

Журнал высшей нервной деятельности им. И.П. Павлова, 2022, T. 72, № 5, стр. 651-665
Использование транскраниальной магнитной стимуляции в исследованиях когнитивного диссонанса при принятии решений
А. Г. Давыдова 1, *, Ю. П. Шеронова 1, В. В. Косоногов 1, А. Н. Шестакова 1, В. А. Ключарев 1
1 Институт Когнитивных Нейронаук, НИУ Высшая школа экономики
Москва, Россия
* E-mail: agdavydova@edu.hse.ru
Поступила в редакцию 15.03.2022
После доработки 08.05.2022
Принята к публикации 27.06.2022
- EDN: MPDJAR
- DOI: 10.31857/S0044467722050057
Аннотация
Теория когнитивного диссонанса (КД) тесно связана с исследованием процесса принятия сложных решений. В частности, КД проявляется в рамках “парадигмы свободного выбора” в переоценке альтернатив в результате выбора между двумя схожими альтернативами как следствие мотивации уменьшить внутренний конфликт. Механизмы КД связывают с активностью задней медиальной и дорсолатеральной префронтальной коры, прилежащего ядра и задней поясной коры, взаимосвязь которых по-разному интерпретируют по результатам нейровизуализационных и стимуляционных исследований. Одним из ключевых и до сих пор не решенных вопросов понимания механизмов КД является нейрохронометрия его возникновения, раскрытие которой возможно с помощью методов транскраниальной магнитной и электрической стимуляции (ТМС и ТЭС). Раскрытие временной последовательности нейрональных механизмов КД позволит не только прояснить фундаментальную природу принятия сложных решений, но и эффективнее манипулировать данными механизмами в различных прикладных задачах. Однако ТМС и ТЭС протоколы влияния на КД различаются такими параметрами, как мишень, момент, частота, сила стимуляции, контрольные условия, что не позволяет напрямую сравнивать результаты исследований. В данном обзоре мы приводим результаты анализа стимуляционных исследований КД с целью выявить временные закономерности влияния стимуляции на изменение предпочтений в результате КД.
ВВЕДЕНИЕ
Психологи давно заметили, что выбор человека не всегда согласован с его поступками. Более того, было также показано, что поступки человека могут влиять на его последующий выбор. Это и было положено в основу теории когнитивного диссонанса (КД). В наши дни она имеет как фундаментальное, так и прикладное значение в изучении природы сложных решений в норме и при патологиях. Леон Фестингер (Festinger, 1957) предположил, что конфликт между “когнициями”, включая конфликт при сложном выборе, вызывает внутренний дискомфорт, который ведет к возможным изменениям в поведении, предпочтениях или аттитюдах с целью снизить данное аверсивное состояние. Теория объясняет ряд феноменов, включая процессы рационализации, поведение людей при различных видах зависимостей и другие, поэтому понимание природы КД чрезвычайно актуально. Российские авторы значительно развили понимание психологических основ выбора, в частности были сформулированы ключевая идея о большей важности того, как субъект выбирает, чем того, что он выбирает, и положение о субъективном конструировании выбора (Леонтьев и др., 2015). Важна роль Ю. Козелецкого (Козелецкий, 1979), выделившего стадии принятия решений. Примечательна гипотеза обратимости Т. Корниловой (Корнилова, 2003), согласно которой выбор совершается, пока альтернативы субъективно обратимы для человека, принимающего решение. Иными словами, осознание необратимости альтернативы означает ее окончательное принятие или отказ от принятия.
Однако нейрофизиологические основы данного феномена остаются не до конца раскрытыми. В целом, данные когнитивной и социальной нейронауки свидетельствуют о том, что активность задней медиальной префронтальной коры (medial prefrontal cortex, МПФК), дорсолатеральной префронтальной коры (dorsolateral prefrontal cortex, ДЛПФК), прилежащего ядра и задней поясной коры (posterior cingulate cortex, ЗПК) лежит в основе изменения предпочтений в результате КД, возникшего в процессе принятия решений (Di Martino et al., 2008; Izuma et al., 2010; Izuma et al., 2015). Однако в предыдущих исследованиях не был предложен однозначный нейрокогнитивный механизм (модель) КД.
Многие нейровизуализационные исследования свидетельствуют о ключевой роли прилежащего ядра, через которое взаимосвязаны задняя МПФК, ДЛПФК и ЗПК (Di Martino et al., 2008). В наших предыдущих работах, включая электроэнцефалографическое исследование КД (Colosio et al., 2017), было показано, что в рамках парадигмы свободного выбора сложные решения приводят к КД, который активирует МПФК в соответствии с механизмом генерации ошибки предсказания при обучении с подкреплением. На основании этого и других исследований (Colosio et al., 2018) мы предположили, что изменение предпочтений в ходе обработки внутреннего конфликта может происходить в соответствии со следующим механизмом: (а) задняя МПФК идентифицирует внутренний конфликт во время сложного выбора; (б) ДЛПФК реализует когнитивный контроль в целях изменения предпочтений и редуцирования КД; (в) прилежащее ядро кодирует предпочтения и запускает обучающий сигнал, который является ключевым для модификации предпочтений и обоснования выбора; (г) ЗПК обеспечивает закрепление измененных предпочтений на длительный срок. В целях разработки и уточнения предложенной модели изменения предпочтений дальнейшим исследователям КД следует учитывать необходимость интеграции различных методов нейровизуализации.
Методы стимуляции позволяют раскрыть нейрохронометрию механизма возникновения КД в процессе принятия решений – понять время и очередность развития нейрофизиологических коррелятов возникновения внутреннего конфликта и последующих изменений в поведении и предпочтениях.
Целью данного обзора является выявление возможностей установления каузальной связи КД и его нейрональных коррелятов, особенностей временной динамики данных коррелят, а также возможностей влияния на механизмы возникновения и последующего снижения КД с помощью методов транскраниальной стимуляции мозга. В этой связи мы сфокусировались на нескольких задачах обзора:
1) проанализировать, насколько существующие парадигмы исследования КД позволяют в экспериментальных условиях оценить психофизиологические механизмы КД и количественно измерить чистый эффект КД (см. раздел 2. Теоретические основы исследований внутреннего конфликта при принятии решений);
2) оценить результаты нейровизуализационных и стимуляционных исследований КД с точки зрения понимания того, какую функцию выполняют различные зоны мозга в процессе возникновения и снижения внутреннего конфликта (см. раздел 3. Области мозга, вовлеченные в процесс обработки внутреннего конфликта);
3) сравнить эффективность использования различных параметров протоколов стимуляции с целью повлиять на процесс развития КД (см. раздел 4. Анализ протоколов стимуляционных исследований внутреннего конфликта);
4) оценить, в какой момент парадигмы свободного выбора исследователям удалось эффективно повлиять на нейрофизиологические корреляты КД (см. раздел 5. Результаты влияния на эффект когнитивного диссонанса при применении различных протоколов стимуляции).
В целом подобный подход позволит прояснить нейрохронометрию КД.
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ ИССЛЕДОВАНИЙ ВНУТРЕННЕГО КОНФЛИКТА ПРИ ПРИНЯТИИ РЕШЕНИЙ
Теория когнитивного диссонанса
Теория КД, созданная социальным психологом Леоном Фестингером (Festinger, 1957), объясняет последствия совершения сложного выбора, вызывающего внутренний дискомфорт. Человек стремится редуцировать или нивелировать несоответствие между личностно значимыми установками (определенное автором как диссонанс) и, соответственно, снизить или полностью избавиться от внутреннего дискомфорта. Для этого он может привести в соответствие диссонирующие взгляды или достичь консонанса, прежде всего, с помощью оправдания собственного выбора и убеждения себя в его правильности. Описание психологических механизмов возникновения внутреннего конфликта и связанного с ним негативного эмоционального состояния потребовало объяснения нейробиологического механизма такой рационализации – изменения привлекательности альтернатив вследствие сложного выбора (снижения значимости отвергнутой опции и усиления привлекательности выбранной). Прояснение нейробиологических механизмов КД требует как релевантной поведенческой парадигмы, так и соответствующего нейрофизиологического метода. В данном разделе статьи приведён анализ парадигмы свободного выбора и ее версий.
Модифицированная парадигма свободного выбора
Парадигма свободного выбора была разработана Дж.В. Бремом (Brehm, 1956), который предлагал участникам выбирать между равно привлекательными для них альтернативами. Согласно результатам исследования, принятие решений в пользу одной из альтернатив приводило к последующей девальвации невыбранной опции и увеличению привлекательности выбранной – к так называемому расхождению альтернатив. Такие изменения предпочтений обнаруживались благодаря следующему этапу парадигмы – повторной оценке тех же альтернатив, которые были предъявлены на стадии выбора. Исследования с использованием парадигмы свободного выбора неоднократно демонстрировали влияние выбора (рационализации) на переоценку альтернатив.
Однако сравнительно недавно К. Чен и Дж. Ризен (Chen, Risen, 2010) подвергли критике традиционную “парадигму свободного выбора”, разработанную Дж.В. Бремом. Основанием критики послужило выявление артефакта, который возникает при повторной оценке опций. Исследователи показали, что повторная оценка находится под влиянием “скрытых” предпочтений участников, которые невозможно абсолютно точно оценить в ходе предварительной оценки опций. В целях более точного измерения расхождения альтернатив, обусловленного исключительно разрешением внутреннего конфликта, авторы предложили модифицированную парадигму свободного выбора (rate-rate-choose paradigm). В рамках данной парадигмы сначала участникам предлагается (1) оценить объекты (опции – товары, изображения или другие объекты) с целью выяснить предпочтения (сессия “Первая оценка”), (2) затем в специально подобранных парах выбрать наиболее предпочтительный объект (сессия “Выбор”), (3) после чего повторно оценить привлекательность объекта (сессия “Повторная оценка”). При этом пары стимулов для второй сессии (“Выбор”) формируются, исходя из разницы в оценках двух объектов. Выбор между парой высоко предпочитаемых объектов, оцененных равным или почти равным образом, представляет собой “сложный выбор”. Согласно теории КД, “сложный выбор” должен вызвать максимальный внутренний конфликт, разрешение которого приводит к расхождению альтернатив. Между тем выбор между объектами с сильно различающимися оценками представляет собой простой выбор, не вызывающий КД. В контрольном условии предлагается вначале также оценить объекты (“Первая оценка”), затем участники повторно оценивают первоначально предъявленные стимулы (“Повторная оценка”) и лишь после этого совершают выбор между парами стимулов (“Поствыбор”), привлекательность которых была оценена ранее два раза подряд. Соответственно, сравнение степени расхождения альтернатив в контрольном и экспериментальном условиях демонстрирует влияние артефакта повторного измерения. Исследования в рамках модифицированной парадигмы свободного выбора показали большее расхождение альтернатив в экспериментальном условии по сравнению с контрольным. Это подтверждает наличие механизма разрешения внутреннего конфликта, который приводит к изменению предпочтений (Chen, Risen, 2010; Izuma et al., 2010; Sharot et al., 2012; Colosio et al., 2017). В нашем исследовании (Colosio et al., 2017), реализованном в рамках модифицированной парадигмы свободного выбора, участникам было необходимо оценить изображения продуктов питания по 8-балльной шкале, затем выбрать между двумя изображениями продуктов питания, предъявленными на первом этапе (при этом в части проб выбор совершал компьютер, и участнику было необходимо подтвердить выбор). Затем участники вновь оценивали сходный набор изображений, а на последнем этапе “Поствыбора” им предлагалось выбрать между изображениями, выбор между которыми на первом этапе был совершeн компьютером. Во время первой сессии выбора изображения были поделены на пары с минимальной разницей в оценке, выбор между которыми вызывает сильный КД, и на те, выбор между которыми в силу большой разницы в оценке не вызывает или вызывает минимальный внутренний конфликт. Пары, выбор между которыми был сделан компьютером, относились к категории “сложного выбора”, соответственно, во время этапа “Поствыбора” участники испытывали сильный КД (см. рис. 1).
Рис. 1.
I. Во время задания “Оценка I” участники оценивали продукты, представленные на экране в течение 3 сек. II. Далее, во время задания “Выбор” в пробах с “самостоятельным выбором” испытуемые свободно выбирали один из двух продуктов (пробы со “сложным” выбором вызывали сильный КД, пробы с “простым” выбором – слабый). В пробах с “выбором компьютера” испытуемые должны были выбрать тот продукт, который был выбран вычислительным алгоритмом (выделен жирной рамкой). III. В задании “Оценка II” участники снова оценивали те же продукты. Участникам напоминали об их выборе (если он был) во время задания “Выбор” (например, “Вы не выбрали эту опцию”). IV. Во время контрольного задания “Поствыбор” участники выбирали продукты из пар, которые были представлены в предыдущих пробах в качестве проб с “выбором компьютера”. Fig. 1. I. During the first rating task participants evaluated products presented during 3 seconds on the screen. II. Further, during the choice task subjects chose one of two products in self-trials (self-difficult trials induced the strong cognitive dissonance, while the self-easy trials evoked the weak one). In computer trials subjects had to choose the item chosen by the computational algorithm (highlighted by the bold frame). III. In the second rating task participants evaluated the same items again. Reminders about participants’ choice during the choice session were presented on the screen (e.g., “You did not chose this option”). IV. During the control Post-ex Choice task participants chose products from pairs, which were presented in the previous computer trials.
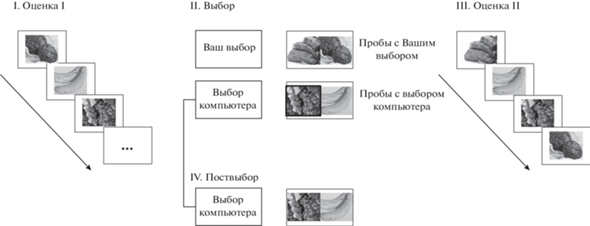
Измененная парадигма широко используется при изучении изменения предпочтений в ходе сложного выбора (Izuma et al., 2010; Sharot et al., 2012; Mengarelli et al., 2015; Colosio et al., 2017).
Парадигма слепого выбора
Интерпретация результатов предыдущих исследований КД, по мнению многих авторов (Izuma et al., 2015), осложняется наличием артефакта, на наличие которого указывают К. Чен и Дж. Ризен (Chen, Risen, 2010). Исследователи показали, что повторная оценка находится под влиянием “скрытых” предпочтений участников, которые невозможно абсолютно точно оценить в ходе предварительной оценки опций. Поэтому ученые предложили альтернативную парадигму – парадигму слепого выбора (Egan et al., 2010). Основное отличие парадигмы слепого выбора от парадигмы свободного выбора состоит в том, что выбор участника либо подменяется экспериментатором (см. подробнее далее), либо выбор участника случайный в силу одинаковости предъявляемых стимулов или кратковременности их предъявления, что не позволяет ни воспринять, ни запомнить сделанный выбор. Таким образом, участник выбирает “вслепую”, то есть либо не осознавая свой выбор, либо ориентируясь на чужой выбор. При этом из-за особенностей экспериментальной парадигмы участник не понимает, что на самом деле его выбор был случайным или выбранные опции подменили. Так, в исследовании КД у детей и обезьян (Egan et al., 2010) участники экспериментов, не зная о сходстве стимулов, сначала совершали выбор (выбор 1) между двумя равно предпочитаемыми объектами, после чего выбирали (выбор 2) между отвергнутой опцией и третьим объектом, также первоначально предпочитаемым на одном уровне с первыми двумя объектами. Контрольная группа осуществляла только выбор 2 в отсутствии выбора 1. В результате в экспериментальной группе наблюдалось обесценивание отвергнутой опции (в ходе второй сессии выбора дети и обезьяны значимо предпочитали третий объект), тогда как в контрольной группе результаты выбора были случайными. Данную парадигму КД использовали также в исследованиях КД у взрослых. Например, Т. Шерот и соавт. (Sharot et al., 2010) предлагали участникам выбрать между стимулами, представляющими собой названия курортных мест. Стимулы предъявлялись в течение 2 мс, и в реальности вместо названий курортов участникам показывали бессмысленный набор букв (участники не имели возможности воспринять предъявляемые стимулы в силу очень короткого периода предъявления). В экспериментальной группе наблюдалось обесценивание отвергнутой опции, в отличие от контрольной, где первый выбор совершал компьютер, что отражает наличие чистого влияния сложного выбора на расхождение альтернатив.
Популярным вариантом парадигмы слепого выбора является незаметная подмена выбора участника. В этом случае участник уверен в том, что его выбор был осознанным и напрямую зависит от его предпочтений (Johansson et al., 2012). Данный подход использовался в исследовании роли задней МПФК в расхождении альтернатив в результате принятия сложного решения в группе К. Изумы (Izuma et al., 2015). В данном исследовании в качестве стимулов были выбраны 50 абстрактных изображений. Они были заранее поделены на 25 пар в соответствии со следующими критериями: 1) изображения должны быть достаточно схожими, чтобы участник эксперимента не заметил подмену выбора после отвлекающего задания; 2) изображения должны в достаточной степени отличаться, чтобы при сравнении пары стимулов участник эксперимента осознанно сделал выбор в пользу одного из изображений, а не совершил случайный выбор в силу отсутствия различия в предпочтениях. Как и в традиционной парадигме, на первом этапе испытуемые оценивали привлекательность изображений. После первой оценки стимулов пары изображений были автоматически распределены на 4 группы: 1) пробы, в которых выбор участника не подменялся (5 пар), 2) пробы, в которых выбор участника подменялся (5 пар); 3) пробы, в которых выбор совершал компьютер (5 пар); 4) прочие контрольные пробы (10 пар). В соответствии с алгоритмом распределения стимулов пары с большей разницей в первоначальной оценке преимущественно были отнесены к первым 3 группам. Далее, во время сессии выбора участник выбирал наиболее привлекательное изображение, а затем выполнял отвлекающую задачу. После этого на экране появлялось истинное изображение, которое было выбрано участником или “подмененное” изображение, которое не было выбрано участником, причем расположение выбранного стимула было изменено так, чтобы заставить участника поверить, что на экране представлен реально выбранный им объект (см. более подробно рис. 2).
Рис. 2.
Иллюстрация трех типов проб в ходе сессии выбора: 1) подмена выбора, 2) выбор без подмены, 3) выбор компьютера. Во время сессии выбора участник сначала выбирал более привлекательное изображение, затем выполнял отвлекающую фланговую задачу, после чего на экране оставалось изображение, которое было либо выбрано участником (1 группа), либо не было выбрано участником, но расположение выбранного стимула было изменено, чтобы заставить участника поверить, что на экране представлен реально выбранный им объект (2 группа), либо выбор был совершен компьютером (3 группа). Во всех типах проб выбор необходимо было подтвердить. Fig. 2. Illustration of three trial types during the choice session: 1) choice switch, 2) choice without switch, 3) computer choice. In the choice task a participant firstly chooses the more attractive picture, then he executes a distracting Flanker task. Afterwards, either a chosen picture (first group), or an unpreferred image, the position of which was substituted to make a participant believe that he indeed chose the shown picture (second group), or the computer made the choice (third group), was shown on the screen. In all trial types participants have to confirm their choice.

Таким образом, необходимо отметить несколько важных характеристик механизма развития КД при принятии решений. Во-первых, возникновение диссонирующих установок сопряжено с появлением психологического дискомфорта, который и порождает дальнейшее стремление привести противоречивые установки в соответствие друг другу. Во-вторых, несмотря на то, что теория КД предполагает необходимость личностной значимости конфликта, экспериментально внутренний конфликт при принятии решений регулярно наблюдался в ситуациях, не представляющих личностной значимости для участника. То есть сам факт выбора между любыми равно предпочитаемыми опциями вызывает стремление к “рационализации” совершенного выбора, что подразумевает возникновение конфликта на неосознаваемом уровне. В-третьих, существует несколько механизмов снижения или избегания внутреннего конфликта, основным из которых является приведение своих взглядов или предпочтений в соответствие с принятым решением, что оказалось возможно количественно измерить с помощью модификации парадигмы свободного выбора.
ОБЛАСТИ МОЗГА, ВОВЛЕЧЕННЫЕ В ПРОЦЕСС ОБРАБОТКИ ВНУТРЕННЕГО КОНФЛИКТА
Медиальная префронтальная кора (МПФК)
Согласно многим исследованиям, МПФК активируется при задачах, которые вызывают конфликт, а именно при классической задаче Струпа и фланговой задаче (Botvinick et al., 2001). Также активность МПФК связывают с мониторингом деятельности и возникновением негативной обратной связи в связи с обнаружением ошибки (Colosio et al., 2018). Одна из ключевых моделей КД – модель ориентации на действие (“action-based model”) – предполагает, что роль задней МПФК состоит в выявлении внутренних конфликтов и уменьшении влияния КД (Harmon-Jones et al., 2009). Кроме того, задняя МПФК вовлечена в процесс генерации ошибки предсказания при обучении с подкреплением, когда результат действия отличается от ожидаемого (Colosio et al., 2018). В исследовании нашей группы мы также показали, что изменение предпочтений в результате КД зависит от динамики нейронов задней МПФК в состоянии покоя (Colosio et al., 2017).
Дорсолатеральная префронтальная кора
По данным исследований в области когнитивной и социальной нейронаук, ДЛПФК играет важную роль в процессе изменения предпочтений в результате сложного выбора, однако в научном сообществе еще не достигнут консенсус относительно того, на какой именно этап изменения предпочтений влияет данная область мозга.
Существуют предположения, что ДЛПФК отвечает за реализацию когнитивного контроля во время принятия решений (Miller, Cohen, 2001). Однако также исследуется роль ДЛПФК в процессе рационализации выбора в состоянии КД (Mengarelli et al., 2015; Qin et al., 2011; Voigt et al., 2019). Так, согласно исследованиям, мозговая активность во фронтальных областях головного мозга может предсказывать индивидуальные различия в степени расхождения альтернатив в результате сложного выбора (Qin et al., 2011). Примечательно также исследование 2008 года под авторством Э. Хармона-Джонса (Harmon-Jones et al., 2008), в котором ему удалось значительно уменьшить изменение предпочтений в ходе сложного выбора, подавляя активность левой ДЛПФК с помощью тренировок с использованием ЭЭГ и биологической обратной связи. Подобный же эффект был достигнут в эксперименте Ф. Менгарелли (Mengarelli et al., 2015), в котором с помощью катодной ТЭС постоянным током левой ДЛПФК получилось уменьшить эффект расхождения альтернатив. При этом аналогичная стимуляция правой ДЛПФК подобного эффекта не дает. Кроме того, исследование с использованием функциональной магнитно-резонансной томографии (фМРТ) показало (Voigt et al., 2020), что активация левой ДЛПФК позволяет предсказать появление последующего эффекта изменения предпочтений.
Тем не менее, не стоит также забывать о другом исследовании Э. Хармона-Джонса (Harmon-Jones et al., 2011), в котором он предполагает, что ДЛПФК не связана с КД напрямую, а ее активность может повышаться независимо от наличия КД, чтобы обеспечить приверженность выбранному плану действий. Также есть предположения, что ДЛПФК является частью более общих механизмов когнитивного контроля (Izuma et al., 2015).
Прилежащее ядро
Прилежащее ядро является важной частью системы вознаграждения и связано непосредственно с ожиданием награды. Кроме того, прилежащее ядро играет ключевую роль при выборе модели поведения в силу того, что интегрирует когнитивную и аффективную информацию, которая обрабатывается в области фронтальной и височной коры головного мозга (Floresco, 2015).
В исследованиях расхождения альтернатив прилежащее ядро также является одной из областей, которая возбуждается во время принятия сложных решений (Izuma et al., 2010; Kitayama et al., 2013). При этом важно заметить, что активность прилежащего ядра не только коррелирует с изменением предпочтений в результате сложного выбора, но и позволяет предсказать расхождение альтернатив. Более того, изменение активности прилежащего ядра для отвергнутых опций меньше, чем для выбранных опций.
Принимая во внимание связь активности прилежащего ядра с ожиданием награды (Knutson et al., 2009), стоит упомянуть, как может быть связано ожидание вознаграждения и совершение выбора при отсутствии очевидной награды. Так, например, Ш. Китаяма в своем исследовании (Kitayama et al., 2013) предполагает, что изменение предпочтений возникает, когда во время принятия решения участник ищет положительные характеристики среди опций, между которыми выбирает, идентифицирует их и успешно связывает с определенными аспектами своей личности. Также, по мнению Ш. Китаямы, успешный поиск характеристик, которые одновременно и уникальны для одной из опций, и релевантны именно для данной личности, может продуцировать стойкое ожидание награды, которая, вероятно, возникнет как результат данного выбора. В том же исследовании Ш. Китаяма обнаруживает положительную корреляцию между усилением предпочтений для выбранных опций и активностью прилежащего ядра.
Несмотря на проведенные исследования, все еще не ясно, на каком именно этапе изменения предпочтений задействовано прилежащее ядро. Чтобы найти ответ на этот вопрос, необходимо множество дополнительных исследований.
Задняя поясная кора
ЗПК связана со множеством когнитивных функций. Как и ДЛПФК, она задействована в процессе когнитивного контроля (Leech et al., 2011). Кроме того, в ряде экспериментов было показано, что ЗПК участвует в кодировании эмоциональных воспоминаний (Maddock et al., 2001), а также извлечении информации из памяти и планировании (Vann et al., 2009). Ряд других исследований также показал, что ЗПК задействована в детекции изменений в окружающей среде и мотивировании последующих изменений в поведении (Pearson et al., 2011), а также в контроле баланса между внешним и внутренним вниманием (Leech, Sharp, 2014).
В исследованиях расхождения альтернатив выдвигаются предположения, что ЗПК может играть центральную роль в регулировании предпочтений. Обнаружена позитивная корреляция между активностью ЗПК и восприятием желанности объектов (Kawabata, Zeki, 2008). Исследования с использованием фМРТ фиксируют значительную активность ЗПК как в процессе принятия сложных решений (Kitayama et al., 2013; Qin et al., 2011; Tompson et al., 2016), так и после сложного выбора (Izuma et al., 2010).
АНАЛИЗ ПРОТОКОЛОВ СТИМУЛЯЦИОННЫХ ИССЛЕДОВАНИЙ ВНУТРЕННЕГО КОНФЛИКТА
Для раскрытия нейрохронометрии развития внутреннего конфликта требуется соответствующий нейрофизиологический инструмент, такой как транскраниальная стимуляция. В рамках обзора использования методов транскраниальной стимуляции мозга для изучения механизмов внутреннего конфликта в процессе принятия решений мы сфокусируемся на протоколах транскраниальной магнитной стимуляции (ТМС) и транскраниальной электрической стимуляции (ТЭС).
ТМС – это метод неинвазивной стимуляции мозга, особенность которого состоит в том, что стимуляционная катушка, расположенная на голове испытуемого, является генератором переменного магнитного поля (до 2 Тесла), которое создает электрический ток, способный деполяризовать или гиперполяризовать нейроны в стимулируемой зоне мозга. ТЭС также является методом неинвазивной транскраниальной стимуляции, который предполагает слабое электрическое воздействие (0.5–2 мА) постоянным (ТСПоТ, tDCS) или переменным (ТСПеТ, tACS) током через 2 электрода, один из которых является стимулирующим, а второй – референтным (при биполярном монтаже размещается над нетаргетируемой зоной мозга, при униполярном монтаже – не на голове).
Далее, мы оценим протоколы ТМС и ТЭС, а именно такие характеристики, как время, частота и сила стимуляции, контрольные условия, зоны мозга – мишени стимуляции, а также проанализируем механизм развития когнитивного конфликта, на который было осуществлено воздействие. Исходя из особенностей методологии нейрохронометрических исследований, рекомендуется применять ТМС как более фокальный метод с точки зрения пространственных и временных характеристик.
Для определения последовательности нейрофизиологических процессов при развитии внутреннего конфликта методами стимуляции мозга требуется определить (i) стимулируемую зону мозга, (ii) момент стимуляции относительно предъявления стимулов, (iii) контрольные условия, (iv) мишень — точку стимуляции, (v) силу стимуляции, (vi) частоту и длительность стимуляции.
Стимулируемая зона мозга – мишень стимуляции
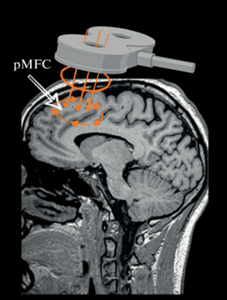
Целый ряд областей мозга связан с различными компонентами развития внутреннего конфликта (см. Colosio et al., 2017, 2018): задняя поясная извилина, прилежащее ядро и передняя островковая зона (расположенные не на поверхности головного мозга), а также задняя МПФК и ДЛПФК. Глубокие области мозга недоступны для ТЭС и ТМС, поэтому основной мишенью стимуляционных исследований внутреннего конфликта являются ЗПК и ДЛПФК (см. рис. 3). Стимуляция данных областей влияет на такие механизмы, как мониторинг результатов деятельности, детекция ошибок, а также общий когнитивный контроль и нивелирование когнитивных конфликтов (Colosio et al., 2018).
Рис. 3.
Мишень стимуляции. MNI координаты точки стимуляции в МПФК (x = 10, y = 48, z = 70). При использовании ТМС через стимуляционную катушку пропускается короткий переменный ток высокого напряжения. При этом в плоскости, перпендикулярной поверхности катушки, возникает переменное магнитное поле (до 2 Тесла). Для стимуляции применяется катушка “восьмерка”, так как данный вид катушки обеспечивает стимуляцию более глубоко расположенных зон головного мозга. Fig. 3. TMS target area. MNI coordinates of the pMFC stimulation point (x = 10, y = 48, z = 70). When using TMS, the short high voltage alternating current is passed through the stimulation coil. At the same time, the changing magnetic field is generated in the plane perpendicular to the coil surface (up to 2 Tesla). The Figure-8 TMS coil is applied since this coil type provides the stimulation of the deeper brain areas.
Момент стимуляции относительно предъявления стимулов
В силу того, что нет однозначного понимания длительности воздействия ТМС и в особенности ТЭС на активность мозга при тех или иных параметрах длительности, частоты и силы стимуляции, стимуляционные исследования не могут однозначно подтвердить точную нейрохронометрию развития КД. Большинство протоколов стимуляции, применяемых при изучении КД, предполагают стимуляцию сразу после стадии выбора (см. рис. 1, т.е. сразу же после стадии II. Выбор) и перед началом повторной оценки (Izuma et al., 2015; Mengarelli et al., 2015). Тем не менее, в некоторых поведенческих, нейровизуализационных и стимуляционных исследованиях авторы исследуют развитие КД в момент принятия решений (Kitayama et al., 2013; Colosio et al., 2018; Voigt et al., 2019; Lee, Daunizeau, 2020). Однако 20-минутная ТЭС на уровне 1.5 мА, предваряющая стадию выбора, может сохранить воздействие также на стадии повторной оценки, так как в некоторых исследованиях эффект модуляции возбудимости МПФК был зафиксирован через несколько часов после стимуляции (Nitsche, Paulus, 2001; Nitsche et al., 2003; Reinhart, Woodman, 2014). Кроме того, стимуляция может повлиять не на процесс детекции и разрешения внутреннего конфликта, а на другие когнитивные процессы, например, на улучшение восприятия собственных предпочтений. Так, согласно некоторым исследованиям, задняя МПФК вовлечена в определение личностной значимости происходящего (Johnson et al., 2002; Northoff et al., 2006). До настоящего момента, по нашим сведениям, ни одно из исследований не фокусировалось на стимуляции в разные моменты парадигмы свободного выбора для прояснения нейрохронометрии процесса. Таким образом, в дальнейших исследованиях требуется провести ТМС в течение разных стадий парадигмы свободного выбора. Ниже мы рассмотрим важные детали, необходимые для будущих нейрохронометрических исследований.
Контрольные условия
Одним из основных параметров оффлайн ТМС протокола, который интересует исследователей с точки зрения решения той или иной исследовательской задачи, является применение различных контрольных условий (плацебо-стимуляции или стимуляции нерелевантной области мозга). Хотя ряд исследований включал только одно контрольное условие (например, только плацебо-стимуляция (Chawke, Kanai, 2016; Holbrook et al., 2016) или только стимуляция контрольной зоны (Holbrook et al., 2020, 2021)), наиболее контролируемые исследования использовали несколько контрольных условий (Klucharev et al., 2011; Izuma et al., 2015).
В связи с тем, что испытуемые могут осознавать тип стимуляции, например, по отсутствию характерных ощущений и звуков, исследователи использовали несколько вариантов плацебо-стимуляции для изучения роли задней МПФК и ДЛПФК в изменении личных предпочтений и взглядов. К. Изума предложил стимулировать участников под углом 90° по отношению к скальпу так, чтобы стимуляция не достигала коры головного мозга (Izuma et al., 2015). В. Ключарев и К. Холбрук снизили силу стимуляции до 10% от максимального значения стимулятора по сравнению со значением экспериментальной стимуляции, которое в среднем составляло 25% от максимума и 80% от активного моторного порога (Klucharev et al., 2011; Holbrook et al., 2016). Третьим вариантом реализации плацебо-стимуляции является минимизация времени стимуляции по сравнению со стимуляцией в экспериментальной группе. В исследовании Ф. Менгарелли и соавт. (Mengarelli et al., 2015) плацебо-стимуляция длилась 15 секунд по сравнению с 15-минутной экспериментальной стимуляцией; в исследовании К. Чоке и Р. Канаи (Chawke, Kanai, 2016) стимуляция длилась 20 секунд, в то время как экспериментальная – 10 минут; в работе М. Колозио и соавт. (Colosio et al., 2018) длительность шам-стимуляции также была уменьшена с 20 минут экспериментальной стимуляции до 30 секунд. Критерии выбора контрольной зоны стимуляции по отношению к задней МПФК также не унифицированы среди исследователей. Это связано с необходимостью выбрать, с одной стороны, зону мозга, на которую не распространяется эффект стимуляции, а с другой стороны, зону, которая не вовлечена в процесс принятия решений и когнитивного контроля. К. Изума в контрольном условии использовал заднюю теменную кору в качестве контрольной области, так как таргетная зона расположена на расстоянии одной трети от назиона на линии между назионом и инионом, а контрольная зона – на расстоянии двух третей (Izuma et al., 2015). В. Ключарёв в рамках контрольного условия стимулировал зону предклинья (Klucharev et al., 2011), а К. Холбрук использовал зрительную зону MT или V5 в правой затылочной коре как контрольную область (Holbrook et al., 2020, 2021). Ф. Менгарелли, который сфокусировался на роли ДЛПФК, выбрал в качестве контрольной зоны стимуляции контралатеральную супраорбитальную область (Mengarelli et al., 2015). Таким образом, в дальнейших исследованиях требуется унификация контрольного условия.
Определение мишени точки стимуляции
От определения конкретной точки стимуляции зависит точность влияния на активность стимулируемой зоны. По этой причине требуется критический анализ наиболее распространенных методов определения мишени стимуляции. Большинство ключевых исследователей внутреннего конфликта не использовали структурных снимков МРТ каждого участника для определения мишени, в отличие, например, от исследователей моторной коры. Однако МРТ использовалась для адаптации координат, определенных на основе международной системы размещения электродов “10–20”. Так, Ф. Менгарелли использовал координаты зон F3 и F4 атласа Талайраха, определенных в соответствии с системой “10–20”, однако адаптировал их по индивидуальному МРТ (Mengarelli et al., 2015). Большинство же исследователей определяли мишень (координаты) стимуляции на основе системы “10–20” и использовали стандартизированную карту мозга в координатной системе Монреальского неврологического института (MNI) либо систему координат Талайраха. Выбирая расположение точки стимуляции, исследователи опирались на результаты предварительных ЭЭГ или фМРТ-исследований (Mengarelli et al., 2015; Chawke, Kanai, 2016; Colosio et al., 2018) или на стандартные краниометрические точки (Klucharev et al., 2011; Holbrook et al., 2020; Izuma et al., 2015). Безусловно, в дальнейших исследованиях для выбора мишени требуется использовать индивидуальные структурные МРТ и координаты, полученные в ходе фМРТ-исследований.
Уровень стимуляции
Стремясь предотвратить изменение предпочтений и взглядов участников через снижение активности областей коры, связанных с обнаружением и обработкой внутреннего конфликта, исследователи снижали возбудимость задней МПФК или ДЛПФК. Они также ориентировались на моторный порог, что является традиционным методом определения уровня стимуляции и в других областях исследования принятия решений методом ТМС (90% исследований, включенных в обзор Turi et al., 2021 с 1991 по 2020 г.). Однако на текущий момент не существует исследований, которые бы доказывали, что пороговое значение в префронтальной коре полностью коррелирует с моторным порогом. В большинстве исследований уровень стимуляции таргетируемой зоны в зависимости от моторного порога варьировал от 80 до 120% (в зависимости от типа стимуляции). При этом при высокочастотной стимуляции, для которой характерны интермиттирующие тета-последовательности стимуляции, наиболее часто применяется уровень стимуляции, составляющий 80% от моторного порога (Turi et al., 2021). В целом, для подавления активности МПФК и ДЛПФК с помощью как высокочастотной, так и низкочастотной стимуляции, ключевыми исследователями был выбран уровень стимуляции в 80% от порогового значения.
Частота и момент стимуляции
Предыдущие стимуляционные исследования КД можно условно разделить на две группы. В первой группе исследований была использована высокочастотная краткая стимуляция с интермиттирующими тета-последовательностями ТМС (серия стимуляции в 50 Гц длительностью 40 с, в течение которой было подано 600 импульсов, была использована в исследованиях Klucharev et al., 2011, Holbrook et al., 2016, 2020; предположительный эффект стимуляции действовал около часа). Во второй же группе исследований использовалась низкочастотная длительная стимуляция (например, К. Изума применял 25-минутную ритмическую стимуляцию частотой 1 Гц, эффект которой предположительно длился около 20 минут, см. Izuma et al., 2015). Безусловными преимуществами высокочастотной стимуляции являются краткость стимуляции как таковой и большая длительность последующего эффекта. Однако исследователи отмечают, что некоторым участникам доставляет дискомфорт высокочастотная стимуляция, что приводит к исключению из выборки ряда потенциальных участников на основе их низкого моторного порога.
РЕЗУЛЬТАТЫ ВЛИЯНИЯ НА ЭФФЕКТ КОГНИТИВНОГО ДИССОНАНСА ПРИ ПРИМЕНЕНИИ РАЗЛИЧНЫХ ПРОТОКОЛОВ СТИМУЛЯЦИИ
В исследовании К. Изумы (Izuma et al., 2015), реализованном в рамках парадигмы слепого выбора, был применен протокол влияния на заднюю МПФК с помощью низкочастотной 25-минутной ТМС. Воздействие с помощью данного протокола снизило степень расхождения альтернатив, а стандартизированный размер эффекта (d Коэна) составил 0.59. Применив альтернативный протокол высокочастотной ТМС в течение 40 секунд для ингибирования задней МПФК, В. Ключарёв и коллеги (Klucharev et al., 2011) снизили уровень социальной конформности. Однако исследователи полагают, что снижение социальной конформности может быть обусловлено редуцированием эффекта КД, вызванного конфликтом с социальными нормами. Сходный высокочастотный протокол использовали К. Холбрук и соавт. (Holbrook et al., 2016, 2020) в целях редуцирования эффекта КД. Примечательно, что они осуществляли воздействие не на степень изменения предпочтений и взглядов в результате разрешения внутреннего конфликта, а на степень усиления приверженности собственным взглядам. Данный эффект был вызван конфликтом из-за предъявления стимулов, представляющих угрозу для идеологических и религиозных взглядов участников. В первом исследовании авторы подавили активность задней МПФК и таким образом добились двух эффектов. Во-первых, исследователи воспрепятствовали усилению приверженности религиозным взглядам после праймингового опроса о восприятии участниками собственной смерти. Во-вторых, авторы сдержали увеличение приверженности собственной социальной группе (“in-group”) после прочтения эссе представителя другой социальной группы (“out-group”/”они”), осуждающего социальную группу участников. Во втором исследовании авторы усовершенствовали протокол, добавив ещё несколько контрольных условий, и не смогли реплицировать эффект КД в условии увеличения приверженности своей социальной группе, в отличие от первого условия. Данное отличие результатов экспериментов, по мнению авторов, обусловлено тем, что если в первом эксперименте под социальной группой подразумевалась американское общество в целом, то во втором – сообщество университета, чувство принадлежности к которому гораздо слабее. Однако в целом результаты исследования свидетельствуют о возможности влияния на разрешение внутреннего конфликта, которое выражается в укреплении прежних религиозных и политических взглядов, с помощью высокочастотной краткосрочной стимуляции. Таким образом, можно говорить об успешности снижения эффекта КД при применении как длительной низкочастотной, так и краткосрочной высокочастотной ТМС.
Некоторые исследователи обнаружили разницу в воздействии на КД в случае применения разных вариантов тех или иных протоколов: анодной/катодной ТЭС (Colosio et al., 2018); воздействия на правую или левую префронтальную кору (Mengarelli et al., 2015); влияния на противоположные взгляды участников (Chawke, Kanai, 2016). М. Колозио и соавт. (Colosio et al., 2018) предполагали, что анодная стимуляция задней МПФК усилит эффект КД, вызванный оценкой и выбором изображений продуктов питания в рамках модифицированной парадигмы свободного выбора. Катодная же стимуляция должна была элиминировать или по крайней мере снизить эффект КД. Однако авторы обнаружили статистически значимый эффект только катодной стимуляции. Подобные результаты могут быть связаны с компенсаторным влиянием ДЛПФК и неоптимальностью точки стимуляции. Кроме того, важно отметить, что исследователи осуществляли стимуляцию до стадии выбора (а не после, как в большинстве исследований КД), соответственно, возможно, раннее вовлечение в механизмы КД способно элиминировать эффект снятия внутреннего конфликта, но не усилить его. Ф. Менгарелли и соавт. (Mengarelli et al., 2015) использовали ТЭС в рамках модифицированной парадигмы свободного выбора, чтобы подавить активность ДЛПФК и, соответственно, снизить степень расхождения альтернатив в результате КД. В результате исследователи обнаружили межполушарные различия во влиянии на обработку внутреннего конфликта: воздействие на левую ДЛПФК снизило эффект КД. В тоже время воздействие на правую ДЛПФК не привело к статистически значимому эффекту по сравнению с плацебо стимуляцией. Исследователи объясняют данное различие несколькими потенциальными причинами: левая префронтальная кора в большей степени вовлечена в процессы рационализации, саморегуляции, самоконтроля, модуляции оценочных суждений, тогда как правая – в процессы детекции несоответствия. К. Чоке и Р. Канаи (Chawke, Kanai, 2016) применили ТЭС с целью повысить возбудимость ДЛПФК участников, которым предъявляли в качестве стимулов видео либеральной или консервативной политической кампании (участников отбирали с помощью пилотных исследований по параметру ярко выраженных либеральных или консервативных политических взглядов). В противовес изначальной гипотезе исследователи выявили, что представители ярко выраженных либеральных взглядов в гораздо большей степени поменяли взгляды в сторону консервативных взглядов в условиях стимуляции по сравнению с плацебо условием, тогда как представители консервативных взглядов не изменили взгляды на либеральные. Авторы полагают, что подобные результаты связаны с тем, что эффект КД обусловлен стремлением к стабильности и избеганию неопределенности. Таким образом, усиленный с помощью стимуляции эффект КД проявился не в изменении взглядов в соответствии с показанным видео, а в обоих случаях в сдвиге взглядов в сторону более консервативных, дающих чувство определенности и стабильности, что имеет большое значение для изучения КД.
ВЫВОДЫ
Одна из наиболее влиятельных теорий в общей и социальной психологии – теория КД – позволяет как предсказывать поведение человека в различных контекстах, так и влиять на его убеждения и предпочтения. Теоретические и методологические подходы, сформулированные в рамках теории КД, оказываются чрезвычайно продуктивными для исследований поведения как психологами, так и нейробиологами. Нейробиология КД оказывается в фокусе нейроэкономических исследований принятия решений и социального влияния. Раскрытие механизмов КД важно и для традиционной экономической науки, в рамках которой принятие решений рассматривается как результат предпочтений индивида, в то время как КД демонстрирует обратный процесс – решения влияют на предпочтения. Для психологических и социальных наук представляется важным создание нейробиологической модели КД. В биологической науке феномен КД является важным теоретическим вызовом для традиционной модели “обучения с подкреплением”, сфокусированной на результате выбора, а не на самом акте выбора. Более того, раскрытие нейробиологических механизмов влияния принятия решений на внутренние конфликты и формирование предпочтений поможет создать новые подходы для диагностики и терапии различных физических и психологических зависимостей (курение, наркомания, ожирение и др.), связанных с состоянием КД.
Несмотря на успехи предыдущих исследований, направленных на нейрокартирование процессов принятия решений, на сегодняшний день остается неясной временная динамика возникновения КД и последующего изменения установок. Большинство исследований КД сфокусированы на изучении его нейробиологических коррелят уже после сложного выбора – при повторном столкновении с отвергнутой альтернативой (Izuma et al., 2015; Mengarelli et al., 2015). Результаты ряда исследований продемонстрировали более ранние корреляты КД, возникающие уже в момент сложного выбора (Colosio et al., 2017; Lieberman et al., 2001). Более того, используя метод долговременных корреляций, М. Колозио и соавт. продемонстрировали, что активация МПФК в покое может служить маркером, отражающим будущие поведенческие реакции в ситуации КД (Colosio et al., 2017). Метод активного нейрокартирования с помощью ТМС и ТЭС – наиболее перспективный подход для проверки гипотезы о том, что ключевые нейробиологические процессы, которые обусловливают возникновение внутреннего конфликта и последующее изменение установок, протекают именно в процессе принятия сложного решения. Оба этих метода активного декодирования мозговой активности неинвазивные и вследствие этого получают все более широкое распространение в фундаментальных и клинических исследованиях мозга, сфокусированных на выявлении причины тех или иных процессов. Исследования с применением “хронометрической” ТМС и классической ТЭС МПФК дают возможность продемонстрировать связь МПФК и КД путем изменения активности МПФК в разные моменты времени, что, в свою очередь, должно повлиять на величину наблюдаемого КД.
Вместе с тем анализ протоколов стимуляции, проведенный в данной работе, позволил нам выделить те ограничения, которые накладывают методы ТЭС и ТМС на раскрытие механизмов обработки внутреннего конфликта: прежде всего, возможна стимуляция только поверхностных областей мозга, вовлеченных во внутренний конфликт, а именно МПФК и ДЛПФК. Это ограничивает степень воздействия стимуляции на такие механизмы, как когнитивный контроль и мониторинг когнитивных конфликтов.
Интерес представляет не только установление пространственно-временных характеристик каузальной связи МПФК и КД, но и получение самой возможности модулировать величину КД как в сторону уменьшения, так и увеличения. Подобные исследования способны изменить имеющиеся представления о механизмах возникновения, развития и модуляции КД и о проблеме адаптивного поведения в целом.
Список литературы
Козелецкий Ю. Психологическая теория решений. М.: Прогресс, 1979.
Корнилова Т.В. Психология риска и принятия решений. М.: Аспект Пресс, 2003.
Леонтьев Д.А., Овчинникова Е.Ю., Рассказова Е.И., Фам А.Х. Психология выбора. М.: Смысл, 2015.
Aronson E. Dissonance theory: Progress and problems. Theories of cognitive consistency: A sourcebook, 1968. 5–27.
Bem D.J. Self-perception theory. Advances in experimental social psychology, 1972. 6: 1–62.
Bem D.J. Self-perception: An alternative interpretation of cognitive dissonance phenomena. Psychological Review, 1967. 74: 183–200.
Brehm J.W. Postdecision changes in the desirability of alternatives. The Journal of Abnormal and Social Psychology, 1956. 52(3): 384–389.
Chawke C., Kanai R. Alteration of Political Belief by Non-invasive Brain Stimulation. Frontiers in Human Neuroscience, 2016. 9: 621.
Chen M.K., Risen J.L. How choice affects and reflects preferences: Revisiting the free-choice paradigm. Journal of Personality and Social Psychology, 2010. 99(4): 573–594.
Colosio M., Shestakova A., Nikulin V.V., Blagovechtchenski E., Klucharev V. Neural Mechanisms of Cognitive Dissonance (Revised): An EEG Study. Journal of Neuroscience, 2017. 37(20): 5074–5083.
Colosio M., Rybina E., Shestakova A., Klucharev V. Neural Mechanisms of Post-Decisional Spreading of Alternatives. Psychology, Journal of the Higher School of Economics, 2018. 15(3): 606–614.
Di Martino A., Scheres A., Margulies D.S., Kelly A.M.C., Uddin L.Q., Shehzad Z., Milham M.P. Functional connectivity of human striatum: A resting state fMRI study. Cerebral Cortex, 2008. 18(12): 2735–2747.
Egan L.C., Bloom P., Santos L.R. Choice-induced preferences in the absence of choice: Evidence from a blind two choice paradigm with young children and capuchin monkeys. Journal of Experimental Social Psychology, 2010. 46(1): 204–207.
Egan L.C., Santos L.R., Bloom P. The origins of cognitive dissonance: Evidence from children and monkeys. Psychological Science, 2007. 18: 978–983.
Festinger L. A theory of cognitive dissonance. Stanford University Press, 1957. 2: 292.
Floresco S.B. The nucleus accumbens: an interface between cognition, emotion, and action. Annual review of psychology, 2015. 66: 25–52.
Harmon-Jones E., Amodio D.M., Harmon-Jones C. Action-based model of dissonance: A review, integration, and expansion of conceptions of cognitive conflict. In M. P. Zanna (Ed.), Advances in experimental social psychology, 2009. 41: 119–166.
Harmon-Jones E., Harmon-Jones C., Serra R., Gable P.A. The effect of commitment on relative left frontal cortical activity: tests of the action-based model of dissonance. Pers Soc Psychol Bull, 2011. 37: 395–408.
Harmon-Jones E., Gerdjikov T., Harmon-Jones C. The effect of induced compliance on relative left frontal cortical activity: A test of the action-based model of dissonance. European Journal of Social Psychology, 2008. 38(1): 35–45.
Holbrook C., Iacoboni M., Gordon C., Proksch S., Makhfi H., Balasubramaniam R. Posterior medial frontal cortex regulates sympathy: A TMS study. Social Neuroscience, 2021. 16(6): 595–606.
Holbrook C., Iacoboni M., Gordon C., Proksch S., Balasubramaniam R. Posterior medial frontal cortex and threat-enhanced religious belief: A replication and extension. Social Cognitive and Affective Neuroscience, 2020. 15(12): 1361–1367.
Holbrook C., Izuma K., Deblieck C., Fessler D.M.T., Iacoboni M. Neuromodulation of group prejudice and religious belief. Social Cognitive and Affective Neuroscience, 2016. 11(3): 387–394.
Izuma K., Akula S., Murayama K., Wu D.-A., Iacoboni M., Adolphs R.A Causal Role for Posterior Medial Frontal Cortex in Choice-Induced Preference Change. Journal of Neuroscience, 2015. 35(8): 3598–3606.
Izuma K., Murayama K. Choice-Induced Preference Change in the Free-Choice Paradigm: A Critical Methodological Review. Frontiers in Psychology, 2013. 4: 41.
Izuma K., Matsumoto M., Murayama K., Samejima K., Sadato N., Matsumoto K. Neural correlates of cognitive dissonance and choice-induced preference change. Proceedings of the National Academy of Sciences, 2010. 107(51): 22014–22019.
Johansson P., Hall L., Chater N. Preference change through choice. Neuroscience of Preference and Choice, 2012. 121–141.
Johnson S.C., Baxter L.C., Wilder L.S., Pipe J.G., Heiserman J.E., Prigatano G.P. Neural correlates of self-reflection. Brain: A Journal of Neurology, 2002. 125(8): 1808–1814.
Kawabata H., Zeki S. The neural correlates of desire. PloS one, 2008. 3 (8): e3027.
Kitayama S., Chua H.F., Tompson S., Han S. Neural mechanisms of dissonance: An fMRI investigation of choice justification. NeuroImage, 2013. 69: 206–212.
Knutso B., Delgado M.R., Phillips P.E. Representation of subjective value in the striatum. In Neuroeconomics, 2009. 389–406.
Lee D., Daunizeau J. Choosing what we like vs liking what we choose: How choice-induced preference change might actually be instrumental to decision-making. PLoS one, 2020. 15(5): e0231081.
Leech R., Sharp D.J. The role of the posterior cingulate cortex in cognition and disease. Brain, 2014. 137(1): 12–32.
Leech R., Kamourieh S., Beckmann C.F., Sharp D.J. Fractionating the default mode network: distinct contributions of the ventral and dorsal posterior cingulate cortex to cognitive control. The Journal of Neuroscience, 2011. 31(9): 3217–3224.
Lieberman M.D., Ochsner K.N., Gilbert D.T., Schacter D.L. Do amnesics exhibit cognitive dissonance reduction? The role of explicit memory and attention in attitude change. Psychological Science, 2001. 12: 135–140.
Maddock R.J., Garrett A.S., Buonocore M.H. Remembering familiar people: The posterior cingulate cortex and autobiographical memory retrieval. Neuroscience, 2001. 104(3): 667–676.
Mengarelli F., Spoglianti S., Avenanti A., di Pellegrino G. Cathodal tDCS over the left prefrontal cortex diminishes choice-induced preference change. Cerebral Cortex, 2015. 25(5): 1219–1227.
Miller E.K., Cohen J.D. An integrative theory of prefrontal cortex function. Annual review of neuroscience, 2001. 24(1): 167–202.
Nitsche M.A., Nitsche M.S., Klein C.C., Tergau F., Rothwell J.C., Paulus W. Level of action of cathodal DC polarisation induced inhibition of the human motor cortex. Clinical Neurophysiology: Official Journal of the International Federation of Clinical Neurophysiology, 2003. 114(4): 600–604.
Nitsche M.A., Paulus W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology, 2001. 57(10): 1899–1901.
Northoff G., Heinzel A., de Greck M., Bermpohl F., Dobrowolny H., Panksepp J. Self-referential processing in our brain — A meta-analysis of imaging studies on the self. NeuroImage, 2006. 31(1): 440–457.
Pearson J.M., Heilbronner S.R., Barack D.L., Hayden B.Y., Platt M.L. Posterior cingulate cortex: Adapting behavior to a changing world. Trends in Cognitive Sciences, 2011. 15(4): 143–151.
Qin J., Kimel S., Kitayama S., Wang X., Yang X., Han S. How choice modifies preference: Neural correlates of choice justification. NeuroImage, 2011. 55(1): 240–246.
Reinhart R.M.G., Woodman G.F. Causal Control of Medial-Frontal Cortex Governs Electrophysiological and Behavioral Indices of Performance Monitoring and Learning. Journal of Neuroscience, 2014. 34(12): 4214–4227.
Rochat P. Five levels of self-awareness as they unfold early in life. Consciousness and Cognition, 2003. 12: 717–731.
Sharot T., Fleming S.M., Yu X., Koster R., Dolan R.J. Is Choice-Induced Preference Change Long Lasting? Psychological Science, 2012. 23(10): 1123–1129.
Sharot T., Velasquez C.M., Dolan R.J. Do decisions shape preference? Evidence from blind choice. Psychological science, 2010. 21(9): 1231–1235.
Tandetnik C., Sohier E., Capelle L., Boullay V. du, Obadia M., Chammat M., Pyatigorskaia N., Naccache L. Cognitive dissonance resolution depends on executive functions and frontal lobe integrity. Cortex, 2021. 139: 1–11.
Tompson S., Chua H.F., Kitayama S. Connectivity between mPFC and PCC predicts post-choice attitude change: The self-referential processing hypothesis of choice justification. Human Brain Mapping, 2016. 37(11): 3810–3820.
Turi Z., Lenz M., Paulus W., Mittner M., Vlachos A. Selecting stimulation intensity in repetitive transcranial magnetic stimulation studies: A systematic review between 1991 and 2020. The European Journal of Neuroscience, 2021. 53(10): 3404–3415.
Vann S.D., Aggleton J.P., Maguire E.A. What does the retrosplenial cortex do? Nature Reviews Neuroscience, 2009. 10(11): 792–802.
Voigt K., Murawski C., Speer S., Bode S. Hard decisions shape the neural coding of preferences. Journal of Neuroscience, 2019. 39(4): 718–726.
Voigt K., Murawski C., Speer S., Bode S. Effective brain connectivity at rest is associated with choice-induced preference formation. Human brain mapping, 2020. 41(11): 3077–3088.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова