

Лесоведение, 2019, № 3, стр. 177-187
Идентификация видов берез в заболоченных лесах центра Русской равнины по результатам молекулярно-генетического анализа
А. А. Маслов a, *, О. Ю. Баранов b, А. А. Сирин a
a Институт лесоведения РАН
143030 Московская обл., п/о Успенское, ул. Советская, 21, Россия
b Институт леса НАН Беларуси
246001 Гомель, ул. Пролетарская, 71,
Беларусь
* E-mail: amaslov@ilan.ras.ru
Поступила в редакцию 26.03.2018
После доработки 23.04.2018
Принята к публикации 03.12.2018
Аннотация
Определен таксономический статус берез (Betula pendula, B. pubescens и их гибриды) на основе молекулярно-генетической оценки (по SSR-маркерам) уровня плоидности в сырых и болотных типах леса Центра Русской равнины на экспериментальных объектах Института лесоведения РАН. Согласно молекулярно-генетическому анализу, на гибриды B. pendula × B. pubescens в объединенной выборке приходится 11% деревьев. Таким образом, гибриды березы являются неотъемлемым компонентом природных сообществ. Впервые показано, что на олиготрофных торфяных болотах среди берез абсолютно преобладает B. pendula, гибриды редки, а B. pubescens встречается единично. В сыром сосняке-черничнике B. pendula входит в первый ярус древостоя, а нижний высотный полог сформирован деревьями B. pubescens и гибридами. Показано, что определение видов березы по ключам из определителей флоры часто приводит к ошибочным результатам, так как значения признаков встречаются в разных комбинациях. Для всех деревьев, отнесенных по результатам молекулярно-генетического анализа к B. pendula, был характерен тип кроны с ветвями “вверх”, без явных признаков “поникания”.
Березы – повислая (Betula pendula Roth) и пушистая (B. pubescens Ehrh.) – повсеместно произрастают в Центре Русской равнины как в виде чистых по составу насаждений, так и совместно с елью, сосной, осиной, липой, черной ольхой и другими породами. В Московской области березняки занимают первое место в структуре лесного фонда, на них приходится 41% от общей площади лесов региона и 37% – от общего запаса насаждений (Распределение площади лесов …, 2018). При почти повсеместном участии березы в составе ельников и сосняков по показателям запаса она существенно превосходит все остальные породы в большинстве областей Центра Русской равнины. Большую роль играет береза в составе болотных и заболоченных лесов – при том, что существенная часть региона характеризуется высокой заболоченностью (Вомперский и др., 2011; Sirin et al., 2017). В Московской области, где общая площадь торфяных болот и торфяников превышает 6% территории (Сирин и др., 2014), береза часто встречается на ненарушенных и антропогенно измененных зарастающих лесом торфяниках.
Морфологические, генетические, экологические различия между Betula pendula и B. pubescens подробно описаны в ботанической и лесоводственной литературе (Бородина и др., 1966; Шиманюк, 1974; Соколов и др., 1977; Атрохин и др., 1982; Абатуров и др., 1982; Лесная энциклопедия, 1985; Данченко, 1990; Ветчинникова, 2004; Денисов, 2009; Beck et al., 2016). Генетические различия выражаются в разном числе хромосом: у B. pendula 2n = 28 (диплоид), у B. pubescens 4n = 56 (тетраплоид). Морфологические различия проявляются в разной форме кроны (ветви – повисшие или направлены вверх), наличии или отсутствии “бородавок” на ветвях, опушенности листьев и побегов, форме листьев (треугольно-ромбические или яйцевидные). Отличаются виды березы и своей экологией. Считается, что B. pendula предпочитает более сухие местообитания и не выносит застойного увлажнения, тогда как B. pubescens хорошо мирится с высокой влажностью почвы и чаще растет на болотах. При этом сырые леса, согласно всем авторам, относятся к тем типам местообитаний, где оба вида произрастают совместно. С хозяйственной точки зрения наибольшую ценность в Центре Русской равнины представляют чистые березняки из B. pendula: их бонитет в среднем на два класса выше, чем у березняков из B. pubescens (Абатуров и др., 1982).
Несмотря на наличие относительно четких ключей во “флорах”, отнесение конкретных деревьев к B. pendula или B. pubescens, по нашему опыту и литературным данным, вызывает затруднение из-за неоднозначного сочетания диагностических признаков. Известно, что вышеуказанные виды образуют гибрид F1 B. pendula × B. pubescens (3n), но о его встречаемости в природе литературные сведения – противоречивы: от “массовой гибридизации” до “редкого явления” – (см. обзор в работе А.М. Данченко, 1990, с. 138). В российском лесоустройстве B. pendula и B. pubescens не различаются, и в материалах учета лесного фонда оба этих вида рассматриваются просто как “береза”. Как следствие, до настоящего времени отсутствуют объективные данные о встречаемости двух видов березы и гибрида F1 в разных типах местообитаний, включая заболоченные леса и болота. В частности, для авторов являлось вопросом, какие виды березы смогли успешно внедриться и постепенно выйти в первый ярус древостоя в сосняке на верховом олиготрофном болоте, для которого присутствие березы обычно не характерно (Маслов, 2001, 2007).
С разработкой новых информативных методов анализа плоидности на основе анализа микросателлитных спектров (без подсчета числа хромосом) появилась возможность быстрого и абсолютно надежного определения таксономического статуса образцов березы (Kulju et al., 2004; Падутов и др., 2007) и внедрения генетических методов в лесной экологии (Маслов, Сирин, 2015). В связи с довольно высокой стоимостью микросателлитного анализа в данной пилотной работе рассмотрен только таксономический статус деревьев березы на экспериментальных объектах Института лесоведения РАН (ИЛАН) – в нескольких заболоченных сосняках и на антропогенно измененных торфяниках с древостоями из березы и ольхи черной (Alnus glutinosa).
Цель работы – установить таксономический статус берез (B. pendula, B. pubescens), наличие или отсутствие их гибридов в заболоченных и иных типах леса (биотопах) на примере экспериментальных объектов ИЛАН с применением методов молекулярно-генетической диагностики. Предполагалось определить, какие таксоны и в какой пропорции произрастают в различных типах рассматриваемых лесов и в объединенной выборке, а также проверить информативность отдельных морфологических признаков в “ключах” из определителей флоры. Предварительные результаты работы уже предлагались для обсуждения (Маслов, Сирин, 2015, 2017).
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили на западе и на севере Московской области – на постоянных пробных площадях (пр. пл.) ИЛАН, приуроченных к заповедным лесным участкам (Маслов, 2009), и объектах мониторинга торфяных болот и антропогенно измененных торфяников (Суворов и др., 2015). Список изученных сообществ и биотопов разной степени заболоченности приводится в табл. 1.
Таблица 1.
Краткая характеристика объектов
| Тип леса/биотоп | Район Московской области | Номер пробной площади | Число образцов |
|---|---|---|---|
| Сосняк-зеленомошник | Клинский | 63–1 | 2 |
| Сосняк-черничник | Клинский | 63–2 | 10 |
| Сосняк-долгомошник | Дмитровский | 67–1 | 6 |
| Сосняк сфагновый | Одинцовский | 1–3 | 17 |
| Сосняк сфагновый | Талдомский | – | 3 |
| Черноольшаник | Талдомский | – | 3 |
| Кавальеры на осушенном торфяном болоте | Талдомский | – | 6 |
Градиент заболачивания представлен в выборке следующими типами леса: зеленомошник, черничник, долгомошник, сфагновый, черноольшаник (болотно-травяной), а также насаждениями из березы, возникшими на кавальерах (насыпях) вдоль дренажных каналов на осушенных торфяных болотах. Подробная характеристика условий произрастания и состава древостоев на постоянных пробных площадях дана в монографии А.А. Маслова (1990). В изученных сосняках береза в составе древостоев (по запасу) представлена единично; отдельные особи выходят в первый ярус, другие присутствуют в нижнем пологе древостоя. Тип леса “сфагновый” в Одинцовском и Талдомском районах представляет собой олиготрофное верховое болото (кустарничково-пушицево-сфагновое) с доминированием сосны. В Одинцовском районе на территории заказника Звенигородской биологической станции МГУ (пр. пл. 1–3) береза за 30 лет наблюдений вышла в первый ярус. Черноольшаник на низинном болоте в составе имеет значительную примесь березы. Насаждения на осушенных торфяных болотах представляют собой практически чистые древостои из березы.
Модельные деревья березы в каждом типе леса отбирали регулярным способом по общему пронумерованному списку. Для каждой модели проводили: измерение высоты и диаметра ствола на высоте 1.3 м, фотографирование кроны и ствола дерева, отбор одного двухлетнего побега (телескопическим сучкорезом, по возможности – из середины кроны). У деревьев с очень высокой кроной в качестве образцов отбирали порослевые (водяные) побеги, весьма характерные для березы в изученных типах леса. В камеральных условиях побеги сканировали для получения выборки по размеру и форме листьев, определяли наличие “бородавок” и опушения побегов (анализ морфологических признаков).
Образцы листьев на ДНК-анализ готовили в виде трех круглых высечек в пробирках 1.5 мл с фиксацией в 70%-м растворе спирта. До проведения молекулярно-генетического анализа образцы хранили при –18°С. Всего подготовлено 47 образцов.
Разделение образцов по плоидности на три группы генотипов – Betula pendula (2n), B. pubescens (4n) и B. pendula × B. pubescens (3n) – выполнялось на основании протокола LOH-анализа для генетического анализатора ABI PRISM 310 (Applied Biosystems, США). В качестве маркеров использованы семь полиморфных SSR-локусов берез – L2.2, L1.10, L7.8, L022, L7.3, L5.4, L3.1. Выбор данных маркеров обусловлен их высоким уровнем изменчивости на основании предварительного анализа показателей полиморфизма (Kulju et al., 2004; Падутов и др., 2007).
В ходе проведенного анализа изучены микросателлитные спектры образцов с различной плоидностью (диплоиды, триплоиды, тетраплоиды, миксоплоиды). Наличие не более двух вариантов аллелей (x, y) соответствовало B. pendula, трех аллелей (x, y, z) – гибридным генотипам (триплоидам), четырех (x, y, z, q) – B. pubescens. Общее заключение об уровне плоидности образцов составлялось на основании результатов анализа всех семи SSR-локусов, представленных в наборе (Баранов, Балюцкас, 2009). Дополнительный количественный анализ SSR-спектров позволил не только подтвердить результаты определения таксономического статуса, основанные на анализе числа аллельных вариантов, но и выявить наличие миксоплоидных тканей, что весьма типично для березы (Буторина, 1985; Машкина и др., 2013).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Соотношение таксонов
Распределение таксонов березы по типам леса (биотопам) и в пределах всей выборки деревьев дано в табл. 2. В объединенной выборке (47 деревьев) по результатам молекулярно-генетического анализа преобладают особи B. pendula (76%). Особи B. pubescens составляют 13%. На гибриды B. pendula × B. pubescens приходится 11%. Конечно, “объединенная выборка” – понятие очень условное – с учетом различий числа деревьев, отобранных в разных типах леса и субъективного выбора собственно типов и биотопов. Однако эти данные ясно свидетельствуют: гибриды B. pendula × × B. pubescens совсем нередко произрастают в природе и могут встречаться в разных типах леса (табл. 2).
Таблица 2.
Соотношение таксонов березы (число деревьев) в разных типах леса (биотопах) по результатам молекулярно-генетического анализа
| Тип леса/биотоп | B. pendula | B. pubescens | B. pendula × B. pubescens |
|---|---|---|---|
| Зеленомошник | 2 | 0 | 0 |
| Черничник | 5 | 3 | 2 |
| Долгомошник | 6 | 0 | 0 |
| Сфагновый (Одинцовский район) | 14 | 1 | 2 |
| Сфагновый (Талдомский район) | 3 | 0 | 0 |
| Черноольшаник | 0 | 2 | 1 |
| Кавальеры на осушенном торфяном болоте | 6 | 0 | 0 |
| Итого: | 36 | 6 | 5 |
Результаты молекулярно-генетической идентификации видов показали, что оба вида берез и гибрид имеют широкий эколого-фитоценотический диапазон. Присутствие B. pendula установлено во всех сообществах, кроме черноольшаника; однако наличие в черноольшанике гибрида B. pendula × B. pubescens (при крайне малом размере выборки) не исключает возможность произрастания B. pendula и в этом типе леса. В березовых насаждениях вдоль дренажных каналов на осушенных торфяниках также преобладает B. pendula: береза выросла здесь на переходных и низинных торфах – вышележащие верховые слои залежи были “сработаны” ранее при добыче торфа. Деревья B. pubescens и гибрида произрастают вместе в трех (из семи) типах леса (табл. 2), преобладая на черноольховом низинном болоте.
Соотношение таксонов березы в трех типах сосняков (с относительно бóльшим размером выборки) показано на рис. 1. Сообщества расположены в порядке увеличения влажности почвы. В черничнике (сыром типе леса с наличием сфагнума), как оказалось, присутствуют все три таксона – с заметным участием в древостое B. pubescens и гибрида, однако с преобладанием B. pendula. Наличие данных по высоте каждого дерева позволило оценить участие разных таксонов в составе высотных ярусов. Средняя высота деревьев B. pendula составила в черничнике 24 м, а высота B. pubescens и гибрида – 13.3 и 12.5 м, соответственно. Таким образом, деревья B. pubescens и гибриды формируют в черничнике нижний ярус древостоя, а не подрост, как считалось ранее, тогда как B. pendula присутствует в первом ярусе. Если объединить B. pubescens и гибриды в одну группу, то по критерию Манна–Уитни высота деревьев этой группы и высота деревьев B. pendula различаются на уровне значимости 5% (P = 0.05).
Рис. 1.
Соотношение таксонов березы в трех типах сосняков по результатам молекулярно-генетического анализа. 1 – Betula pendula, 2 – B. pubescens, 3 – B. pendula × B. pubescens.
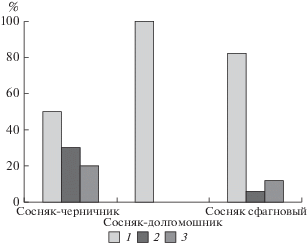
В сфагновом типе на олиготрофном верховом болоте довольно большой размер выборки при проведении молекулярно-генетической идентификации позволил наиболее точно определить соотношение между таксонами. Абсолютно преобладает в сообществе B. pendula; B. pubescens и гибрид присутствуют здесь единично (рис. 1). Разницы между таксонами по высоте ствола (с учетом близкого возраста всех деревьев) здесь обнаружить не удалось.
В занимающем среднее положение (по влажности почвы) долгомошнике, как оказалось, присутствуют только особи B. pendula (рис. 1). Возможно, это связано с локальными факторами, малым размером выборки или иными причинами.
Морфологические различия таксонов
Объективное разделение деревьев березы на три таксона, согласно результатам молекулярно-генетического анализа, позволяет оценить информативность известных диагностических признаков, которые применяются “в поле” при отнесении конкретных особей к тому или иному виду. К таким признакам почти все руководства относят: форму кроны, форму листа, опушение листьев и молодых побегов, наличие “бородавок” на побегах. Данные показатели определялись нами для дерева в целом (крона), а также побегов и листьев, отобранных в качестве образцов.
По форме кроны абсолютно все деревья во всех типах леса и биотопах, по данным анализа фотографий, имели ветви, направленные вверх. Это относится и к особям Betula pubescens, и к гибридам, и к особям B. pendula. Лишь у отдельных деревьев B. pendula единичные ветви III порядка имели признаки “повисания”. Фотография типичной кроны B. pendula в долгомошном типе леса приведена на рис. 2а.
Рис. 2.
Типичная форма кроны Betula pendula в изученных типах леса (а) и по литературным данным (б). а – фотография сделана в сосняке-долгомошнике 13.06.2015 (пр. пл. 67-1, береза № 116); б – источник: “Лесная энциклопедия” (1985), с. 68.

В отличие от формы кроны, параметры формы листьев внутри трех таксонов сильно варьируют. Это касается основания листовой пластинки, длины черешка и “оттянутости” вершинки. Типичная форма листа Betula pendula (в пределах изученной выборки) показана на рис. 3б. Однако листья B. pendula крайне редко имели клиновидное основание, подобно показанному на рис. 3а. Анализ возможностей диагностики видов березы по измерениям формы листа будет предпринят нами в дальнейшем. Наличие “бородавок” на стебле молодых побегов отмечено у большинства (но далеко не у всех) особей Betula pendula. Аналогичная ситуация отмечена и с опушением у особей B. pubescens.
Рис. 3.
Форма листа березы. а – Betula pendula, источник: “Лесная энциклопедия” (1985, с. 68); б – скан листа Betula pendula № 134 в сосняке-черничнике; в – Betula pubescens, источник: И.А. Губанов и др. (2003, с. 29).

Betula pendula и B. pubescens, с одной стороны, имеют четкие систематические признаки, но в то же время весьма полиморфны (Махнев, 1987; Данченко, 1990; Atkinson, 1992; Howland et al., 1995; Ветчинникова, 2004; Maliouchenko et al., 2007; Коропачинский, 2013). В составе природных популяций широко распространены деревья с промежуточными признаками, которые часто считали “гибридами”. В наиболее популярном справочнике-определителе середины XX века “Деревья и кустарники СССР” отмечается, что “у гибридов часто настолько перепутаны признаки обоих видов, что даже лесоводы-ботаники затрудняются иногда решить, к какому виду отнести то или другое дерево” (Бородина и др., 1966, с. 300). Поэтому при изучении березы и березняков в природе лишь немногие авторы называли виды березы или давали весьма приближенную оценку их участия в древостоях (Абатуров и др., 1982). В результате мы до сих не знаем о реальном соотношении Betula pendula, B. pubescens и гибрида B. pendula × B. pubescens в природных сообществах региона, так как определение видов (в тех случаях, когда оно проводилось) осуществлялось по морфологическим признакам согласно определителям.
Как оказалось, определение видов березы по ключам определителей флоры часто приводит к ошибочным результатам, поскольку значения диагностических признаков встречаются в разных комбинациях. Например, деревья березы на верховом болоте в сфагновом типе леса по строению кроны и форме листьев ранее были отнесены нами к Betula pubescens (Маслов, 2001, 2007). Результаты молекулярно-генетической диагностики показали, что это не так, и на болоте преобладают деревья B. pendula (табл. 2).
“Повислая” форма кроны – один из самых пародоксальных диагностических признаков – с учетом того, что прилагательное “повислая” присутствует в самом названии Betula pendula – как на русском, так и на латинском языке. Повислая крона показана как типичная на иллюстрации к “Лесной энциклопедии” (1985) (рис. 2б). Однако все изученные нами деревья березы имели кроны с ветвями “вверх”, включая все особи, отнесенные к B. pendula. Возможно, деревья березы “с поникающими тонкими ветвями” (Иллюстрированный определитель …, 2003, с. 31), действительно, однозначно диагностируют B. pendula. Однако в природных лесных популяциях у деревьев данного вида абсолютно преобладает “непоникающая” форма кроны (рис. 2а), и этот факт необходимо учитывать. Аналогично “треугольно-ромбические” листья (рис. 3а) не могут считаться необходимым диагностическим признаком Betula pendula (см., например, Маевский, 1964, с. 177). Типичная форма листьев B. pendula в изученных типах леса показана на рис. 3б. По визуальной оценке, она занимает промежуточное положение, между крайними формами листа деревьев, однозначно отнесенных к Betula pendula и B. pubescens (рис. 3а, 3в).
Вопрос возможности разделения деревьев березы на Betula pendula и B. pubescens по результатам морфометрического анализа листьев имеет важное практическое значение, так как молекулярно-генетический метод пока остается достаточно сложной и относительно дорогой процедурой. Успешные результаты по разделению видов по форме листа были получены путем применения дискриминантных методов (Русанович, Скворцов, 1981; Atkinson, Colding, 1986). Однако в данных работах отсутствуют указания на то, каким образом эталонные образцы относились к тому или иному виду березы. Вероятно, выделение эталонов проводилось по совокупности морфологических признаков, что априори закладывает возможность ошибки. Кроме того, в указанных двух работах ничего не сказано о гибридах, которые вполне могли присутствовать в анализируемой выборке. Разделение образцов по результатам молекулярно-генетической идентификации (Балюцкас и др., 2010; Wang et al., 2014) позволит корректнее оценить возможности разделения видов на основе дискриминантных методов.
Схема границ эколого-фитоценотических ареалов Betula pendula и B. pubescens в Центре Русской равнины и положение изученных типов леса показаны на рис. 4. По мнению Ю.Д. Абатурова с соавт. (1982, с. 14), березняки с преобладанием B. pendula, как правило, занимают относительно сухие и хорошо дренированные эдатопы; наоборот, в условиях избыточного увлажнения и неустойчивой аэрации доминирует B. pubescens. Заболоченные леса и торфяники на схеме относятся к зоне господства B. pubescens (рис. 4). Данное мнение является общепринятым и абсолютно преобладает в литературе. Приведем еще одно свидетельство со стороны известного флориста и систематика рода Betula А.К. Скворцова: “Чем сырее местообитание и чем менее проточно увлажнение, тем больше преобладает B. pubescens и отступает В. pendula. Наконец, на малопроточных эутрофных или переходных болотах и в заболоченных лесах остается только B. pubescens” (Соколов и др., 1977, с. 98).
Рис. 4.
Схема границ эдафических ареалов Betula pendula и B. pubescens в координатах эколого-фитоценотических рядов В.Н. Сукачева (1938), по: (Абатуров и др., 1982, с. 15), с изменениями. Горизонтальной штриховкой показана зона господства B. pendula (1), вертикальной – зона господства B. pubescens (2). Обозначения типов леса: з – зеленомошник, ч – черничник, д – долгомошник, сф – сфагновый, чол – черноольшаник, к – кисличник, н – неморальный.
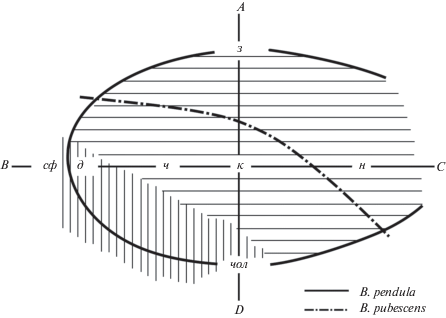
Материалы генетико-таксономической идентификации (табл. 2, рис. 1) одновременно и подтверждают, и опровергают традиционные представления. Вполне ожидаемым можно считать преобладание Betula pendula в зеленомошнике и B. pubescens – в черноольшанике. Хорошо согласуется со схемой, представленной на рис. 4, почти равное участие двух видов в черничнике. Подтверждается мнение, что в смешанных насаждениях черничных типов леса B. pubescens имеет более низкий бонитет и занимает более низкий вертикальный полог (Абатуров и др., 1982; Денисов, 2009). Не противоречит устоявшимся представлениям и факт преобладания В. pendula на кавальерах осушенных торфяных болот с благоприятным режимом аэрации и нередким наличием минеральных добавок в торфе.
Неожиданным результатом, по данным молекулярно-генетического анализа, стала таксономическая структура древостоя березы в сфагновых типах леса. Оказалось, что на олиготрофных верховых болотах (при наличии в древостое или подросте березы) абсолютно преобладают особи Betula pendula, гибриды редки, а деревья B. pubescens встречаются единично (табл. 2, рис. 1). Преобладание B. pendula на верховом торфяном болоте установлено для двух достаточно удаленных точек – в Одинцовском и Талдомском районах. На пр. пл. 1–3 в Одинцовском районе B. pendula не только смогла внедриться в сообщество сосняка кустарниково-пушицево-сфагнового после частичного распада соснового древостоя, но и за 30 лет вышла в первый ярус. Не соответствует распространенному представлению и факт доминирования B. pendula в долгомошном типе леса.
Результаты исследования впервые дают представление о встречаемости гибридов B. pendula × × B. pubescens в лесных сообществах Центра Русской равнины. Несмотря на то, что по морфологическим признакам большинство деревьев объединенной выборки можно отнести к переходной форме между двумя видами березы, согласно результатам молекулярно-генетического анализа гибриды B. pendula × B. pubescens составили 11% от всех деревьев. Конечно, этот процент зависит от выбора типов леса и размера выборки в конкретных сообществах. Но в целом результаты работы показывают, что гибриды березы являются неотъемлемым компонентом природных сообществ, и это необходимо иметь в виду при изучении популяций березы. По морфологическим признакам гибридные особи больше напоминают Betula pubescens – как было показано в работе I.R. Brown с соавт. (1982). Это подтверждается нашими данными, включая и характер роста в высоту гибридных особей в черничном типе леса.
Заключение. Новые эффективные и информативные методы анализа плоидности на основе SSR–анализа ДНК дают возможность быстрого и надежного определения таксономического статуса образцов березы и внедрения генетических методов в лесной экологии. В результате работы установлен процент присутствия разных таксонов березы в природных сообществах заболоченных и болотных типов леса Центра Русской равнины. Впервые показано, что на олиготрофных торфяных болотах среди берез абсолютно преобладает Betula pendula, гибриды редки, а B. pubescens встречается единично. Заметное участие всех трех таксонов показано для сырого черничного типа леса; при этом B. pendula формирует верхний ярус древостоя, а B. pubescens и гибриды произрастают в нижнем пологе.
Несмотря на то, что по морфологическим признакам большинство деревьев объединенной выборки можно отнести к переходной форме между двумя видами березы, согласно результатам молекулярно-генетического анализа, гибриды B. pendula × B. pubescens составили 11% от всех деревьев. Таким образом, нечеткость морфологических признаков видов формируется не в результате массовой гибридизации, а является следствием высокой полиморфности и B. pendula, и B. pubescens. С другой стороны, впервые достоверно показано, что в заболоченных и болотных типах леса региона гибриды березы являются неотъемлемым компонентом природных сообществ.
Как оказалось, определение видов березы по ключам из определителей флоры часто приводит к ошибочным результатам, поскольку значения диагностических признаков встречаются в разных комбинациях. Для всех деревьев, отнесенных по результатам молекулярно-генетического анализа к B. pendula, был характерен тип кроны с ветвями, направленными вверх, без явных признаков “поникания”. Ошибочным оказалось типичное представление о березе повислой как о дереве с “поникающей” кроной. Достаточно редко у особей, отнесенных к данному виду, встречались и листья треугольно-ромбической формы, как это описано во многих определителях.
Анализ встречаемости таксонов березы в заболоченных типах леса, рассмотренный в данной работе, основан на небольшом числе конкретных примеров и может считаться предварительным. Для более детального изучения проблемы потребуется взятие большого числа образцов в различных типах сообществ.
Авторы выражают благодарность Н.Г. Улановой за ценные советы в процессе работы над публикацией.
Список литературы
Абатуров Ю.Д., Зворыкина К.В., Ильюшенко А.Ф. Типы березовых лесов центральной части южной тайги. М.: Наука, 1982. 156 с.
Атрохин В.Г., Калуцкий К.К., Тюриков Ф.Т. Древесные породы мира. Т. 3. Древесные породы СССР. М.: Лесн. пром-сть, 1982. 264 с.
Балюцкас В., Балюцкене А., Баранов О.Ю. Морфолого-генетическая характеристика межвидовых гибридов (B. pendula × pubescens) в полусибсовом потомстве березы повислой // Наука о лесе XXI века: Матер. Междунар. научно-практической конф. (17–19 ноября 2010 г., Гомель). Гомель: Институт леса НАН Беларуси, 2010. С. 130–134.
Баранов О.Ю., Балюцкас В. Использование молекулярно-генетических маркеров для анализа плоидности осины и березы // Проблемы лесоведения и лесоводства. Гомель: Институт леса НАН Беларуси, 2009. Вып. 69. С. 129–135.
Бородина Н.А., Некрасов В.И., Некрасова Н.С., Петрова И.П., Плотникова Л.С., Смирнова Н.Г. Деревья и кустарники СССР. М.: Мысль, 1966. 637 с.
Буторина А.К. Цитогенетика хозяйственно-ценных форм карельской березы // Генетика, 1985. Т. 21. № 7. С. 1192–1198.
Ветчинникова Л.В. Береза: вопросы изменчивости (морфо-физиологические и биохимические аспекты). М.: Наука, 2004. 183 с.
Вомперский С.Э., Сирин А.А., Сальников А.А., Цыганова О.П., Валяева Н.А. Оценка площади болотных и заболоченных лесов России // Лесоведение. 2011. № 5. С. 3–11.
Данченко А.М. Популяционная изменчивость березы. Новосибирск: Наука, 1990. 205 с.
Денисов С.А. Динамика формационной структуры березняков в связи с их онтогенезом в различных эдатопах в Среднем Поволжье // Вестник Марийского государственного технического университета. Серия: Лес. Экология. Природопользование, 2009. № 3. С. 13–27.
Губанов И.А., Киселёва К.В., Новиков В.С., Тихомиров В.Н. Иллюстрированный определитель растений Средней России. Т. 2: Покрытосеменные (двудольные: раздельнолепестные). М.: Товарищество научных изданий КМК. 2003. 665 с.
Коропачинский И.Ю. Естественная гибридизация и проблемы систематики берез Северной Азии // Сибирский экологический журн. 2013. Т. 20. № 4. С. 459–479.
Лесная энциклопедия. М.: Советская энциклопедия. 1985. Т. 1. 563 с.
Маевский П.Ф. Флора средней полосы европейской части СССР. Изд. 9-е. Л.: Колос, 1964. 880 с.
Маслов А.А. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука, 1990. 160 с.
Маслов А.А. Динамика древостоя и нижних ярусов на олиготрофном лесном болоте близ Звенигорода: концептуальная модель процессов в масштабе десятилетий // Бюллетень МОИП. Отдел биологический. 2001. Т. 106. № 4. С. 71–77.
Маслов А.А. Механизмы динамики лесного сообщества в фазе климакса на примере олиготрофного сосняка пушицево-сфагнового // Актуальные проблемы геоботаники. III Всероссийская школа-конференция. Лекции. Петрозаводск: Изд-во Карельского НЦ РАН, 2007. С. 349–364.
Маслов А.А. Мониторинг биоразнообразия и процессов природной динамики в заповедных лесных участках: программа и итоги работ за 25 лет // Структура и функции лесов Европейской России. М.: Товарищество научных изданий КМК, 2009. С. 172–190.
Маслов А.А., Сирин А.А. Определение видов березы по морфологическим признакам и путем анализа ДНК // Наука – инновационному развитию лесного хозяйства. Гомель: Институт леса НАН Беларуси, 2015. С. 346–348.
Маслов А.А., Сирин А.А. Береза повислая и береза пушистая в заболоченных биотопах: традиционный подход и современный ДНК-анализ // Генетика популяций: прогресс и перспективы. Матер. Междунар. научной конференции (17–21 апреля 2017 г., Звенигородская биологическая станция им. С.Н. Скадовского Биологического факультета МГУ им. М.В. Ломоносова). М.: Ваш Формат, 2017. С. 163–165.
Махнев А.К. Внутривидовая изменчивость и популяционная структура берез секции Albae и Nanae. М.: Наука, 1987. 129 с.
Машкина О.С., Табацкая Т.М., Баранов О.Ю., Зеленина Е.А. Цитогенетические и молекулярно-генетические особенности коллекции ценных генотипов березы, длительно культивируемой в условиях in vitro // Биология клеток растений in vitro и биотехнология: Матер. X Междунар. конференции (14–18 октября 2013 г., Казань). Казань: Казанский институт биохимии и биофизики Казанского НЦ РАН, 2013. С. 301.
Падутов В.Е., Баранов О.Ю., Воропаев Е.В. Методы молекулярно-генетического анализа. Минск: Юнипол, 2007. 176 с.
Распределение площади лесов и запасов древесины по преобладающим породам и группам возраста. М. Комитет лесного хозяйства Московской области, 2018. –электронный ресурс: URL: klh.mosreg.ru/dokumenty/otkrytye-dannye/raspredelenie-ploshchadi-lesov-i-zapasov-drev/12-02-2018-09-30-00-forma-2-glr-na-01-01-2017. Дата обращения: 20.08.2018.
Русанович И.И., Скворцов А.К. Использование дискриминантного анализа для нахождения разграничительных видовых характеристик формы листа берез // Журн. общей биологии. 1981. Т. 42. № 5. С. 762–770.
Сирин А.А., Маслов А.А., Валяева Н.А., Цыганова О.П., Глухова Т.В. Картографирование торфяных болот Московской области по данным космической съемки высокого разрешения // Лесоведение. 2014. № 5. С. 65–71.
Соколов С.Я., Связева О.А., Кубли В.А., при участии Скворцова А.К., Грудзинской И.А., Огуреевой Г.Н. Тиссовые – Кирказоновые. Т. 1. Л.: Наука, 1977. 164 с.
Суворов Г.Г., Чистотин М.В., Сирин А.А. Потери углерода при добыче торфа и сельскохозяйственном использовании осушенного торфяника в Московской области // Агрохимия. 2015. № 11. С. 51–62.
Шиманюк А.П. Дендрология. Изд. 2-е М.: Лесн. пром-сть, 1974. 264 с.
Atkinson M.D. Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. // Journal of Ecology. 1992. V. 80. № 4. P. 837–870.
Atkinson M.D., Codling A.N. A reliable method for distinguishing between Betula pendula and B. pubescens // Watsonia. 1986. V. 16. № 1. P. 75–76.
Beck P., Caudullo G., de Rigo D., Tinner W. Betula pendula, Betula pubescens and other birches in Europe: distribution, habitat, usage and threats // European atlas of forest tree species. Luxembourg: Publications Office of the European Union, 2016. P. 70–73.
Brown I.R., Kennedy D., Williams D.A. The occurrence of natural hybrids between Betula pendula Roth and B. pubescens Ehrh. // Watsonia. 1982. V. 14. P. 133–145.
Howland D., Oliver R., Davy A. Morphological and molecular variation in natural populations of Betula // New Phytologist, 1995. V. 130. P. 117–124.
Kulju K.K.M., Pekkinen M., Varvio S. Twenty-three microsatellite primer pairs for Betula pendula (Betulaceae) // Molecular Ecology Notes. 2004. V. 4. № 3. P. 471–473.
Maliouchenko O., Palmé A.E., Buonamici A., Vendramin G.G., Lascoux M. Comparative phylogeography and population structure of European Betula species, with particular focus on B. pendula and B. pubescens // J. Biogeography. 2007. V. 34. № 9. P. 1601–1610.
Sirin A., Minayeva T., Yurkovskaya T., Kuznetsov O., Smagin V., Fedotov Yu. Russian Federation (European Part) // Mires and peatlands of Europe: Status, distribution and conservation. Stuttgart: Schweizerbart Science Publishers, 2017. P. 589–616.
Wang N., Borrell J., Buggs R. Is the Atkinson discriminant function a reliable method for distinguishing between Betula pendula and B. pubescens (Betulaceae)? // New J. Botany. 2014. V. 4. P. 90–94.
Дополнительные материалы отсутствуют.