

Журнал высшей нервной деятельности им. И.П. Павлова, 2023, T. 73, № 1, стр. 3-23
Зрительные механизмы второго порядка: обзор исследований
В. В. Бабенко *
Академия психологии и педагогики, Южный федеральный университет
Ростов-на-Дону, Россия
* E-mail: babenko@sfedu.ru
Поступила в редакцию 30.03.2022
После доработки 06.06.2022
Принята к публикации 27.06.2022
- EDN: GIYDEO
- DOI: 10.31857/S0044467723010057
Аннотация
Обзор посвящен анализу исследований, направленных на изучение зрительных фильтров второго порядка, как механизма преаттентивного группирования локальной зрительной информации, выделяемой стриарными нейронами (фильтрами первого порядка). Анализируется развитие представлений о возможных механизмах объединения первичных зрительных признаков, при этом акцент делается на исследованиях свойств фильтров второго порядка. Рассматриваются вопросы их пространственной организации и принципов взаимодействия фильтров первого и второго порядка. Описываются результаты, указывающие на многоканальность механизма группирования и возможную связь частотных и ориентационных настроек фильтров первого и второго порядка. Приводится модель организации фильтров второго порядка. Анализируются возможные нейрофизиологические корреляты указанных механизмов. Рассматривается возможная роль механизмов второго порядка в зрительном восприятии.
Исследованные Д. Хьюбелом и Т. Визелом стриарные нейроны реагировали на градиенты яркости, которые были признаны элементарными признаками изображений. В этой ситуации фактически любые варианты последующего объединения локальных признаков претендовали на роль “признаков второго порядка”. Речь могла идти об объединении градиентов яркости в крестообразные структуры (Шевелев, 2010) или об интеграции информации разной пространственной частоты или ориентации определенной локализации (Olzak, Thomas, 1992). Однако общим для такого рода операций было то, что они не выходили за рамки локальных. С другой стороны, интерес представляли и механизмы пространственной интеграции, обеспечивающие описание ансамблей объектов и вычисления их сводных статистик (Яковлев и др., 2020). Однако эти операции могут быть реализованы, по всей видимости, лишь после привлечения механизмов внимания и репрезентации отдельных объектов (Baek, Chong, 2020).
В этом обзоре речь пойдет о механизмах, которые являются первым шагом к выходу за пределы локальной обработки. Мы будем говорить об операциях, которые осуществляются в протяженной пространственной области, но при этом остаются автоматическими, преаттентивными. Именно эти процессы в последние годы стали относить к так называемому зрению второго порядка. В этой связи мы будем придерживаться следующих устоявшихся определений: информация первого порядка – это изменения яркости, информация второго порядка – это пространственно-временная модуляция информации первого порядка (Huang, Chen, 2014). Заметим, что данное определение не включает объединение коллинеарных градиентов яркости и операции наращивания контура. По-видимому, этот механизм следует рассматривать как самостоятельный (Field et al., 1993).
ЛОКАЛЬНАЯ ОБРАБОТКА В ЗРИТЕЛЬНОЙ СИСТЕМЕ ЧЕЛОВЕКА
В своей, ставшей классической, работе Ф. Кэмпбелл и Дж. Робсон (Campbell, Robson, 1968) сравнивали обнаружимость синусоидальных и прямоугольных решеток разной пространственной частоты. Полученный результат был первым убедительным свидетельством того, что паттерновое зрение на уровне порога обеспечивается набором независимых линейных каналов, настроенных на разные диапазоны пространственных частот. Эта гипотеза получила дополнительное подтверждение в их более поздних работах (напр., (Campbell et al., 1978)), в которых сравнивалась обнаружимость и измерялась различимость синусоидальных, прямоугольных решеток и решеток с удаленной основной частотой. Главный вопрос, который ставился в этих исследованиях, сводился к тому, могут ли результаты, полученные для синусоидальных стимулов, предсказать обнаружимость других решеток. Авторы выяснили, что яркостные паттерны, состоящие из множества гармоник, обнаруживались, когда одна из гармоник достигала порогового уровня.
Анализ линейных систем оказался удивительно успешным при моделировании порогового зрения. До 1980 г. огромное число исследований было посвящено детальному изучению свойств линейных полосовых фильтров, а многоканальная теория зрительной обработки добилась статуса базовой доктрины.
В те годы модель множества фильтров, настроенных на локализацию, ориентацию и пространственную частоту, представлялась очень хорошей моделью паттернового зрения, особенно когда речь шла об экспериментах, использующих зрительные стимулы порогового контраста. Ее представления базировались на классической модели одного из типов нейронов, открытых Д. Хьюбелом и Т. Визелом в стриарной коре, которые были названы ими простыми клетками (Hubel, Wiesel, 1959, 1962). Эти фильтры срабатывали автоматически при попадании в их рецептивное поле градиента яркости. Чтобы из множества фильтров получить ответ испытуемого, модель использовала правило решения, которое было весьма простой комбинацией выходов этих фильтров: например, испытуемый принимает решение, что паттерн является вертикальным тогда и только тогда, когда фильтр с наибольшим выходом имеет пиковую чувствительность к вертикальной ориентации. Когда же речь шла об увеличении контрастной чувствительности при возрастании числа циклов в синусоидальной решетке, это объяснялось вероятностной суммацией (Hoekstra et al., 1974; King-Smith, Kulikowski, 1975). Лишь значительно позднее были получены экспериментальные доказательства того, что увеличение чувствительности в данном случае связано с работой особого преаттентивного механизма, интегрирующего в пространстве градиенты яркости (Бабенко, 2001; Usher et al., 1999).
Линейные модели, хорошо зарекомендовавшие себя в условиях околопорогового восприятия, столкнулись также с проблемой объяснения эффектов нелинейности, возникавших при увеличении контраста стимулов. Нелинейность носила характер ускорения (расширения) при низких контрастах и замедления (сжатия) – при высоких (Legge, Foley, 1980). В нейрофизиологических исследованиях также была обнаружена нелинейность контрастного отклика нейронов зрительной коры кошек и приматов (Albrecht, Hamilton, 1982; Barlow et al., 1987; Dean, 1981; Maffei, Fiorentini, 1973; Ohzawa et al., 1985).
Еще одна проблема, с которой столкнулись линейные модели, – это сегментация текстур. Данные о преаттентивном разделении текстур сначала пытались объяснить, основываясь на паттерне выходов пространственно-частотных каналов. Планомерные попытки решения этой проблемы были предприняты Б. Джулесом (Caelli, Julesz, 1978; Julesz, 1980; Julesz et al., 1973), который предположил, что для разделения текстур зрительная система может использовать информацию о линейных сегментах и их окончаниях, вычисляя их число. Однако на каждое очередное предположение относительно использования статистик все более высокого порядка находились все новые контраргументы (Julesz et al., 1978). В итоге Б. Джулес предположил, что к разделению текстур может приводить разница в статистике “текстонов” по обе стороны текстурной границы (Julesz, 1981). Однако так и не удалось определить, что представляют собой текстоны и как должно быть организовано вычисление.
Вместе с тем появлялись все новые примеры разделения текстур, которые нельзя было объяснить их различиями в Фурье-спектрах. Например, речь может идти об обнаружении границы между двумя текстурами равной средней яркости, составленными из по-разному ориентированных Габоровских микропаттернов (рис. 1а). Хотя области по обеим сторонам границы имеют одинаковый Фурье-спектр, разделение текстур происходит автоматически, преаттентивно.
Рис. 1.
Текстурные стимулы второго порядка. Граница в Габоровской текстуре (а); случайно-точечные паттерны, синусоидально модулированные по яркости (б) и контрасту (в); синусоидальная решетка, модулированная по контрасту, с наложенным на нее рецептивным полем фильтра 1-го порядка (г); полосовые текстуры, синусоидально модулированные по пространственной частоте (д), ориентации (е) и контрасту (ж). Fig. 1. The second-order textures. Border in Gabor texture (а); random dot patterns sinusoidally modulated in luminance (б) and contrast (в); contrast-modulated sinusoidal grating with a 1st-order filter receptive field superimposed on it (г); stripe textures sinusoidally modulated in spatial frequency (д), orientation (е), and contrast (ж).

Если посмотреть на изображения, показанные на рис. 1б и в, легко заметить, что обе текстуры неоднородны. Модуляцию яркости (1б) обнаруживают фильтры 1-го порядка. Однако они не могут обнаружить модуляцию контраста (1в). Поскольку выходной сигнал фильтра 1-го порядка представляет собой линейное суммирование ответов от возбуждающих и тормозных областей, линейные фильтры не могут использоваться зрительной системой для восприятия модуляций контраста, так как в данном случае каждое их субполе (и возбудительное и тормозное) стимулируется одинаковым количеством светлого и темного (рис. 1г).
Эти и многие другие факты поставили вопрос о том, достаточно ли локальных линейных фильтров, которые впоследствии были названы механизмами 1-го порядка, чтобы объяснить работу зрительной системы человека на преаттентивном уровне обработки. Становилось все более очевидным, что необходим дополнительный механизм, группирующий пространственно распределенные градиенты яркости. По-видимому, первое предположение относительно преаттентивного объединения локальных фильтров принадлежит Г. Хеннингу с соавторами (Henning et al., 1975).
ПРОБЛЕМА ПРОСТРАНСТВЕННОГО ОБЪЕДИНЕНИЯ ЛОКАЛЬНОЙ ЗРИТЕЛЬНОЙ ИНФОРМАЦИИ
Для решения многих зрительных задач информация, выделяемая линейными фильтрами 1-го порядка, должна быть сгруппирована. Это необходимо и для выделения объектов из фона, и для сегментации текстур, и для восприятия движущихся объектов (проблема апертуры), и для многих других задач. А. Трейзман и Г. Гелад в своей теории интеграции признаков предположили, что эта операция реализуется с помощью избирательного внимания. То есть само внимание является механизмом связывания “свободно плавающих признаков”, выделяемых преаттентивно (Treisman, Gelade, 1980).
Главным инструментом экспериментальной проверки этой теории стал зрительный поиск. Если увеличение числа дистракторов не приводило к возрастанию времени поиска цели, результат интерпретировался как доказательство того, что целевой признак обнаруживается автоматически. А если при инвертировании цели и дистракторов (дистрактор становился целью, а целевые объекты – дистракторами) поиск становился последовательным, признак, обнаруживаемый автоматически, оценивался как “базовый” (Treisman, Souther, 1985; Wolfe, 2001). Эффект асимметрии поиска стал широко использоваться для выявления все новых базовых признаков, однако очень скоро их число насчитывало уже около двух десятков. Причем далеко не во всех этих случаях можно было говорить о действительно простых признаках, выделяемых начальными уровнями корковой обработки (напр., (Не, Nakayama, 1992; Levin et al., 2001)). Это поставило под сомнение “примитивность” большинства базовых признаков и возможность их обнаружения с помощью известных преаттентивных механизмов.
В конце 80-х годов прошлого века почти одновременно появляется несколько работ, которые предположили возможность группирования пространственно распределенной локальной зрительной информации уже на стадии, предшествующей избирательному вниманию.
Так, Ч. Чуб и Г. Сперлинг предложили новый вид стимула, который они назвали стимулом случайного “сбалансированного смещения” (a “drift-balanced” random stimulus) (Chubb, Sperling, 1988). Этот стимул представляет собой текстуру, состоящую из неподвижных элементов, внутри которой смещается область, отличающаяся по контрасту от остальной части текстуры. Такое смещение тут же замечается наблюдателем, хотя линейные фильтры первого порядка нечувствительны к этому типу движения. Поскольку стимулы такого типа не имеют энергии в Фурье-области на соответствующей пространственной частоте, авторы назвали такое движение не-Фурье-движением (движением 2-го порядка), в отличие от Фурье-движения – 1-го порядка. Как показали авторы этой работы, движение 2-го порядка может быть обнаружено только в результате объединения выходов линейных фильтров на так называемых механизмах 2-го порядка. В. Бабенко, со своей стороны, предложил схему группирования механизмов первого порядка, способную объяснить механизм масштабной инвариантности зрительной сегментации (Бабенко, 1989). А для объяснения механизма разделения текстур в раннее зрение была включена вторая стадия ориентационно- и частотно-избирательной фильтрации, на которой группируются выходы фильтров первой стадии (Sutter et al., 1989).
Надо сказать, что модель организации преаттентивной обработки, предполагающая объединение выпрямленных выходов фильтров 1-го порядка на так называемых фильтрах 2-го порядка, оказалась весьма подходящей для объяснения множества феноменов зрительного восприятия: определения центра тяжести кластера точек противоположного контраста (Morgan, Glennerster, 1991), разделения текстур с одинаковой амплитудой Фурье-спектров (Graham et al., 1992), различения крупномасштабных локализаций (Hess, Holliday, 1992), обнаружения иллюзорных контуров (Wilson, Richards, 1992), вычисления формы из текстуры (Sakai, Finkel, 1995) и др. Этот класс моделей можно было использовать также и для объяснения обнаружения в текстурах синусоидальных модуляций контраста (Sutter et al., 1995), ориентации (Kingdom, Keeble, 1996) и пространственной частоты (Arsenault et al., 1999) (см. рис. 1 (д–ж)).
Потенциальная возможность объяснения многих феноменов зрительного восприятия с использованием механизма преаттентивного пространственного группирования послужила толчком к появлению в 90-х гг. большого числа работ, направленных на доказательство реальности этого механизма. Как правило, в такого рода исследованиях изучалась обработка сложных границ второго порядка (напр., (Rubenstein, Sagi, 1993)). Эти границы могли быть контрастными, текстурными (изменения ориентации, размера или пространственной частоты элементов), движущимися и т.д. Но какие бы стимулы ни использовались, информация 2-го порядка представляла собой пространственную модуляцию информации 1-го порядка. Таким образом, любые стимулы 2-го порядка состоят из несущей (это, как правило, синусоидальная решетка либо текстура), которая модулирована огибающей.
Итак, к середине 90-х годов сложилось устойчивое представление о том, что обнаружение структур второго порядка предполагает существование двух стадий фильтрации, разделенных нелинейностью. При этом на фильтрах 2-го порядка объединяются выпрямленные выходы фильтров 1-го порядка, отличающихся только локализацией (Graham, 1994; Kingdom et al., 1995). Но чтобы доказать правомерность этой модели, в первую очередь следовало убедиться в том, что обработка 2-го порядка действительно выполняется механизмом, отличным от механизма, отвечающего за обработку 1-го порядка.
Этот вопрос возник в том числе и в связи с тем, что попытки обнаружить нейроны, реагирующие исключительно на стимулы второго порядка, не дали ожидаемого результата. Было показано, что около 50% нейронов полей 17 и 18 кошки (Mareschal, Baker, 1998a; Zhou, Baker, 1993) и около 30% нейронов V2 приматов (Li et al., 2014) реагируют на огибающую. Но эти же нейроны реагировали и на несущую. С другой стороны, было бы странно ожидать, что клетки, избирательные к стимулам 2-го порядка, не будут активироваться стимулами 1-го порядка. Ведь, согласно существующим представлениям, их вход сформирован нейронами, реагирующими на отдельные градиенты яркости. Как и следовало ожидать, в последующих исследованиях зрительных механизмов второго порядка основная ставка была сделана на психофизические показатели.
НЕЗАВИСИМОСТЬ МЕХАНИЗМОВ ПЕРВОГО И ВТОРОГО ПОРЯДКА
Один из принципиальных вопросов относительно восприятия стимулов 2-го порядка заключался в том, действительно ли для этого необходим специальный механизм. К. Лэнгли с соавторами (Langley et al., 1996) адаптировали испытуемых к синусоидальным решеткам, а затем измеряли, насколько повышался порог обнаружения контрастно-модулированных решеток. Максимально порог повышался (примерно в 16 раз), когда ориентация и пространственная частота адаптирующей решетки совпадала с несущей, но не с огибающей. Этот результат подтверждал, что обнаружению структур 2-го порядка предшествует полосовая фильтрация по ориентации и пространственной частоте, и указывал на независимость 1-й и 2-й стадии. Отсутствие эффектов взаимной адаптации стимулов 1-го и 2-го порядка было показано также в работе С. Нишида с соавторами (Nishida et al., 1997).
В другом похожем исследовании использовали эффект пьедестала (Schofield, Georgeson, 1999). Облегчающая часть этого эффекта может использоваться для исследования того, обрабатываются ли целевые и маскирующие стимулы одним и тем же механизмом. Если облегчение имеет место, когда маска и цель представляют разные типы информации, можно сделать вывод, что они обрабатываются перекрывающимися механизмами. Если перекрестного облегчения нет, тогда маска и цель, вероятно, обрабатываются раздельно. Следуя этому обоснованию, авторами было показано, что модуляция яркости 1-го порядка и модуляция контраста 2-го порядка обрабатываются разными механизмами. Отсутствие взаимного маскирующего эффекта между стимулами 1-го и 2-го порядка показано также в работе Р. Алларда и Дж. Фоберта (Allard, Faubert, 2007).
Было также продемонстрировано, что обнаружение контрастно-модулированных стимулов нечувствительно к структуре и спектру несущей (частотные характеристики шума). Это является еще одним свидетельством того, что модуляции 1-го и 2-го порядка обрабатываются разными механизмами, “каждый из которых защищен от влияния другого” (Schofield, Georgeson, 2003). Независимость этих механизмов была подтверждена и данными об отсутствии переноса перцептивного обучения с системы 1-го порядка на систему 2-го порядка (Gong, Olzak, 2014).
Свой вклад в решение этого вопроса внесли также электрофизиологические исследования. Регистрация нейронов 17-го и 18-го полей кошки (Mareschal, Baker, 1998b), а также зрительных ВП человека (Calvert et al., 2005) продемонстрировала более длительную задержку при обработке стимулов 2-го порядка по сравнению со стимулами 1-го порядка.
Таким образом, были получены убедительные доказательства того, что обработка 2-го порядка выполняется иным механизмом, нежели механизм, отвечающий за обработку 1-го порядка. А разделение фильтров 1-го и 2-го порядка обеспечивается промежуточной нелинейностью. Нелинейность между двумя стадиями линейного фильтра необходима, потому что в противном случае объединение выходных сигналов от разных линейных фильтров первой стадии будет нивелировано и два последовательных линейных фильтра сведутся до одного линейного фильтра. Модуляция 2-го порядка не будет извлечена из входного сигнала таким фильтром.
Для объяснения разделяющего механизма почти все исследователи привлекали некоторую форму статической нелинейности, следующую вслед за линейным фильтром 1-й стадии, а некоторые также включали и контроль прироста: за выходом линейного фильтра 1-й стадии следует его статическая нелинейность с последующей нормализацией объединенным ответом других фильтров. Н. Грэхэм и А. Саттер предположили, что таким контролем прироста является торможение между каналами, как в сети нормализации (Graham, Sutter, 2000). Более поздние исследования подтвердили это предположение (Huang, Chen, 2014; Wang et al., 2012).
ПЕРЕДАТОЧНАЯ ФУНКЦИЯ МОДУЛЯЦИИ ГРАДИЕНТОВ ЯРКОСТИ И МНОГОКАНАЛЬНОСТЬ ЗРИТЕЛЬНЫХ МЕХАНИЗМОВ ВТОРОГО ПОРЯДКА
Изучение зрительных механизмов 2-го порядка началось, как в свое время и изучение механизмов 1-го порядка, с определения передаточной функции. И хотя в разных работах имели место некоторые различия в выборе стимулов и диапазона пространственных частот несущей и огибающей, передаточная функция модуляции контраста оказалась низкопередаточной (Jamar, Koenderink, 1985; Schofield, Georgeson, 1999, 2003). Правда, она была более широкополосной, чем аналогичная функция для механизмов 1-го порядка (Sutter et al., 1995). Похожая картина наблюдалась и для модуляции ориентации (Kingdom et al., 1995). Ф. Киндом с соавторами использовали синусоидальную модуляцию ориентации в микропаттерновых текстурах и обнаружили, что кривая была практически плоской, хотя и с тенденцией в сторону низкочастотной передачи огибающей с пиком между 0.06 и 0.2 ц/град (в зависимости от размера микропаттерна). М. Лэнди и П. Орук (Landy, Oruç, 2002) установили, что чувствительность к модуляции и контраста, и ориентации была почти плоской в диапазоне 5 октав. Однако она становилась полосовой, когда отображалась в виде эффективности (относительно идеального наблюдателя, решающего ту же задачу).
Естественно, возник вопрос, является ли такая передаточная функция отражением работы одного широкополосного канала или набора узкополосных каналов, настроенных на разные пространственные частоты и ориентации огибающей.
Избирательность механизмов второго порядка к пространственной частоте и ориентации была доказана с использованием методов адаптации (Kwan, Regan, 1998), маскировки (Arsenault et al., 1999) и околопороговой суммации (Landy, Oruc, 2002). Эти и другие подобные исследования явились убедительным свидетельством многоканальности на уровне механизмов 2-го порядка.
Д. Элемберг с соавторами (Ellemberg et al., 2006) сравнили число пространственно-частотных и ориентационных механизмов, лежащих в основе обработки 1-го и 2-го порядка путем измерения различения Габоровских стимулов 1-го и 2-го порядка на уровне порога обнаружения. Стимулы 2-го порядка были созданы путем умножения синусоидального компонента Габора на двумерный бинарный шум. Определялись пороговые различия этих стимулов по частоте и ориентации. Число ориентационных каналов оказалось одинаковым для фильтров 1-го и 2-го порядка. Что касается частотных каналов, то для паттернов 2-го порядка число каналов было тем же, что и для решеток 1-го порядка, вплоть до частоты 2 ц/г. Однако их было больше для 1-го порядка на более высоких пространственных частотах. Это согласуется с тем, что вторая стадия фильтрации смещается к более низким частотам. Самый низкочастотный механизм оказался настроен на 0.1 ц/г, а самый высокочастотный – на 6 ц/г. Всего пространственно-частотных каналов 2-го порядка было выявлено 5, а ориентационных – 6.
Хотя данная парадигма является инструментом определения числа каналов, она не позволяет измерять их полосы пропускания. Потребовались дополнительные исследования, которые показали, что фильтры 2-го порядка имеют ширину настройки на пространственную частоту огибающей порядка 1 октавы (Landy, Henry, 2007), а на ориентацию огибающей – около 30 градусов (Reynaud, Hess, 2012).
БАЗОВАЯ МОДЕЛЬ МЕХАНИЗМА ВТОРОГО ПОРЯДКА
Психофизические исследования показали, что фильтры 1-го порядка, которые формируют вход фильтров 2-го порядка, настроены на ориентацию и пространственную частоту градиентов яркости и делают свертку входного изображения в наборе пространственных шкал и ориентаций (Dakin, Mareschal, 2000; Graham et al., 1993; Hunt, Meinhardt, 2021; Motoyoshi, Nishida, 2004).
Выход первой стадии фильтрации подвергается нелинейным преобразованиям – полноволновому выпрямлению – суммированию полуволновых выпрямленных ответов в парах on- и off- центральных фильтров. Нелинейность делает все ответы положительными, тем самым препятствуя их взаимному уничтожению при линейном объединении на второй стадии.
Фильтры 2-го порядка суммируют нелинейно трансформированные выходы фильтров 1-го порядка и имеют более крупные рецептивные поля (рис. 2 (а)). Промежуточная нелинейность передает на вторую стадию энергетический ответ, сглаживая мелкие детали, а фильтры 2-го порядка обнаруживают разницу в ответах фильтров 1-го порядка.
Рис. 2.
(а) – схема формирования рецептивного поля фильтра 2-го порядка (модель “фильтрация-выпрямление-фильтрация”). Выходы фильтров 1-го порядка (стриарных нейронов), рецептивные поля которых отличаются только локализацией в поле зрения, после выпрямления объединяются на одном из фильтров 2-го порядка (постстриарном нейроне), формируя рецептивное поле с оппонентной организацией. (б) – показано положение рецептивного поля фильтра 2-го порядка относительно контрастно-модулированной текстуры, при котором суммарный возбудительный вход, формирующий центр рецептивного поля, не компенсируется торможением с его флангов. Fig. 2. (а) Scheme of the formation of the receptive field of the 2nd order filter (filtering-rectifying-filtering model). The outputs of the 1st order filters (striate neurons), whose receptive fields differ only in their localization in the visual field, are combined on one of the 2nd order filters (poststriate neuron) after rectifying and form a receptive field with an opponent organization. (б) – It is demonstration of the receptive field position of the 2nd order filter relative to the contrast-modulated texture, at which the total excitatory input, which forms the center of the receptive field, is not compensated by inhibition from its flanks.
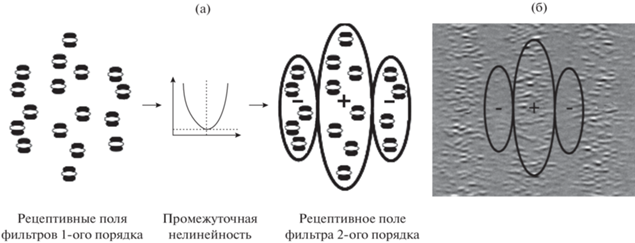
Характерная особенность модели “фильтрация-выпрямление фильтрация” состоит в том, что фильтр 2-го порядка не реагирует на однородную текстуру. Это объясняется тем, что все его субполя формируются выходами фильтров 1-го порядка с одинаковыми частотными и ориентационными настройками (отличающимися только локализацией). В результате в случае однородной (немодулированной) текстуры возбудительный вход центрального субполя компенсируется тормозным входом фланговых субполей. Фильтр 2-го порядка активируется, когда сигналы с флангов рецептивного поля снижаются, как, например, в случае, показанном на рис. 2 (б).
Поскольку модуляция контраста текстуры приводит к снижению контраста в областях, совпадающих с позицией тормозных флангов соответствующих фильтров, возбудительный вход превалирует над тормозным, и это обусловливает активацию данного механизма 2-го порядка. Следует подчеркнуть, что базовая модель “фильтрация-выпрямление-фильтрация” не избирательна к тому, какой параметр текстуры модулируется. Фильтры будут активироваться при модуляции любой размерности: и контраста, и ориентации, и пространственной частоты градиентов яркости.
Итак, базовая модель преаттентивной фильтрации предполагает, что сначала стимул фильтруется набором линейных фильтров 1-й стадии, настроенных на различные ориентации и пространственные частоты. Эта часть модели аппроксимирует вейвлет-трансформацию. Затем выходы этих фильтров являются объектом нелинейности (в разных вариантах модели это может быть возведение в квадрат, полу- или полноволновое выпрямление, нормализация и др.). После этого карты энергии ответов фильтров 1-го порядка на каждой масштабной шкале и ориентации фильтруются механизмами 2-го порядка с более крупными рецептивными полями.
СВЯЗЬ ЧАСТОТНЫХ И ОРИЕНТАЦИОННЫХ НАСТРОЕК ФИЛЬТРОВ ПЕРВОГО И ВТОРОГО ПОРЯДКА
Инвариантность восприятия к масштабным изменениям является одним из фундаментальных механизмов зрительной системы (Cannon, Fullenkamp, 1991; Jamar, Koenderink, 1983; Landy, Bergen, 1991; Savoy, McCann, 1975; Virsu, Rovamo, 1979). Именно благодаря такой инвариантности мы одинаково воспринимаем объекты, находящиеся от нас на разном расстоянии.
Поскольку речь идет о восприятии протяженных паттернов, этот механизм не может быть реализован на уровне локальных фильтров, описывающих отдельные градиенты яркости. Модели организации начальных уровней зрительной системы, способные объяснить инвариантность к масштабным преобразованиям, были сформулированы на рубеже 80-х и 90-х гг. (Бабенко, 1989; Шелепин и др., 1992; Babenko, 1991). В частности, была предложена схема, в которой двумерные матрицы одинакового числа элементов 1-го уровня (стриарных нейронов) с одинаковым размером рецептивных полей объединяются на элементах 2-го уровня. Второй уровень на каждом участке поля зрения представлен полями разного размера (пропорционального величине полей входной матрицы), описывающими входное изображение с разным разрешением. На этом уровне обработки происходит сегментация изображения на отдельные объекты (более крупными полями) и на фрагменты этих объектов (более мелкими полями). Изменение размера воспринимаемого объекта сопровождается его “переходом” в другое, соответствующее по размеру, рецептивное поле второго уровня. Благодаря универсальному принципу формирования этих полей изменение размера анализируемого объекта не отражается на его описании.
Такая организация связей, обеспечивающая инвариантность к масштабным преобразованиям, фактически является моделью формирования возбудительного (центрального) субполя фильтра 2-го порядка и основана на связи между шкалами фильтров 1-го и 2-го порядка. Пирамидальная организация зрительной системы, в которой взаимосвязаны пространственные шкалы анализа локальных градиентов яркости и глобальных градиентов текстур, была также предложена Дж. Бергеном (Bergen, 1991). Таким образом, как было предсказано теоретически, именно связь частотных настроек фильтров первой и второй стадии может обеспечить масштабную инвариантность.
Последующие исследования подтвердили существование такой связи (Kingdom, Keeble, 1999; Sutter et al., 1995). Масштабная инвариантность была продемонстрирована в задачах обнаружения модуляции контраста (Dakin, Mareschal, 2000; Sutter et al., 1995) и ориентации (Kingdom et al., 1995). Еще одним подтверждением этого стал и тот факт, что шкалы 1-го и 2-го порядка с эксцентриситетом меняются синхронно (Solomon, Sperling, 1995; Vakrou et al., 2007).
Естественно, возник вопрос, какова взаимосвязь частотных настроек фильтров 1-го и 2-го порядка в количественном выражении. Данные, полученные разными авторами, отличались, но все они говорили о наличии “жесткой” связи. Так, Д. Саги оценил отношение масштабов 1-го и 2-го порядка как 1/12 (Sagi, 1990), Н. Грэхэм – как 1/3 – 1/4 (Graham, 1994), а А. Саттер с соавторами оценили это соотношение как 1/8 : 1/16 (Sutter et al., 1995). Расхождение полученных соотношений вероятнее всего связано с различиями в использованных стимулах и с особенностями решаемой испытуемыми зрительной задачи. Так, Д. Саги использовал Габоровские патчи в задаче зрительного поиска, Н. Грэхэм применяла упорядоченные текстуры и задачу обнаружения, а А. Саттер с соавторами использовали полосовой изотропный шум, модулированный Габором, в задаче различения ориентаций. Последующее исследование В. Бабенко с соавторами (Бабенко и др., 2001) было попыткой прояснить этот вопрос, учтя возможные методические проблемы первых лет изучения механизмов 2-го порядка (см. Dakin, Mareschal, 2000). В результате было установлено, что пространственная интеграция градиентов яркости в центральном субполе фильтров 2-го порядка составляет 3–4 цикла несущей. То есть частотные настройки фильтров 1-го и 2-го порядка соотносятся как 1/7 : 1/8. Более того, они сохранялись при изменении пространственной частоты несущей в широком частотном диапазоне. Впоследствии такое соотношение частоты несущей и огибающей было подтверждено Пенг Сан и А. Шофилдом (Sun, Schofield, 2011).
Если относительно жесткой взаимосвязи частотных настроек фильтров 1-го и 2-го порядка сомнений практически не оставалось (Graham, 1994; Kingdom, Keeble, 1999; Sagi, 1990; Sutter et al., 1995; Wenderoth et al., 2001 и др.), то в отношении соответствия их ориентационных настроек картина оказалось иной. Данные одних авторов указывали на совпадение ориентационных настроек фильтров 1-го и 2-го порядка (Wolfson, Landy, 1995; Graham, Wolfson, 2001). Данные других свидетельствовали о взаимной перпендикулярности ориентационных настроек (Dakin, Mareschal, 2000). Третьи же говорили об отсутствии какой-либо связи ориентации огибающей с ориентацией несущей (Arsenault et al., 1999).
Такое расхождение выводов может быть связано с тем, что данные были получены при использовании в этих экспериментах пространственных модуляций разных размерностей. Так, С. Вольфсон и М. Лэнди использовали текстуры с модуляцией ориентации, С. Дакин и И. Марешал – с модуляцией контраста, а А. Арсенолт с соавторами – с модуляцией пространственной частоты. Правда, механизм “фильтрация-выпрямление-фильтрация” не должен делать различий в отношении размерностей модуляций. А значит, таких расхождений, как описано выше, быть не должно. Но они становятся возможны, если модуляции контраста, ориентации и пространственной частоты обнаруживаются разными фильтрами 2-го порядка.
СПЕЦИФИЧНОСТЬ ЗРИТЕЛЬНЫХ МЕХАНИЗМОВ ВТОРОГО ПОРЯДКА
Одним из самым дискуссионных стал вопрос о специфичности механизмов второго порядка. Впервые предположение о том, что модуляции контраста, ориентации и пространственной частоты обрабатываются разными независимыми механизмами, было высказано Ф. Киндомом с соавторами (Kingdom et al., 2003). В этом исследовании тестировался эффект пьедестала между различными типами модуляций второго порядка и не было обнаружено облегчающего эффекта, когда маска пьедестала и цель отличались размерностью модуляции. Однако в более позднем исследовании Ф. Шофилд и Т. Йейтс (Schofield, Yates, 2005) обнаружили слабое облегчение между обработкой модуляции контраста и модуляции ориентации. Они предположили, что отсутствие облегчения в работе Ф. Киндома с соавторами могло быть связано с фазовыми отношениями несущей и огибающей, а также c использованными уровнями пьедестала. В то же время ими не исключалось, что интеграция информации о модуляции ориентации и контраста возможна лишь на вышележащих уровнях обработки (Cruickshank, Schofield, 2005). Более того, последующие результаты все же убеждали в разделении механизмов, чувствительных к модуляциям контраста и ориентации (Schofield et al., 2007). Очевидно, что этот вопрос требовал решения. В нашей лаборатории была предпринята серия исследований, ставящих своей целью дать ответ на этот вопрос.
В психофизических исследованиях было показано, что эффекты адаптации и маскировки в задаче обнаружения пространственной модуляции имеют место только в том случае, когда в обоих взаимодействующих стимулах модулируется один и тот же параметр текстуры (Babenko, Yavna, 2008). Это прямо указывало на специфичность фильтров 2-го порядка. С другой стороны, было обнаружено, что зрительный поиск цели второго порядка в условиях ее неопределенности (размерность модуляции меняется в ходе эксперимента) не зависит от числа дистракторов и является эффективным (Babenko, Yavna, 2010). Этот результат исключал возможность перепрограммирования механизмов обнаружения определенной модуляции “сверху” и позволил сделать вывод о том, что механизмы второго порядка представляют собой фиксированный набор преаттентивных фильтров, специфичных к размерности модуляции. Психофизические доказательства специфичности зрительных механизмов второго порядка были дополнены электрофизиологическими данными, которые продемонстрировали, что модуляции контраста, ориентации и пространственной частоты вызывали зрительные ВП, существенно отличающиеся по своим характеристикам (Babenko, Ermakov, 2015). Таким образом, полученные результаты указывали на избирательность зрительных механизмов второго порядка к модулируемому параметру текстуры.
Доказательство специфичности зрительных механизмов второго порядка вновь поставило вопрос о возможных особенностях их организации. И действительно было обнаружено разное оптимальное соотношение частот несущей и огибающей для модуляций контраста, ориентации и пространственной частоты (Бабенко и др., 2010). А это, в свою очередь, вновь свидетельствовало в пользу избирательности фильтров 2-го порядка к модулируемому параметру текстуры (рис. 3).
Рис. 3.
Зависимость пороговой модуляции от соотношения частот огибающей и несущей. Колонка (а) – модуляция контраста, (б) – модуляция ориентации, (в) – модуляция пространственной частоты. Сплошная линия – ось модуляции и ориентация элементов текстуры перпендикулярны, пунктир – параллельны. Ось абсцисс – отношение частот огибающей и несущей. Ось ординат – пороговая амплитуда модуляции (дБ) (из статьи Бабенко и др., 2010). Минимальные значения пороговой амплитуды модуляции достигаются при оптимальном соотношении пространственных частот огибающей и несущей. Fig. 3. The dependence of threshold modulation on the frequency ratio of envelope and carrier. Column (а) is contrast modulation, (б) is orientation modulation, (в) is spatial frequency modulation. The solid line is when the axis of modulation and axis of the texture elements orientation are perpendicular, the dotted line is when they are parallel. The x-axis is the frequency ratio of the envelope and carrier. The y-axis is the threshold modulation amplitude (dB) (from Babenko et al., 2010). The minimum values of the threshold amplitude of the modulation occur when the frequency ratio of the envelope and the carrier is optimal.
Факты, указывающие на специфичность фильтров 2-го порядка, ставили вопрос о внесении изменений в базовую модель этих механизмов. В этой связи было показано, что избирательность к размерности модуляции достигается благодаря разной организации входов, формирующих фланговые субполя фильтров 2-го порядка (Явна, 2012).
Следует заметить, что специфичность фильтров 2-го порядка выглядит более целесообразным вариантом их организации, чем неспецифичность, поскольку модуляции разной размерности содержат информацию разного назначения. Модуляции контраста могут быть важны для выделения объектов из фона и их сегментации на фрагменты (Бабенко, Кульба, 2002), модуляции ориентации – для сегментации поверхностей в естественных сценах (DiMattina, Baker, 2021), а изменения пространственной частоты вызывают сильное впечатление сжатия поверхности (Sakai, Finkel, 1995) и относительной удаленности (третье измерение).
ВОЗМОЖНАЯ РОЛЬ МЕХАНИЗМОВ ВТОРОГО ПОРЯДКА В ЗРИТЕЛЬНОМ ВОСПРИЯТИИ
Фильтры 2-го порядка традиционно исследовались как механизм сегментации текстур. Несмотря на то, что текстуры – частый компонент самых разных естественных изображений, все же возникает вопрос: неужели их роль в зрительном восприятии ограничивается решением такой весьма узкой задачи, как разделение текстур? Только лишь для этого эволюция создала эти механизмы?
Давайте зададимся вопросом: а что это такое – модуляции градиентов яркости, которые обнаруживаются фильтрами 2-го порядка? Очевидно, что речь идет об областях пространственной неоднородности. А ведь именно области изображения, отличающиеся от окружения, представляют наибольший интерес (Gao et al., 2009; Gao, Vasconcelos, 2007; Itti, Koch, 2001; Itti et al., 1998). Общепринятой стала точка зрения, что мерой “визуальной заметности” элемента сцены является его отличие от окружения (Hou et al., 2013; Perazzi et al., 2012). Как мы выяснили выше, фильтры 2-го порядка реагируют на пространственные неоднородности в изображении. Эти неоднородности распределены по полю зрения и имеют разную выраженность. Реакции фильтров 2-го порядка будут пропорциональны амплитуде модуляции градиентов яркости в их рецептивных полях. Благодаря механизму латерального торможения дополнительное преимущество получат наиболее активированные фильтры. Это согласуется с выводом о том, что в системе зрительных механизмов 2-го порядка действует правило “победитель получает все” (Westrick et al., 2013). Именно эти “фильтры-победители” выделяют области изображения, которые и являются наиболее перцептивно заметными.
С другой стороны, очевидно, что внимание должно направляться на наиболее информативные участки изображения (Bruce, 2005; Bruce, Tsotsos, 2006). Очевидно, что способность выбирать в изображении области дальнейшей обработки оптимизирует процесс обработки (Mnih et al., 2014).
Смещения взора – очевидный показатель сдвига внимания в поле зрения. И локализация начальных точек фиксации (за исключением “центральной тенденции”) коррелирует с отличимостью областей от окружения (Parkhurst et al., 2002; Бабенко и др., 2020). При этом чем больше отличие, тем выше вероятность фиксации взора ('t Hart et al., 2013). Таким образом, целями непроизвольного внимания являются области пространственной неоднородности. Хорошо известно, например, что непроизвольный сдвиг внимания вызывают области изображения, отличающиеся от окружения по ориентации градиентов яркости (Ben-Shahar et al., 2007; Fuchs et al., 2011; Joseph, Optican, 1996) или их контрасту (Açık et al., 2009; Einhauser, Konig, 2003).
Итак, внимание направляется на области изображения, которые находят механизмы второго порядка. Могут ли фильтры 2-го порядка быть “окнами”, через которые внимание получает информацию и передает ее на постаттентивные уровни обработки?
МЕХАНИЗМЫ ВТОРОГО ПОРЯДКА И ВОСПРИЯТИЕ ЕСТЕСТВЕННЫХ СЦЕН
До сих пор мы говорили о работе фильтров 2-го порядка главным образом применительно к задаче сегментации текстур. А как поведут себя эти фильтры при восприятии естественных изображений, таких, например, как сцены? Очевидно, что фильтры “найдут” во входном изображении области пространственных неоднородностей разного размера. Наибольшей активностью будут обладать фильтры, соответствующие по размеру выделенным фрагментам. Самые крупные механизмы выделят крупные фрагменты сцены и отдельные объекты, более мелкие обнаружат неоднородности внутри объектов. Все эти области, как мы отмечали выше, являются, по всей видимости, наиболее информативными участками изображений и способны притягивать внимание.
Как этот механизм может быть реализован на нейронном уровне? Клетки со свойствами фильтров 2-го порядка были обнаружены в различных зрительных областях коры и у обезьян (Albright, 1995; Chaudhuri, Albright, 1997; Geesaman, Anderson, 1996; Grosof et al., 1993; O’Keefe, Movshon, 1998; Olavarria et al., 1992; Peterhans, von der Heydt, 1991; Sary et al., 1995), и у кошек (Leventhal et al., 1998; Mareschal, Baker, 1998; Zhou, Baker, 1994). Причем большинство ответов на модулированные стимулы были полосовыми (Mareschal, Baker, 1998). И регистрация реакций отдельных нейронов (Albright, 1992; Zhou, Baker, 1993), и исследования с использованием фМРТ (Larsson et al., 2006, 2010) продемонстрировали, что в обработку стимулов 2-го порядка включены не только стриарные, но и экстрастриарные области коры. Было также показано, что зрение 1-го и 2-го порядка имеет разные корковые генераторы вызванных потенциалов (Calvert, Manahilov, 2002). Начальная активация стриарных нейронов, в рецептивные поля которых попали градиенты яркости, происходит в первые 100 мс. После этого включается механизм внимания с использованием обратных связей (Mehta et al., 2000). Последующая реакция нейрона зависит от того, совпадает ли направление внимания с локализацией его рецептивного поля (Vidyasagar, 1998). После этого активность стриарных нейронов передается на нейроны V4, которые реагируют на стимулы, отобранные в ходе параллельной обработки (Motter, 1994). Их активность существенно зависит от привлечения к этим стимулам внимания.
Действительно, многие исследователи склоняются к тому, что важную роль в обработке стимулов 2-го порядка играет область V4. Так, исследование повреждений различных зон коры у обезьян продемонстрировало критическую роль V4 в различении текстур второго порядка (Merigan, 2000). А. Тильшер и Х. Ньюман на основании полученных ими результатов предположили, что нейроны V4 связаны с операцией пространственного группирования, определяющего вход внимания (Thielscher, Neumann, 2007).
Исследования с использованием зрительных ВП также указывают на то, что негативность рассогласования, связанная с привлечением внимания к целевому объекту (компонент N2pc), начинает формироваться примерно через 180–200 мс после его появления (Luck, 2012). То есть примерно в то время, когда происходит реактивация стриарных нейронов и активируются клетки V4. Примерно в это же время формируется компонент ВП, связанный с размерностью модуляции в стимуле второго порядка (Бабенко, Ермаков, 2015).
Как отмечает Дж. Тиус (Theeuwes, 1993), непроизвольное внимание автоматически привлекается к локализации, имеющей наиболее высокую активацию. При этом выбор полностью определяется физическими свойствами стимула. Объект в этой позиции автоматически выбирается независимо от намерения субъекта. Приоритеты выбора не могут быть изменены сверху. Сверху можно изменить только размер окна внимания. Поэтому первоначальный отбор полностью определяется стимулом (Theeuwes, 2010; Van der Stigchel et al., 2009).
ЭКСПЕРИМЕНТАЛЬНЫЕ ДОКАЗАТЕЛЬСТВА РОЛИ ФИЛЬТРОВ ВТОРОГО ПОРЯДКА, КАК ОКОН ВНИМАНИЯ
Выше мы говорили о возможном участии фильтров 2-го порядка в привлечении внимания теоретически, опираясь на косвенные экспериментальные данные. Ниже речь пойдет о прямой экспериментальной проверке этого предположения.
Для этого мы решили использовать в своих экспериментах “лицевые” стимулы. Это обусловлено тем, что лица – это своеобразный класс изображений. С одной стороны, они отличаются особой сложностью. С другой стороны, однотипность пространственной организации делает задачу их различения нетривиальной. Вместе с тем быстрота обнаружения и идентификации лиц, встроенных в сцены (Cauchoix et al., 2014; Crouzet, Thorpe, 2011), по всей видимости, подтверждает важную роль преаттентивных процессов обработки в их восприятии (VanRullen, 2006; Vuilleumier et al., 2001).
Мы начали с того, что создали градиентный оператор, имитирующий работу фильтра 2-го порядка, чувствительного к пространственной модуляции контраста. Это было сделано, чтобы выделять в изображениях области пространственной неоднородности и понять, полезна ли эта информация для категоризации лиц. Принимая в расчет сведения о том, что для восприятия лиц достаточно диапазона низких пространственных частот до 4 циклов на изображение (Ginsburg, 1980), в котором содержится около 75% всей энергии изображения (Guyader et al., 2017), мы задали частоту фильтрации для всех фильтров разного размера, равную 4 циклам на размер фильтра (центральная область рецептивного поля). Ширина частотной настройки всех фильтров составляла 1 октаву. Принцип работы оператора изложен в статье В. Бабенко с соавторами (Babenko et al., 2021).
Исходя из предположения, что чем выше неоднородность области изображения, тем она более информативна, необходимо было доказать, что более полезными в категоризации лиц являются области именно с наибольшей амплитудой модуляции контраста. В качестве исходных были использованы изображения мужских и женских лиц, европейской и азиатской расы, нейтральные и радостные. Из каждого исходного изображения, после обработки моделью, были созданы три стимула, состоящие только из областей с наибольшим, наименьшим и промежуточным приростом контраста. Лицевые стимулы предъявлялись в случайной последовательности в череде объектов других категорий, обработанных тем же способом. Задачей испытуемых было обнаружить лицо и дифференцировать его по полу, расе и выражению.
Следует отметить, что, поскольку фильтры 2-го порядка имеют разный размер, более крупный фильтр выделит лицо целиком, а меньшие по размеру – все более мелкие фрагменты. Соответственно, и пространственная частота (в циклах на градус), с которой описываются выделенные участки стимула, будет возрастать по мере уменьшения размера этих областей. Нами было задано, что, поскольку на каждом следующем шаге частота фильтрации возрастает вдвое, число выделяемых на этой частоте областей также увеличивается вдвое.
Учитывая существующие представления о целостном характере восприятия лиц, особый интерес представляли результаты распознавания, когда стимулы были сформированы одним фильтром, выделяющим изображение лица целиком. Оказалось, что этой информации достаточно, чтобы уверенно обнаруживать лица в череде других стимулов (выше 75%), но недостаточно, чтобы дифференцировать их по расе, полу и эмоции (Бабенко и др., 2021). Стало очевидным, что для решения этих задач необходимо расширение частотного диапазона за счет фильтров, выделяющих фрагменты изображения. Сравнение результатов, полученных для лицевых стимулов, сформированных из областей с разной амплитудой нелокального контраста, показало, что чем выше прирост контраста в выделенных областях, тем выше результат. И если при минимальном контрасте областей, из которых синтезирован стимул, процент правильных ответов был близок к нулю (допускался ответ “не знаю”), то при наибольшем он достигал более 60% (Babenko et al., 2021a). Полученные результаты показали, что более информативными областями лица являются области с большим приростом нелокального контраста.
Следующим шагом было доказательство того, что именно эти области изображения являются целями внимания. Мы использовали изображения реальных лиц с отображением 6 базовых эмоций по Экману и нейтральным выражением. Испытуемые в течение 700 мс рассматривали предъявленное изображение и определяли его экспрессию. Во время решения задачи регистрировались движения глаз. Оказалось, что начальные области фиксации взора совпадают с областями изображения, характеризующимися наибольшим приростом нелокального контраста (Babenko et al., 2021b). Интересно отметить, что области с наибольшим приростом контраста в диапазоне частот от 8 до 32 циклов на лицо, который, как известно, является наиболее информативным при восприятии лиц (Collin et al., 2006; Gold et al., 1999; Nasanen, 1999), локализуются в области глаз и рта. В этих же областях лица локализуется и большинство начальных фиксаций.
Таким образом, полученные результаты свидетельствуют о том, что целями внимания при решении задачи определения экспрессии могут быть области с наибольшим приростом суммарного контраста, которые преаттентивно выделяются зрительными механизмами второго порядка.
КАКАЯ ИНФОРМАЦИЯ МОЖЕТ БЫТЬ ПЕРЕДАНА ЧЕРЕЗ ФИЛЬТРЫ ВТОРОГО ПОРЯДКА?
Репрезентации, сохраняемые в рабочей, а затем и в долговременной памяти, формируются из информации, которая извлекается с уровня преаттентивной обработки. Что же это за информация, пропускаемая фильтрами 2-го порядка?
Самые крупные по величине своих рецептивных полей фильтры охватывают значительную часть поля зрения. Они способны выделять части зрительной сцены и могут передать информацию об их статистических свойствах. Меньшие по размеру фильтры выделяют в сцене отдельные объекты, целостное описание которых является низкочастотным (возможно, до 4 циклов на объект). Эта информация представляется важной с точки зрения формирования прототипов категорий. Более мелкие фильтры выделяют наиболее информативные фрагменты изображений. Чем меньше выделяемый фрагмент, тем с большим разрешением он описывается (при постоянном числе циклов на “окно” фильтра с уменьшением диаметра “окна” разрешение возрастает). Выделяемые фрагменты могут быть использованы вышележащими уровнями анализа для формирования признаков объектов. В результате создается возможность описания зрительного входа с разной степенью детализации: на уровне статистических свойств участка сцены, на уровне грубого описания отдельных объектов, а также на признаковом уровне, позволяющем детализировать объекты с необходимой точностью. При приближении или удалении объекта его описание остается неизменным, поскольку все фильтры имеют одинаковую организацию. Просто теперь как целый объект, так и его фрагменты будут выделяться фильтрами другого размера.
Данный принцип описания входного изображения позволяет представить любой объект в виде набора рекурсивно вложенных друг в друга репрезентаций. То, что в коре хранятся репрезентации как целостных образов, так и их фрагментов, подтверждают данные, полученные с использованием фМРТ. Например, для лиц было показано, что в одних областях коры (FFA) хранятся их целостные описания, тогда как в других (OFA и fSTS) – репрезентации их строительных блоков (глаза, нос, рот) (Liu et al., 2010; Maher et al., 2016; Nichols et al., 2010).
ПРОБЛЕМЫ И ПЕРСПЕКТИВЫ
Несложно заметить, что многие результаты, касающиеся свойств зрительных фильтров второго порядка, согласуясь в целом, иногда не совпадают в деталях (см. напр., (Westrick et al., 2013)). Скорее всего, объясняется это тем, что речь идет о механизмах, находящихся в так называемой серой зоне. Несомненно, легче изучать процессы, реализуемые на входе и на выходе сенсорной системы, для которых проще контролировать параметры входных сигналов и ответных реакций. А вот для механизмов, реализуемых на промежуточных стадиях обработки, бывает сложно учесть все возможные переменные. Вероятно, поэтому конкретный результат изучения тех или иных свойств механизмов второго порядка зависит от особенностей используемых стимулов и задач, которые стоят перед испытуемыми. Еще одна особенность этого этапа обработки состоит в том, что она находится на стыке стадий пред- и поствнимания. А значит, ее результаты зависят не только от восходящих, но и от нисходящих процессов. Взаимодействие этих потоков способно дать большое разнообразие эффектов. Тем не менее за последние без малого 30 лет сделан огромный шаг в изучении этих механизмов, и эти работы продолжаются.
В последние годы внимание исследователей было сосредоточено на изучении роли механизмов 2-го порядка в восприятии симметричных паттернов (Bellagarda et al., 2021), на исследовании закономерностей интеграции контуров 2-го порядка (Baldwin et al., 2020), на выяснении влияния старения на чувствительность к стимулам 2-го порядка, как показателя коркового дефицита (Reynaud et al., 2019), на оценке роли механизмов 2-го порядка в восприятии пространственной глубины (Asher, Hibbard, 2018), на изучении природы нелинейности на выходах фильтров 2-го порядка (Yi Gao, 2017). Предложен новый подход к машинному обучению, позволяющий моделировать зрительную обработку 2-го порядка (DiMattina, Baker, 2019).
Несмотря на то, что сегодня мы уже многое знаем о механизмах второго порядка, еще больше предстоит узнать. Сегодня мы подошли вплотную к пониманию их значения не только в восприятии текстур, но и всего многообразия окружающего мира. Однако требуется ответить еще на множество вопросов. Распространяются ли закономерности, установленные при восприятии лиц, на объекты других категорий? Какую роль в зрительном восприятии играют другие типы модуляций: ориентации и пространственной частоты градиентов яркости? Как используется информация из разных частотных диапазонов? Полученные в последние годы результаты указывают на то, что роль механизмов 2-го порядка в зрительном восприятии значительно шире, чем представлялось ранее.
ЗАКЛЮЧЕНИЕ
Если градиенты яркости разного контраста, ориентации и пространственной частоты, которые часто называют первичными признаками, являются своего рода “атомами” зрительного восприятия, то что представляют собой его “молекулы”? Вопрос очень сложный, и в попытках найти на него ответ предлагались разные варианты решения. Хотя по большому счету решений всего два – либо этим процессом управляют высшие уровни анализа, формируя “молекулы” с помощью внимания, либо это автоматический процесс, реализуемый по универсальному алгоритму. Конечно, и во втором варианте без внимания не обойтись. Но оно обращается к уже сформированным “молекулам” и объединяет их затем в разных сочетаниях. Первый вариант удобен своей “гибкостью” и долгое время рассматривался как приоритетный. Однако накапливалось все больше экспериментальных данных, которые сложно было объяснить без привлечения дополнительного преаттентивного механизма группирования. И на рубеже 80-х и 90-х гг. появляются теоретические концепции, которые предлагают вариант решения этого вопроса. Все они сходились на том, что локальная зрительная информация, выделяемая механизмами 1-го порядка, автоматически группируется на следующем этапе преаттентивной обработки механизмами 2-го порядка. Эта модель оказалась настолько удобной для интерпретации многих необъяснимых эффектов, что ей даже дали шутливое название “модель заднего кармана”, подчеркивая, что, если возникают затруднения с объяснением некоторых фактов, данная модель окажется весьма кстати. Во второй половине 90-х и начале 2000-х годов наблюдается всплеск исследований “зрительных механизмов второго порядка”. Обсуждаются вопросы их пространственной организации, независимость и принципы взаимодействия фильтров 1-го и 2-го порядка, настроечные характеристики и многоканальность механизмов группирования, избирательность этих механизмов к размерности модуляции. Однако к концу 2000-х годов интерес к этой проблеме стал угасать. Эти механизмы казались предназначенными для решения лишь одной зрительной задачи – разделения текстур, где они действительно показали себя в высшей степени эффективно. Задачи, несомненно, важной, но весьма специфичной. Интерес к операциям преаттентивного пространственного группирования вновь возрос, когда появились предположения о более важной роли этих механизмов в зрительном восприятии. Поскольку пространственные модуляции градиентов яркости содержатся не только в текстурах, но и в изображениях любых реальных объектов, фильтры 2-го порядка могут использоваться для их сегментации и выделения областей интереса. Появились первые экспериментальные работы, указывающие на возможную роль фильтров 2-го порядка в восходящем управлении вниманием. И вполне может оказаться, что зрительные механизмы, которые ранее представлялись своего рода функциональным довеском, на самом деле являются весьма важным звеном в цепи процессов обработки зрительной информации.
Список литературы
Бабенко В.В. Новый подход к вопросу о механизмах зрительного восприятия. Проблемы нейрокибернетики. Ростов-на-Дону: из-во РГУ, 1989. 10–11 с.
Бабенко В.В., Ермаков П.Н., Божинская М.А. Соотношение пространственно-частотных настроек зрительных фильтров первого и второго порядка. Психол. журн. 2010. 31 (2): 48–57.
Бабенко В.В., Кульба С.Н. Модель механизма зрительной сегментации. Сенс. сист. 2002. 16 (3): 179–189.
Бабенко В.В., Кураев Г.А., Кульба С.Н. Механизмы повышения контрастной чувствительности при увеличении протяженности синусоидальной решетки. Сенс. сист. 2001. 15 (1): 3–10.
Бабенко В.В., Явна Д.В., Анохина П.В. Роль нелокального контраста в обнаружении лиц и идентификации эмоций. Первый Национальный конгресс по когнитивным исследованиям, искусственному интеллекту и нейроинформатике. Девятая международная конференция по когнитивной науке. Отв. ред. В.Л. Ушаков, И.И. Русак. М.: НИЯУ МИФИ, 2021. Часть 2. 414–416 с.
Бабенко В.В., Явна Д.В., Родионов Е.Г. Вклад различных пространственных модуляций градиентов яркости в управление зрительным вниманием. Журн. высш. нервн. деят. им. И.П. Павлова. 2020. 70 (2): 183–193.
Шевелёв И.А. Нейроны-детекторы зрительной коры. М: Наука, 2010. 183 с.
Шелепин Ю.Е., Макаров Ф.Н., Трифонов М.И. Пирамидальная организация первичной зрительной системы человека. Проблемы нейрокибернетики. Ростов-на-Дону: из-во РГУ, 1989. 186–187 с.
Явна Д.В. Психофизиологические особенности зрительного восприятия пространственно модулированных признаков. Автореф. дис. … канд. психол. наук, Ростов-на-Дону: ЮФУ, 2012. 23 с.
Яковлев А.Ю., Тюрина Н.А., Уточкин И.С. Зрительное восприятие ансамблей: обзор исследований. Российский журнал когнитивной науки. 2020. 7 (3): 4–24.
Açık F., Onat S., Schumann F., Einhäuser W., König P. Effects of luminance contrast and its modifications on fixation behavior during free viewing of images from different categories. Vis. Res. 2009. 49 (12): 1541–1553.
Albrecht D.G., Hamilton D.B. Striate cortex of monkey and cat: contrast response function. J Neurophysiol. 1982. 48 (1): 217–237.
Albright T.D. Form-cue invariant motion processing in primate visual cortex. Science. 1992. 255 (5048): 1141–1143.
Allard R., Faubert J. Double dissociation between first- and second-order processing. Vis. Res. 2007. 47 (9): 1129–1141.
Arsenault A.S., Wilkinson F., Kingdom F.A. Modulation frequency and orientation tuning of second-order texture mechanisms. J Opt. Soc. Am. A. 1999. 16 (3): 427–435.
Asher J.M., Hibbard P.B. First- and second-order contributions to depth perception in anti-correlated random dot stereograms. Sci. Rep. 2018. 8: Art. 14120.
Babenko V.V. A model of an object-form description mechanism. Perception. 1991. 20 (1): 70.
Babenko V., Ermakov P. Specificity of brain reactions to second-order visual stimuli. Vis. Neurosci. 2015. 32: Art. e011.
Babenko V., Yavna D. Specificity of the visual second-order mechanisms. Perception. 2008. Suppl.: 78.
Babenko V., Yavna D. Second-order visual mechanisms are fixed filters. Perception. 2010. Suppl.: 194.
Babenko V.V., Yavna D.V., Ermakov P.N., Anokhina P.V. Nonlocal contrast calculated by the second order visual mechanisms and its significance in identifying facial emotions. F1000Res. 2021a. 10: 274.
Babenko V., Yavna D., Vorobyeva E., Denisova E., Ermakov P., Kovsh E. Relationship Between Facial Areas With the Greatest Increase in Non-local Contrast and Gaze Fixations in Recognizing Emotional Expressions. Int. J Cognit. Res. in Science, Engineering and Education. 2021b. 9 (3): 359–368.
Baek J., Chong S.C. Distributed attention model of perceptual averaging. Attent. Percept. Psychophys. 2020. 82 (1): 63–79.
Bahcall D.O., Kowler E. Attentional interference at small spatial separations. Vis. Res. 1999. 39 (1): 71–86.
Baldwin A.S., Kenwood M., Hess R.F. Integration of contours defined by second-order contrast-modulation of texture. Vis. Res. 2020. 176: 1–15.
Barlow H.B., Kaushal T.P., Hawken M., Parker A.J. Human contrast discrimination and the threshold of cortical neurons. J Opt. Soc. Am. A. 1987. 4 (12): 2366–2371.
Bellagarda C.A., Dickinson J.E., Bell J., Badcock D.R. The temporal integration windows for visual mirror symmetry. Vis. Res. 2021. 188: 184–192.
Ben-Shahar O., Scholl B.J., Zucker S.W. Attention, segregation, and textons: Bridging the gap between object-based attention and texton-based segregation. Vis. Res. 2007. 47 (6): 845–860.
Bergen J.R. Theories of visual texture perception. Vision and visual dysfunction. Ed. Regan D. New York: MacMillan, 1991. V. 10B. 114–134 pp.
Bruce N.D.B. Features that draw visual attention: an information theoretic perspective. Neurocomputing. 2005. 65–66: 125–133.
Bruce N.D., Tsotsos J.K. Saliency based on information maximization. Adv. Neural Inf. Process. Syst. 2006. 18: 5–162.
Caelli T., Julesz B. On perceptual analyzers underlying visual texture discrimination. Biol Cybern. 1978. 28 (1): 167–175.
Campbell F.W., Robson J.G. Application of Fourier analysis to the visibility of gratings. J Physiol. 1968. 197 (3): 551–566.
Campbell F.W., Howell E.R., Johnstone J.R. A comparison of threshold and suprathreshold appearance of gratings with components in the low and high spatial frequency range. J Physiol. 1978. 284 (1): 193–201.
Calvert J., Manahilov V., Simpson W.A., Parker D.M. Human cortical responses to contrast modulations of visual noise. Vis. Res. 2005. 45 (17): 2218–2230.
Cannon M.W., Fullenkamp S.C. Spatial interactions in apparent contrast: inhibitory effects among grating patterns of different spatial frequencies, spatial positions and orientations. Vis. Res. 1991. 31 (11): 1985–1998.
Castiello U., Umilta C. Size of the attentional focus and efficiency of processing. Acta Psychol. (Amst). 1990. 73 (3): 195–209.
Cauchoix M., Barragan-Jason G., Serre T., Barbeau E.J. The Neural Dynamics of Face Detection in the Wild Revealed by MVPA. J Neurosci. 2014. 34 (3): 846–854.
Cave R.R., Kosslin S.M. Varieties of size-specific visual selection. J Exp. Psychol. Gen. 1989. 118 (2): 148–164.
Cave K.R., Pashler H. Visual selection mediated by location: Selecting successive visual objects. Percept. Psychophys. 1995. 57 (4): 421–432.
Chaudhuri A., Albright T.D. Neuronal responses to edges defined by luminance vs. temporal texture in macaque area V1. Vis. Neurosci. 1997. 14: 949–962.
Chen Z. Not all features are created equal: Processing asymmetries between location and object features. Vis. Res. 2009. 49 (11): 1481–1491.
Chubb C., Sperling G. Drift-balanced random stimuli: a general basis for studying non-Fourier motion perception. J Opt. Soc. Am. A. 1988. 5 (11): 1986–2007.
Collin C.A., Therrien M., Martin C., Rainville S. Spatial frequency thresholds for face recognition when comparison faces are filtered and unfiltered. Percept. Psychophys. 2006. 68 (6): 879–889.
Crouzet S.M., Thorpe S.J. Low level cues and ultra-fast face detection. Frontiers in Psychology. 2011. 2: Art. 342.
Cruickshank A.G., Schofield A.J. Transfer of tilt after-effects between second-order cues. Spatial Vision. 2005. 18 (4): 379–397.
Dakin S.C., Mareschal I. Sensitivity to contrast modulation depends on carrier spatial frequency and orientation. Vis. Res. 2000. 40 (3): 311–329.
Dean A.F. The relationship between response amplitude and contrast for cat striate cortical neurones. J Physiol. 1981. 318: 413–427.
DiMattina C., Baker C.L.Jr. Modeling second-order boundary perception: A machine learning approach. PLoS Comput. Biol. 2019. 15 (3): Art. e1006829.
DiMattina C., Baker C.L.Jr. Segmenting surface boundaries using luminance cues Scientific Reports. 2021. 11: Art. 10074.
Ellemberg D., Allen H.A., Hess R.F. Second-order spatial frequency and orientation channels in human vision. Vis. Res. 2006. 46 (17): 2798–2803.
Eriksen C.W., Yeh Y.Y. Allocation of attention in the visual field. J Exp. Psychol. Hum. Percept. Perform. 1985. 11 (5): 583–597.
Field D.J., Hayes A., Hess R.F. Contour integration by the human visual system: Evidence for a local ‘association field’. Vis. Res. 1993. 33: 173–193.
Fuchs I., Ansorge U., Redies C., Leder H. Salience in Paintings: Bottom-Up Influences on Eye Fixations. Cognitive Computation. 2011. 3 (1): 25–36.
Gao Yi . On the processing of second-order information in normal and amblyopic vision. A thesis submitted to McGill University in partial fulfillment of the requirements of the degree of doctor of philosophy. 2017.
Gao D., Han S., Vasconcelos N. Discriminant saliency, the detection of suspicious coincidences, and applications to visual recognition. IEEE Trans. Pattern Anal. Mach. Intell. 2009. 31 (6): 989–1005.
Gao D., Vasconcelos N. Bottom-up saliency is a discriminant process. IEEE Int’l Conf. Computer Vision (ICCV). 2007: 6 pp.
Geesaman B.J., Anderson R.A. The analysis of complex motion patterns by form0cue invariant MSTd neurons. J Neurosci. 1996. 16: 4716–4732.
Ginsburg A.P. Specifying relevant spatial information for image evaluation and display design: An explanation of how we see certain objects. Proceedings of the SID. 1980. 21: 219–227.
Gold J., Bennett P.J., Sekule A.B. Identification of band-pass filtered letters and faces by human and ideal observers. Vis. Res. 1999. 39 (21): 3537–3560.
Gong M., Olzak L. Perceptual learning transfer between first- and second-order fine orientation discriminations. J Vision. 2014. 14: Art. 77.
Goolkasian P. Size scaling and spatial factors in visual attention. Am. J Psychol. 1997. 110 (3): 397–415.
Graham N. Non-linearities in texture segregation. CIBA Foundation Symposium. Eds: Bock G.R., Goode J.A. New York: Wiley, 1994. 184: 309–329.
Graham N. Non-linearities in texture segregation. CIBA Foundation Symposium. Eds: Bock G.R., Goode J.A. New York: Wiley, 1994. 184: discussion 323–338.
Graham N., Beck J., Sutter A. Nonlinear processes in spatial-frequency channel models of perceived texture segregation. Vis. Res. 1992. 32: 719–743.
Graham N., Sutter A. Normalization: contrast-gain control in simple (Fourier) and complex (non-Fourier) pathways of pattern vision. Vis. Res. 2000. 40 (20): 2737–2761.
Graham N., Sutter A., Venkatesan C. Spatial-frequency- and orientation-selectivity of simple and complex channels in region segregation. Vis. Res. 1993. 33 (14): 1893–1911.
Graham N., Wolfson S.S. A note about preferred orientations at the first and second stages of complex (second-order) texture channels. J Opt. Soc. Am. A. 2001. 18 (9): 2273- -2281.
Grosof D.H., Shapley R.M., Hawken M.J. Macaque V1 neurons can signal ‘illusory’ contours. Nature. 1993. 365: 550–552.
Guyader N., Chauvin A., Boucart M., Peyrin C. Do low spatial frequencies explain the extremely fast saccades towards human faces? Vis. Res. 2017. 133: 100–111.
Han B., Tcheang L., Walsh V., Gao X. A Novel Feature Combination Methods for Saliency-Based Visual Attention. Natural Computation, ICNC '09. 2009. 18–22 pp.
He Z.J., Nakayama K. Surface versus features in visual search. Nature. 1992. 359: 231–233.
Henning G.B., Hertz B.G., Broadbent D.E. Some experiments bearing on the hypothesis that the visual system analyzes spatial patterns in independent bands of spatial frequency. Vis. Res. 1975. 15: 887–899.
Hess R.F., Holliday I.E. The coding of spatial position by the human visual system: Effects of spatial scale and contrast. Vis. Res. 1992. 32: 1085–2097.
Hoekstra J., van der Goot D.P.J., van der Brink G., Bilzen F.A. The influence of the number of cycles upon the visual contrast threshold for spatial sine wave patterns. Vis. Res. 1974. 14: 365–368.
Hou W., Gao X., Tao D., Li X. Visual saliency detection using information divergence. Pattern Recognition. 2013. 46 (10): 2658–2669.
Huang P.C., Chen C.C. A comparison of pedestal effects in first- and second-order patterns. J Vision. 2014. 14 (1): Art. 9.
Hubel D.H., Wiesel T.N. Receptive fields of single neurons in the cat’s striate cortex. J Physiol. 1959. 148: 574–591.
Hubel D.H., Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J Physiol. 1962. 160: 106–154.
Hunt C., Meinhardt G. Synergy of spatial frequency and orientation bandwidth in texture segregation. J Vision. 2021. 21 (2): Art. 5.
Itti L., Koch C. Computational modeling of visual attention. Nature Reviews Neurosci. 2001. 2 (3): 194–203.
Itti L., Koch C., Niebur E. A model of saliency-based visual attention for rapid scene analysis. IEEE Trans. Pattern Anal. Mach. Intell. 1998. 20 (11): 1254–1259.
Jamar J.H., Koenderink J.J. Sine-wave gratings: scale invariance and spatial integration at suprathreshold contrast. Vis. Res. 1983. 23 (8): 805–810.
Jamar J.H., Koenderink J.J. Contrast detection and detection of contrast modulation for noise gratings. Vis. Res. 1985. 25 (4): 511–521.
Joseph J.S., Optican L.M. Involuntary attentional shifts due to orientation differences. Percept. Psychopchys. 1996. 58 (5): 651–665.
Julesz B. Spatial nonlinearities in the instantaneous perception of textures with identical power spectra. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980. 290 (1038): 83–94.
Julesz B., Gilbert E.N., Shepp L.A., Frisch H.L. Inability of humans to discriminate between visual textures that agree in second-order statistics-revisited. Perception. 1973. 2: 391–405.
Julesz B., Gilbert E.N., Victor J.D. Visual discrimination of textures with identical third order statistics. Biol. Cybern. 1978. 31: 137–140.
Julesz B. Textons, the elements of texture perception and their interactions. Nature. 1981. 290: 91–97.
Kingdom F.A., Keeble D.R. A linear systems approach to the detection of both abrupt and smooth spatial variations in orientation-defined textures. Vis. Res. 1996. 36 (3): 409–420.
Kingdom F.A., Keeble D., Moulden B. Sensitivity to orientation modulation in micropattern-based textures. Vis. Res. 1995. 35 (1): 79–91.
Kingdom F.A., Keeble T. On the mechanism for scale invariance in orientation-defined textures. Vis. Res. 1999. 39 (8): 1477–1489.
Kingdom F.A., Prins N., Hayes A. Mechanism independence for texture-modulation detection is consistent with a filter-rectify-filter mechanism. Vis. Neurosci. 2003. 20: 65–76.
King-Smith P.E., Kulikowski J.J. The detection of gratings by independent activation of line detectors. J Physiol. 1975. 247 (2): 237–271.
Kwan L., Regan D. Orientation-tuned spatial filters for texture-defined form. Vis. Res. 1998. 38 (24): 3849–3855.
LaBerge D. Spatial extent of attention to letters and words. J Exp. Psychol. Hum. Percept. Perform. 1983. 9 (3): 371–379.
Landy M.S., Bergen J.R. Texture segregation and orientation gradient. Vis. Res. 1991. 31 (4): 679–691.
Landy M.S., Henry C.A. Critical-band masking estimation of 2nd-order filter properties. Perception. 2007. 36 (Suppl.): 61.
Landy M.S., Oruç P. Properties of second-order spatial frequency channels. Vis. Res. 2002. 42 (19): 2311–2329.
Langley K., Fleet D.J., Hibbard P.B. Linear filtering precedes nonlinear processing in early vision. Curr. Biol. 1996. 6 (7): 891–896.
Larsson J., Heeger D.J., Landy M.S. Orientation selectivity of motion-boundary responses in human visual cortex. J Neurophysiol. 2010. 104 (6): 2940–2950.
Larsson J., Landy M.S., Heeger D.J. Orientation-selective adaptation to first- and second-order patterns in human visual cortex. J Neurophysiol. 2006. 95 (2): 862–881.
Legge G.E., Foley J.M. Contrast masking in human vision. J Opt. Soc. Am. 1980. 70 (12): 1458–1471.
Leventhal A.G., Wang Y., Schmolesky M.T., Zhou Y. Neural correlates of boundary perception. Vis. Neurosci. 1998. 15: 1107–1118.
Levin D.T., Takarae Y., Miner A.G., Keil F. Efficient visual search by category: specifying the features that mark the difference between artifacts and animals in preattentive vision. Percept. Psychophys. 2001. 63 (4): 676–697.
Li G., Yao Z., Wang Z., Yuan N., Talebi V., Tan J., Baker C.L.Jr. Form-cue invariant second-order neuronal responses to contrast modulation in primate area v2. J Neurosci. 2014. 34 (36): 12 081–12 092.
Liu J., Harris A., Kanwisher N. Perception of Face Parts and Face Configurations: An fMRI Study. J Cognit. Neurosci. 2010. 22 (1): 203–211.
Luck S.J. Electrophysiological correlates of the focusing of attention within complex visual scenes: N2pc and related ERP components. The Oxford handbook of event-related potential components. Oxford: Oxford University Press, 2012. 329–360 pp.
Maffei L., Fiorentini A. The visual cortex as a spatial frequency analyser. Vis. Res. 1973. 13 (7): 1255–1267.
Maher S., Ekstrom T., Tong Y., Nickerson L.D., Frederick B., Chen Y. Greater sensitivity of the cortical face processing system to perceptually-equated face detection. Brain Res. 2016. 1631: 13–21.
Mareschal I., Baker C.L.Jr. Temporal and spatial response to second-order stimuli in cat area 18. J Neurophysiol. 1998a. 80 (6): 2811–2823.
Mareschal I., Baker C.L.Jr. A cortical locus for the processing of contrast-defined contours. Nat Neurosci. 1998b. 1 (2): 150–154.
Mehta A.D., Ulbert I., Schroeder C.E. Intermodal Selective Attention in Monkeys. II: Physiological Mechanisms of Modulation. Cerebral Cortex. 2000. 10 (4): 359–370.
Merigan W.H. Cortical area V4 is critical for certain texture discriminations, but this effect is not dependent on attention. Vis. Neurosci. 2000. 17 (6): 949–958.
Mnih V., Heess N., Graves A., Kavukcuoglu K. Recurrent Models of Visual Attention. Advances in Neural Information Processing Systems 27 (NIPS 2014). Eds: Ghahramani Z., Welling M., Cortes C., Lawrence N.D., Weinberger K.Q. Curran Associates, Inc., 2014. 2204–2212 pp.
Morgan M.J., Glennerster A. Efficiency of locating centers of dot-clusters by human observers. Vis. Res. 1991. 31: 2075–2083.
Motoyoshi I., Nishida S. Cross-orientation summation in texture segregation. Vis. Res. 2004. 44 (22): 2567–2576.
Motter B.C. Neural correlates of attentive selection for color or luminance in extrastriate area V4. J Neurosci. 1994. 14 (4): 2178–2189.
Nasanen R. Spatial frequency bandwidth used in the recognition of facial images. Vis. Res. 1999. 39 (23): 3824–3833.
Nichols D.F., Betts L.R., Wilson H.R. Decoding of faces and face components in face-sensitive human visual cortex. Front. Psychol. 2010. 1: Art. 28.
Nishida S., Ledgeway T., Edwards M. Dual multiple-scale processing for motion in the human visual system. Vis. Res. 1997. 37 (19): 2685–2698.
Ohzawa I., Sclar G., Freeman R.D. Contrast gain control in the cat’s visual system. J Neurophysiol. 1985. 54 (3): 651–667.
O’Keefe L.P., Movshon J.A. Processing of first- and second-order motion signals by neurons in area MT of the macaque monkey. Vis. Neurosci. 1998. 15: 305–317.
Olavarria J.F., DeYoe E.A., Knierim J.J., Fox J.M., Van Essen D.C. Neural responses to visual texture patterns in middle temporal area of the macaque monkey. J Neurosci. 1992. 68: 164–181.
Olzak L.A., Thomas J.P. Configural effects constrain Fourier models of pattern discrimination. Vis. Res. 1992. 32 (10): 1885–1898.
Parkhurst D., Law K., Niebur E. Modeling the role of salience in the allocation of overt visual attention. Vis. Res. 2002. 42 (1): 107–123.
Perazzi F., Krahenbuhl P., Pritch Y., Hornung A. Saliency Filters: Contrast Based Filtering for Salient Region Detection. Computer Vision and Pattern Recognition (CVPR), IEEE Conference on Computer Vision and Pattern Recognition. 2012. 733–740 pp.
Peterhans E., von der Heydt R. Subjective contours-bridging the gap between psychophysics and physiology. Trends in Neurosci. 1991. 14: 112–119.
Reynaud A., Hess R.F. Properties of spatial channels underlying the detection of orientation-modulations. Exp. Brain Res. 2012. 220 (2): 135–145.
Reynaud A., Tang Y., Zhou Y., Hess R.F. Second-order visual sensitivity in the aging population. Aging Clin. Exp. Res. 2019. 31: 705–716.
Rubenstein B.S., Sagi D. Effects of foreground scale in texture discrimination tasks: performance is size, shape, and content specific. Spat. Vis. 1993. 7 (4): 293–310.
Saarinen J., Rovamo J., Virsu V. Texture discrimination at different eccentricities. J Opt. Soc. Amer. A. 1987. 4 (8): 1699–1703.
Sagi D. Detection of an orientation singularity in Gabor textures: effect of signal density and spatial frequency. Vis. Res. 1990. 30 (9): 1377–1388.
Sakai K., Finkel L.H. Characterization of the spatial-frequency spectrum in the perception of shape from texture. J Opt. Soc. Amer. 1995. 12: 1208–1224.
Sary G., Vogels R., Kovacs G., Orban G.A. Responses of monkey inferior temporal neurons to luminance-, motion-, and texture defined gratings. J Neurophysiol. 1995. 73: 1341–1354.
Savoy R.L., McCann J.J. Visibility of low-spatial-frequency sine-wave targets: Dependence on number of cycles. J Opt. Soc. Amer. 1975. 65 (3): 343–350.
Schofield A.J., Georgeson M.A. Sensitivity to modulations of luminance and contrast in visual white noise: separate mechanisms with similar behavior. Vis. Res. 1999. 39 (16): 2697–2716.
Schofield A.J., Georgeson M.A. Sensitivity to contrast modulation: the spatial frequency dependence of second-order vision. Vis. Res. 2003. 43 (3): 243–259.
Schofield A.J., Ledgeway T., Hutchinson C.V. Asymmetric transfer of the dynamic motion aftereffect between first- and second-order cues and among different second-order cues. J Vision. 2007. 7 (8): Art. 1.
Schofield A.J., Yates T.A. Interactions between orientation and contrast modulations suggest limited cross-cue linkage. Perception. 2005. 34 (7): 769–792.
Shih S.-I., Sperling G. Is there feature-based attentional selection in visual search. J Exp. Psychol.: Hum. Percept. Perform. 1996. 22 (3): 758–779.
Solomon J.A., Sperling G. 1st- and 2nd-order motion and texture resolution in central and peripheral vision. Vis. Res. 1995. 35 (1): 59–64.
Steinman B.A., Steinman S.B., Lehmkuhle S. Visual attention mechanisms show a center-surround organization. Vis. Res. 1995. 35 (13): 1859–1869.
Sun Y., Fisher R. Hierarchical Selectivity for Object-Based Visual Attention. Int. Workshop on Biologically Motivated Computer Vision. BMCV 2002: Biologically Motivated Computer Vision. 2002. 427–438 pp.
Sun P., Schofield A.J. The efficacy of local luminance amplitude in disambiguating the origin of luminance signals depends on carrier frequency: Further evidence for the active role of second-order vision in layer decomposition. Vis. Res. 2011. 51 (5): 496–507.
Sutter A., Beck J., Graham N. Contrast and spatial variables in texture segregation: testing a simple spatial-frequency channels model. Percept. Psychophys. 1989. 46 (4): 312–332.
Sutter A., Sperling G., Chubb C. Measuring the spatial frequency selectivity of second-order texture mechanisms. Vis. Res. 1995. 35 (7): 915–924.
Tannazzo T., Kurylo D.D., Bukhari F. Perceptual grouping across eccentricity. Vis. Res. 2014. 103: 101–108.
't Hart B.M., Schmidt H.C.E.F., Roth C., Einhäuser W. Fixations on objects in natural scenes: dissociating importance from salience. Front. Psychol. 2013. 4: Art. 455.
Theeuwes J. Visual selective attention: a theoretical analysis. Acta Psychol. (Amst). 1993. 93 (2): 93–154.
Theeuwes J. Top–down and bottom–up control of visual selection. Acta Psychologica. 2010. 135 (2): 77–99.
Thielscher A., Neumann H. A computational model to link psychophysics and cortical cell activation patterns in human texture processing. J Comput. Neurosci. 2007. 22 (3): 255–282.
Treisman A.M., Gelade G. A feature-integration theory of attention. Cognit. Psychol. 1980. 12 (1): 97–136.
Treisman A., Souther J. Search asymmetry: a diagnostic for preattentive processing of separable features. J Exp. Psychol. Gen. 1985. 114 (3): 285–310.
Usher M., Bonneh Y., Sagi D., Herrmann M. Mechanisms for spatial integration in visual detection: a model based on lateral interactions. Spatial Vis. 1999. 12 (2): 187–209.
Vakrou C., Whitaker D., McGraw P.V. Extrafoveal viewing reveals the nature of second-order human vision. J Vision. 2007. 7 (14): Art. 13.
Van der Stigchel S., Belopolsky A.V., Peters J.C., Wijnen J.G., Meeter M., Theeuwes J. The limits of top-down control of visual attention. Acta Psychologica. 2009. 132 (3): 201–212.
VanRullen R. On second glance: Still no high-level pop-out effect for faces. Vis. Res. 2006. 46 (18): 3017–3027.
Vidyasagar T.R. Gating of neuronal responses in macaque primary visual cortex by an attentional spotlight. Neuroreport. 1998. 22 (9): 1947–1952.
Virsu V., Rovamo J. Visual resolution, contrast sensitivity, and the cortical magnification factor. Exp. Brain Res. 1979. 37 (3): 475–494.
Vuilleumier P., Sagiv N., Hazeltine E., Poldrack R.A., Swick D., Rafal R.D., Gabrieli J.D. Neural fate of seen and unseen faces in visuospatial neglect: A combined event-related functional MRI and event-related potential study. Proceedings of the National Academy of Science USA. 2001. 98 (6): 3495–3500.
Wang H.X., Heeger D.J., Landy M.S. Responses to second-order texture modulations undergo surround suppression. Vis. Res. 2012. 62: 192–200.
Wenderoth P., Clifford C.W.G., Wyatt A.M. Hierarchy of spatial interactions in the processing of contrast-defined contours. J Opt. Soc. Amer. A. 2001. 18 (9): 2190–2196.
Westrick Z.M., Henry C.A., Landy M.S. Inconsistent channel bandwidth estimates suggest winner-take-all nonlinearity in second-order vision. Vis. Res. 2013. 81 (5): 58–68.
Wilson H.R., Richards W.A. Curvature and separation discrimination at texture boundaries. J Opt. Soc. Amer. A. 1992. 9: 1653–1662.
Wolfe J.M. Asymmetries in visual search: An introduction. Percept. Psychophys. 2001. 63 (3): 381–389.
Wolfson S.S., Landy M.S. Discrimination of orientation-defined texture edges. Vis. Res. 1995. 35 (20): 2863–2877.
Zhou Y.X., Baker C.L.Jr. A processing stream in mammalian visual cortex neurons for non-Fourier responses. Science. 1993. 261 (5117): 98–101.
Zhou, Y.X., Baker C.L.Jr. Envelope-responsive neurons in areas 17 and 18 of cat. J Neurophysiol. 1994. 72: 2134–2150.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова