

Лесоведение, 2019, № 1, стр. 38-48
Разложение коры в составе порубочных остатков после сплошных рубок в среднетаежных лесах
Е. А. Капица a, b, *, Е. В. Шорохова a, b, c, И. В. Ромашкин b, Н. А. Галибина b, К. М. Никерова b, И. А. Казарцев a, b, d
a Санкт-Петербургский государственный лесотехнический университет
194021 Санкт-Петербург, Институтский пер., 5,
Россия
b Карельский научный центр Российской академии наук
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, 11,
Россия
c Институт Природных Ресурсов Финляндии (Луке)
FI-00790 Хельсинки, ул. Виикинкаари, 4, Финляндия
d Всероссийский научно-исследовательский институт защиты растений
196608 Санкт-Петербург, Пушкин, шоссе Подбельского, 3, Россия
* E-mail: kapitsa@list.ru
Поступила в редакцию 16.11.2017
После доработки 11.12.2017
Принята к публикации 08.10.2018
Аннотация
Роль коры как фракции фитомассы в биологическом круговороте веществ изучена недостаточно. В работе исследована шестилетняя динамика физических и химических свойств коры, оставшейся после сплошной рубки леса. Масса коры в составе порубочных остатков составила 20.8 м3 га–1, или пятую часть от запаса крупных древесных остатков, который варьировал от 105 до 127 м3 га–1. Начальная удельная масса коры составила в среднем 0.601 г cм–2 для осины, 0.346 г cм–2 для сосны, 0.446 г cм–2 для березы и 0.233 г cм–2 для ели. Начальная плотность коры варьировала от 0.51 до 0.71 г см–3 для осины, от 0.56 до 0.80 г см–3 для березы, от 0.39 до 0.64 г см–3 для сосны и от 0.60 до 0.73 г см–3 для ели; начальные концентрации азота и углерода – от 0.23 до 0.53% и от 42 до 48%, соответственно. Изменение во времени удельной массы коры, процента флоэмы и толщины коры зависело от древесной породы. Максимальная скорость потери массы отмечена у еловой коры, тогда как максимальная скорость потери флоэмы – у березовой коры. За 6 лет, прошедших после рубки, удельная масса коры уменьшилась на 32–38%, плотность – на 10–17%, процент флоэмы – на 78–99%. Концентрации азота и углерода в коре не изменились. Сохранение слоя коры на поверхности почвы после рубки леса частично компенсирует потери азота вследствие вывоза срубленной древесины.
Кора представляет собой совокупность тканей, выполняющих, главным образом, защитные функции от: 1) неблагоприятных климатических условий, предохраняя ткани дерева от чрезмерного перегрева летом и охлаждения зимой; 2) биологических агентов, таких как дереворазрушающие грибы, бактерии и вирусы; 3) механических повреждений и атмосферного загрязнения, длительное время оберегая дерево от воздействия поллютантов (Sawidis et al., 2011; Dossa et al., 2016). После отмирания дерева все фракции фитомассы, в том числе и кора, вовлекаются в биологический круговорот. Роль коры в возврате минеральных элементов в лесную экосистему до конца неясна в связи с ее сложным анатомическим строением. Отличия анатомического строения и минерального состава коры и древесины (Поздняков и др., 1974; Казимиров, Морозова, 1975; Лотова, 1999) не позволяют объединить эти фракции при изучении биологического круговорота. Различия луба и корки по плотности (Полубояринов, Сорокин, 1997) делают процесс деструкции неравномерным. Луб как наиболее доступный и питательный пищевой ресурс активно поедают насекомые и разлагают микроорганизмы, в то время как богатая смолами и таннинами корка длительное время сохраняет свою структуру (Рипачек, 1967; Parameswaran et al., 1976).
Ранее для таежных лесов были определены скорости потери массы и объема прикрепленной к стволу коры (Tarasov, Birdsey, 2001; Шорохова, Капица, 2007), а также образцов коры, выложенных на поверхность почвы и зарытых в подстилку (Wilhelm, 1976) или почву (Kuhlman, 1970), а также выявлены факторы, регулирующие эти процессы, к которым относятся древесная порода (Shorohova, Kapitsa, 2014), секция ствола (Шорохова, Капица, 2007), лесорастительные условия, воздействие регулируемого пала (Shorohova et al., 2008a, b). На вырубках рассчитаны константы разложения коры, прикрепленной к пням (Shorohova et al., 2008a). Вместе с тем, разложение коры, оставшейся после сплошной рубки леса на поверхности почвы, не изучено. Количественно не оценены объем и масса коры как фракции подстилки. Использование констант, характеризующих процесс разложения коры, прикрепленной к пням, невозможно в силу различий микроклиматических условий (Дымов, Старцев, 2016; Lukina et al., 2017).
Химический состав коры определяется породой (Поздняков и др., 1974), почвенными условиями (Gustafsson, Eriksson, 1995; Орлова и др., 2016), наличием в воздухе загрязнителей (Cekstere et al., 2015), а также временем и интенсивностью процесса разложения (Palviainen et al., 2010). Содержание азота и углерода в коре, как правило, выше чем в древесине (Palviainen et al., 2010). Установлено, что по мере деструкции древесины и прикрепленной к пню коры концентрация азота (на единицу сухой массы) в первые 10 лет увеличивается, что связано с низкой скоростью его вымывания, а также с фиксацией азота из атмосферы азотфиксирующими бактериями или его переносом филаментными грибами из других природных резервуаров (например, из почвы) в зону биодеструкции (разрушения). По истечении периода освоения коры и древесины происходит плавное снижение концентрации азота. Содержание углерода в коре (на единицу сухой массы), прикрепленной к пню, практически не меняется (Palviainen et al., 2010). Однако динамика углерода и азота в коре как фракции подстилки остается неизученной.
Несмотря на свою высокую биологическую и сырьевую ценность (Lambert, 1981), кора остается слабо используемым ресурсом. В странах Скандинавии интенсификация лесопользования подразумевает использование всех фракций фитомассы, включая кору, корни и ветви, что ведет к истощению продуктивности лесных экосистем и повышению кислотности почв (Jönsson, 2000). Масштабы снижения продуктивности “истощенных экосистем” в долговременной перспективе пока не оценены. В России же наблюдается обратная ситуация – удаленность заготовительных и лесоперерабатывающих предприятий и, как следствие, отсутствие рынка сбыта древесных отходов приводят к образованию отвалов, содержащих продукты лесопиления, большую часть которых составляет кора. В процессе ее разложения высвобождаются фенольные и другие соединения, которые отрицательно влияют на грунтовые воды и лесные экосистемы, таким образом нарушая санитарное состояние территории. В связи с этим важно определить темпы естественного разложения древесной коры и оценить факторы, влияющие на скорость этого процесса.
Целью исследования являлась характеристика динамики физических свойств коры в процессе ее разложения в составе порубочных остатков после сплошных рубок в смешанных среднетаежных лесах. Решались следующие задачи: 1) оценить проективное покрытие, запас и массу коры, оставшейся на лесосеке после сплошной рубки леса, и сравнить их c таковыми для крупных древесных остатков; 2) выявить закономерности изменения физических характеристик коры, концентраций углерода (С) и азота (N) в процессе разложения и рассчитать параметры моделей, описывающих эти изменения во времени. В процессе работы была протестирована гипотеза об определяющей роли древесной породы в динамике изменения физических и химических характеристик коры на вырубках.
ОБЪЕКТЫ И МЕТОДИКА
Исследование было выполнено в 2015–2016 гг. на вырубках в Республике Карелия. Территория приурочена к среднетаежной подзоне растительности (Курнаев, 1973). Среднегодовая температура составляет +2.4°C, количество осадков – 584 мм год–1. Разница температур между самым теплым (июль) и самым холодным (январь) месяцами равна 34.1°C. Продолжительность вегетационного периода составляет 80 дней.
Почва временны́х пробных площадей, на которой происходило разложение порубочных остатков, была классифицирована как подзол иллювиально-железисто-гумусовый (Соломатина, 2009). Почвообразующая порода представлена пылевато-песчаной мореной.
Сплошная узколесосечная рубка на всех пробных площадях проведена харвестером. Трелевка сортиментов осуществлялась по волокам (форвардером), с временны́м размещением сортиментов на погрузочной площадке. Порубочные остатки (отходы древесины, образующиеся на лесосеке при валке и трелевке деревьев, очистке стволов от сучьев (Острошенко, 2005)) распределены по пробным площадям относительно равномерно, с незначительной аккумуляцией на волоках, где отходы лесозаготовки используют для повышения проходимости техники.
Полевые исследования. Образцы коры были отобраны на четырех временны́х пробных площадях, заложенных на площадях с разной давностью сплошной рубки леса: 6 месяцев (Ялгуба-1, Березовые мосты), 2 года (Гомсельга) и 6 лет (Ялгуба-2) (табл. 1). На двух пробных площадях (Гомсельга, Березовые мосты) преобладающей породой до рубки леса была ель (Picea abies (L.) H.Karst), сопутствующими – сосна (Pinus sylvestris L.), береза (Betula pubescens Ehrh. и Betula pendula Roth.), осина (Populus tremula L.). В напочвенном покрове преобладали виды, характеризующие средние по богатству и влажности почвенные условия таежного леса: черника (Vaccinium myrtillus L.), брусника (Vaccínium vítis-idaéa L.) и др. На третьей пробной площади (Ялгуба-1) доминировала сосна, на четвертой (Ялгуба-2) – осина. В напочвенном покрове более богатых по лесорастительным условиям третьей и четвертой пробных площадях преобладала черника (Vaccinium myrtillus L.), значительные площади занимали кислица (Óxalis acetosélla L.), костяника (Rúbus saxátilis L.) и папоротник (Dryópteris fílix-mas (L.) Schott).
Таблица 1.
Характеристика временны́х пробных площадей
| Пробная площадь, год вырубки | Географические координаты | Тип леса | Состав древостоя до рубки* | Запас древостоя до рубки, м3 га–1 | Запас КДО** на вырубке, м3 га–1*** |
|---|---|---|---|---|---|
| Гомсельга, 2013 г. | 62°06′ с.ш., 33°99′ в.д. | Черничный | 5Е 100–140 2Ос 60–70 2С 50–60 1Б 50–60 |
447 | 17 + 64 + 17 + 29 = 127 |
| Березовые мосты, 2015 г. |
61°90′ с.ш., 34°57′ в.д. | Черничный | 6Е 120–140 2Б 70–80 1С 120–140 1Ос 60–70 |
433 | 14 + 54 + 20 + 17 = 105 |
| Ялгуба-1, 2015 г. | 61°94′ с.ш., 34°44′ в.д. | Кислично-черничный | 5С 100–160 3Б 30–40 1Е 120 1Ос 60–70 |
253 | 55 + 22 + 15 + 12 + 18 = 122 |
| Ялгуба-2, 2010 г. | 61°91′ с.ш., 34°57′ в.д. | Кислично-черничный | 4Ос 60–70 3Е 100–140 2С 120–160 1Б 60–70 |
346 | **** |
Запас крупных древесных остатков (КДО) оценивали на каждой вырубке на четырех 25-метровых трансектах, заложенных с севера на юг (2 трансекты) и с запада на восток (2 трансекты), захватывающих как пасеку, так и волоки. Для валежа и зависших деревьев учитывали породу, класс разложения (Шорохова, Шорохов, 1999) и диаметр в месте пересечения ходовой линии. Объем КДО по породам и классам разложения рассчитывали по формуле (Stahl et al., 2001):
(1)
$V = {{\left( {{{{{\pi }^{{\text{2}}}}} \mathord{\left/ {\vphantom {{{{\pi }^{{\text{2}}}}} {{\text{8}}\Sigma d_{i}^{{{\text{2 }}}}S}}} \right. \kern-0em} {{\text{8}}\Sigma d_{i}^{{{\text{2 }}}}S}}} \right)} \mathord{\left/ {\vphantom {{\left( {{{{{\pi }^{{\text{2}}}}} \mathord{\left/ {\vphantom {{{{\pi }^{{\text{2}}}}} {{\text{8}}\Sigma d_{i}^{{{\text{2 }}}}S}}} \right. \kern-0em} {{\text{8}}\Sigma d_{i}^{{{\text{2 }}}}S}}} \right)} {\Sigma {{L}_{j}}}}} \right. \kern-0em} {\Sigma {{L}_{j}}}},$Кору отбирали с поверхности почвы, на некотором расстоянии (до 50 см) от пня. Данный метод отбора позволяет быть уверенным в том, что процесс биогенного разложения коры на протяжении всего периода проходил на поверхности почвы.
Для исследования были отобраны 140 образцов коры: 40 – ели, 30 – сосны, 33 – осины и 37 – березы. Данные о возрасте рубки (а, значит, и моменте формирования слоя коры вырубки) получены из лесничеств. В качестве контроля (начальной точки разложения) использовали образцы коры, собранные с валежа текущего года (со свежей листвой и хвоей) или с живого дерева (для сосны) в непосредственной близости от временны́х пробных площадей. Для всех образцов коры в полевых условиях измеряли площадь поверхности.
Для оценки запаса коры на свежей вырубке (возраст рубки 0 лет) были заложены две 25-метровые трансекты: одна – с севера на юг, другая с запада на восток. На каждой трансекте с шагом 1 м оценивали процент покрытия коры на поверхности почвы на площадке 1 м2; в общей сложности обследовали 12 площадок. Толщина слоя составляла в среднем 1 см.
Массу слоя коры для исследуемых вырубок рассчитывали, исходя из доли участия древесных пород до рубки леса в составе древостоя, путем умножения объема слоя коры на свежей вырубке на базисную плотность коры.
Массу углерода и азота на свежей вырубки рассчитывали как процент содержания соответствующего элемента в коре от общей массы коры на вырубке.
Лабораторные исследования и расчеты. В лаборатории кору высушивали в сушильном шкафу при температуре 103°С в течение 48 ч до абсолютно сухого состояния, взвешивали (для оценки удельной массы коры), парафинировали, взвешивали после парафинирования для вычитания массы парафина и определяли объем методом гидростатического взвешивания (Полубояринов, 1976).
Для определения содержания N и C все растительные образцы сразу после отбора замораживали в жидком азоте, лиофильно высушивали и измельчали до однородной массы на лабораторной мельнице. Элементоорганический анализ проводили с помощью CHNS/O-анализатора Perkin Elmer 2400 Series II (USA). На анализ отбирали навеску 1.5–2.5 мг абсолютно сухого образца. После каждых 10–20 образцов использовали органический аналитический стандарт ацетанилида (Perkin Elmer № 0204-1121) для калибровки CHNS/O-анализатора.
Скорость потери массы коры рассчитывали на единицу площади (методика разработана на основании ранее проведенных исследований (Шорохова, Капица, 2007)). Массу коры на единицу площади (Mb, г м–2) (удельную массу) вычисляли по формуле:
где M – масса образца коры в абсолютно сухом состоянии, мг; S – площадь поверхности образца, см2.Долю флоэмы (%) абсолютно сухого образца измеряли как процент от толщины коры. Толщину коры (см) рассчитывали делением объема образца на площадь его поверхности. Плотность коры в абсолютно сухом состоянии (ρ, г м–3) находили делением массы образца в абсолютно сухом состоянии на его объем в абсолютно сухом состоянии. Объем абсолютно сухого образца рассчитывали методом гидростатического взвешивания (Полубояринов, 1976).
Статистический анализ данных. Зависимости массы на единицу площади (Mb), плотности (ρ), доли флоэмы и концентрации С и N от древесной породы, времени рубки и типа лесорастительных условий (средние и богатые) оценивали с использованием дисперсионного анализа (ANOVA) и теста Дункана. По результатам статистического анализа данные были разделены на группы. Динамику вышеперечисленных показателей анализировали с помощью линейной множественной регрессионной и генерализированной линейной множественной регрессионной моделей среды R (функции lm, glm), (R Core Team, 2013).
Построение графиков осуществляли в среде R (ggplot2), (R Core Team, 2013).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Вклад слоя коры в круговорот веществ на вырубке. Запас коры, оставшейся после рубки леса, составил 20.8 м3 га–1, из которых 2.1–10.4 м3 га–1 – кора сосны, 2.1–12.5 м3 га–1 – кора ели, 2.1–6.2 м3 га–1 – кора березы, 2.1–8.3 м3 га–1 – кора осины (табл. 2). Запасы крупных древесных остатков на пробных площадях варьировали от 105 до 127 м3 га–1. Оставленные порубочные остатки были представлены в основном доминирующей породой (елью или сосной) и составили 22–64 м3 га–1 для ели и 14–55 м3 га–1 для сосны (табл. 1). Если учесть, что в коренных сосняках средней тайги на долю крупных древесных остатков приходится от 2 до 92 м3 га–1, а в ельниках – от 32 до 328 м3 га–1 (Капица и др., 2012б), оставленные 20.8 м3 га–1 коры являются значительным вкладом в биологический круговорот после сплошнолесосечной рубки леса, особенно с учетом того, что кора богаче биогенными элементами, чем древесина (Palviainen et al., 2010).
Таблица 2.
Объем и масса коры, масса азота (N) в коре, масса углерода (C) в коре в составе порубочных остатков на свежей вырубке
| Показатель | Осина | Береза | Сосна | Ель |
|---|---|---|---|---|
| Запас коры на вырубке, м3 га–1 | 2.1–8.3 | 2.1–6.2 | 2.1–10.4 | 2.1–12.5 |
| Масса коры на вырубке, кг га–1 | 1331–5262 | 1514–4470 | 1045–5179 | 1403–8350 |
| Масса N в коре на вырубке, кг га–1 | 7.1–27.9 | 6.8–20.1 | 2.4–11.9 | 6.0–35.9 |
| Масса C в коре на вырубке, кг га–1 | 559–2210 | 727–2146 | 502–2486 | 645–3841 |
Физические характеристики контрольных образцов. Средний процент флоэмы у контрольных образцов уменьшался в ряду: береза, осина, ель, сосна (табл. 3). Минимальный процент флоэмы был у контрольных образцов сосны – 21% от общей толщины коры, максимальный – у коры березы (71%). Полученные результаты близки к данным, полученным О.И. Полубояриновым и А.М. Сорокиным (1997), согласно которым процент флоэмы коры на высоте 1.3 м также уменьшался в ряду береза, осина, ель, сосна и составил 72.4% (против 71% по нашим результатам), 49.7 (против 55), 31.6 (против 48), 8.72% (против 21%), соответственно. Незначительные различия связаны с тем, что процент флоэмы от общей толщины коры определяется также возрастом дерева, частью ствола и условиями произрастания (Полубояринов, Сорокин, 1997).
Таблица 3.
Средние значения физических и минеральных характеристик коры контрольных образцов
| Показатель | Осина | Береза | Сосна | Ель |
|---|---|---|---|---|
| Базисная плотность p0, г см–3 | 0.634 (0.025) | 0.721 (0.023) | 0.498 (0.072) | 0.668 (0.022) |
| Удельная масса Mb/S, г см–2 | 0.601 (0.073) | 0.446 (0.057) | 0.346 (0.107) | 0.233 (0.012) |
| Толщина, см | 1.101 (0.179) | 0.618 (0.068) | 0.602 (0.053) | 0.348 (0.013) |
| Флоэма, % | 55 (6) | 71 (3) | 21 (10) | 48 (3) |
| Концентрация С, % | 42 (1) | 48 (1) | 48 (3) | 46 (3) |
| Концентрация N, % | 0.53 (0.06) | 0.45 (0.04) | 0.23 (0.04) | 0.43 (0.10) |
| С/N | 88 (12) | 109 (7) | 230 (17) | 135 (37) |
Толщина коры контрольных образцов варьировала от 0.31 до 1.82 см: 0.33–0.98 см для березы, 0.31–0.39 см для ели, 0.53–1.82 см для осины и 0.52–0.70 см для сосны.
Значения начальной плотности коры (p0) варьировали: от 0.51 до 0.71 для осины, от 0.56 до 0.80 для березы, от 0.39 до 0.64 для сосны и от 0.60 до 0.73 г см–3 для ели. Средняя базисная плотность контрольных образцов уменьшалась в ряду: береза, ель, осина, сосна (табл. 3).
Полученные концентрации углерода и азота несколько ниже по сравнению с данными М. Palviainen с соавт. (2010) для пней южной тайги. Так, концентрация углерода для ели, сосны и березы равна 46, 48 и 48% (наши данные), в то время как на вырубках южнотаежной подзоны – 53, 56, 52%; концентрация азота – 0.43, 0.23, 0.45% (наши данные) против 0.31, 0.28, 0.45% (Palviainen et al., 2010). Процентное соотношение химических элементов в коре определяется породой (Palviainen et al., 2010), диаметром ствола, возрастом дерева, частью ствола (Harmon et al., 1986), плодородием почвы (Werkelin et al., 2011), временем года (Helmisaari, Siltala, 1989) и климатом (Berg, McClaugherty, 2003).
Изменение физических показателей коры во времени. Различия в динамике потери удельной массы коры, уменьшении пропорции флоэмы и толщины зависели от породы и времени. Наиболее активно происходило истончение коры осины и сосны, практически вдвое медленнее – коры березы (табл. 4). Процент флоэмы линейно уменьшался со временем разложения (рис. 1).
Таблица 4.
Изменение физических и химических характеристик коры в зависимости от времени и породы
| Зависимая переменная | Модель, распределение, функция связи | F | df | Средние значения коэффициентов моделей для породы | Средние значения коэффициентов моделей для времени | Z | Множественный R2: скорректированный R2 |
|---|---|---|---|---|---|---|---|
| Плотность p, г см–3 | Линейная регрессия, нормальное, тождество | 49.76 | 4 | Свободный член (Осина): 0.61 (0.18) Сосна: –0.19 (0.02) Ель: 0.06 (0.02) Береза: –0.09 (0.02) |
– | – | 0.565 : 0.554 |
| Удельная масса Mb/S, г см–2 | Линейная регрессия, нормальное, тождество | 11.85 | 4 | Свободный член (Осина): 0.47 (0.03) Ель: –0.15 (0.04) |
–0.026 (0.006) | – | 0.234 : 0.217 |
| Толщина, cм | Линейная регрессия, нормальное, тождество | 11.86 | 4 | Свободный член (Осина): 0.84 (0.06) Ель: –0.34 (0.08) Береза: –0.16 (0.08) |
–0.04 (0.01) | – | 0.237 : 0.217 |
| Флоэма, % | Генерализированная линейная, Пуассона, тождество | 1564 (время) 1129 (порода) |
1 3 |
Свободный член (Осина): 3.89 (0.04) Сосна: –1.78 (0.12) Ель: –0.33 (0.06) Береза: 0.15 (0.05) |
–0.30 (0.13) | 93 –14 –5 3 |
– |
| Концентрация N, % | Линейная регрессия, нормальное, тождество | 8.82 | 4 | Свободный член (Осина): 0.65 (0.05) Сосна: –0.33 (0.07) Ель: –0.13 (0.06) |
– | – | – |
| С : N | Линейная регрессия, нормальное, тождество | 4.92 | 4 | Свободный член (Осина): 85.20(10.27) Сосна: 41.79 (13.03) |
– | – | – |
Рис. 1.
Изменение физических характеристик (а – удельной массы, б – доли флоэмы) коры осины, березы, ели и сосны в процессе разложения.
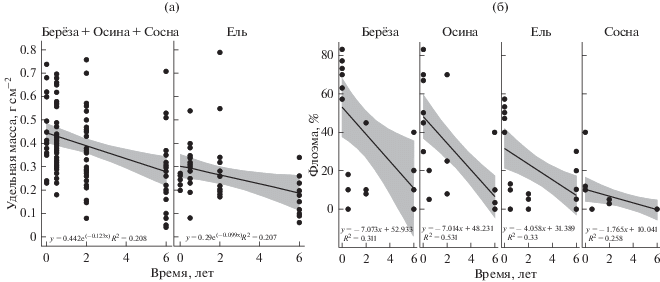
За шесть лет, прошедших после рубки, удельная масса коры осины, березы и сосны уменьшилась практически вдвое. Изменение удельной массы коры ели протекало менее интенсивно (рис. 1). Высокая скорость изменения удельной массы коры лиственных пород может быть связана с отсутствием смолистых соединений в их строении (в отличие от хвойных пород). Смолистые вещества, пронизывающие древесину и кору сосны и ели, выполняют роль биологической защиты от патогенных организмов: монотерпены, входящие в состав смолы, обладают свойствами контактных ядов для насекомых-ксилофагов, которые, фрагментируя кору, ускоряют ее разложение (Рожков, 1972; Ludley et al., 2008). Различия в скорости изменения удельной массы коры сосны и ели объясняются способностью коры сосны расслаиваться больше коры ели. Процесс потери массы на единицу площади связан не только с деструкцией коры дереворазрушающими организмами, но и с отслаиванием коры по мере ее высыхания.
Анатомическое строение коры определяет ее плотность и, как следствие, различия в скорости биологической деструкции (табл. 4). Изменение плотности коры во времени статистически недостоверно, что связано с низкой плотностью флоэмы, которая хоть и быстро разлагается, но вносит незначительный вклад в общую плотность коры. Более того, быстрое разложение неплотной флоэмы повышает плотность коры в целом спустя 6 лет, что, в свою очередь, нивелирует влияние времени на динамику плотности. Отсутствие зависимости изменения плотности от времени также может быть связано с незначительным временем исследования, которого оказалось недостаточно для освоения трудноразлагаемых элементов коры (корки) дереворазрушающими организмами.
Динамика углерода в коре не зависела от времени и породы (рис. 2); концентрация азота в коре на вырубках определялась только древесной породой (табл. 4). Полученные результаты подтверждают теорию о 5–6-летнем лаг-периоде, в течение которого скорость биологической деструкции незначительна. Так, согласно литературным данным (Palviainen et al., 2010), через 5 лет после рубки концентрация углерода в коре пней снижается лишь на 7 (ель) и 1% (сосна). Концентрация азота, вопреки нашим ожиданиям, не увеличилась, что также, возможно, связано с наличием периода “освоения” коры.
Рис. 2.
Концентрации минеральных элементов (а – углерода, б – азота) по мере разложения коры в составе порубочных остатков.
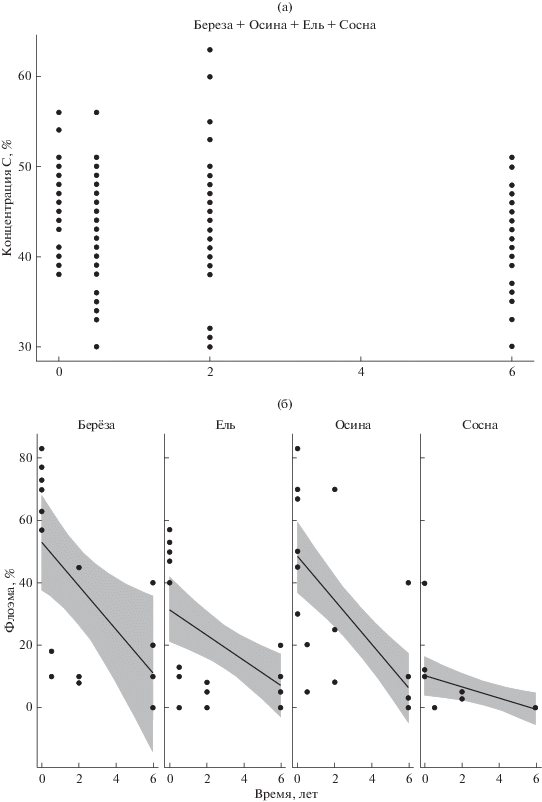
Тип леса (черничный vs. кислично-черничный) не влиял на скорость биогенной деструкции коры на поверхности почвы (удельная масса: F = = 1.706; p = 0.203; плотность: F = 0.018; p = 0.893; толщина коры: F = 4.070; p = 0.058). Гипотеза о более активной деструкции коры на поверхности более богатых почв (Прокушкин и др., 2014) не нашла подтверждения в нашем исследовании. Микроклимат под пологом и на вырубках сильно различается (влажность, температура) (Мурзаева, 1978). На вырубке скорость деструкции коры определяют климатические характеристики района, которые более значимы, чем лесорастительные особенности биотопа.
Заключение. Кора в составе порубочных остатков играет значительную роль в обеспечении устойчивого лесопользования, препятствуя обеднению почв, – запасы коры на вырубке составляют около 1/5 от объема крупных древесных остатков. Особенно существенна роль коры в азотном цикле, т. к. она более богата азотом по сравнению с древесиной. Различия в анатомическом строении древесных пород определяют скорость изменения плотности, процента флоэмы, толщины, удельной массы и азота в коре, а, следовательно, и ее роль как части подстилки в биологическом круговороте.
Мы благодарим за помощь в сборе данных и подборе пробных площадей Алексея Полевого, Анастасию Мамай, Елену Мошкину, Нилу Сироткину, Дарью Глазунову и Никиту Баклаженко.
Список литературы
Дымов А.А., Старцев В.В. Изменение температурного режима подзолистых почв в процессе естественного лесовозобновления после сплошнолесосечных рубок // Почвоведение. 2016. № 5. С. 599–608.
Казимиров Н.И., Морозова Р.М. Биологический круговорот веществ в ельниках Карелии // Л.: Наука, 1973. 175.
Капица Е.А., Трубицына Е.А., Шорохова Е.В. Биогенное разложение стволов, ветвей и корней основных лесообразующих пород темнохвойных северотаежных лесов // Лесоведение. 2012а. № 3. С. 51–58.
Капица Е.А., Шорохова Е.В., Кузнецов А.А. Пул углерода крупных древесных остатков в коренных лесах северо-запада Русской равнины // Лесоведение. 2012б. № 5. С. 36–43.
Курнаев С.Ф. Лесорастительное районирование СССР. М.: Наука. 1973. 203 с.
Лотова Л.И. Микроструктура коры основных лесообразующих лиственных деревьев и кустарников Восточной Европы. М.: КМК. 1999. 114 с.
Мурзаева М.К. Особенности микроклимата на лесосеках различных способов рубок // Леса Урала и хозяйство в них. Свердловск: Сред.-Урал. кн. изд-во. 1978. Вып. 11. С. 73–77.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. Влияние ели на кислотность и содержание элементов питания в почвах северотаежных ельников кустарниково-зеленомошных // Почвоведение. 2016. № 11. С. 1355–1367.
Острошенко В.В. Краткий словарь основных лесоводственно-экономических терминов. Уссурийск: Приморская гос. с.-х. академия, 2005. 196 с.
Поздняков Л.К., Щербаков Н.М., Зябченко С.С., Куликова В.К. Лесные растительные ресурсы Карелии. Петрозаводск: Карельск. филиал АН СССР. 1974. 135 с.
Полубояринов О.И. Плотность древесины. М.: Наука, 1976. 159 с.
Полубояринов О.И., Сорокин А.М. Физические свойства сосновой коры и ее компонентов // Известия высших учебных заведений. Лесной журнал. 1997. № 3. С. 70–74.
Прокушкин С.Г., Прокушкин А.С., Сорокин Н.Д. Интенсивность разложения отдельных компонентов фитодетрита в лиственничниках криолитозоны Средней Сибири // Известия РАН. Серия биол. 2014. № 1. С. 76–85.
Рипачек В. Биология дереворазрушающих грибов. М.: Лесн. пром-сть, 1967. 258 с.
Рожков А.С. Анатомические, гистохимические и биологические преобразования у лиственницы при повреждении насекомыми. Иркутск: АН СССР. Сиб. отд-ние. Сиб. ин-т физиологии и биохимии растений. 1972. 125 с.
Соломатова Е.А. Экология и география почв // Почвы еловых лесов Восточной Фенноскандии. Петрозаводск: Кар НЦ РАН, 2009. С. 66–90.
Шорохова Е.В., Капица Е.А. Фрагментация и разложение еловой коры в лесах южной и средней тайги // Лесоведение. 2007. № 5. С. 22–26.
Шорохова Е.В., Шорохов А.А. Характеристика классов разложения древесного детрита ели, березы и осины в ельниках подзоны средней тайги // Тр. СПб. НИИ лесн. хоз-ва. 1999. Вып. 1(2). С. 17–23.
Berg B., McClaugherty C. Plant litter: decomposition, humus formation, carbon sequestration. Berlin – Heidelberg: Springer Verlag, 2003. 286 p.
Čekstere G., Laiviņš M., Osvalde A. Chemical composition of Scots pine bark as a bioindicator of enviromental quality in Riga, Latvia // Proceedings of the Latvian academy of science. Section B. Natural, Exact, and Applied Sciences. 2015. V. 69. № 3. P. 87–97.
Dossa G.G.O., Paudel E., Cao K., Schaefer D., Harrison R.D. Factors controlling bark decomposition and its role in wood decomposition in the tropical tree species. Scientific Scientific Reports. 6:34153. doi 10.1038/srep34153
Gustafsson L., Eriksson I. Factors of importance for the epiphytic vegetation of aspen Popullus tremula with special emphasis on bark chemistry and soil chemistry. J. Applied Ecology. 1995. V. 32. № 2. P. 412–424.
Harmon M.E., Cromack Jr K., Smith B.G. Coarse woody debris in mixed-conifer forests, Sequoia National Park, California // Canadian J. Forest Research. 1987. V. 17. № 10. P. 1265–1272.
Helmisaari H.S., Siltala T. Variation in nutrient concentrations of Pinus sylvestris stems // Scandinavian J. Forest Research. 1989. V. 4. № 1–4. P. 443–451.
Jönsson A.M. Mineral nutrients of beech (Fagus sylvatica) bark in relation to frost sensitivity and soil treatments in southern Sweden // Annals of Forest Science. 2000. V. 57. № 1. P. 1–8.
Kuhlman E.G. Decomposition of loblolly pine bark by soil- and root-inhabiting fungi // Canadian Journal of Botany 1970. V. 48. № 10. P. 1787–1793.
Lambert M.J. Inorganic constituents in wood and bark of New South Wales forest tree species // Forestry commission of New South Wales research note № 45. Sydney. 1981. 43 p.
Ludley K.E., Robinson C.H., Jickells S., Chamberlain P.M., Whitaker J. Differential response of ectomycorrhizal and saprotrophic fungal mycelium from coniferous forest soils to selected monoterpenes // Soil Biology and Biochemistry. 2008. V. 40. Is. 3. P. 669–678.
Lukina N.V., Orlova M.A., Steinnes E., Artemkina N.A., Gorbacheva T.T., Smirnov V.E., Belova E.A. Mass-loss rates from decomposition of plant residues in spruce forests near the northern tree line subject to strong air pollution // Environmental Science and Pollution Research. 2017. V. 24. № 24. P. 19874–19887.
Palviainen M., Finér L., Laiho R., Shorohova, E., Kapitsa E., Vanha-Majamaa I. Carbon and nitrogen release from decomposing Scots pine, Norway spruce and Silver birch stumps // Forest Ecology & Management. 2010. V. 259. № 3. P. 390–398.
Parameswaran N., Wilhelm G.E., Liese W. Ultrastructural aspects of beech bark degradation by fungi // European J. Forest Pathology. 1976. V. 6. № 5. P. 274–286.
Sawidis T., Breuste J., Mitrovic M., Pavlovic P., Tsigaridas K. Trees as bioindicator of heavy metal pollution in three European cities // Environmental Pollution. 2011. V. 159. № 12. P. 3560–3570.
Shorohova E., Kapitsa E., Vanha-Majamaa I. Decomposition of stumps 10 years after partial and complete harvesting in southern boreal forest in Finland // Canadian J. Forest Research. 2008a. V. 38. № 9. P. 2414–2421.
Shorohova E., Kapitsa E., Vanha-Majamaa I. Decomposition of stumps in a chronosequence after clear-felling vs. clear-felling with prescribed burning in southern boreal forest in Finland // Forest Ecology and Management. V. 255. № 10. 2008b. P. 3606–3612.
Shorohova E., Kapitsa E. Influence of the substrate and ecosystem attributes on the decomposition rates of coarse woody debris in European boreal forests // Forest Ecology and Management. 2014. V. 315. № 1. P. 173–184.
Ståhl G., Ringvall A., Fridman J. Assessment of coarse woody debris – a methodological overview // Ecological Bulletins. 2001. V. 49. P. 57–70.
Tarasov M.E., Birdsey R.A. Decay rate and potential storage of course woody debris in the Leningrad region // Ecological Bulletins. 2001. V 49. P. 137–147.
Werkelin J., Lindberg D., Boström D., Skrifvars B.-J., Hupa M. Ash-forming elements in four Scandinavian wood species. Part 3. Combustion of five spruce samples // Biomass & Bioenergy. 2011. V. 35. № 1. P. 725–733.
Wilhelm G.E. Űber die Zersetzung von Buchen- und Fichtenrinde unter natűrlichen Bedingungen // European Journal of Forest Pathology. 1976. V. 6. № 2. P. 80–91.
Дополнительные материалы отсутствуют.