Navicula определены мезогалобы Navicula peregrina, N. diluviana, а также галофил Gyrosigma acuminatum, несколько обломков крупной створки Dydimosphaenia sp. Важно отметить присутствие в виде обломков солоноватоводно-морских форм Coscinodiscus sp., Nitzschia sp., Epithemia sp. (глубина 480–475 см). Выше по разрезу (глубина 460–455 см) встречаются корродированные створки морских форм рода Pinnularia sp., а также целые створки Diploneis smithi, Cymbopleura inaequalis, Cocconeis scutellum. Эти единичные створки морских и солоноватоводных видов следует отнести к переотложенным, при доминировании пресноводных рода Staurosira. Отложения формировались в бореальное время.
Анализируя эколого-географические характеристики диатомового комплекса и рН-спектр, можно отметить, что на данном этапе господствовали космополитные формы, составляя более 70% флоры (рис. 4).
Рис. 4.
Эколого-географические характеристики и рН-спектр диатомовой флоры донных осадков оз. Южное Хаугилампи. Fig. 4. Ecological and geographical characteristics and pH – spectrum of diatom flora of bottom sediments of the lake Yuzhnoye Haugilampi.
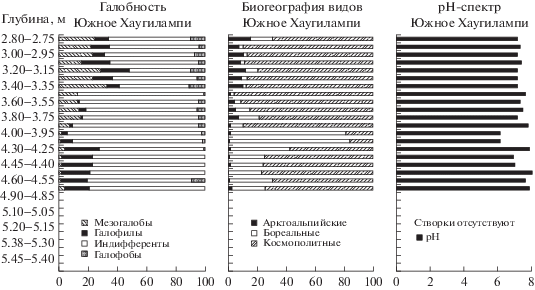
В структуре видов по галобности преобладали индифференты, однако доля галофилов составляла 20% при незначительном количестве галофобов. При этом среда была щелочной, показатель рН достигал максимальных значений 8. Таким образом, состав диатомового комплекса DZ 1 свидетельствует о мелководности хорошо прогреваемого водоема со щелочной средой, что характерно для бореального времени.
DZ 2 (450–425 см). В структуре диатомового комплекса отмечаются значительные колебания процентного соотношения отдельных групп водорослей. В начале зоны возрастает доля донных форм рода Navicula sp. (до 50%), примерно такого же количества достигают и формы обрастаний Staurosira sp. К концу этапа участие Staurosira sp. снижается с 80 до 50%. Все остальные виды присутствуют единично. Однако на глубине 445–435 см донные диатомеи рода Navicula sp. вытесняются формами обрастаний Cymbopleura inaequalis (почти 50%). В конце DZ 2 появляются планктонные виды Handmania comta, а также Aulacoseira granulata + angustissima, Stephanodiscus hantzschii, St. astraea (до 20%). Содержание форм обрастаний Staurosira sp. постоянно и составляет также около 20%. Возрастает роль видов рода Cymbella sp., представленных главным образом Cymbella obtusa, а также Cymbopleura inaequalis. Вновь появляются донные виды рода Navicula sp. (10%), единичные створки галофилов Epithemia zebra, E. zebra var. saxonica, E. muellery, Melosira moniliformis. Несмотря на относительно высокое содержание галофилов (до 30%) при доминировании индифферентных форм, рН резко снижается до нейтральных значений (рис. 4). К концу этапа возрастает роль бореальных видов, что связано, скорее всего, с нарастанием увлажненности климата в начале атлантического времени.
DZ 3 (425–395 см) отличается полным господством видов рода Cymbella sp. (около 80%), среди которых выделяются Cymbella laevis, C. obtusa, снижением содержания видов рода Staurosira sp. (до 10–15%). Остальные представители диатомового комплекса Handmania comta, Navicula sp., Gyrosigma sp., Achnanthes sp., Diploneis sp. и другие выявлены единично. Отличительной особенностью диатомового комплекса данной DZ выступает резкое повышение содержания бореальных форм и сокращение роли космополитов (до 20%). При этом, в результате смены гидрохимических условий со щелочных на кислые (значения рН снизились с 8 до 6.1) заметно уменьшилось количество галофилов. Возросло участие планктонного сообщества (с 2 до 20%), предположительно, в результате обводнения водоема, возможно за счет увлажнения климата во второй половине атлантического периода.
DZ 4 (395–335 см) отличается господством донных форм диатомовых водорослей, представленных видами родов Navicula sp. (70–90%), Stauroneis sp. (10%), Neidium sp. (5–10%), незначительным количеством обрастателей Cymbella sp., Cymbopleura in-aequalis (5–10%) и единичным участием Achnanthes sp., Cocconeis sp., Stauroneis sp., Eunotia sp. В структуре комплекса по галобности возрастает содержание мезогалобов (15% и более) и, наоборот, снижается доля галофильных форм по сравнению с предыдущим этапом. Вместе с тем заметно повышаются значения рН (колеблются в пределах 7–7.5), указывая на слабощелочную среду. В конце этапа в небольших количествах появляются, вероятно, переотложенные створки морского планктонно-литорального вида Paralia sulcata (до 5%), что может быть связано с потеплением климата в конце атлантического периода, усилением процессов питания водоема подземными более минерализованными водами, возможным поступлением с водораздела ранее отложенных четвертичных осадков.
DZ 5 (335–275 см). На данном этапе развития водоема в структуре диатомового комплекса происходят заметные изменения, связанные со значительным увеличением роли обрастателей Staurosira sp. (с 2–5 до 50%), снижением содержания донных форм с 90 до 40%. Кроме этого в небольшом количестве выявлены новые виды: мезогалоб Mastogloia smithi var. lacustris (10%), слабый кальцефил Anomoeoneis zellensis, а также виды родов Amphora sp., Neidium sp. (до 10%). Формы из обрастаний Cymbella sp., Achnanthes sp., Nitzschia kuetzingiana и др. составляют не более 2–5%. Увеличение роли видов рода Staurosira sp. может свидетельствовать о лучшей прогреваемости водоема, а также, вероятно, появлении новых мелководий из-за снижения уровня воды в результате сухости климата в суббореальное время. На это указывает сокращение до 20% доли арктоальпийских и бореальных форм, расширение спектра космополитов. В структуре комплекса по галобности повышается содержание мезогалобов (с 10 до 20%). Вместе с галофилами в середине этапа они достигают 50%. Показатель рН практически не меняется и близок к нейтральному. Все это может указывать на относительно теплые климатические условия суббореального времени.
Таким образом, отличительной особенностью суббореальных отложений изученного водоема является высокое содержание в составе диатомового комплекса мезогалобов и галофилов (суммарно 50%). Такого большого содержания видов этих групп не выявлено для этого времени ни в одном водоеме на территории Карелии. Большое содержание данных видов в составе диатомового комплекса можно объяснить: а) переотложением из ранее не размытых морских осадков (микулинских или моря Портландия); б) развитием видов в этом периоде. Ранее установленный уровень трансгрессии Беломорского бассейна (моря Портландия) достигал современных абсолютных отметок 150–175 м [21, 24]. Подтверждением переотложения створок служит сходство составов диатомовых комплексов моря Портландия и даже микулинских (рис. 5).
Рис. 5.
Переотложенные створки диатомовых водорослей: 1 – Paralia sulcata; 2 – Hyalodiscus scoticus; 3 – Mastogloia elliptica; 4 – Campilodiscus sp.; 5 – Grammatophora oceanica; 6 – Navicula peregrina. Fig. 5. Redeposited valves of diatoms.
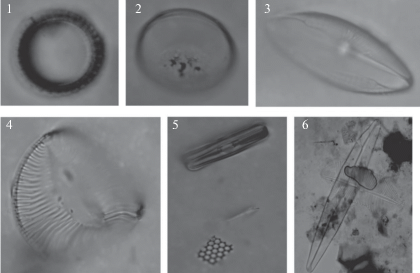
Среди типичных видов, отмеченных в суббореале в озере Южное Хаугилампи, необходимо выделить мезогалобы Navicula halophila, N. peregrina, Navicula sp., Cocconeis scutellum; галофилы Staurosirella pinnata, Navicula oblonga, Navicula (Sellaphora) pupula var. rectangularis, Diploneis sp. Кроме этого важно заметить, что перечисленные виды отличаются более плохой сохранностью по сравнению с пресноводными индифферентами и галофобами. Присутствие видов Paralia sulcata, Pinnularia quadratarea, Cocconeis costata, Cocconeis scutellum – типичных представителей литоральной зоны Белого моря − еще больше подтверждает наше предположение. А их совместное существование с галофобами хорошей сохранности (Neidium amphilatum, Neidium bisulcatum), вообще не совместимых по экологии с морскими формами, является явным свидетельством переотложения. На рис. 5 представлены фотографии со светового микроскопа, свидетельствующие о плохой сохранности переотложенной флоры и наличии морских форм, несовместимых по обитанию с пресноводными.
Осадки субатлантического времени не зафиксированы в разрезе. Вероятно, с помощью ручного бура их поднять не удалось из-за разжиженной консистенции.
ВЫВОДЫ
Согласно полученным данным, установлено, что осадконакопление в водоеме началось в позднем дриасе. Базальные отложения, представленные гравийной смесью с включением мелкой гальки, лишены микрофоссилий и их возраст не может быть установлен биостратиграфическим методом. В результате спорово-пыльцевого анализа выделены отложения заключительного этапа позднего дриаса, пребореального, бореального, атлантического периода и нерасчлененные осадки позднего голоцена.
Тундровые ландшафты позднего дриаса постепенно замещались березовыми редколесьями. Направленное потепление и увлажнение климата началось в пребореальном периоде, что нашло отражение в составе СПС. Предположительно во второй половине пребореала береза становится доминирующей породой в северотаежных лесах изучаемой территории. В их составе все большее участие начинает принимать сосна, образуя сосново-березовые палеоценозы. В бореальное время максимального развития достигли сосновые среднетаежные леса, как монодоминантные, так и с участием березы. В атлантическом периоде они сменяются южнотаежными сосновыми и еловыми лесами с участием термофильных пород. В позднем голоцене леса начали приобретать среднетаежный облик.
По данным диатомового анализа выделяются позднеледниковые отложения, сменяющиеся осадками от пребореального до суббореального периодов. Позднеледниковые отложения практически не содержат ископаемой диатомовой флоры. В них выявлены лишь единичные морские створки Pinnularia quadratarea, Nitzschia sp. В пребореальном периоде в водоеме в незначительном количестве развивались пионерные виды из обрастаний Staurosira venter, St. pinnata, Fragilaria construens, Achnanthes linearis, Ach. minutissima, а также планктонные виды Discotella stelligera, Cyclotella kuetzingiana, донные виды Navicula radiosa и другие. Массовая вегетация диатомовых водорослей началась лишь в бореальное время во время максимального для этого периода потепления. В донных отложениях начала бореала при доминировании эпифитов Staurosira sp. выявлены также единичные сильно корродированные створки видов Ellerbeckia arenaria, Pseudostaurosira brevistriata, морская, вероятно, переотложенная форма Grammatophora oceanica, мелкий обломок Coscinodiscus sp., мезогалобные донные формы Mastogloia sp., галофилы Navicula (Sellaphora) pupula var. pupula и десмидиевая водоросль Ephidatia muellery.
В атлантическом периоде наблюдается подъем уровня, фиксирующийся по повышению содержания планктонных форм и диатомей из обрастаний.
В суббореальный период максимального содержания (50%) в диатомовом комплексе достигли мезогалобы в сумме с галофилами. Такое высокое их содержание в суббореале Карелии зафиксировано впервые и не наблюдалось ранее ни в одном из изученных водоемов Карелии. Однако, в это время отмечается рост количества данных видов во всех водоемах, что объясняется сухостью климата и усилением подземного питания, повышением минерализации воды [20]. Но в изученном озере такое высокое для Карелии содержание мезогалобов и галофилов, по нашему мнению, не связано с названными причинами, а выступает следствием переотложения и выщелачивания ранее сформированных и не до конца выветрелых морских микулинских (?) осадков, или осадков моря Портландия.
Результаты диатомового анализа донных отложений, начиная с бореального времени, подтверждаются данными спорово-пыльцевого анализа и радиоуглеродного датирования, таким образом, доказывая надежность комплексных исследований.
В соответствии с предложенной ранее схемой [23] образование малых озер, расположенных между конечными моренами сямозерской (невской) и ругозерской стадиями оледенения, происходило в аллереде. Согласно полученным данным, начало формирования донных отложений в озере Южное Хаугилампи, находящемся почти на одинаковом (~60 км) расстоянии от конечных морен невской и ругозерской стадий, относится к позднему дриасу (не ранее 11 000 14С л. н./12 900 кал. л. н.), что свидетельствует о запаздывании образования озера относительно края отступающего ледника. Дополнительные разносторонние комплексные исследования донных отложений малых озер этого региона позволят получить более достоверную дополнительную информацию о времени зарождения озер и об особенностях ископаемой диатомовой флоры для каждого палеоклиматического периода голоцена.
Работа выполнена в рамках государственной темы ИГ Кар НЦ РАН № АААА-А18-118020690231-1 и гранта РНФ 18-17-00176.
Список литературы
Баринова С.С., Медведева Л.А., Анисимова О.В. Биоразнообразие водорослей – индикаторов окружающей среды. Тель-Авив: PilliesStudio, 2006. 498 с.
Бобров А.Е., Куприянова Л.A., Литвинцева М.В., Тарасевич В.Ф. Споры папоротникообразных и пыльца голосеменных и однодольных растений флоры европейской части СССР. Л.: Наука: Ленингр. отд-ние, 1983. 208 с.
Генкал С.И., Трифонова И.С. Диатомовые водоросли планктона Ладожского озера и водоемов его бассейна. Рыбинск: Изд-во “Рыбинский дом печати”, 2009. 72 с.
Генкал С.И., Чекрыжева Т.А. Центрические диатомовые водоросли (Bacillariophyta, Centrophyceae) водоемов Карелии // Биол. внутр. вод. 2011. № 1. С. 5–16.
Генкал С.И., Куликовский М.С., Михеева Т.М., Кузнецова И.В., Лукьянова Е.В. Диатомовые водоросли планктона реки Свислочь и ее водохранилищ. М.: Научный мир, 2013. 236 с.
Генкал С.И., Чекрыжева Т. А., Комулайнен С.Ф. Диатомовые водоросли водоемов и водотоков Карелии. М.: Научный мир, 2015. 202 с.
Давыдова Н.Н. Диатомовые водоросли – индикаторы природных условий водоемов в голоцене. Л.: Наука, 1985. 244 с.
Демидов И.Н. Деградация поздневалдайского оледенения в бассейне Онежского озера // Геология и полезные ископаемые Карелии. Вып. 8. Петрозаводск: Карел. науч. центр РАН, 2005. С. 134–142.
Диатомовый анализ. Определитель ископаемых и современных диатомовых водорослей. Л.: Государственное издательство геологической литературы, 1949–1950. Кн. 1. 240 с. Кн. 2. 288 с. Кн. 3. 399 с.
Елина Г.А. Принципы и методы реконструкции и картирования растительности голоцена. Л.: Наука, 1981. 160 с.
Куликовский М.С., Глушенко А.М., Генкал С.И. Кузнецова И.В. Определитель диатомовых водорослей России. Ярославль: Филигрань, 2016. 804 с.
Куприянова Л.А., Алешина Л.А. Пыльца и споры растений флоры европейской части СССР. Т. 1. Л.: Наука, 1972. 171 с.
Куприянова Л.А., Алешина Л.А. Пыльца двудольных растений флоры европейской части СССР. Lamiaceae, Zygophyllaceae. Л.: Наука, 1978. 184 с.
Лийвранд Э. Методические проблемы палиностратиграфии плейстоцена. Таллинн, 1990. 176 с.
Определитель пресноводных водорослей СССР. М.; Л.: Изд-во АН СССР, 1951. Вып. 4. 619 с.
Пыльцевой анализ / Под ред. И.М. Покровской. М.: Гос. изд-во геологической литературы, 1950. 571 с.
Субетто Д.А. Донные отложения озер: палеолимнологические реконструкции. СПб.: Из-во РГПУ им. Герцена, 2009. 339 с.
Филимонова Л.В., Климанов В.А. Изменение количественных показателей палеоклимата в среднетаежной подзоне Карелии за последние 11000 лет // Труды Карельского научного центра РАН. 2005. Вып. 8. С. 112–120.
Хотинский Н.А. Голоцен Северной Евразии. М.: Наука, 1977. 200 с.
Шелехова Т.С. Эволюция малых озер охраняемых территорий Карелии (по данным диатомового анализа донных отложений): Автореф. дис. канд. геогр. наук. СПб., 1998. 26 с.
Шелехова Т.С. Четвертичные отложения и геоморфология Зеленого Пояса Фенноскандии // Труды Карельского научного центра РАН. № 4. 2019. С. 41–50. https://doi.org/10.17076/them1007
Шелехова Т.С., Лаврова Н.Б. Палеогеографические реконструкции развития территории Северо-западной Карелии в голоцене (по данным изучения донных отложений малых озер) // Труды Карельского научного центра РАН. 2020. № 9. С. 101–122. https://doi.org/10.17076/lim1268
Экман И.М., Лукашов А.Д., Колканен А.М., Лийва А.А. Динамика развития озер и озерности Карельской АССР по данным радиохронологии (за последние 13000–14000 лет) // Изотопно-геохронологические исследования в Прибалтике и Белоруссии. Таллинн, 1988. С. 237.
Ekman I., Iliyn V. (1995). Deglatiation, the Younger Dryas end moraines and their correlation in Russian Karelia and adjacent areas // Glacial deposits in North-East Europe / J. Ehlers, S. Kozarski, P. Gibbard (Eds.). Rotterdam-Brookfield: A.A. Balkema. P. 195–209.
Grimm E.C. Tilia and TiliaGraph PC spreadsheet and graphics software for pollen data. INQUA, working group on data-handlingmethods. Newsletter 4. 1999. P. 5–7.
Grimm E.C. TGView. Illinois State Museum, Research and Collections Center, Springfield, 2004.
Komarek J., Jankovska V. Review of the Green Algal Genus Pediastrum: Implication for Pollen analytical Research. Bibliotheca Phycologica. Band XX. Berlin, Stuttgart: Cramer, 1999. 127 p.
Krammer K., Lange-Bertalot H. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae // Sűswasserflora von Mitteleuropa. Stuttgart; Jena: Gustav Fischer Verlag, 1991. 576 p.
Lunkka J.P., Johansson P., Saarnisto M., Sallasmaa O. Glaciation of Finland // Quaternary Glaciation – Extent and Chronology / J. Ehlers, P.L. Gibbard (Eds.). Elsevier Science, 2004. P. 93–100.
Mazei N.G., Kusilman M.V., Novenko E.Y. The Occurrence of Carpinus, Fagus, Tilia, and Quercus Pollen in Subrecent Spore-Pollen Spectra from the East European Plain: On the Possibility of Long-Distance Pollen Transfer // Russian J. Ecology. 2018. V. 49 (6). P. 484–491. https://doi.org/10.1134/S1067413618050077
Moore P.D., Webb J.A., Collinson M.E. Pollen analysis. Second edition. London, Malden, Carlton: Blackwell Science, 1991. 216 p.
Mölder K., Tynni R. Uber Finnlands rezente und subfossile Diatomeen. I–VII // Bull. Geol. Soc. Finland. 1967. V. 39. P. 199–217; 1968. V. 40. P. 151–170; 1969. V. 41. P. 235–251; 1970. V. 42. P. 129–144; 1971. V. 43. P. 203–220; 1972. V. 44. P. 141–149; 1973. V. 45. P. 159–179.
Niemelӓ J., Ekman I., Lukashov A. (Eds.). Quaternary deposits of Finland and Northwestern part of Russian Federation and their resources. Scale 1 : 1000000. Espoo: Geological Survey of Finland, 1993.
Saarnisto M., Saarinen T. Deglaciation chronology of the Scandinavian Ice Sheet from the Lake Onega Basin to the Salpausselkä End Moraines // Global and Planetary Change. 2001. V. 31(1–4). P. 387–405. https://doi.org/10.1016/S0921-8181(01)00131-X
Tynni R. Uber Finnlands rezente und subfossile Diatomeen VIII–XI // Geol. Surv. Finland Bull. 1975. V. 274. P. 1–55; 1976. V. 284. P. 1–37; 1978. V. 296. P. 1–55; 1980. V. 312. P. 1–93.
Дополнительные материалы отсутствуют.
Инструменты
Известия Русского географического общества