

Лесоведение, 2019, № 2, стр. 138-156
Микробиологическая оценка состояния почв хвойных лесов Средней Сибири после пожаров разной интенсивности
А. В. Богородская a, *, Е. А. Кукавская a, О. П. Каленская b, Л. В. Буряк b, c
a Институт леса им. В.Н. Сукачева Сибирского отделения Российской академии наук –
обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
b Сибирский государственный университет науки и технологий
им. М.Ф. Решетнева
660049 Красноярск, просп. Мира, 82, Россия
c Филиал ФБУ ВНИИЛМ “Центр лесной пирологии”
660022 Красноярск, ул. Крупской, 42, Россия
* E-mail: anbog@ksc.krasn.ru
Поступила в редакцию 12.02.2018
После доработки 06.03.2018
Принята к публикации 08.10.2018
Аннотация
Дана оценка послепожарной трансформации некоторых физико-химических и микробиологических свойств почв среднетаежных хвойных насаждений Средней Сибири. Низовые пожары в кедровых, лиственничных и производных хвойно-лиственных насаждениях приводили к увеличению функциональной активности, численности эколого-трофических групп микроорганизмов, запасов Смик и микробной продукции СО2 в верхних минеральных почвенных горизонтах, причем данный эффект более очевиден после высокоинтенсивных пожаров. В подстилках и верхнем 0–5 сантиметровом подзолистом горизонте почв кедровников через месяц после высокоинтенсивных пожаров наблюдалось увеличение значения qСО2 в 2–3 раза, тогда как низкоинтенсивные пожары в изучаемых насаждениях не вызвали значительного изменения устойчивости почвенного микробоценоза. С течением времени, пройденного после пожара, значения qСО2 снижаются. В подстилке лиственничника вейникового через четыре года после высокоинтенсивного пожара отмечено снижение содержания микробной биомассы в 2 раза, а также численности аммонификаторов и повышение олиготрофности. Послепожарная динамика направлена на снижение общей микробной продукции СО2 почвами изучаемых насаждений, при этом даже спустя четыре года после пожаров в лиственничниках, независимо от интенсивности, она в 2 раза выше и отстает от уровня повышения общих запасов микробной биомассы.
Пожары – один из основных естественных циклических факторов, вызывающих изменение и современную эволюцию таежных ландшафтов (Фуряев, 1996). В бореальных лесах Сибири пожары ежегодно охватывают несколько миллионов гектар (Kukavskaya et al., 2013) и являются основным фактором нарушения функционирования лесных экосистем.
В России ареал светлохвойных насаждений с преобладанием сосны и лиственницы в большей степени подвергается пожарам (Барталев и др., 2015), что связано с их высокой природной пожарной опасностью (Иванова и др., 2014). Темнохвойные леса, напротив, подвержены меньшей пожароопасности вследствие высокой влажности лесных горючих материалов (Софронов и др., 2005). Однако при потеплении климата и в периоды длительной засухи возникает вероятность распространения пожаров в темнохвойных лесах, и их последствия могут оказаться разрушительными (Седых, 2014).
Особо охраняемые природные территории, являются эталонами современного состояния таежных экосистем и характеризуются высокой комплексностью почвенного и растительного покрова, при этом, в поиске фактора, нарушающего таежные леса и, в том числе на особо охраняемых природных территориях, можно назвать пожары. Ведущей причиной возникновения пожаров на территории государственного природного биосферного заповедника “Центральносибирский” являются грозы (Иванов, Иванова, 2010). Следует отметить, что в последнее десятилетие отмечено увеличение горимости лесов заповедника (Кукавская и др., 2017).
Развитие летних устойчивых пожаров на территории заповедника связано с широким распространением темнохвойных насаждений моховых типов леса, характеризующихся сомкнутой структурой древостоев, что обусловливает поздний сход снегового покрова, повышенное влагосодержание напочвенных горючих материалов, и, соответственно, более позднее пожарное созревание участков лесных земель (Кукавская и др., 2017). При развитии устойчивых низовых пожаров огонь заглубляется в подстилку, уже просохшую к летнему периоду. Вследствие высокой плотности мощного горизонта подстилки горение происходит в режиме тления, за счет чего увеличивается резидентное время воздействия огня и возрастает степень повреждения как напочвенного покрова и почвы, так и деревьев, имеющих в данных условиях местопроизрастания поверхностную корневую систему. Показано, что низовые устойчивые пожары даже слабой интенсивности могут быть губительны для кедровых лесов (Семечкин и др., 1985).
Послепожарная трансформация почв (физико-химические свойства, механический состав, водно-воздушный и гидротермический режимы) изучалась многими авторами в зоне распространения умеренных и бореальных лесов России, Северной Америки и Европы (Попова, 1997; Nearly et al., 1999; Безкоровайная, и др., 2005; Certini, 2005; De Marco et al., 2005; Thiffault et al., 2008). Особое внимание было уделено изучению биологических свойств лесных почв после пожаров (Сорокин, 1983; Hernandez et al., 1997; Yeager et al., 2005; Богородская, 2006; Ginzburg, Steinberger, 2012; Heydari et al., 2017; Sharma et al., 2017).
Исследования влияния пожаров на почвы темнохвойных лесов немногочисленны. Так, изучено послепожарное формирование почв кустарничково-зеленомошных кедровников в южном Прибайкалье (Краснощеков, Чередникова, 2012; Краснощеков и др., 2013). Авторы отмечают, что низовые пожары подстилочно-гумусового вида трансформируют органогенные горизонты подзолов грубогумусовых и приводят к формированию маломощных пирогенных горизонтов, химические, физико-химические свойства и биологический круговорот зольных элементов которых существенно отличаются от природных аналогов. После пожаров происходит смена темнохвойных лесов мелколиственными производными, что, в свою очередь, сопровождается значительным изменением живого растительного покрова, трансформацией грубогумусовых и формированием органогенных пирогенных горизонтов. Отмечено, что нарушенные пожарами почвы кедровых лесов в южном Прибайкалье имеют длительный период восстановления.
Изучение послепожарного состояния горно-таежных ожелезненных почв кедровых и пихтовых лесов Прибайкальской провинции свидетельствует о значимости пирогенного фактора в их динамике (Стефин, 1981). В работе отмечено, что в почве 10–20 летних гарей, представленных производными березняками, отношение С : N в верхних горизонтах возрастает, а в нижних – уменьшается. Это, по-видимому, связано со значительной потерей азота в гумусовом горизонте и с увеличением скорости минерализации гумуса в нижнем. Отмечено, что после пожаров кислотность почв близка к нейтральным значениям, объемный вес лесной подстилки возрастает, влага в почве перераспределяется. В почве пихтарника при неоднократном воздействии огня слабой силы не отмечали потерь азота, изменения С : N и актуальной кислотности, однако объемный вес подстилки возрастал (Стефин, 1981). В целом показано, что высокоинтенсивные пожары снижают противоэрозионную устойчивость почв невозобновившихся гарей, а низкоинтенсивные, но повторяющиеся, создают благоприятные условия для возобновления темнохвойных лесов под материнским пологом.
Показано, что после верхового пожара в северо-таежном ельнике спустя 9 лет образовался постпирогенный лиственный молодняк, в дерново-подзолистой почве которого морфологические пирогенные признаки выявлены до глубины 20–30 см (Дымов и др., 2015). Подстилка лиственного молодняка насыщена основаниями и обогащена азотом по сравнению с коренным ельником. Существенных пирогенных изменений в запасах углерода и азота не выявлено, но уменьшается вклад подстилки в общие запасы углерода в профиле почв.
Внимание к изучению послепожарных изменений почвенных микробоценозов определяется их ключевой ролью в цикле биогенных элементов и продукционно-деструкционных процессах органического вещества. Показана высокая вариабельность отклика почвенных микроорганизмов на пожар (Dooley, Treseder, 2012; Sharma et al., 2017). Исследователи отмечают как кратковременное увеличение численности и биомассы почвенных микроорганизмов после пожаров (Vazquez et al., 1993; Grasso et al., 1996; Wuthrich et al., 2002), так и снижение этих показателей (Fritze et al., 1993; Pietikainen, Fritze, 1993, 1995; Hernandez et al., 1997; Nearly et al., 1999; Certini, 2005; Holden et al., 2016). Однако отмечают, что спустя нескольких дней или месяцев биомасса почвенных микроорганизмов восстанавливается до исходного уровня или даже превышает его. Показано также, что уменьшение содержания микробной биомассы и скорости микробного дыхания почвы определяется интенсивностью пожара и потерей ее органического углерода (Nearly et al., 1999; Holden et al., 2016; Heydari et al., 2017). Выявлено, что восстановление функционирования почвенных микробоценозов после пожаров зависит от содержания органического вещества почвы, его качественного состава, физико-химических и гидротермических свойств, интенсивности пожара, продолжительности его воздействия и глубины прогорания (Nearly et al., 1999; Certini, 2005; Holden et al., 2016). Отмечают влияние и послепожарных сукцессий напочвенного покрова (Thiffault et al., 2008; Богородская и др., 2011; Cutler et al., 2017).
Целью исследований являлась оценка структурно-функционального состояния микробоценозов почв после пожаров разной интенсивности в среднетаежных хвойных насаждениях Средней Сибири. Полученные данные могут быть использованы в биодиагностике послепожарного состояния почв и мониторинге нарушенных лесных экосистем.
ОБЪЕКТЫ И МЕТОДИКА
Почвенно-микробиологические исследования проводились в хвойных насаждениях государственного природного биосферного заповедника “Центральносибирский”, локализованного в умеренно континентальном климате. Средняя температура января составляет –25°С, июля – 16°С. Количество выпадающих осадков – 400–580 мм в год. Продолжительность безморозного периода 65–96 дней, средняя высота снегового покрова достигает 94–114 см (Горшков и др., 2003). Почвообразование протекает в условиях промывного типа водного режима, тепловой режим – длительно сезонно-промерзающий.
Согласно схеме лесорастительного районирования России (Коротков, 1994), исследованные лесные участки расположены в Приенисейской лесорастительной провинции в пределах Средне-Сибирской плоскогорной лесорастительной области, где произрастают кедровые, пихтовые, еловые, лиственничные и березовые насаждения при доминировании темнохвойных лесов.
В правобережной части бассейна р. Енисей было заложено 4 пробных площади (пр. пл.), на каждой из которых выделяли участки лесных земель, пройденные пожарами от слабой (С) до высокой (В) интенсивности (табл. 1). Для каждой пробной площади подбирали контрольное насаждение (К) (длительно негоревшее), которое располагалось в непосредственной близости от участков, пройденных огнем, в одном лесном массиве и одинаковых лесорастительных условиях, а древостои изначально имели схожие лесотаксационные характеристики.
Таблица 1.
Местоположение и лесотаксационная характеристика насаждений пробных площадей, пройденных пожарами разной силы и давности
| № пр.пл. | Категория участка* | Абсолютная высота, м | Крутизна склона, °; экспозиция | Пожар | Характеристика древостоя** | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| давность, лет | сила | тип леса*** | состав**** | полнота | запас, м3 га–1 (доля отпада, %) |
бонитет | |||||
| 1 | С | 73 | 3; ЮЗ | 0 | Слабая | $\frac{{{\text{К р т р з м }}}}{{{\text{К м р т }}}}$ | $\frac{{{\text{8К 2Л ц }} + {\text{Е , е д }}{\text{.П , Б }}}}{{{\text{8К 2Л ц }} + {\text{Е , е д }}{\text{.П , Б }}}}$ | $\frac{{1.2}}{{1.1}}$ | $\frac{{560}}{{506}}$ | (10) | IV |
| В | 54 | 10; ЮЗ | Сильная | $\frac{{{\text{К р т р з м }}}}{{{\text{Л ц м р т }}}}$ | $\frac{{{\text{4К 4Л ц 2Е }} + {\text{Б }}}}{{{\text{5Л ц 3К 2Е }} + {\text{Б }}}}$ | $\frac{{1.3}}{{0.8}}$ | $\frac{{472}}{{287}}$ | (39) | III | ||
| 2 | В | 234 | – | Сильная | $\frac{{{\text{К л ш з м }}}}{{{\text{К м р т }}}}$ | $\frac{{{\text{8К 1Е 1Б }}}}{{{\text{8К 1Е 1Б }}}}$ | $\frac{{0.5}}{{0.3}}$ | $\frac{{230}}{{154}}$ | (33) | IV | |
| 3 | С | 133 | 5; СВ | 2 | Слабая | $\frac{{{\text{Б з м }}}}{{{\text{Б х в в е й н }}}}$ | $\frac{{{\text{4Б 3Л ц 2Е 1К }} + {\text{П }}}}{{{\text{5Б 2Е 2К 1Л ц , е д }}{\text{.П }}}}$ | $\frac{{0.7}}{{0.4}}$ | $\frac{{310}}{{199}}$ | (36) | IV |
| 4 | С | 91 | 15; Ю | 4 | От слабой до средней | $\frac{{{\text{Л ц з м }}}}{{{\text{Л ц к и п р }}}}$ | $\frac{{{\text{8Л ц 2К }} + {\text{Е , е д }}{\text{.П }}}}{{{\text{9Л ц 1К }}}}$ | $\frac{{1.3}}{{0.5}}$ | $\frac{{544}}{{216}}$ | (60) | III |
| В | 81 | 15; Ю | Сильная | $\frac{{{\text{Л ц з м }}}}{{{\text{Л ц в е й н }}}}$ | $\frac{{{\text{7Л ц 2Е 1К , е д }}{\text{.П }}}}{{{\text{9Л ц 1К }}}}$ | $\frac{{1.3}}{{0.3}}$ | $\frac{{465}}{{111}}$ | (76) | IV | ||
* С – участок леса, пройденный низкоинтенсивным пожаром; В – участок леса, пройденный высокоинтенсивным пожаром. ** В числителе – до пожара; в знаменателе – после пожара. *** К ртр зм – кедровник разнотравно-зеленомошный; К мрт – кедровник мертвопокровный; Лц мрт – лиственничник мертвопокровный; К лш зм – кедровник лишайниково-зеленомошный; Б зм – березняк зеленомошный; Б хв вейн – березняк хвощево-вейниковый; Лц зм – лиственничник зеленомошный; Лц кипр – лиственничник кипрейный; Лц вейн – лиственничник вейниковый. **** К – кедр; Лц – лиственница; Е – ель; П – пихта; Б – береза.
Исследуемые насаждения до пирогенного воздействия представлены смешанными хвойными древостоями с преобладанием кедра сибирского (Pinus sibirica Du Tour) (пр. пл. 1–2), лиственницы сибирской (Larix sibirica Ledeb.) (пр. пл. 4) и участием в составе ели сибирской (Picea obovata Ledeb.), пихты сибирской (Abies sibirica Ledeb.) и березы пушистой (Betula pubescens Ehrh.). Насаждение на пр. пл. 3 представлено производным хвойно-лиственным древостоем с преобладанием в его составе березы. Данная пробная площадь включена в исследование, так как представляет собой характерную стадию послепожарной сукцессии, где в настоящее время происходит формирование хвойного насаждения. Периодичность пожаров на изученных площадях (определено методом перекрестного датирования) составляла от 100 до 200 лет. Средний диаметр древостоя темнохвойных и лиственничных насаждений – 27–37 см, высота – 22–24 м, возраст – 250 лет, в производном березняке – 19 см, 20 м и 110 лет соответственно. Класс бонитета изученных насаждений III–IV.
Исследовали насаждения, пройденные низовыми пожарами в год исследования и спустя 1, 2 и 4 года после пирогенного воздействия. Пожары распространялись в летний период (июль–август), их интенсивность варьировала от слабой до сильной. На пр. пл. 2 пожар развил беглую форму, на остальных – устойчивую. Отпад деревьев по запасу варьировал от 10 до 76% в зависимости от интенсивности и давности пожаров (табл. 1).
Живой напочвенный покров в ненарушенных пожаром насаждениях (контрольные пробные площади) представлен мхами (Pleurozium schreberi, Hylocomium splendens, Polytrichum commune), проективное покрытие которых достигает 100% (табл. 2). На пр. пл. 2 присутствуют и лишайники (Cladonia rangiferina, C. arbuscula, C. gracilis), в составе трав доминируют Equisetum sylvaticum, Carex globularis, Rubus saxatilis, Pyrola rotundifolia, кустарничков – Vaccinium vitis-idaea, Linnaea borealis, V. myrtillus. На участках лесных земель, пройденных пожарами, живой напочвенный покров сгорает полностью или погибает в результате теплового воздействия. Исключение составляют долгомошниковые или сфагновые синузии, приуроченные к понижениям нанорельефа. Запасы живого напочвенного покрова снижаются на 45–78%. Участки лесонасаждения, пройденные пожарами в год исследования, представлены мертвопокровным типом. Через 2–4 г. после пожаров проективное покрытие живого напочвенного покрова образованного, в основном, травами, на изученных участках достигает 60–100%, а его запасы составляют 2.2 т га–1 (табл. 2).
Таблица 2.
Характеристика напочвенного покрова на пробных площадях
| № пр. пл. | Категория участка* | Преобладающие виды** | Мощность, см | Запас, г м–2 | ||||
|---|---|---|---|---|---|---|---|---|
| травы, кустарнички | мхи, лишайники | мох, лишайник | подстилка | травы, кустарнички | мох, лишайник | подстилка | ||
| 1 | К | Equisetum sylvaticum, Carex globularis, Maianthemum bifolium, Vaccinium vitis-idaea, V. myrtillus, Pyrola rotundifolia, Linnaea borealis, Calamagrostis obtusata, Rubus saxatilis, Gymnocarpium dryopteris (90) | Pleurozium schreberi, Hylocomium splendens, Polytrichum commune (100) | 11.6 ± 1.6 | 13.6 ± 2.1 | 23 ± 2 | 1030 ± 296 | 3485 ± 386 |
| С | Carex globularis, Vaccinium vtis-idaea, Maianthemum bifolium, Linnaea borealis, Equisetum sylvaticum (10) | Polytrichum commune (10) | 1.3 ± 0.06 | 7.2 ± 0.1 | 4 ± 2 | 317 ± 144 | 2312 ± 227 | |
| В | – | – | – | 5.2 ± 0.1 | – | – | 1615 ± 334 | |
| 2 | К | Equisetum sylvaticum, Ledum palustre, Vaccinium vitis-idaea, V. myrtillus, Pyrola rotundifolia, Linnaea borealis, Calamagrostis obtusata (70) | Pleurozium schreberi,
Hylocomium splendens, Cladonia rangiferina, C. arbuscula, C. gracilis (100) |
5.7 ± 0.7 | 10.6 ± 2.4 | 57 ± 13 | 531 ± 62 | 2768 ± 329 |
| В | – | – | – | 4.5 ± 0.8 | – | – | 1730 ± 64 | |
| 3 | К | Calamagrostis obtusata, Equisetum sylvaticum Chamaenerion angustifolium, Pteridium aquilinum, Geranium sylvaticum, Sanguisorba officinalis, Rubus saxatilis (90) | Pleurozium schreberi, Hylocomium splendens (60) | 4.7 ± 0.3 | 4.2 ± 0.5 | 68 ± 9 | 389 ± 45 | 1534 ± 142 |
| С | Ledum palustre, Vaccinium vitis-idaea, V. myrtillus, Calamagrostis obtusata, Pyrola rotundifolia, Linnaea borealis, Maianthemum bifolium, Equisetum sylvaticum, Rubus saxátilis (60) | – | – | 3.4 ± 0.9 | 43 ± 1 | – | 1161 ± 225 | |
| 4 | К | Equisetum sylvaticum, Vaccinium vitis-idaea, V. myrtillus, Calamagrostis obtusata, Lathyrus pratensis saxatilis (90) | Hylocomium splendens, Pleurozium schreberi (100) | 10.5 ± 1.3 | 5.0 ± 0.6 | 17 ± 6 | 1350 ± 537 | 2164 ± 341 |
| С | Chamaenerion angustifolium, Calamagrostis obtusata, Equisetum sylvaticum, Pyrola rotundifolia, Viola uniflora (80) | Dicranum undulatum (40) | 0.5 ± 0.2 | 2.8 ± 0.2 | 215 ± 46 | 8 ± 6 | 1386 ± 179 | |
| В | Calamagrostis obtusata, Chamaenerion angustifolium, Equisetum sylvaticum, Maianthemum bifolium, Pyrola rotundifolia, Viola uniflora (70) | Dicranum undulatum (80) | 0.5 ± 0.3 | 1.4 ± 0.2 | 121 ± 45 | 11 ± 7 | 746 ± 228 | |
В кедровых и лиственничных насаждениях зеленомошных и разнотравно-зеленомошных (пр. пл. 1, 4) представлены подзолистые иллювиально-железистые среднесуглинистые с микропрофилем подзола почвы (Шишов и др., 2004). Строение почвенного профиля имеет следующий вид: О(0–11)–EL[e-hf](11–13)–BEL(13–31)–BT(31–97)–C(97–106 см). Почва кедровника лишайниково-зеленомошного (пр. пл. 2) – подзолисто-глеевая среднесуглинистая O(0–10)–EL(10–13)–BELg(13–25)–G(25–47)–CG(47–60), в производном хвойно-лиственном насаждении (пр. пл. 3) – литозем темногумусовый легкосуглинистый (О(0–4)–AU(4–12)–C(12–14)–M(14–16).
Для определения физико-химических свойств почв образцы отбирались из почвенных разрезов по генетическим горизонтам. Для микробиологических анализов отбирали образцы подстилки и минерального горизонта почвы до 10 см, для которого многими исследователями показано влияние пожаров на почвенный микробоценоз (Nearly et al., 1999; Богородская, Сорокин, 2006). Определяли влажность почвы (термовесовой метод), температуру (портативный термометр “Checktemp”) и плотность (бур Н.А. Качинского). На пр. пл. 1 отбор образцов почвы проводили в год пожара и через год. Физико-химические свойства почв определяли стандартными методами (Аринушкина, 1970).
Почвенную суспензию высевали на разные твердые питательные среды для подсчета: аммонификаторов (мясо-пептонный агар, МПА); прототрофов, использующих минеральные формы азота (крахмало-аммиачный агар, КАА); олиготрофов (почвенный агар, ПА); олигонитрофилов (среда Эшби), численность которых выражали колоние-образующими единицами (КОЕ) в 1 грамме сухой почвы (Методы .., 1991). Структуру эколого-трофических групп микроорганизмов (ЭКТГМ) оценивали отношением их численности.
Определяли экофизиологические параметры функциональной активности почвенных микроорганизмов: содержание углерода микробной биомассы (Смик) методом субстрат-индуцированного дыхания (СИД) (Anderson, Domsch, 1978; Ананьева, 2003) и базальное дыхание (БД, почва без корней). БД и СИД определяли по разности концентраций СО2 в начале и конце инкубации при помощи газового хроматографа “Agilent Technologies 6890N”, снабженного пламенно-ионизационным детектором и метанатором (“Hewlett-Packard”, США). Методика и рабочие параметры хроматографа описаны ранее (Богородская, Кукавская, 2016). Смик определяли путем пересчета скорости СИД по формуле: Смик (мкг С г–1 почвы) = (мкл СО2 г–1 почвы ч–1) × × 40.04 + 0.37. Скорость базального дыхания выражали в мкг СО2–С г–1 ч–1.
Микробный метаболический коэффициент qСО2 (мкг СО2–С мг–1 Смик ч–1) или удельное дыхание микробной биомассы рассчитывали как отношение БД ${\text{C}}_{{{\text{м и к }}}}^{{ - 1}}$ (Anderson, Domsch, 1993). Рассчитывали запас углерода микробной биомассы Смик (г С м–3) и скорость базального дыхания БД (мг СО2–С м–3 ч–1) в исследуемых горизонтах почв, с учетом их объемного веса (p, г см–3) и объема (V, м3) (Сусьян и др., 2009; Стольникова и др., 2011). Также рассчитывали суммарные запасы Смик и общего БД (микробное продуцирование СО2) исследуемого профиля почв.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Послепожарная трансформация свойств почв хвойных насаждений
Послепожарная трансформация физико-химических свойств почвы рассматривалась для кедровника разнотравно-зеленомошного (пр. пл. 1). В длительно негоревшем кедровнике (контроль) реакция почвы кислая (pH водной вытяжки не превышает 4.0), что обусловлено доминированием в напочвенном покрове мхов и хвойного опада. Разложение органических остатков подстилки в условиях благоприятного сочетания тепла и влаги на фоне промывного водного режима способствует образованию кислых продуктов и развитию процессов подзолообразования.
Подзолистая почва длительно негоревшего кедровника бедна гумусом, содержание которого в горизонте EL не превышает 0.83%, и оно снижается с глубиной. Гумус почв очень слабо обогащен азотом, валовое содержание которого в горизонте EL составляет 0.04%, в BEL – 0.007% (табл. 3).
Таблица 3.
Изменение физико-химических свойств почв кедровника разнотравно-зеленомошного после пожаров разной интенсивности на пр. пл. 1
| Категория участка* | Период после пожара, лет | Горизонт | Глубина, см | pH водный | pH солевой | Валовые | К2О, мг кг–1 |
Р2О5, мг кг–1 |
|
|---|---|---|---|---|---|---|---|---|---|
| гумус, % | азот, % | ||||||||
| К | 0 | EL | 11–13 | 4.0 | 3.7 | 0.8 | 0.04 | 419 | 230 |
| ВEL | 13–31 | 3.3 | 2.8 | 0.6 | 0.007 | 80 | 265 | ||
| С | 0 | EL | 5–7 | 4.1 | 3.3 | 4.9 | 0.03 | 288 | 155 |
| ВEL | 7–24 | 4.3 | 3.4 | 3.4 | 0.004 | 93 | 156 | ||
| 1 | EL | 6–9 | 3.9 | 2.9 | 2.7 | 0.02 | 201 | 142 | |
| ВEL | 9–28 | 3.3 | 3.2 | 2.3 | 0.005 | 59 | 188 | ||
| В | 0 | Opir | 1–3 | 5.3 | 4.4 | 5.0 | 0.02 | 194 | 57 |
| EL | 3–7 | 5.3 | 4.5 | 4.6 | 0.01 | 100 | 36 | ||
| ВEL | 7–25 | 5.5 | 4.5 | 1.2 | 0.006 | 105 | 121 | ||
| 1 | Opir | 0.5–3 | 5.1 | 3.9 | 5.4 | 0.02 | 126 | 99 | |
| EL | 3–7 | 4.2 | 3.9 | 5.0 | 0.01 | 95 | 70 | ||
| ВEL | 7–25 | 4.5 | 4.1 | 1.6 | 0.005 | 73 | 122 | ||
* Обозначения см. табл. 2.
После пожаров на поверхность почвы поступает большое количества золы, что приводит к изменению физико-химических свойств верхних почвенных горизонтов. Их кислотность снижается активнее после высокоинтенсивного пожара (до 5.3 pH водной вытяжки, до 4.4 – солевой), тогда как после низкоинтенсивного снижение кислотности незначительное и сравнимо с контролем (табл. 3). Однако спустя год после высокоинтенсивного пожара кислотность этих горизонтов постепенно нарастает, что можно объяснить поступлением кислых продуктов разложения опада хвои (фульвокислоты) с поврежденных деревьев на поверхность почвы.
После пожаров содержание гумуса в подзолистом горизонте кедровника увеличивается до 4.6–4.9%, в пирогенном достигает 5% (табл. 3). Известно, что при пожарах происходит потеря органического азота почвами (Nearly et al., 1999). Показано, что между температурой прогрева подстилок бореальных лесов Канады и выходом пирогенного углерода имеется четкая корреляция (Santín et al., 2016). Часть общего запаса углерода подстилок (24%) переходит в пирогенный органический материал, который может оказывать влияние на их химические свойства. Так называемый “черный углерод” (“вlack carbon”) устойчив к окислению и вовлечению в биологический круговорот, он депонируется в почве на тысячи лет, являясь долговременным стоком углерода. В свою очередь этот углерод приводит к деградации почвенного гумуса через активацию почвенной микробиоты (Wardle et al., 2008). Повышение содержания гумуса в почве после пожаров отражает не процесс гумификации, а накопление пирогенного углерода в почвенном профиле. Кроме того, послепожарное появление травянистых растений вместо зеленых мхов активизирует дерновый процесс, что проявляется в накоплении в почве азота и гумуса (Попова, 1997). Через год после высокоинтенсивного пожара содержание гумуса в аккумулятивном пирогенном горизонте составляло 5.4%, а в подзолистом – 5%.
В кедровнике разнотавно-зеленомошном после высокоинтенсивного пожара содержание валового азота снижается до 0.01–0.02%, при низкоинтенсивном пожаре – до 0.03%. Через год после пожаров содержание валового азота также снижено (табл. 3). Несмотря на снижение валового азота пожары способствуют повышению содержания в почве доступных форм азота и скорости его минерализации (Попова, 1997; Thiffault et al., 2008; Sharma et al., 2017).
Отмечено, что после пожаров в почве возрастает концентрация калия, кальция, магния и фосфора, которые содержатся в золе сгоревших растений (Hernandez et al., 1997; Neary et al., 1999). Дальнейшее снижение их содержания зависит от выщелачиваемости ионов и их взаимодействия с минеральной частью почвы (De Marco et al., 2005).
Содержание подвижных форм калия и фосфора в почве контрольного участка в 1.5–2 раза больше такового после пожаров. Так, содержание подвижного фосфора связано с низкой щелочностью, которая повышает растворимость соединений и переход их в доступные формы (Мякина, Аринушкина, 1979). В почвах отмечается небольшое содержание калия. Основная часть этого элемента сосредоточена во вторичных глинистых минералах. Так как почвы сформировались на песках и супесях водно-ледникового происхождения, преимущественно представленных первичными минералами, вероятно, это и обусловливает низкое содержание данного элемента (Мякина, Аринушкина, 1979).
Таким образом, проведенные исследования показали неоднозначный характер влияния пожаров на физико-химические свойства подзолистых почв кедровников разнотравно-зеленомошных. Пожары высокой интенсивности приводят к трансформации органогенных горизонтов. Большое содержание золы вызывает снижение актуальной и обменной кислотности, рост содержания гумуса, при этом содержание валового азота, подвижных форм калия и фосфора в почве сокращается.
Изменения экофизиологических показателей (Смик, БД, qСО2) микробоценозов почв после пожаров
Содержание Смик в подстилке изученных насаждений варьировало от 1610 мкг С г–1 в кедровнике лишайниково-зеленомошном до 3230 мкг С г–1 в лиственничнике зеленомошном (рис. 1). Скорость БД подстилок контрольных насаждений составила 6–8.3 мкг СО2–С г–1 ч–1. В верхних 0–5 см гумусового горизонта литозема темногумусового в производном березняке зеленомошном показатели Смик и БД достигали максимальных величин: 610 мкг С г–1 и 2.15 мкг СО2–С г–1 ч–1 соответственно. В подзолистом горизонте кедровников и лиственничников показатели изменялись и составили 310–500 мкг С г–1 и 1.1–1.5 мкг СО2–С г–1 ч–1. Микробиологические процессы в почвах кедровых и лиственничных насаждений на контрольных участках хорошо сбалансированы, экофизиологические показатели постепенно снижаются с глубиной почвенного профиля и имеют тесную корреляцию с температурой почвенных горизонтов (r = 0.96) и их влажностью (r = 0.91).
Рис. 1.
Содержание углерода микробной биомассы (гистограммы) и интенсивность базального дыхания (графики) в почвах хвойных насаждений заповедника, пройденных пожарами разной интенсивности. К – контроль; С – участок леса, пройденный слабоинтенсивным пожаром, В – участок леса, пройденный высокоинтенсивным пожаром. А – пр. пл. 1, кедровник разнотравно-зеленомошный; Б – пр. пл. 2, кедровник лишайниково-зеленомошный; В – пр. пл. 3, березняк зеленомошный; Г – пр. пл. 4, лиственничник зеленомошный.
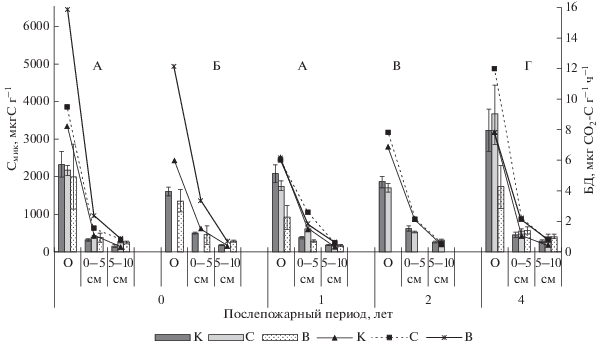
Через месяц после пожара высокой интенсивности в пирогенно преобразованной подстилке кедровника разнотравно-зеленомошного (пр. пл. 1) содержание Смик снижалось на 25% по сравнению с контролем, скорость БД возрастала почти в 2 раза и достигала 15.9 мкг СО2–С г–1 ч–1. В минеральных горизонтах подзолистой почвы рассматриваемого кедровника содержание Смик возрастало на 25–65% по сравнению с контролем, а БД – более чем в 2 раза. После низкоинтенсивного пожара в подстилке этого типа насаждений не отмечено достоверных изменений величин Смик и БД, а в минеральных горизонтах они выше контроля в 1.5–2 раза.
Спустя месяц после высокоинтенсивного пожара в кедровнике лишайниково-зеленомошном в подстилке содержание Смик уменьшалось на 30% от контроля, скорость БД, напротив, возрастала более чем в 2 раза, достигая 12.15 мкг СО2–С г–1 ч–1, что может отражать стрессовое состояние микробоценоза почвы после пирогенного воздействия. В подзолистом горизонте подзолисто-глеевой среднесуглинистой почвы содержание Смик снижено, а в горизонте 5–10 см оно выше контроля на 60%. Величина БД минеральных горизонтов выше контроля в 1.5–2 раза.
Итак, в год после высокоинтенсивных пожаров в кедровниках заповедника не наблюдалось значительного снижения содержания микробной биомассы, интенсивность же микробного дыхания значительно возрастала. В сосняках лишайниково-зеленомошных Средней Сибири через сутки после высокоинтенсивных экспериментальных пожаров наблюдалось снижение Смик в подстилке в 8–10 раз и интенсивности БД – в 1.5–3 раза (Богородская, 2006; Богородская, Сорокин, 2006). В минеральных горизонтах почвы до 10 см аналогичные параметры также значительно снижались, что объясняется термическим воздействием огня: в процессе горения подстилки на поверхности развиваются высокие температуры, достигающие 900°С и более, а на глубине 10 см – не более 100–150°С, в верхнем 1 см минерального горизонта почвы температура не превышает 45°С, в большинстве случаев поднимается лишь на 2–3°С (Nearly et al., 1999; Валендик и др., 2001; Богородская, 2006).
Дополнительным буфером, предохраняющим почву от действия высоких температур, служит лесная подстилка (Курбатский, 1964). Если она имеет значительную мощность, как в изучаемых кедровниках, где вместе с напочвенным покровом она достигала более 25 см, при пожаре сгорает только ее часть, почва и нижняя часть подстилки при этом, как правило, остаются влажными. В такой ситуации прямой эффект на почву незначителен (Pietikainen, Fritze, 1995; Grasso et al., 1996; Валендик и др., 2001).
Через год после высокоинтенсивного пожара в кедровнике разнотравно-зеленомошном содержание Смик в подстилке ниже контроля в 2 раза, в минеральном горизонте 0–5 см – на 25%, тогда как скорость БД сравнима с контролем. В нижележащем горизонте 5–10 см скорость БД возрастала в 2 раза (рис. 1). После низкоинтенсивного пожара рассматриваемые величины в подстилке сравнимы с контролем, а в минеральном горизонте 0–5 см они выше, чем на контрольном участке в 2 раза.
Через два года после слабоинтенсивного пожара в производном хвойно-лиственном насаждении, представленном березняком зеленомошным, достоверных изменений параметров функциональной активности микробоценоза не выявлено, что согласуется с последствиями низкоинтенсивных контролируемых выжиганий в светлохвойных лесах Нижнего Приангарья, после которых экофизиологическое состояние микробных комплексов восстанавливалось в течение одного–двух лет (Богородская, 2006; Богородская, Сорокин, 2006; Богородская и др., 2011, 2016).
Через четыре года после низкоинтенсивного пожара в подстилке лиственничника кипрейного отмечено незначительное увеличение содержания Смик и повышенная на 50% от контроля скорость БД, тогда как после высокоинтенсивного пожара в подстилке лиственничника вейникового содержание Смик почти в 2 раза ниже контроля, что является следствием как значительного прогорания подстилок лиственничников, так и послепожарного изменения количества и качества растительного опада (Fritze et al., 1993; Pietikainen, Fritze, 1993, 1995; Certini, 2005; Sharma et al., 2017). В минеральных горизонтах подзолистой почвы содержания Смик после пожаров выше контроля в 1.3–2 раза, скорость БД – в 2 раза.
Значительное увеличение базального дыхания почвами было отмечено в первые месяцы и даже сутки после пожаров и связано с увеличением количества доступного органического вещества, минеральных элементов, доступных форм азота, увеличения рН почвы и прочих химических изменений, связанных с горением (Wuthrich et al., 2002; Bárcenas-Moreno et al., 2011; Masyagina et al., 2016). Высокие температуры во время горения оказывают стерилизующее действие на верхние органогенные горизонты почвы, но повторный засев может произойти очень быстро в течение нескольких суток после пожара за счет принесенных ветром спор и прочих остатков и вследствие вторжения из нижележащих горизонтов. Большое значение при этом имеет влага: увлажнение почвы и выпадение дождей после пожара оказывает благоприятное действие на повторный засев и увеличение популяций микроорганизмов (Vazquez et al., 1993; Hernandez et al., 1997; Bárcenas-Moreno et al., 2011; Sharma et al., 2017).
По мере исчерпания доступных субстратов биологическая активность почв после высокоинтенсивных пожаров снижается, и уже через год ее уровень значительно ниже, чем в почвах негоревших насаждений. Дальнейшее восстановление микробоценозов почв будет определяться послепожарными сукцессиями растительности и гидротермическими свойствами почв (Nearly et al., 1999; Богородская, 2006; Богородская и др., 2011; Bárcenas-Moreno et al., 2011; Holden et al., 2016; Cutler et al., 2017).
Через месяц после пожаров в хвойном лесу Северного Израиля количество общего растворимого азота в 10 см минерального горизонта почвы увеличивалось в 5 раз и вызывало значительное снижение содержания микробного азота и базального дыхания. По мере снижения количества общего растворимого азота уровень микробной биомассы восстанавливался до первоначального в течение двух лет (Ginzburg, Steinberger, 2012). В кедровнике разнотравно-зеленомошном через месяц после пожаров содержание общего азота снижалось (табл. 3), не оказывая ингибирующего действия на микробную активность.
Микробный метаболический коэффициент (qСО2) является важным индикатором эффективности использования субстрата и, тем самым, микробного стресса (Anderson, Domsch, 1993; Ананьева, 2003). В устойчивой “зрелой” почвенной экосистеме на поддержание единицы углерода микробной биомассы выделяется меньше СО2, чем в “юной” или нарушенной, что позволяет оценивать равновесное состояние и устойчивость микробоценоза почвы (Сусьян и др., 2009). Выявлено, что в подстилках исследуемых насаждений показатель qСО2 варьировал от 2.43 мкг СО2–С мг–1 Смик ч–1 в лиственничнике зеленомошном до 3.68 мкг СО2–С мг–1 Смик ч–1 в производном березняке зеленомошном (рис. 2). В верхнем минеральном горизонте 0–5 см почв контрольных насаждений его величины находятся в этом же диапазоне, а в нижележащем горизонте 5–10 см снижаются до 1.4–2.3 мкг СО2–С мг–1 Смик ч–1. Прохождение насаждений пожарами высокой и низкой интенсивности вызывало повышение величины коэффициента, причем максимальное увеличение наблюдалось после пожаров высокой интенсивности – в 2–3 раза в подстилке и верхнем горизонте 0–5 см минеральной части почвы (рис. 2). В нижележащем горизонте 5–10 см его значения после пожаров высокой интенсивности повышались не столь значительно, либо оставались на уровне контроля после низкоинтенсивных пожаров. Наблюдался тренд снижения значений qСО2 с течением времени, пройденного после пожара.
Рис. 2.
Микробный метаболический коэффициент в почвах хвойных насаждений, пройденных пожарами разной интенсивности. Обозначения см. рис. 1.
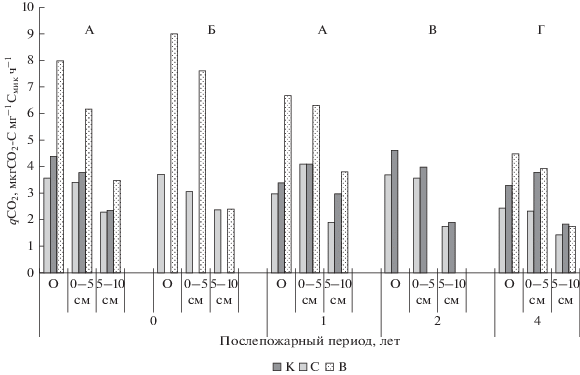
Увеличение значений микробного метаболического коэффициента, связанное с возрастанием дыхательной активности микрофлоры после пожаров, объясняется преобладанием на начальных этапах сукцессий R-стратегов в микробных популяциях, которые быстро используют ресурс и нуждаются в большом количестве энергии для поддержания своей биомассы (Pietikainen, Fritze, 1993; Wuthrich et al., 2002; Thiffault et al., 2008). Обогащение почвы легкоминерализуемыми источниками углерода и минеральными элементами, а также изменение гидротермического режима почв после пожаров вызывает повышение минерализационной активности и снижение устойчивости в почвенном микробном блоке. Дальнейшее послепожарное сукцессионное развитие микробоценозов направлено на преобладание медленнорастущих К-стратегов, полнее усваивающих субстрат. В итоге энергетический баланс системы выравнивается, что отражается в снижении значений qСО2 (Fritze et al., 1993).
Запасы углерода микробной биомассы и микробное продуцирование СО2 в профиле почв после пожаров
Максимум запасов микробной биомассы приходится на верхние 0–5 см почвенного профиля кедровых (34–50% от общих запасов Смик) и березового (46%) насаждений. В лиственничнике зеленомошном запас Смик выше в горизонте 5–10 см (рис. 3а). В подстилках запасы Смик оценивались в 3.5–8 г С м–3 в ненарушенных насаждениях, тогда как минеральный горизонт почвы до 10 см содержал 16–32 г С м–3. Наибольшее микробное продуцирование СО2 приходится на 0–5 см почв контрольных насаждений (28–58 мг СО2–С м–3 ч–1), что составляло 37% от общего в подзолистом горизонте кедровника разнотравно-зеленомошного и до 60% в гумусовом горизонте литозема темногумусового в березняке зеленомошном (рис. 3б). Подстилки выделяли 12–29 мг СО2–С м–3 ч–1, тогда как минеральный 10-сантиметровый горизонт почв – 46–86 мг СО2–С м–3 ч–1. Полученные данные по запасам микробной биомассы и продуцированию СО2 почвами изученных насаждений значительно ниже, чем в почвах южно- и среднетаежных лесов Европейской России (Сусьян и др., 2009; Стольникова и др., 2011) и в 3–10 раз ниже запасов Смик в гумусовом горизонте дерново-подзолистых почв светлохвойных лесов Средней Сибири (Богородская, Кукавская, 2016) и серых почв Красноярской лесостепи (Богородская и др., 2017).
Рис. 3.
Запасы углерода микробной биомассы (а) и продуцирование СО2 почвами (б) после пожаров разной интенсивности. Обозначения см. рис. 1.
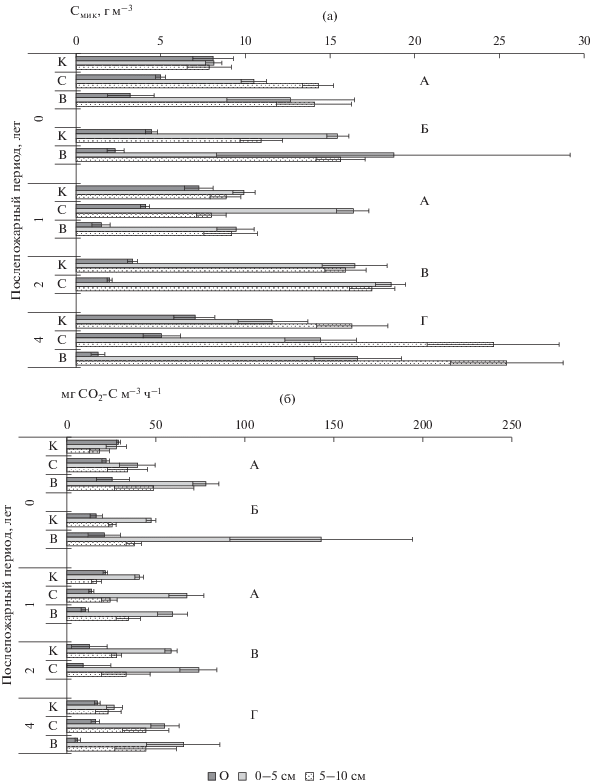
После пожаров в хвойных насаждениях заповедника происходит трансформация профильного распределения запасов Смик и микробной продукции углекислого газа (рис. 3). В первый год после пожаров в подстилках кедровников сокращается в 1.6–2.5 раза запас Смик, тогда как в верхних 0–5 см минеральной почвы он увеличивается на 20–60%, а в нижележащем – на 45–80%. Микробное продуцирование СО2 в подстилках мало изменяется, а в минеральных горизонтах увеличивается в 1.5–3 раза, особенно после высокоинтенсивных пожаров, достигая максимальной величины 143 мг СО2–С м–3 ч–1 в кедровнике лишайниково-зеленомошном.
Через год после высокоинтенсивного пожара в кедровнике разнотравно-зеленомошном запас Смик в подстилке снижен в 5 раз, тогда как в верхних горизонтах минеральной почвы сравним с контролем. После низкоинтенсивного пожара сохранена тенденция, отмеченная в первый год (рис. 3а). Микробное продуцирование СО2 снижается по сравнению с первым годом, но также выше контроля, особенно после высокоинтенсивного пожара (рис. 3б).
В березняке зеленомошном через два года после низкоинтенсивного пожара изменений в профильных запасах Смик и продукции СО2 не выявлено. В лиственничнике вейниковом спустя четыре года после пожара высокой интенсивности запас Смик в подстилке снижен в 5.5 раз, тогда как в минеральных горизонтах он выше в 1.5 раза вне зависимости от интенсивности пожаров. Микробная продукция СО2 также меньше контроля в 3 раза в подстилке после высокоинтенсивного пожара, а в минеральных горизонтах выше в 2–2.5 раза (рис. 3).
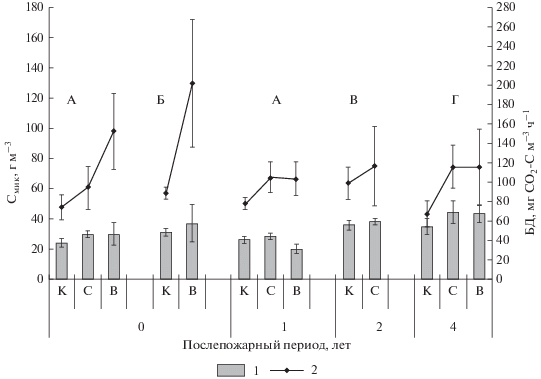
Суммарные запасы углерода микробной биомассы в профиле до 10 см в год после пожаров в кедровниках выше контроля на 20% вне зависимости от интенсивности горения, тогда как общая микробная продукция СО2 увеличивалась на 30% после пожара низкой интенсивности и более чем в 2 раза после высокоинтенсивных пожаров, достигая 200 СО2–С м–3 ч–1 в кедровнике лишайниково-зеленомошном (рис. 4). Уже через год после высокоинтенсивного пожара в кедровнике разнотравно-зеленомошном суммарный запас Смик снижался на 20% от контроля, а продуцирование СО2 увеличивалось на 30% после всех пожаров. В березняке через два года после низкоинтенсивного пожара рассматриваемые величины сравнимы с контролем, тогда как в лиственничнике вне зависимости от интенсивности пожаров через четыре года суммарный запас Смик выше контроля на 25%, а микробное продуцирование СО2 – почти в 2 раза (рис. 4).
Рис. 4.
Суммарные запасы углерода микробной биомассы (1) и общее микробное продуцирование СО2 (2) в профиле до 10 см почв после пожаров разной интенсивности. Обозначения см. рис. 1.
Изменение структуры и численности ЭКТГМ почв после пожаров
Наибольшая численность гетеротрофных микроорганизмов обнаружена в подстилках исследуемых насаждений, при этом доминирующее положение занимали микроорганизмы, использующие минеральные формы азота и олиготрофные группировки (рис. 5). При переходе от органогенных к минеральным горизонтам численность КОЕ разных групп микроорганизмов резко снижалась, что характерно для почв таежных экосистем.
Рис. 5.
Численность ЭКТГМ в подстилках (а), минеральном горизонте почв 0–5 см (б), 5–10 см (в) хвойных насаждений, пройденных пожарами разной интенсивности. 1 – аммонификаторы; 2 – прототрофы; 3 – олиготрофы; 4 – олигонитрофилы. Обозначения см. рис. 1.
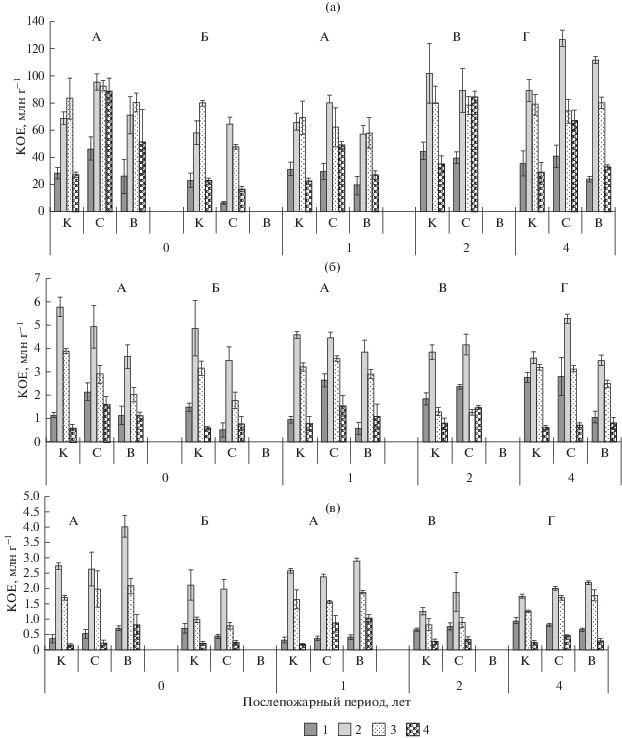
Биогенность верхнего минерального горизонта почв выше в березняке зеленомошном и лиственничнике лишайниково-зеленомошном, где отмечена высокая численность аммонификаторов и прототрофов, достигающая 2.8–3.9 млн КОЕ г–1, а олиготрофность почв низкая. В кедровых насаждениях преобладают олиготрофные микроорганизмы (рис. 5б).
Послепожарная трансформация структуры и численности ЭКТГМ затрагивает как подстилку, так и минеральные горизонты почв хвойных насаждений. В подстилке кедровника разнотравно-зеленомошного в год после пожара низкой интенсивности численность аммонификаторов увеличивалась в 2 раза, олигонитрофилов – в 3 раза, тогда как после пожара высокой интенсивности отмечено лишь достоверное увеличение численности олигонитрофилов (рис. 5). В подзолистом горизонте 0–5 см после низкоинтенсивного пожара численность КОЕ аммонификаторов и олигонитрофилов возрастала в 2–2.5 раза, в нижележащем она не изменялась. После высокоинтенсивного пожара, напротив, численность этих ЭКТГМ повышалась в 2 раза в минеральном горизонте 5–10 см.
В подстилке кедровника лишайниково-зеленомошного в год пожара численность КОЕ аммонификаторов снижена почти в 3.5 раза, олиготрофных групп – на 30–40%, при этом значительное преобладание микроорганизмов, требующих для роста минеральных источников азота, свидетельствует об активизации процессов минерализации пирогенного органического вещества. В минеральном горизонте почвы 0–5 см численность всех ЭКТГМ снижена в 1.5–3 раза, а в нижележащем сравнима с контролем. Вероятно, присутствие лишайника в напочвенном покрове, который обладает меньшей влажностью и лучше горит (Богородская, 2006), вызвало большее термическое влияние пожара на почву.
Через год после низкоинтенсивного пожара в подзолистом горизонте кедровника разнотравно-зеленомошного в 2–3 раза возрастала численность ЭКТГМ, олиготрофность почв снижалась. После высокоинтенсивного пожара численность ЭКТГМ была незначительно снижена в подстилке и верхнем минеральном горизонте 0–5 см, тогда как в нижележащем она возрастала (рис. 5в).
Увеличение численности гетеротрофных микроорганизмов после воздействия пожаров связано с созданием благоприятных гидротермических условий при прогревании достаточно влажной подстилки в совокупности с обогащением почвы легкогидролизуемым органическим веществом, азотом и зольными элементами (Fritze et al., 1993; Vazquez et al., 1993; Попова, 1997; Безкоровайная и др., 2005; De Marco et al., 2005; Thiffault et al., 2008; Sharma et al., 2017). Также после низкоинтенсивных пожаров в смешанных хвойных лесах отмечено возрастание аммонифицирующей микрофлоры и снижение олиготрофности почв (Yeager et al., 2005). Смена мохового покрова на травянистый предопределяет переход бактериального сообщества от олиготрофной к копитрофной стратегии жизнеобеспечения, предполагая улучшение условий роста микробных популяций в течение 20–30 лет после пожаров (Cutler et al., 2017).
Спустя 2–4 года после низкоинтенсивных пожаров в березняке и лиственничнике зеленомошных численность ЭКТГМ сравнима с контролем, тогда как через 4 г. после высокоинтенсивного пожара в подстилке и минеральном горизонте 0–5 см численность аммонификаторов снижена при повышенной олиготрофности почв, что является отражением качественного изменения органического вещества (Fritze et al., 1993; Thiffault et al., 2008; Yeager et al., 2005).
Заключение. Низовые пожары в кедровых, лиственничных и производных хвойно-лиственных насаждениях Средней Сибири приводили к увеличению функциональной активности, численности эколого-трофических групп микроорганизмов, запасов Смик и микробной продукции СО2 в верхних минеральных почвенных горизонтах, причем данный эффект более очевиден после высокоинтенсивных пожаров. В подстилках и в верхнем 0–5 см подзолистом горизонте почв кедровников через месяц после высокоинтенсивных пожаров наблюдалось увеличение значения qСО2 в 2–3 раза, тогда как низкоинтенсивные пожары в изучаемых насаждениях не вызвали значительного изменения устойчивости почвенного микробоценоза. С течением времени, пройденного после пожара, значения qСО2 снижаются. В подстилке лиственничника вейникового через четыре года после высокоинтенсивного пожара отмечено снижение уровня микробной биомассы (в 2 раза) и численности аммонификаторов, а также повышение олиготрофности. Отмеченная трансформация структуры, численности ЭКТГМ и параметров функциональной активности микробоценозов почв кедровых насаждений после высокоинтенсивных пожаров гораздо меньше, чем аналогичные изменения после средне- и высокоинтенсивных пожаров в южно- и среднетаежных сосняках лишайниково-зеленомошных с менее мощным напочвенным покровом, предохраняющим от негативного воздействия высоких температур при горении, и сопоставимы с изменениями в светлохвойных лесах названной лесорастительной зоны после низкоинтенсивных пожаров, где экофизиологическое состояние и структура микробных комплексов почв восстановились в течение одного–двух лет (Богородская, 2006; Богородская, Сорокин, 2006; Богородская и др., 2016). Степень трансформации микробоценоза почвы лиственничника зеленомошного через четыре года после высокоинтенсивного пожара сопоставима с аналогичными изменениями в смешанном насаждении с преобладанием лиственницы в Нижнем Приангарье, где нарушение гомеостатического состояния микробоценоза наблюдалось в течение трех лет после высокоинтенсивного пожара с дальнейшей тенденцией к уравновешиванию микробиологических показателей (Богородская и др., 2011). Послепожарная динамика направлена на снижение общей микробной продукции СО2 почвами, при этом даже спустя четыре года после пожаров в лиственничниках независимо от интенсивности она в 2 раза выше и отстает от уровня повышения общих запасов микробной биомассы. Преобладание минерализационных процессов в почвах кедровников и лиственничников в послепожарный период отражает смену эдафических и трофических условий почв и свидетельствует об активном вовлечении травяного опада и органического вещества в биотический круговорот.
Список литературы
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 222 с.
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1970. 487 с.
Барталев С.А., Стыценко Ф.В., Егоров В.А., Лупян Е.А. Спутниковая оценка гибели лесов России от пожаров // Лесоведение. 2015. № 2. С. 83–94.
Безкоровайная И.Н., Иванова Г.А., Тарасов П.А., Сорокин Н.Д., Богородская А.В., Иванов В.А., Конард С.Г., Макрае Д.Дж. Пирогенная трансформация почв сосняков средней тайги Красноярского края // Сибирский экологический журн. 2005. № 1. С. 143–152.
Богородская А.В. Влияние пожаров на микробные комплексы почв сосновых лесов Средней Сибири: Автореф. дис. … канд. биол. наук: 03.00.16, 03.00.07. Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 2006. 24 с.
Богородская А.В., Сорокин Н.Д. Микробиологическая диагностика состояния пирогенно-измененных почв сосняков Нижнего Приангарья // Почвоведение. 2006. № 10. С. 1258–1266.
Богородская А.В., Иванова Г.А., Тарасов П.А. Послепожарная трансформация микробных комплексов почв лиственничников Нижнего Приангарья // Почвоведение. 2011. № 1. С. 56–63.
Богородская А.В., Кукавская Е.А. Состояние микробных сообществ в почвах лиственных и светлохвойных лесов Средней Сибири после рубок и пожаров // Лесоведение. 2016. № 5. С. 383–396.
Богородская А.В., Пономарева Т.В., Ефимов Д.Ю., Шишикин А.С. Трансформация эколого-функциональных параметров микробоценозов почв на просеках линий электропередач в условиях Средней Сибири // Почвоведение. 2017. № 6. С. 731–743.
Валендик Э.Н., Векшин В.Н., Иванова Г.А., Кисиляхов Е.К., Перевозникова В.Д., Брюханов А.В., Бычков В.А., Верховец С.В. Контролируемые выжигания на вырубках в горных лесах. Новосибирск: Изд-во СО РАН, 2001. 172 с.
Горшков С.П., Ванденберг Дж., Алексеев Б.А., Мочалова О.И., Тишкова М.А. Климат, мерзлота и ландшафты Среднеенисейского региона. М.: Изд-во МГУ, 2003. 81 с.
Дымов А.А., Дубровский Ю.А., Габов Д.Н., Жангуров Е.В., Низовцев Н.А. Влияние пожара в северотаежном ельнике на органическое вещество почвы // Лесоведение. 2015. № 1. С. 52–62.
Иванов В.А., Иванова Г.А. Пожары от гроз в лесах Сибири. Новосибирск: Наука, 2010. 160 с.
Иванова Г.А., Конард С.Г., Макрае Д.Д., Безкоровайная И.Н., Богородская А.В., Жила С.В., Иванов В.А., Иванов А.В., Ковалева Н.М., Краснощекова Е.Н., Кукавская Е.А., Орешков Д.Н., Перевозникова В.Д., Самсонов Ю.Н., Сорокин Н.Д., Тарасов П.А., Цветков П.А., Шишикин А.С. Воздействие пожаров на компоненты экосистемы среднетаежных сосняков Сибири / Под ред. М.Д. Евдокименко. Новосибирск: Наука, 2014. 232 с.
Коротков И.А. Лесорастительное районирование России и республик бывшего СССР // Углерод в экосистемах лесов и болот России. Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 1994. С. 29–47.
Краснощеков Ю.Н., Чередникова Ю.С. Постпирогенная трансформация почв кедровых лесов в южном Прибайкалье // Почвоведение. 2012. № 10. С. 1057–1067.
Краснощеков Ю.Н., Евдокименко М.Д., Чередникова Ю.С. Лесоэкологические последствия пожаров в кедровниках Южного Прибайкалья // География и природные ресурсы. 2013. № 1. С. 33–42.
Кукавская Е.А., Буряк Л.В., Каленская О.П., Зарубин Д.С. Трансформация напочвенного покрова при низовых пожарах и оценка пирогенной эмиссии углерода в темнохвойных лесах Средней Сибири // Сибирский экологический журнал. 2017. № 1. С. 72–82.
Курбатский Н.П. Проблема лесных пожаров // Возникновение лесных пожаров. М.: Наука. 1964. С. 5–60.
Методы почвенной микробиологии и биохимии / Под ред. Д.Г. Звягинцева. М.: Изд-во Моск. университета, 1991. 304 с.
Мякина Н.Б., Аринушкина Е.В. Методическое пособие для чтения результатов химических анализов почв. М.: Изд-во МГУ, 1979. 62 с.
Попова Э.П. Пирогенная трансформация свойств лесных почв Среднего Приангарья // Сибирский экологический журнал. 1997. № 4. С. 413–418.
Седых В.Н. Динамика равнинных кедровых лесов Сибири. Новосибирск: Наука, 2014. 232 с.
Семечкин И.В., Поликарпов Н.П., Ирошников А.И., Бабинцева Р.М., Воробьев В.Н., Дашко Н.В., Иванов В.В., Кондаков Ю.П., Коротков И.А., Мурина Т.К., Назимова Д.И., Попов В.Е., Попова Ю.М., Соколов Г.А., Софронов М.А., Смагин В.Н., Спиридонов Б.С., Чебакова Н.М., Чередникова Ю.С. Кедровые леса Сибири / Под ред. А.С. Исаева. Новосибирск: Наука, 1985. 258 с.
Сорокин Н.Д. Влияние лесных пожаров на биологическую активность почв // Лесоведение. 1983. № 4. С. 24–28.
Софронов М.А., Голдаммер И.Г., Волокитина А.В., Софронова Т.М. Пожарная опасность в природных условиях. Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 2005. 330 с.
Стефин В.В. Антропогенные воздействия на горно-лесные почвы. Новосибирск: Наука, 1981. 169 с.
Стольникова Е.В., Ананьева Н.Д., Чернова О.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов Европейской территории России // Почвоведение. 2011. № 4. С. 479–494.
Сусьян Е.А., Ананьева Н.Д., Гавриленко Е.Г., Чернова О.В., Бобровский М.В. Углерод микробной биомассы в профиле лесных почв южной тайги // Почвоведение. 2009. № 10. С. 1233–1240.
Фуряев В.В. Роль пожаров в процессе лесообразования. Новосибирск: Наука, 1996. 253 с.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Anderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biology and Biochemistry. 1978. V. 10. № 3. P. 215–221.
Anderson T.-H., Domsch K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental condition, such as, pH, on the microbial biomass of forest soil // Soil Biology and Biochemistry. 1993. V. 25. № 3. P. 393–395.
Bárcenas-Moreno G., García-Orenes F., Mataix-Solera J., Mataix-Beneyto J., Bååth E. Soil microbial recolonisation after a fire in a Mediterranean forest // Biology and Fertility of Soils. 2011. V. 47. № 3. P. 261–272.
Certini G. Effects of fire on properties of forest soils: a review // Oecologia. 2005. V. 143. № 1. P. 1–10.
Cutler N.A., Arróniz-Crespo M., Street L.E., Jones D.L., Chaput D.L., Deluca T.H. Long-term recovery of microbial communities in the Boreal bryosphere following fire disturbance // Microbial Ecology. 2017. V. 73. № 1. P. 75–90.
De Marco A., Gentile A.E., Arena C., De Santo A.V. Organic matter, nutrient content and biological activity in burned and unburned soils of a Mediterranean maquis area of southern Italy // International J. Wildland Fire. 2005. V. 14. № 4. 14. P. 365–377.
Dooley S.R., Treseder K.K. The effect of fire on microbial biomass: a meta-analysis of field studies // Biogeochemistry. 2012. V. 109. № 1. P. 49–61.
Fritze H., Pennanen T., Pietikainen J. Recovery of soil microbial biomass and activity from prescribed burning // Canadian J. Forest Research. 1993. V. 23. № 7. P. 1286–1290.
Ginzburg O., Steinberger Y. Effects of forest wildfire on soil microbial-community activity and chemical components on a temporal-seasonal scale // Plant and Soil. 2012. V. 360. № 1. P. 243–257.
Grasso G.H., Ripabelli G., Sammareo M.L., Mazzoleni S. Effect of heating on the microbial population of grassland soil // The International Journal of Wildland Fire. 1996. V. 6. № 2. P. 67–70.
Hernandez T., Garcia C., Reinhardt I. Short-term effect of wildfire on the chemical, biochemical and microbiological properties of Mediterranean pine forest soils // Biology and Fertility of Soils. 1997. V. 25. № 2. P. 109–116.
Heydari M., Rostamy A., Najafi F., Dey D.C. Effect of fire severity on physical and biochemical soil properties in Zagros oak (Quercus brantii Lindl.) forests in Iran // Journal of Forestry Research. 2017. V. 28. № 1. P. 95–104.
Holden S.R., Rogers B.M., Treseder K.K., Randerson J.T. Fire severity influences the response of soil microbes to a boreal forest fire // Environmental Research Letters. 2016. V. 11. № 3. P. 035004.
Kukavskaya E.A., Buryak L.V., Ivanova G.A., Conard S.G., Kalenskaya O.P., Zhila S.V., McRae D.J. Influence of logging on the effects of wildfire in Siberia // Environmental Research Letters. 2013. V. 8. № 4. P. 045034.
Masyagina O.V., Tokareva I.V., Prokushkin A.S. Post fire organic matter biodegradation in permafrost soils: Case study after experimental heating of mineral horizons // Science of the Total Environment. 2016. V. 573. P. 1255–1264.
Nearly D.G., Klopatek C.C., DeBano L.F., Ffolliott P.F. Fire effects on bellowground sustainability: review and synthesis // Forest Ecology and Management. 1999. V. 122. № 1. P. 51–71.
Pietikainen J., Fritze H. Clear-cutting and prescribed burning in coniferous forest: comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification // Soil Biology and Biochemistry. 1995. V. 27. № 1. P. 101–109.
Pietikainen J., Fritze H. Microbial biomass and activity in the humus layer following burning: short-term effects of two different fires // Canadian Journal of Forest Research. 1993. V. 23. № 7. P. 1275–1285.
Santín C., Doerr S.H, Merino A., Bryant R., Loader N.J. Forest floor chemical transformations in a boreal forest fire and their correlations with temperature and heating duration // Geoderma. 2016. V. 264. Part A. P. 71–80.
Sharma U., Garima, Sharma J.C., Devi M. Effect of Forest fire on soil nitrogen mineralization and microbial biomass: A review // J. Pharmacognosy and Phytochemistry. 2017. V. 6. № 3. P. 682–685.
Thiffault E., Hannam K.D., Quideau S.A., Pare D., Belanger N., Oh S.-W., Munson A.D. Chemical composition of forest floor and consequences for nutrient availability after wildfire and harvesting in the boreal forest // Plant and Soil. 2008. V. 308. № 1–2. 308. P. 37–53.
Vazquez F.J., Acea M.J., Carhallas T. Soil microbial populations after wildfire // FEMS Microbiology Ecology. 1993. V. 13. № 2. P. 93–103.
Wardle D.A., Nilsson M.-C., Zackrisson O. Fire-derived charcoal causes loss of forest humus // Science. 2008. V. 320. № 5876. P. 629–629.
Wüthrich C., Schaub D., Weber M., Marxer P., Conedera M. Soil respiration and soil microbial biomass after fire in a sweet chestnut forest in southern Switzerland // Catena. 2002. V. 48. № 3. P. 201–215.
Yeager C.M., Northup D.E., Grow C.C., Barns S.M., Kuske C.R. Changes in nitrogen-fixing and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire // Applied and Environmental Microbiology. 2005. V. 71. № 5. P. 2713–2722.
Дополнительные материалы отсутствуют.