

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 1, стр. 71-85
ИССЛЕДОВАНИЕ ОБУЧЕНИЯ НАВЫКАМ РАЗНОЙ СТЕПЕНИ СЛОЖНОСТИ У ПАВИАНОВ ГАМАДРИЛОВ (PAPIO HAMADRYAS)
А. Е. Аникаев 1, *, В. Г. Чалян 1, Н. В. Мейшвили 1
1 ФГБНУ “Научно-исследовательский институт медицинской приматологии”
Сочи, Россия
* E-mail: mg_anykey@mail.ru
Поступила в редакцию 18.04.2019
После доработки 30.08.2019
Принята к публикации 16.09.2019
Аннотация
Проведено исследование способности к обучению навыкам трех уровней сложности у 69 особей павианов гамадрилов (Papio hamadryas). Формировались навыки, в которых в качестве стимулов использовались: положение и цвет (навык 1), только положение (навык 2) и только цвет (навык 3). Оценка результатов осуществлялась по следующим критериям: обучаемость, обученность, экспериментальная активность и динамика обучения. Зафиксирован высокий уровень экспериментальной активности по трем навыкам. Отмечены высокие значения показателей обучаемости и обученности и по первому навыку, и второму навыку. Установлено, что исследуемые животные в большинстве своем не способны обучиться третьему навыку. Результаты обсуждаются.
ВВЕДЕНИЕ
Исследование когнитивных способностей приматов является одним из приоритетных направлений современной науки. Данные, полученные в этой области, являются источником информации для решения проблем эволюции психики [Barney et al., 2015; Beran et al., 2016; Byrne, Bates, 2010; Seed, Tomasello, 2010; van Schaik, Pradhau, 2003]. Исследования когнитивных способностей лабораторных приматов служат выявлению особенностей функционирования мозга [Boschin et al., 2015; Byrne, Corp, 2004; Deaner et al., 2007; Gale et al., 2013; Paton et al., 2006; Wilson et al., 2017], установлению эффектов фармацевтических препаратов [Curtis et al., 2015; Sandin et al., 1990; Zhou et al., 2015], оценке характера влияния на когнитивные функции различных воздействий [Беляева и др., 2017; Krause et al., 2017; Rahman et al., 2013; Schneider et al., 2013], старения [Callahan et al., 2013; Joly et al., 2014; Nagahara et al., 2010]. Проведение подобных исследований с использованием лабораторных приматов должно опираться на знание когнитивных показателей животных, находящихся в нормальном физиологическом состоянии. Данное исследование в первую очередь направлено на анализ такой характеристики, как способность к обучению у павианов гамадрилов (Papio hamadryas).
Со времен применения методики “установки на обучение” (objet learning-set formation) Харлоу [Harlow, 1949] по данной тематике накоплен определенный багаж данных. Исследования способности к обучению у приматов проводятся с помощью разных методических приемов, начиная от классических установок до современных систем, предполагающих работу животных с экраном компьютера как в индивидуальных условиях содержания, так и в условиях содержания социальной группой [Fagot, Bonte, 2010; Fagot et al., 2013; Zurcher et al., 2010]. Объектом подобных исследований являются различные виды приматов, от лемуров, капуцинов и макаков до антропоидов. Наиболее хорошо исследованы способности к обучению шимпанзе, горилл и орангутанов, что позволило установить их превосходство в этих способностях по сравнению с другими видами обезьян [Natale et al., 1986; Rumbaugh, Pate, 2014]. В последнее время особое внимание в качестве объекта исследований способностей к обучению привлекают разные виды павианов: гвинейские павианы (Papio papio) [Fagot, Cook, 2006; Fagot, Lillo, 2011; Flemming et al., 2013; Goujon, Fagot, 2013; Hannagan et al., 2014; Huguet et al., 2014; Poletiek et al., 2016], анубисы (Papio anubis) [Schmidt, Fisher, 2009; Schmidt et al., 2012; Vick et al., 2001, 2003] и чакма (Papio ursinus) [Bergman, Kitchen, 2009; Noser, Byrne, 2015]. В частности, у гвинейских павианов (Papio papio) обнаружена способность научиться различать 4-буквенные слова на экране компьютера [Linke et al., 2017]. Число слов, которые выучили 6 самцов павианов после 60 000 проб, варьировало от 81 до 307. В экспериментах по исследованию способностей к научению павианами запоминания пространственного положения обнаружено высокое сходство механизмов этого процесса с таковым у человека [Goujon, Fagot, 2013]. В ряде сравнительных исследований установлено, что павианы демонстрируют относительно более высокие показатели способностей к научению по сравнению с другими видами низших обезьян.
Павианы гамадрилы ввиду особенностей своей экологии в определенной степени являются уникальным видом приматов. Особенности образа жизни павианов гамадрилов, а именно жизнь большими стадами, включающими несколько сотен особей на открытых засушливых равнинах, предъявляют гораздо более серьезные требования к когнитивным способностям этих животных, чем таковые у обезьян, живущих небольшими семейными группами в изобилующих источниками пищи тропических лесах. Выживание павианов в суровых условиях среды требует способностей к запоминанию и узнаванию многочисленных особей своего стада, к распознаванию хищников и способностей к групповой обороне, к ориентации на местности, лишенной надежных ориентиров, к запоминанию места расположения источников пищи и воды, зачастую удаленных на несколько километров [Чалян, Мейшвили, 2019]. Проведенные нами ранее исследования показали, что павианы гамадрилы значительно опережают макаков резусов по многим когнитивным характеристикам: уровню исследовательской активности [Аникаев и др., 2016] и выполнению пространственных тестов. Наши исследования показали также, что павианы гамадрилы обладают в экспериментах высоким уровнем любознательности и быстротой реакции на требования экспериментатора, демонстрируют своего рода “сотрудничество” с экспериментатором. Именно это делает их более удобным объектом, по сравнению с макаками, при проведении экспериментов, требующих оценки когнитивной функции. При этом следует отметить, что когнитивные способности павианов гамадрилов мало изучены. В частности, отсутствуют исследования, связанные с обучением у павианов гамадрилов.
Целью данной работы является анализ обучения павианов гамадрилов навыкам разной степени сложности.
МЕТОДИКА
Объект исследования – 69 самцов павианов гамадрилов. Все самцы были рождены и содержались до начала эксперимента в семейных группах павианов гамадрилов в вольерах Научно-исследовательского института медицинской приматологии. Важно отметить, что в институте используется методика содержания обезьян, предполагающая минимальное вмешательство персонала в их жизнь, которое ограничивается ежедневным кормлением, уборкой помещений и ежегодным профилактическим осмотром группы. Фактически с точки зрения опыта контакта с людьми содержащиеся в вольерах Адлерского центра обезьяны соответствуют диким обезьянам, привезенным из мест естественного обитания. Исследование начиналось с отсаживания животных из родных групп и помещения их в индивидуальные клетки, где первоначально они проходили этап предварительного привыкания к индивидуальному содержанию и постоянному присутствию и близости человека, без какой-либо дрессировки и приручения. Никаких предварительных тестов и наблюдений, позволяющих оценить уровень их когнитивных способностей, не проводилось. После двухнедельного этапа привыкания обезьян к условиям индивидуального содержания начиналось тестирование животных. Наблюдения за ходом выполнения тестов осуществлялись с помощью видеокамеры. Тестирование начиналось с теста на взятие приманки, назначение которого состояло в проверке готовности животного к выполнению задач, его заинтересованности и мотивации. В качестве приманки использовались нарезанные яблоки.
Для обучения павианов был использован метод формирования навыка, который является частным случаем методики “установки на обучение” (object learning-set formation) [Harlow, 1949]. Данный метод широко применяется для оценки степени влияния на животных воздействий различного качества, анализа межвидовых, возрастных и половых различий [Chelonis, 2003; Jentsch et al., 2002]. Суть методики заключается в следующем: животному предоставляют возможность выбрать из двух емкостей, в одну из которых помещено пищевое подкрепление, продемонстрированное заранее. Проводилось обучение трем навыкам. При обучении первому навыку (Рис. 1 (а)), в качестве стимула выступали цвет и положение емкости с подкреплением: использовались черная и желтая емкость, подкрепление помещалось с левой от животного стороны в черную емкость. При обучении второму навыку (Рис. 1 (б)) стимулом являлось только положение емкости с подкреплением. Использовались две белые емкости, подкрепление помещалось с правой от животного стороны. При обучении третьему навыку (рис. 1 (в)) в качестве стимула применялся только цвет емкости с подкреплением. Использовались красная и синяя емкости, подкрепление помещалось под синюю емкость, положение которой периодически менялось, предъявление емкости с одной стороны подряд не превышало трех раз. Каждый навык выполнялся в течение одного дня по 50 предъявлений каждый, в случае с третьим навыком по 25 предъявлений на каждую сторону. На выполнение одного предъявления отводилась 1 мин. В течение одного дня каждое животное выполняло только один навык, обучение следующему навыку происходило через 2–3 дня. Обучение навыкам проводилось в последовательности, соответствующей их номерам. Фиксировались следующие варианты выполнения: верное решение (+), неверное решение (–) и отказ от решения (0).
Рис. 1.
Формируемые навыки: (а) – навык 1. (б) – навык 2. (в) – навык 3.
Fig. 1. Formed skills: (а) – skill 1, (б) – skill 2, (в) – skill 3.

Анализ полученных результатов осуществлялся по трем основным параметрам:
1. Успешность. Оценка успешности каждого животного осуществлялась с помощью трех показателей. Для установления потенциала способности каждого животного к обучению, т.е. оценки его обучаемости, в % подсчитывалось количество верных решений из определенного числа предъявлений (здесь и далее показатель № 1). Уровень обученности животного, характеризующий результат его обучения на определенном этапе, оценивался с помощью двух показателей – среднего значения числа верных решений, образующих непрерывные интервалы (здесь и далее показатель № 2) и максимального количества непрерывных верных решений (здесь и далее показатель № 3). Справедливость объединения показателей № 2 и № 3 в одну характеристику как критерия обученности, обоснована высокой корреляцией между ними (табл. 1).
Таблица 1.
Значения коэффициентов корреляции между показателями экспериментальной активности и успешности Table 1. The values of the correlation coefficients between the indicators of experimental activity and success
| Навык | 1 | 2 | 3 |
|---|---|---|---|
| Показатель успешности № 1/Показатель успешности № 2 | 0.5 | 0.7 | 0.8 |
| Показатель успешности № 2/Показатель успешности № 3 | 0.8 | 0.9 | 0.9 |
| Показатель успешности № 1/Показатель успешности № 3 | 0.7 | 0.8 | 0.7 |
| Показатель экспериментальной активности/показатель успешности № 1 | –0.9 | –0.7 | –0.9 |
| Показатель экспериментальной активности/показатель успешности № 2 | –0.3 | –0.4 | –0.6 |
| Показатель экспериментальной активности/показатель успешности № 3 | –0.6 | –0.4 | –0.5 |
2. Активность. Анализ данного параметра проводился путем подсчета числа отказов от выполнения задачи, что характеризует уровень поведенческой активности животного в эксперименте (далее экспериментальная активность). Как показывают наши исследования [Аникаев и др., 2016], данный фактор может выступать не только как индивидуальный, но и как видовой показатель.
3. Динамика. Анализ параметра осуществлялся путем вычисления числа верных решений и отказов в отдельности по каждому из 50 предъявлений по всем обезьянам с последующим формированием динамических рядов. Также проводился подсчет среднего значения по интервалам в 10 предъявлений, вычисление общего коэффициента прироста успешности по навыку и темпа прироста успешности по интервалам.
Статистический анализ полученных данных осуществлялся путем вычисления стандартной ошибки среднего, критерия Манна–Уитни (U), критерия Стьюдента (t), коэффициента корреляции Пирсона (R), коэффициента вариации (V) и коэффициента прироста (Kпр) пакетами программного обеспечения Microsoft Offiсe Excel 2010 и Statistica 6.0.
Все эксперименты проводились в соответствии с международными правилами по использованию лабораторных животных [National Research Council, 2010]. На проведение исследования было получено разрешение Комиссии по этике ФГБНУ “Научно-исследовательский институт медицинской приматологии”.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Основные показатели, полученные при анализе успешности, представлены в табл. 2. Как видно из таблицы, павианы гамадрилы обнаруживают способность к выработке навыков на положение приманки и цвет. Тем не менее очевидно, что величина показателей обучаемости и обученности животных варьирует в зависимости от сложности навыка. Так, при выполнении первого навыка, очень высокую обучаемость продемонстрировали около трети всех самцов, показавших от 76 до 100% правильных решений (рис. 2а). Самую большую категорию, включающую в себя 42% исследованных самцов, составили животные с высокой обучаемостью, которые сделали правильный выбор в пределах 51–75% предъявлений (рис. 2а). К числу наиболее плохо обучающихся животных можно было отнести 15% всех самцов, правильно выполнившие первый навык не более, чем в 25% всех предъявлений. Только 11% самцов сделали правильный выбор в интервале 26–50% предъявлений, что можно рассматривать как умеренную обучаемость. При выполнении второго навыка доля животных с очень высоким, высоким и умеренным уровнем обучаемости почти не отличается и приближается к 30%. При выполнении третьего навыка очень высокий уровень обучаемости был обнаружен только у 3 самцов (5.8%). Большинство животных могло быть отнесено по этому навыку к числу животных с высоким (55%) и умеренным (27.5%) уровнем обучаемости. Полученные результаты указывают на определенный спад успешности при выполнении навыков различной степени сложности. При этом по среднему числу верных решений (табл. 2) достоверные отличия имеются только между результатами по наиболее простому первому навыку и наиболее сложному третьему навыку (критерий Стьюдента, t = 2.15, p < 0.05).
Таблица 2.
Показатели успешности и экспериментальной активности при обучении навыкам Table 2. Indicators of success and experimental activity in learning skills
| Навык | 1 | 2 | 3 |
|---|---|---|---|
| Среднее количество верных решений (%) (показатель успешности № 1) | 58.6 ± 3.4 | 52.8 ± 3.3 | 49.7 ± 2.5 |
| Среднее количество верных решений, образующих непрерывные интервалы (%) (показатель успешности № 2) | 14.2 ± 2.0 | 8.5 ± 1.0 | 4.0 ± 0.3 |
| Среднее значение максимального количества непрерывных верных решений (%) (показатель успешности № 3) | 32.4 ± 2.9 | 21.5 ± 2.1 | 10.7 ± 1.0 |
| Среднее количество отказов от выполнения (%) (показатель экспериментальной активности) | 24.1 ± 3.8 | 13.5 ± 3.4 | 12.2 ± 3.7 |
Рис. 2.
Анализ распределения обезьян в зависимости от величины показателя: (а) – показатель успешности № 1. (б) – показатель успешности № 2. (в) – показатель успешности № 3. (г) – показатель экспериментальной активности.
Fig. 2. Analysis of the distribution of monkeys depending on the value of the indicator: (а) – success indicator No. 1, (б) – success indicator No. 2, (в) – success indicator No. 3, (г) – the indicator of experimental activity.
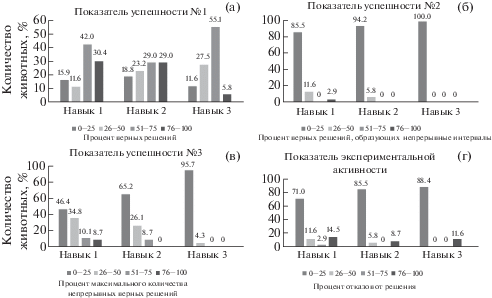
На рис. 2б показано в процентах число животных, демонстрировавших правильное решение в течение разного числа непрерывных подходов, при выполнении первого навыка у 85.5% животных продолжительность непрерывного ряда правильных решений варьировала от 0 до 25% общего числа предъявлений, и только 14.5% самцов демонстрировали более длинные непрерывные ряды успешно решенных предъявлений. При выполнении второго навыка соответствующее число животных составило 94.2 и 5.8%. При выполнении третьего навыка все 100% самцов демонстрировали только короткие ряды непрерывных успешных решений с численностью не более 25% общего числа предъявлений. При этом по среднему числу верных решений, образующих непрерывные интервалы, как и по средним значениям максимального числа непрерывных решений, различия между первым, вторым и третьим навыками достоверны и соответственно составили для второго показателя: 5.7% между первым и вторым навыком (критерий Стьюдента, t = 2.48, p < < 0.05), 4.5% между вторым и третьим навыком (критерий Стьюдента, t = 4.20, p < 0.05), 10.2% между первым и третьим навыком (критерий Стьюдента, t = 4.96, p < 0.05); для третьего показателя: 10.9% между первым и вторым навыком (критерий Стьюдента, t = = 3.47, p < 0.05), 10.8% между вторым и третьим навыком (критерий Стьюдента, t = 4.98, p < 0.05), 21.7% между первым и третьим навыком (критерий Стьюдента, t = 7.55, p < 0.05).
Совокупность этих данных позволяет заключить, что самцы павианов гамадрилов однозначно менее успешны при обучении третьему навыку по сравнению с первым и вторым навыком. Тенденция снижения успешности обучения от первого к третьему навыку подтверждается также данными коэффициента прироста, вычисленными по результатам анализа данных, полученных по всем трем навыкам. Коэффициент прироста (Кпр) среднего числа правильных решений, подсчитанный по результатам анализа последовательно трех навыков, составил Кпр = –8.0, Кпр величины непрерывных рядов правильных решений – –46.6, Кпр максимальной величины рядов правильных решений – –42.5.
Анализ коэффициентов корреляции между различными показателями успешности самцов павианов гамадрилов (табл. 1) показывает, что по всем трем навыкам наблюдается высокая степень зависимости между исследуемыми показателями. Эти данные свидетельствуют об очевидном факте: уровень обученности напрямую зависит от уровня обучаемости, результат зависит от потенциала.
Для анализа динамики успешности при обучении навыкам производился подсчет суммы верных решений в отдельности по каждому из 50 предъявлений по всем обезьянам. Установлено, что значимые различия между суммами верных решений наблюдаются между рядами значений по первому и второму навыку (критерий Стьюдента, t = 2.44, p < 0.05) и по первому и третьему навыку (критерий Стьюдента, t = 3.14, p < 0.05). На рис. 3а показано выраженное в процентах среднее число верных решений по пяти интервалам в 10 предъявлений каждый, а в табл. 3 представлены значения темпов прироста успешности, вычисленные для каждого из пяти интервалов. Анализ динамики успешности при обучении первому навыку показывает, что имеет место резкое возрастание числа правильных решений павианов в первых 20 предъявлениях, что соответствует коэффициенту прироста во втором интервале в максимальные 48.4%. Начиная с 21-го предъявления темпы возрастания успешности снижаются и достигают плато на 40-м предъявлении. Динамика формирования второго навыка практически полностью соответствует динамике первого навыка. При этом статистически значимые различия в обоих случаях наблюдаются только между данными по третьему и четвертому интервалу. В целом коэффициент прироста процента верных решений составил 3.5 по первому навыку, 1.7 – по второму навыку, что свидетельствует о тенденции роста в числе правильных решений при их выполнении.
Рис. 3.
Анализ динамики при обучении навыкам: (а) – динамика успешности (по среднему количеству верных решений за интервал). (б) – динамика экспериментальной активности (по среднему количеству отказов от решения за интервал).
Fig. 3. Analysis of the dynamics in learning skills: (а) – dynamics of success (according to the average number of correct decisions per interval). (б) – dynamics of experimental activity (according to the average number of refusals from the solution for the interval).
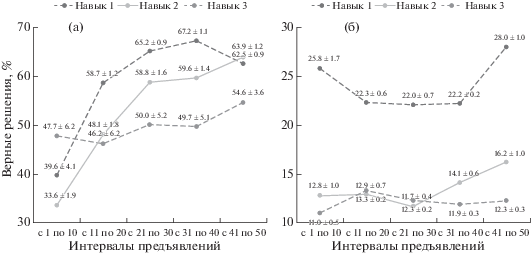
Таблица 3.
Значения коэффициента вариации по исследуемым параметрам Table 3. The values of the coefficient of variation for the parameters studied
| Навык | 1 | 2 | 3 |
|---|---|---|---|
| Коэффициент вариации (V) по показателю успешности № 1 (%) | 47.7 | 51.1 | 41.3 |
| Коэффициент вариации (V) по показателю успешности № 2 (%) | 118.6 | 100.3 | 56.7 |
| Коэффициент вариации (V) по показателю успешности № 3 (%) | 75.1 | 82.0 | 75.5 |
| Коэффициент вариации (V) по показателю экспериментальной активности (%) | 131.3 | 208.3 | 255.3 |
В отличие от двух первых навыков, динамика формирования третьего навыка развивается особенным образом. Начиная с первых предъявлений, павианы выходят первоначально на относительно высокий уровень успешности, который плавно возрастает по мере возрастания числа предъявлений, но не достигает уровня успешности, отмечаемой в соответствующих предъявлениях по первому и второму навыку. Кпр, вычисленный для третьего навыка, в целом составил –0.2. На фоне визуально наблюдаемого роста показатели темпа прироста и критерия Манна–Уитни (табл. 3) указывают на стойкое достоверное колебание среднего числа верных решений по этому навыку.
Сходство и различия в динамике формирования трех разных навыков подтверждаются анализом коэффициентов корреляции между рядами значений сумм верных решений по каждому предъявлению. Имеется сильная взаимосвязь (R = 0.8) между значениями сумм верных решений по каждому предъявлению по первому и второму навыку, тогда как между данными по второму и третьему, первому и третьему навыками коэффициенты корреляции (R = 0.1) указывают на практически полное отсутствие зависимости. Эти данные можно интерпретировать следующим образом: показатели динамики успешности обучения первому и второму навыку в определенной степени сходны, но отличны от таковых по третьему навыку.
В целом совокупность представленных данных позволяет утверждать наличие сходной положительной динамики успешности обучения первому и второму навыку у исследуемых животных. При этом данные, полученные по третьему навыку, не позволяют сделать вывод о положительной динамике, но обнаруживают закономерное колебание числа верных решений по мере обучения.
Для анализа индивидуальных различий при обучении навыкам в качестве показателя степени разброса и сравнительной характеристики использовался коэффициент вариации. Допущена следующая интерпретация данного показателя: чем ниже значение коэффициента, тем более однородны данные и, следовательно, меньше проявление индивидуальных различий. Следует отметить, что в некоторых случаях данный показатель превышает 100%, это объясняется наличием в выборке значений, сильно отличающихся от среднего. В целом вычисленные коэффициенты вариации по всем показателям (табл. 4) указывают на высокую степень индивидуальных различий. Однако значения коэффициента вариации по показателю № 1 и № 3 указывают на сравнительно более низкий размах изменчивости в успешности у исследуемой группы при обучении первому и третьему навыку, чем при обучении второму навыку. При этом значения коэффициента по показателю № 2 свидетельствуют о постепенном спаде уровня индивидуальных различий от первого к третьему навыку. Эти данные подтверждаются анализом распределения испытуемых животных в зависимости от величины каждого определенного показателя, выраженного в процентах (рис. 2б, 2в). В целом можно заключить, что наиболее низкий уровень индивидуальной изменчивости наблюдается при выполнении третьего навыка, при этом большинство животных относится к категории с крайне низкими показателями успешности № 2 и № 3. Этим дополнительно подтверждается сложность третьего навыка.
Таблица 4.
Показатели достоверности динамики успешности и активности Table 4. Indicators of the reliability of the dynamics of success and activity
| Показатель | Разница между средними (%) | Темп прироста (Тпр) (%) | Критерий Манна–Уитни (U) | ||
|---|---|---|---|---|---|
| Успешность | Навык 1 | 1/2 интервал | 19.1 | 48.4 | 4.0 (p > 0.05) |
| 2/3 интервал | 6.5 | 11.1 | 8.0 (p > 0.05) | ||
| 3/4 интервал | 2.0 | 3.1 | 32.5 (p < 0.05)* | ||
| 4/5 интервал | –4.8 | –7.1 | 15.0 (p > 0.05) | ||
| Навык 2 | 1/2 интервал | 14.5 | 43.1 | 5.5 (p > 0.05) | |
| 2/3 интервал | 10.7 | 22.3 | 6.0 (p > 0.05) | ||
| 3/4 интервал | 0.7 | 1.2 | 44.5 (p < 0.05)* | ||
| 4/5 интервал | 4.3 | 7.3 | 21.5 (p > 0.05) | ||
| Навык 3 | 1/2 интервал | –1.4 | –3.0 | 47.0 (p < 0.05)* | |
| 2/3 интервал | 3.8 | 8.2 | 42.0 (p < 0.05)* | ||
| 3/4 интервал | –0.3 | –0.6 | 47.0 (p < 0.05)* | ||
| 4/5 интервал | 4.9 | 9.9 | 39.5 (p < 0.05)* | ||
| Активность | Навык 1 | 1/2 интервал | –3.5 | –13.5 | 28.0 (p < 0.05)* |
| 2/3 интервал | –0.3 | –1.3 | 47.0 (p < 0.05)* | ||
| 3/4 интервал | 0.1 | 0.7 | 44.0 (p < 0.05)* | ||
| 4/5 интервал | 5.8 | 26.1 | 0.0 (p > 0.05) | ||
| Навык 2 | 1/2 интервал | 0.1 | 1.1 | 41.5 (p < 0.05)* | |
| 2/3 интервал | –1.2 | –9.0 | 30.0 (p < 0.05)* | ||
| 3/4 интервал | 2.3 | 19.8 | 16.0 (p > 0.05) | ||
| 4/5 интервал | 2.2 | 15.5 | 29.5 (p < 0.05) | ||
| Навык 3 | 1/2 интервал | 2.3 | 21.1 | 12.0 (p > 0.05) | |
| 2/3 интервал | –1.0 | –7.6 | 20.0 (p > 0.05) | ||
| 3/4 интервал | –0.4 | –3.5 | 32.5 (p < 0.05)* | ||
| 4/5 интервал | 0.4 | 3.7 | 36.5 (p < 0.05)* | ||
Исходя из выше изложенных данных, можно сделать вывод, что большинство из исследуемых павианов гамадрилов успешно обучаются первому и второму навыку и хуже обучаются третьему навыку (за исключением нескольких “выдающихся” особей). Причем наши наблюдения показывают, что показатели отдельных особей не обязательно подчиняются общей тенденции спада успешности при последовательном обучении навыкам. В ряде случаев животные были малоуспешны при обучении первому и третьему навыку, но давали хорошие результаты при выполнении второго навыка, в редких случаях у обезьян обнаруживался рост успешности от первого к третьему навыку. Все это свидетельствует о том, что способности к обучению разным навыкам слабо связаны.
Для исследования экспериментальной активности павианов были проанализированы случаи отказов от выполнения экспериментальной задачи. Данные, полученные при анализе количества отказов от выполнения задачи (табл. 2), указывают на то, что в среднем самцы павианов гамадрилов демонстрируют высокий уровень экспериментальной активности. При этом по мере обучения навыкам наблюдается сокращение числа отказов и соответствующий рост активности. Этот факт подтверждается низким значением коэффициента прироста средних показателей числа отказов при выполнении каждого навыка (Кпр = –28.9). В среднем наибольшее число отказов было зафиксировано при выполнении первого навыка (24.1 + 3.8) (табл. 2). При выполнении второго навыка среднее число отказов было достоверно меньше (критерий Стьюдента, t = 2.06, p < 0.05), также, как и при выполнении третьего навыка (критерий Стьюдента, t = 2.23, p < 0.05). При этом разница между средним число отказов по второму и третьему навыку составила 1.3% (критерий Стьюдента, t = 0.27, p > 0.05).
Значения коэффициентов вариации по показателю экспериментальной активности (табл. 4) позволяют заключить следующее. Во-первых, имеет место широкий размах индивидуальных различий по данному критерию. Во-вторых, размах индивидуальных различий в проявлении экспериментальной активности явно возрастает по мере усложнения навыка. В-третьих, в исследуемой группе павианов гамадрилов имеются особи, чьи результаты по числу отказов резко отличаются от среднего значения. Анализ распределения обезьян в зависимости от числа отказов от выполнения (рис. 2г) указывает на постепенное разделение по мере выполнения навыков на 2 категории, соответствующие двум крайним значениям показателя экспериментальной активности. Основное число животных при обучении всем трем навыкам демонстрируют низкий процент отказов и соответственно высокий уровень экспериментальной активности. При этом если в случае с первым и вторым навыком имеются особи также со средним и низким уровнем активности, то в случае с третьим навыком исследуемые животные строго разделились на категорию с высоким уровнем активности (88.4%) и крайне низким уровнем активности (11.6%). Следует отметить, что не все животные проявляли активность в точности в соответствии с обнаруженной общей тенденцией роста после первого навыка. Некоторые животные демонстрировали высокий уровень экспериментальной активности (0% отказов) по всем трем навыкам, небольшое число особей показали нулевой уровень активности по всем трем навыкам (100% отказов), имелись животные, у которых по мере обучения наблюдался спад активности.
Для анализа динамики активности при обучении каждому из навыков производился подсчет суммы отказов в отдельности по каждому из 50 предъявлений по всем обезьянам. При этом значимые различия наблюдаются по рядам значений, полученным по всем трем навыкам. Критерий Стьюдента составил t = 15.47 (p < 0.05) между рядами значений по первому и второму навыку, t = 3.07 (p < 0.05) – по второму и третьему навыку и t = 20.61 (p < < 0.05) – по первому и третьему навыку. Это может свидетельствовать о том, что среднее число отказов по каждому предъявлению при обучении первому, второму и третьему навыку достоверно различно, что в целом соотносится с результатами, полученными при анализе средних значений экспериментальной активности.
Данные коэффициентов корреляции между рядами значений сумм отказов по каждому предъявлению по разным навыкам свидетельствуют о наличии умеренной (R = 0.5) взаимосвязи между рядами значений по первому и второму навыку. В то же время между данными по второму и третьему, первому и третьему навыкам коэффициенты корреляции (R = 0.1 и R = 0.0) указывают на практически полное отсутствие зависимости. Эти данные можно интерпретировать следующим образом: показатели динамики активности при обучении первому и второму навыку в некоторой степени сходны, но отличны от таковых по третьему навыку.
Значения коэффициента вариации процента отказов по отдельным предъявлениям указывают на незначительные колебания активности животных по разным предъявлениям. Так, сравнительно наибольший размах изменчивости процента отказов по разным предъявлениям отмечается для второго навыка (V = 21.0), затем для первого навыка (V = = 16.1) и для третьего навыка (V = 10.5). Данные коэффициента прироста также не указывают на существенные изменения в динамике (Кпр = 1.1 для первого навыка, Кпр = 1.3 для второго навыка и Кпр = 1.0 для третьего навыка). Тем не менее визуальный анализ данных (рис. 3б), а также показатели темпа прироста и критерия Манна–Уитни (табл. 4) действительно указывают на сходство динамики активности по первому и второму навыку и отличие таковой по третьему навыку. Так, по первому и второму навыку наблюдается достоверный спад числа отказов в первой половине предъявлений и небольшой статистически незначимый рост во второй половине. Другими словами, активность в течение выполнения первого и второго навыка сначала растет и затем ближе к концу обучения незначительно падает. При этом динамика активности по третьему навыку отличается резким (следует отметить, недостоверным ростом числа отказов) и последующим плавным (достоверным) спадом этого показателя. То есть в начале обучения третьему навыку активность резко падает и затем плавно поднимается до значения, близкого к исходному.
Степень зависимости уровня успешности от уровня экспериментальной активности по коэффициентам корреляции отражена в табл. 3. Наиболее чувствительным к уровню экспериментальной активности является показатель обучаемости. То есть число верных решений, что вполне закономерно, обратно пропорционально числу отказов, и чем активнее животные, тем выше процент верных решений. Причем данная взаимосвязь проявляется на высоком уровне вне зависимости от выполняемого навыка. Менее чувствительны к уровню активности оба показателя обученности. В данном случае значения корреляции колеблются от низкого до среднего, что соответствует сравнительно слабому влиянию уровня активности животного на число непрерывно выполненных верных решений. Эти данные позволяют заключить, что степень активности животного в эксперименте является важным фактором, влияющим на обучаемость, но в меньшей степени сказывается на уровне обученности.
Данные по корреляции между показателями динамики успешности и динамики активности для всех трех навыков указывают на крайне низкую взаимосвязь или ее отсутствие (R = –0.23 для первого навыка, R = 0.11 для второго навыка и R = –0.08 для третьего навыка). То есть динамика успешности по числу верных решений в каждом предъявлении слабо связана с динамикой активности, выраженной в числе отказов в каждом предъявлении, вне зависимости от формируемого навыка.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
При анализе полученных результатов следует прежде всего обратить внимание на высокую активность павианов гамадрилов. Экспериментально наивные животные, впервые попавшие в условия индивидуального содержания, в большинстве случаев уже с первого предъявления демонстрировали высокую активность по выбору емкости с приманкой. Следует отметить, что при среднем высоком уровне активности павианов гамадрилов отмечается большой размах индивидуальных различий по всем трем навыкам. От 71 до 88% (в зависимости от навыка) особей демонстрировали менее 25% отказов от выполнения задачи каждый, что соответствует высокому уровню экспериментальной активности. Остальные особи распределились по категориям средней и низкой активности. Сравнение показателей активности в динамике показывает возрастание экспериментальной активности обезьян в ходе эксперимента, что объясняется привыканием животных к его условиям. То есть отказы животных от выполнения задачи, наблюдаемые чаще всего при обучении первому навыку, следует, по-видимому, рассматривать в качестве реакции на новую для них ситуацию, в частности, на присутствие экспериментатора во время постановки задачи и наличие самой экспериментальной установки.
Наиболее примечательным результатом проведенного исследования является установленный факт очевидного отличия результатов, полученных при обучении павианов гамадрилов третьему навыку, от результатов обучения первому и второму навыку, как по показателям успешности, так и по динамике формирования. Вероятной причиной обнаруженной сложности в обучении павианов гамадрилов третьему навыку, по нашему мнению, являются особенности стимулов, используемых для выработки навыков. В случае с первым навыком у животных имелись два стимула: цвет – желтый или черный и положение емкости, при формировании второго навыка стимулом служило только положение, при формировании третьего только цвет – красный либо синий. Результаты экспериментов показывают, что у исследуемых животных наиболее успешно формируется ассоциация с положением емкости, подкрепленная желтым или черным цветом (навык № 1), несколько менее успешно – ассоциация только с положением емкости (навык № 2). Минимальная успешность обнаруживается в отношении выбора емкостей, окрашенных в красный или синий цвет (навык № 3), способность к научению различения которых была обнаружена только у трех самцов (4.3% исследованных самцов). Затруднения, возникающие у павианов при выборе красной или синей емкости, подтверждаются следующим наблюдением: при обучении третьему навыку для животных были типичными две стратегии. Первая: после одного-двух выборов верной емкости на третье предъявление павианы выбирали емкость с тем же положением, но отличающуюся по цвету, после чего на четвертое предъявление искали подкрепление в противоположном направлении, в случае ее нахождения ситуация повторялась. Вторая (наблюдалась реже): независимо от цвета животные в течение всего процесса формирования навыка выбирали емкость, находящуюся строго в одном положении. Обе стратегии в совокупности отразились на графическом анализе динамики обучения третьему навыку, где четко прослеживается периодичность подъемов и спадов среднего числа верных решений без явного роста успешности.
Таким образом, анализ показывает, что различные признаки имеют принципиально разное значение в качестве стимулов при научении павианов. Наиболее значимым, по-видимому, является стимул положения приманки. Использование этого стимула в сочетании с использованием желтого и черного цветов (навык № 1) и в чистом виде (навык № 2) достаточно легко приводит к формированию навыка. Практически малозначимым в качестве стимулов, по-видимому, является красный и синий цвета, различение которых без подкрепления положением было затруднительным для павианов.
Обнаруженные различия в отношении павианов гамадрилов к разным стимулам биологически обоснованы. В естественных условиях, павианы гамадрилы живут в достаточно сложной среде в условиях полупустыни, где ресурсы, а именно источники еды и воды, а также укрытия для ночлега распределены крайне неравномерно. Исходя из этого очевидно, что способность к запоминанию мест локализации пищи и воды, мест ночлега является для них жизненно важной стратегией, определяющей саму возможность их выживания. Как показали полевые наблюдения [Schreier, Grove, 2010; Sigg, Stolba, 1981], павианы обладают хорошей пространственной памятью и способностью к созданию “ментальной карты”, позволяющей в разное время года находить на местности необходимые ресурсы, совершая для этого многокилометровые переходы. Обнаруженную в эксперименте способность павианов к выработке навыка на положение приманки следует, по-видимому, рассматривать как проявление эволюционно выработанных способностей к запоминанию положения целей и ориентации на местности.
Аналогично биологически обоснованной представляется способность павианов гамадрилов к различению емкостей желтого и черного цветов. Как известно, большинство приматов, прежде всего приматов Старого Света, трихроматичны [Osorio, Vorobyev, 2008]. Некоторое исключение из общей трихроматичности приматов обнаруживается у лемуров и обезьян Нового Света, у которых обнаружена либо трихроматичность (Alouatta caraya) [AraUJo et al., 2008], (Saguinus oedipus) [Savage et al., 1987], либо сочетание дихроматичных самцов и дихроматичных/трихроматичных самок (Callithrix penicillata) [Pessoa et al., 2005]. Существует ряд гипотез, раскрывающих эволюционное значение трихроматичности приматов [Rowe 2002; Fernandez, Morris, 2007]. В наиболее распространенном варианте трихроматичность приматов рассматривается как адаптивное приспособление к обнаружению спелых фруктов среди листвы [Smith et al., 2003]. В исследовании когнитивной деятельности макак-резусов (Macaca mulatta) и бурых капуцинов (Cebus apella) при дифференцировании по величине пар двумерных и трехмерных зрительных стимулов и при распознавании множеств в виде плоскостных и объемных геометрических фигур разного цвета обнаруживаются свидетельства того, что выполнение задач осуществлялось успешно независимо от цвета [Малюкова и др., 2006а; Малюкова и др., 2006б]. Адаптивные преимущества трихроматичности приматов подтверждены в ряде экспериментов с обезьянами тех видов, у которых встречается и дихроматичность, и трихроматичность. Установлено в частности, что трихроматичные особи тамаринов более эффективно осуществляли поиск и отбор зрелых плодов по сравнению с дихроматичными особями того же вида [Smith et al., 2003]. Трихроматичные тамарины более эффективно, чем дихроматы, осуществляли различение стимулов оранжевого и зеленого цвета [Pessoa et al., 2003]. Вместе с тем в работе, посвященной изучению способности трихроматичных тамаринов использовать различные ориентиры для поиска пищи, исследователи указывают, что при использовании в качестве стимула только цвета у животных возникали определенные сложности [Deipolyi et al., 2001].
Павианы гамадрилы являются трихроматами и демонстрируют в естественных условиях обитания преимущественную фруктоядность. Спелые плоды деревьев являются для них предпочитаемым видом корма, ради которого они совершают многокилометровые переходы. Можно предположить, что способность восприятия разных цветов у них варьирует в пользу тех цветов, различение оттенков которых позволяет им более эффективно отыскивать спелые плоды в листве деревьев, отличать созревшие плоды от незрелых. Биологически более значимые цвета, в данном случае желтый, то есть цвет спелых фруктов, могут быть приоритетными для обезьян. Соответственно желтый цвет одной из емкостей, использующихся при выработке первого навыка, может рассматриваться как фактор, облегчающий научение, который выступает в данном случае в качестве дополнительного стимула к основному – положению приманки. В случае с выработкой третьего навыка отсутствие основного стимула (фиксированного положения приманки) и использование малозначимых для павианов цветов емкостей (красного и синего) приводят к неуспешности большинства животных.
Если отличия в процессе и результатах обучения третьему навыку носят очевидный характер, то различия между данными по первому и второму навыку не столь явно выражены. Хотя по второму навыку относительно первого навыка имеет место спад значений по всем показателям успешности, отсутствуют достоверные отличия показателя обучаемости (среднего значения верных решений) на фоне достоверных отличий в показателях обученности (числе непрерывных верных решений). Эти различия свидетельствуют о возрастании числа ошибок у павианов при научении второму навыку, что является, по-видимому, следствием трудностей, возникающих у них при переходе от научения более простому навыку, ориентирами при выполнении которого являются цвет и положение (первый навык), к более сложному навыку с ориентиром – только положением (второй навык). Следует отметить, что переход к обучению второму навыку сопровождается достоверным ростом показателя экспериментальной активности, который в дальнейшем (при обучении третьему навыку) остается на относительно постоянном высоком уровне. Таким образом, при переходе от первого навыка ко второму навыку рост активности сопровождается спадом успешности, что выражается, в частности, достоверным снижением среднего количества верных решений, образующих непрерывные интервалы и среднего значения максимального числа непрерывных верных решений. То есть можно утверждать о наличии спада обученности на фоне роста экспериментальной активности. Тем не менее значения коэффициентов корреляции свидетельствуют о том, что чем активнее животное ведет себя в эксперименте, тем выше уровень его обучаемости, что, в свою очередь, существенно влияет на уровень обученности. Однако степень взаимосвязи между активностью и обученностью животного слабая. В связи с этим можно утверждать, что при обучении павианов гамадрилов демонстрируемый высокий уровень экспериментальной активности не является гарантом достижения ими высоких значений показателей обученности. Исходя из этого, вероятно, является целесообразным рассматривать рост активности и спад успешности по мере обучения, как два не связанных друг с другом события.
Определенный вклад в снижение успешности обезьян при обучении второму навыку, по-видимому, вносит тот факт, что второй навык по своей постановке являлся в некотором роде обратным первому навыку, так как положение емкости с приманкой менялось на противоположное. Соответственно можно предположить, что задача для животных усложнялась необходимостью переучивания. Подтверждением этому могут служить результаты небольшого числа особей, которые действительно в процессе обучения второму навыку продолжали стойко (иногда в течение всех 50 предъявлений) выбирать емкость, расположенную с той стороны, где подкрепление находилась при обучении первому навыку. В ряде исследований, посвященных проблемам переучивания, указывается на существенное отставание обезьян при обучении обратному навыку [Bartus et al., 1979; Costa et al., 2015; Chau et al., 2015; Judge et al., 2011; Lai et al., 1995; Makori et al., 2013]. Устойчивость сформированных навыков и степень индивидуальных различий по данному показателю у павианов гамадрилов, несомненно, имеют место и требуют отдельных исследований.
Помимо анализа процесса обучения навыкам, в данной работе мы хотели бы уделить внимание методической составляющей эксперимента и позволить себе озвучить некоторые рекомендации.
Как уже отмечалось, павианы гамадрилы, несомненно, демонстрируют высокий уровень экспериментальной активности, более высокий по сравнению с другими популярными лабораторными приматами – макаками резусами. В совокупности со сравнительно высокими показателями по обучаемости и обученности, полученными в данной работе, этот факт делает их более предпочтительной моделью для осуществления различного рода когнитивных исследований.
В качестве анализа процесса обучения использовались два критерия, обозначенные как “обучаемость” и “обученность”. Как уже указывалось выше, свидетельств подобного разбора показателей успешности ранее нами обнаружено не было. При этом показатели обученности являются более точным отражением результата обучения, особенно в экспериментах по обучению с фиксированным количеством предъявлений.
Результаты данного исследования показали, что павианы гамадрилы способны выдержать обучение навыку в течение 50 предъявлений в одном сеансе. Для большинства особей этого количества предъявлений может быть достаточно для формирования простого навыка. Однако анализ динамики экспериментальной активности указывает на резкий достоверный рост отказов (по первому и второму навыку) в интервале от 30-го до 40-го предъявления. Данный факт говорит об очевидной нецелесообразности повышения количества предъявлений за один сеанс и возможной необходимости его уменьшения.
ЗАКЛЮЧЕНИЕ
В рамках данной работы у 69 взрослых самцов павианов гамадрилов зафиксирован высокий уровень экспериментальной активности по трем навыкам. От первого к третьему навыку наблюдается повышение уровня экспериментальной активности исследуемых обезьян.
Отмечены высокие значения показателей успешности по первому навыку, где в качестве стимула использовались положение и цвет (желтый и черный) емкости с подкреплением, и по второму навыку, в котором в качестве стимула использовалось только положение емкости с подкреплением. Установлено, что исследуемые животные значительно хуже обучаются третьему навыку, где в качестве стимула использовался только красный и синий цвет емкости с подкреплением.
По мере обучения навыкам от первого к третьему зафиксирован спад по показателям обучаемости и обученности, что подтверждает предполагаемое возрастание уровня сложности заложенных в их основу принципов. Обнаружена высокая степень зависимости между уровнем экспериментальной активности и обучаемостью, а также между обучаемостью и обученностью. При этом между экспериментальной активностью и обученностью корреляция слабая.
В исследуемой группе павианов гамадрилов установлен широкий размах индивидуальных различий в разной степени по всем анализируемым показателям.
Грант: Российский фонд фундаментальных исследований 15-04-05283.
Список литературы
Аникаев А.Е., Чалян В.Г., Мейшвили Н.В. Исследование способности к формированию навыка у павианов гамадрилов. “Фундаментальные и прикладные аспекты медицинской приматологии”. 2016. 231–239.
Беляева А.Г., Штемберг А.С., Носовский А.М., Васильева О.Н., Гордеев Ю.В., Кудрин В.С., Навкевич В.В., Красавин Е.А., Тимошенко Г.Н., Лапин Б.А., Базян А.С. Воздействие высокоэнергетических протонов и ионов углерода на когнитивные функции обезьян и содержание моноаминов и их метаболитов в периферической крови. Нейрохимия. 2017. 11 (1): 1–8.
Малюкова И.В., Черникова Н.А., Сикетин В.А. Особенности распознавания и сопоставления по величине двумерных изображений и трехмерных объектов разного цвета у низших обезьян. Журнал эволюционной биохимии и физиологии. 2006. 42 (3): 257–262.
Малюкова И.В., Черникова Н.А., Сикетин В.А. Исследование интеллектуальной деятельности при распознавании счетных множеств двумерных и трехмерных стимулов разного цвета у низших обезьян. Журнал эволюционной биохимии и физиологии. 2006. 42 (5): 491–500.
Чалян В.Г., Мейшвили Н.В. Павианы гамадрилы в лесах Черноморского побережья Кавказа. Изд-во “ЯСК”. 2019.
AraUJo A.C., Didonet J.J., AraUJo C.S., Saletti P.G., Borges T.R., Pessoa V.F. Color vision in the black howler monkey (Alouatta caraya). Visual neuroscience. 2008. 25 (3): 243–248.
Barney B.J., Amici F., Aureli F., Call J., Johnson V.E. Joint baynesian modeling of binomial and rank data for primate cognition. Journal of the American Statistical Association. 2015. 110 (510): 573–582.
Bartus R.T., Dean III R.L., Fleming D.L. Aging in the rhesus monkey: effects on visual discrimination learning and reversal learning. Journal of Gerontology. 1979. 34 (2): 209–219.
Beran M.J., Menzel Ch.R., Parrish A.E., Perdue B., Sayers K., Smith J.D., Washburn D.A. Primate cognition: attention, episodic memory, prospective memory, self-control, and metacognition as examples of cognitive control in non-human primates. Wiley Interdisciplinary Reviews: Cognitive Science. 2016. 7 (5): 294–316.
Bergman Th.J., Kitchen D.M. Comparing responses to novel objects in wild baboons (Papio ursinus) and geladas (Theropithecus gelada). Animal Cognition. 2009. 12: 63–73.
Boschin E.A., Piekema C., Buckley M.J. Essential functions of primate frontopolar cortex in cognition. Proceedings of the National Academy of Sciences. 2015. 112 (9): 1020–1027.
Byrne R.W., Bates L.A. Primate social cognition: uniquely primate, uniquely social, or just unique? Neuron. 2010. 65 (6): 815–830.
Byrne R.W., Corp G.R. Neocortex size predicts deception rate in primates. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2004. 271 (1549): 1693–1699.
Callahan P.M., Hutchings E.J., Kille N.J., Chapman J., Terry A.J. Positive allosteric modulator of alpha 7 nicotine-acetylcholine receptors, PNU-120596 augments the effects of doneperezil on learning and memory in aged rodents and non-human primates. Neuropharmacology. 2013. 67: 201–212.
Chau B.K., Sallet J., Papageorgiou C.K., Noonan M.P., Bell A.H., Waiton M.E., Rusworth M.F.S. Contrasting roles for orbitofrontal cortex and amygdala in credit assignment and learning in macaques. Neuron. 2015. 87 (5): 1106–1118.
Chelonis J.J., Gillam M.P., Paule M.G. The effects of prenatal cocaine exposure on reversal learning using a simple visual discrimination task in rhesus monkeys. Neurotoxicology and teratology. 2003. 25 (4): 437–446.
Costa V.D., Trau V.L., Turchi J., Averbeck B.B. Reversal learing and dopamine: a Bayesian perspective. The Journal of Neuroscience. 2015. 35 (6): 2407–2416.
Curtis B., Liberato N., Rulieu M., Morrisroe K., Kenney C., Yutuc V., Ferrier C., Marti C.N., Mandell D., Burbacher Th.M., Sackett G.P., Hewitson L. Examination of the safety of pediatric vaccine schedules in a nonhuman primate model: assessments of neurodevelopment, learning and social behavior. Environ. Health Perspect. 2015. 123 (6): 579–589.
Deaner R.O., Isler K., Burkart J., van Schaik C. Overall brain size and not encephalisation quotient, best predicts cognitive ability across non-human primates. Brain, Behavior and Evolution. 2007. 70 (2): 115–124.
Deipolyi A., Santos L., Hauser M.D. The role of landmarks in cotton-top tamarin spatial foraging: evidence for geometric and non-geometric features. Animal Cognition. 2001. 4 (2): 99–108.
Fagot J., Bonte E. Automated testing of cognitive perfomance in monkeys: Use of a battery of computerized test systems by a troop of semi-free-ranging baboons (Papio papio). Behaviour Research Methods. 2010. 42 (2): 507–516.
Fagot J., De Lillo C. A compatible study of working memory: Immediate serial spatial recall in baboons (Papio papio) and humans. Neuropsychologia. 2011. 49 (14): 3870–3880.
Fagot J., Gulistrand J., Kemp C., Defiles C., Meraouche M. Effects of freely accessible computerized test systems on the spontaneous behaviors and stress level of Guinea baboons (Papio papio). American Journal of Primatology. 2014. 76 (1): 56–64.
Fagot J., Cook R.G. Evidence for large long-term memory capacities in baboons and pigeons and its implications for learning and the evolution of cognition. Proceedings of the National Academy of Sciences. 2006. 103 (46): 17564–17567.
Fernandez A.A., Morris M.R. Sexual selection and trichromatic color vision in primates: statistical support for the preexisting-bias hypothesis. The American Naturalist. 2007. 170 (1): 10–20.
Flemming T.M., Thompson R.L.R., Fagot J. Baboons like humans solve analogy by categorical abstraction of relations. Animal cognition. 2013. 16 (3): 519–524.
Gale J.T., Lee K.H., Amirnovin R., Roberts D.W., Williams Z.M., Blaha Ch.D., Eskaudar E.N. Electrical stimulation – evoked dopamine release in the primate striatum. Stereotactic and Functional Neurosurgery. 2013. 91 (6): 355–363.
Goujon F., Fagot J. Learning of spatial statistcs in nonhuman primates: contextual cueing in baboons (Papio papio). Behavioural Brain Research. 2013. 247: 101–109.
Hannagan Th., Ziegler J., Dufau S., Fagot J., Grainger J. Deep learning of orphographic representations in baboons. Plos One. 2014. 9 (1): e84843.
Harlow H.F. The formation of learning sets. Psychological review. 1949. 56 (1): 51.
Huguet P., Barbet I., Belleitier C., Monteil J.M. Cognitive control under social influence in baboons. Journal of Experimental Psychology: General. 2014. 143 (6): 2067–2073.
Jentsch J.D., Olausson P., De La Garza II R., Taylor J.R. Impairments of reversal learning and response perseveration after repeated, intermittent cocaine administrations to monkeys. Neuropsychopharmacology. 2002. 26 (2): 183–190.
Joly M., Ammersdorfer S., Schmidtke D., Zimmermann E. Touchscreen Based cognitive task reveal age-related impairment in a primate aging model, the Grey Mouse Lemur (Microcebus murinus). PLOS ONE. 2014. 9 (10): e109393.
Judge P.G., Evans D.W., Schroepfer K.K., Gross A.C. Perseveration on a reversal-learning task correlates with rates of self-directed behavior in nonhuman primates. Behavioural Brain Research. 2011. 222 (1): 57–65.
Krause M.R., Zanos Th.P., Csorba B.A., Pilly P.K., Choe J., Phillips M.F., Datta A., Pack Ch.C. Transcranial direct current stimulation facilitates associative learning and alters functional connectivity in the primate brain. Current Biology. 2017. 27 (20): 3086–3096.
Lai Z.C., Moss M.B., Killiany R.J., Rosene D.L., Herndon J.G. Executive system dysfunction in the aged monkey: spatial and object reversal learning. Neurobiology of aging. 1995. 16 (6): 947–954.
Linke M., Broker F., Ramskar M., Baayen H. Are baboons learning “orthographic” representations? Probably not. Plos One. 2017. 12 (8): e0183876.
Makori N., Watson R.E., Hogrefe C.F., Lalayeva N., Oneda S. Object discrimination and reversal learning in infant and non-human primates in a non-clinical laboratory. Journal of medical primatology. 2013. 42 (3): 147–157.
Nagahara A.H., Bernot T., Tuszynski M.H. Age-related cognitive deficits in rhesus monkeys mirror human deficits on an automated test battery. Neurobiology of aging. 2010. 31 (6): 1020–1031.
Natale F., Antinucci F., Spinozzi G., Poti P. Stage 6 object concept in nonhuman primate’s cognition: a comparison between gorilla (Gorilla gorilla gorilla) and japanese macaque (Macaca fuscata). Journal of Comparative Psychology. 1986. 100 (4): 335–339.
National Research Council. Guide for the care and use of laboratory animals. National Academies Press. 2010.
Noser R., Byrne R.W. Wild chacma baboons (Papio ursinus) remember single foraging episodes. Animal. Cognition. 2015. 18 (4): 921–929.
Osorio D., Vorobyev M. A review of the evolution of animal colour vision and visual communication signals. Vision research. 2008. 48 (20): 2042–2051.
Paton J., Belova M.A., Morrison S.E., Salzman C.D. The primate amygdala represents the positive and negative value of visual stimulus during learning. Nature. 2006. 439 (7078): 865–870.
Pessoa D.M., Araujo M.F., Tomaz C., Pessoa V.F. Colour discrimination learning in black-handed tamarin (Saguinus midas niger). Primates. 2003. 44 (4): 413–418.
Pessoa D.M., Cunha J.F., Tomaz C., Pessoa V.F. Colour discrimination in the black-tufted-ear marmoset (Callithrix penicillata): ecological implications. Folia Primatologica. 2005. 76 (3): 125–134.
Poletiek F.H., Fitz H., Bocanegra B.B. What baboon can (not) tell us about natural language grammars. Cognition. 2016. 151: 108–112.
Rahman A., Languille S., Lamberty Y., Babiloni Cl., Perret M., Bordet P., Blin O., Jacob T., Auffret A., Schenker E., Richardson J., Pifferi F., Aujard F. Sleep deprivation impairs spatial retrieval but not spatial learning in the nonhuman primates grey mouse lemur. PLOS ONE. 2013. 8 (5): e64493.
Rowe M.H. Trichromatic color vision in primates. Physiology. 2002. 17 (3): 93–98.
Rumbaugh D.M., Pate J.L. 13 Primates’ Learning By Levels. Behavioral Evolution and Integrative Levels: The Tc Schneirla Conferences Series. Psychology Press. 2014. 1: 221.
Sandin M., Jasmin S., Levere T.E. Aging and cognition: Facilitation of recent memory in aged nonhuman primates by nimodipine. Neurobiology of Aging. 1990. 11 (5): 73–575.
Savage A., Dronzek L.A., Snowdon C.T. Color discrimination by the cotton-top tamarin (Saguinus oedipus oedipus) and its relation to fruit coloration. Folia Primatologica. 1987. 49 (2): 57–69.
Schmidt V., Fisher J. Inferential reasoning and modality dependent discrimination learning in olive baboons (Papio hamadryas anubis). Journal of Comparative Psychology. 2009. 123 (3): 316–325.
Schmidt V., Pankau B., Fisher J. Old World monkeys compare to apes in the Primate Cognition Rest Battery. Plos One. 2012. 7 (4): e32024.
Schneider M.L., Larson J.A., Rypstat C.W., Resch L.M., Roberts A., Moore C.F. Moderate level prenatal alcohol exposure enhances acoustic startle magnitude and disrupts prepulse inhibition in adult rhesus monkeys. Alcoholism: Clinical and Experimental Research. 2013. 37 (10): 1729–1736.
Schreier A.L., Grove M. Ranging patterns of hamadryas baboons: random walk analyses. Animal Behaviour. 2010. 80 (1): 75–87.
Seed A., Tomasello M. Primate cognition. Topics in Cognitive Science. 2010. 2: 407–419.
Smith A.C., Buchanan-Smith H.M., Surridge A.K., Osorio D., Mundy N.I. The effect of colour vision status on the detection and selection of fruits by tamarins (Saguinus spp.). Journal of Experimental Biology. 2003. 206 (18): 3159–3165.
Sigg H., Stolba A. Home range and daily march in a hamadryas baboon troop. Folia Primatologica. 1981. 36 (1–2): 40–75.
Van Schaik C.P., Pradhan G.R. A model for tool – use traditions in primates: implications for the coevolution of culture and cognition. Journal of Human Evolution. 2003. 44 (6): 645–664.
Vick S.J., Anderson J.R. Use of human visual attention cues by olive baboons (Papio anubis) in a competitive task. Journal of Comparative Psychology. 2003. 117 (2): 209.
Vick S.J., Bovet D., Anderson J.R. Gaze discrimination learning in olive baboons (Papio anubis). Animal Cognition. 2001. 4 (1): 1–10.
Wilson B., Marslen-Wilson W.D., Petkov C.I. Conserved sequence processing in primate frontal cortex. Trends in neurosciences. 2017. 40 (2): 72–82.
Zhou L., Wang Z., Liu T., Lu F., Wang Sh., Li J., Peng Sh., Zuo Zh. Neonatal exposure to sevoflurane may not cause learning and memory deficits and behavioural abnormality in the childhood of Cynomolgus monkeys. Scientific Reports. 2015. 5: 111–145.
Zurcher N., Rodriguez J.S., Jenkins S.L., Keenach K., Bartlett Th.Q., McDonald Th.J., Nathanielsz P.W., Nijland M.J. Perfomance of juvenile baboons on neuropsychological tests assessing associative learning motivation and attention. Journal of neuroscience methods. 2010. 188 (2): 219–225.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова