

Лесоведение, 2019, № 3, стр. 208-214
Популяционная изменчивость ели финской по форме семенных чешуй на севере Архангельской области
С. Н. Тарханов *
Федеральный исследовательский центр комплексного изучения Арктики РАН
163000 Архангельск, Наб. Северной Двины, 23,
Россия
* E-mail: tarkse@yandex.ru
Поступила в редакцию 16.04.2015
После доработки 04.04.2017
Принята к публикации 05.06.2018
Аннотация
Изучена изменчивость ели (Picea obovata Ledeb. × Picea abies (L.) Karst.) по морфометрическим параметрам зрелых шишек и показателям формы семенных чешуй в бассейне рек Северной Двины, Онеги и Пинеги на севере Архангельской области. Установлено доминирование по численности (70–95%) в северотаежных популяциях формы “сибирского” типа и близкой к ней формы “промежуточного” типа. Не выявлено существенных различий ели с разным типом семенных чешуй (в спелом возрасте) по размерам ствола, кроны, численности боковых побегов и почек, продолжительности жизни хвои. Более северные (64°–65° с.ш.) и восточные (43°–44° в.д.) популяции ели существенно уступают более южным (63° с.ш.) и западным (38° в.д.) популяциям по длине и сухой массе шишек. Линейные связи морфометрических параметров зрелых шишек с показателем формы (коэффициентом вытянутости) семенных чешуй внутри популяции отсутствуют.
Виды ели, в том числе ель европейская и ель сибирская, подверглись сильной интрогрессивной гибридизации. Биологический смысл данного явления заключается в воспроизведении естественных гибридов двух контактирующих видов путем возвратных скрещиваний с одним или обоими родительскими видами, что приводит к переносу генов от одного из них к другому и преодолению барьеров, препятствующих скрещиванию (Anderson, 1949; Бобров, 1971, 1972; Грант, 1984). Обобщив значительный материал по изменчивости популяций ели, Л.Ф. Правдин (1975) выделил три зоны: с безраздельным господством ели сибирской; господства ели европейской; интрогрессивной гибридизации ели сибирской и ели европейской. Последняя занимает широкую полосу Восточно-Европейской равнины – от Уральских гор до Среднерусской возвышенности и представлена симпатрическими популяциями гибридных форм, описанных ранее под названием P. × fennica Rql. (Kоm.), P. × uralensis Tepl., P. × medioxima Nyl., P. × uwarоwii Kaufm. Эти гибридные формы были объединены под общим названием – ель финская (Picea fennica (Regel) (Бобров, 1978). Родительские виды в этой зоне, по данным Л.Ф. Правдина (1975), отсутствуют, так как они были вытеснены гибридной формой.
Форма семенных чешуй у многих видов семейства Pinaceae несет наибольший объем генетической информации, свойственной как самому виду, так и его внутривидовым формам, являясь надежным феном при установлении таксономического положения (Абатурова, 1978). Целью настоящей работы является изучение изменчивости ели (Picea obovata Ledeb. × P. abies (L.) Karst.) по форме семенных чешуй и параметрам шишек в бассейне рек Северной Двины, Онеги и Пинеги, что позволит более объективно определять их таксономическую принадлежность и выявлять более продуктивные формы для данных лесорастительных условий.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены на севере Архангельской области в устье р. Северной Двины (64°–65° с.ш., 40°–41° в.д., усть-двинская популяция), бассейне р. Онеги (63°–64° с.ш., 38°–40° в.д., онежская и плесецкая популяции) и р. Пинеги (64°–65° с.ш., 43°–44° в.д., пинежская популяция), в наиболее распространенных типах леса − ельниках черничных свежих, типичных для лесорастительных условий северной тайги. На заложенных в соответствии с принятыми стандартами пробных площадях определяли частоту встречаемости деревьев с разным типом семенных чешуй (Правдин, 1975). В усть-двинской популяции у 25 деревьев каждой формы ели определяли высоту и диаметр ствола на высоте 1.3 м, размеры кроны. На 20 ветвях первого порядка определяли размеры побегов, число боковых почек и побегов в “мутовке”, длину и предельный возраст хвои. Для оценки индивидуальной изменчивости женских генеративных органов у 50 деревьев отбирали индивидуальные образцы по 10 зрелых (урожая прошлого года) шишек на каждом дереве. Для каждого образца определяли длину и массу шишки, форму семенных чешуй (Правдин, 1975), коэффициент вытянутости семенных чешуй (Ср). Показатель Ср определяли в процентах по измерениям проекции семенной чешуи из средней части шишки как отношение расстояния от верхнего края до максимальной ширины к последней (Попов, 1999). Уровни изменчивости признаков определяли согласно шкале С.А. Мамаева (1972).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты исследований показали, что в северных районах Архангельской области по численности доминирует промежуточная форма с преобладанием признаков ели сибирской (западные районы) и форма типичной ели сибирской (восточная часть). В устье Северной Двины соотношение этих форм примерно одинаково (рис. 1). Это соответствует результатам ранее проведенных исследований (Попов, 2010а, б; Коропачинский и др., 2012). Согласно данным П.П. Попова (2011), на северной окраине ареала ели в Восточной Европе средняя длина шишек в популяциях обычно не превышает 60–65 мм. По сведениям А.А. Молчанова и И.Ф. Преображенского (1957), средняя длина шишек в ельниках черничных в Архангельской области составляет 73–75 мм, а по данным Н.П. Пастуховой (цит. по: Попов, 2011) – 67–78 мм. По нашим данным (рис. 2), средняя длина шишек в изучаемых районах увеличивается от 62–63 мм в более северных популяциях до 69 мм в более западном районе и до 72 мм в более южной популяции. Таким образом, предоставленные в разные годы данные довольно сходны, несмотря на вполне вероятные различия в методике сбора образцов шишек. При этом велика вероятность того, что их длина не зависит от формы семенных чешуй и видовой принадлежности ели. Следовательно, можно согласиться с авторами (Попов, 2011) в том, что размеры шишек обусловлены влиянием многих факторов (наследственных, географических, почвенно-климатических, метеорологических, физиолого-биохимических и других). Средняя масса шишки в сухом состоянии в исследуемых популяциях колеблется от 5.0–6.1 г в северных районах до 7.5 г в более западном районе и до 7.8 г в более южном. Следовательно, ее изменение в разных районах соответствует динамике длины шишек.
Рис. 1.
Частота встречаемости ели с разным типом семенных чешуй (по Л.Ф. Правдину, 1975) в районах исследований. 1 – типичная ель сибирская, 2 – ель гибридная с преобладанием признаков ели сибирской, 3 – ель гибридная с преобладанием признаков ели европейской.
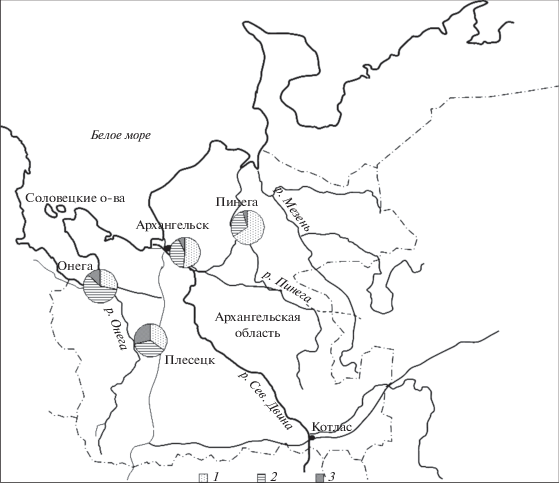
Рис. 2.
Морфоструктурные признаки (среднее значение с ошибкой) зрелых шишек в популяциях ели. 1 – усть-двинская, 2 – пинежская, 3 – плесецкая, 4 – онежская.
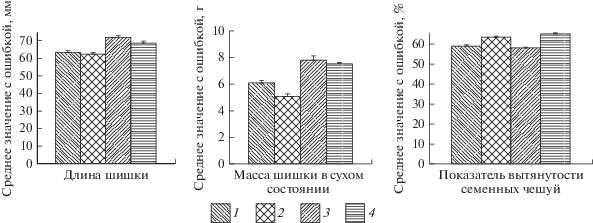
При сравнении морфоструктурных признаков шишек в различных географических районах получены следующие результаты. Усть-двинская и пинежская популяции существенно уступают плесецкой и онежской ели по длине и сухой массе шишек (t-критерий, р < 0.05). Коэффициент вытянутости семенных чешуй в пинежской и онежской популяциях достоверно (на 5%-ом уровне значимости) превышает его величину в усть-двинской и плесецкой популяциях (рис. 2). Однофакторный дисперсионный анализ подтверждает географическую изменчивость длины и массы шишек (F0.05 = 2.65; F = 22.5–26.7), показателя вытянутости семенных чешуй (F0.05 = 2.65; F = 37.2).
Уровни индивидуальной изменчивости длины шишки в популяциях ели низкие (C.V. < 12%), а сухой массы – довольно высокие (C.V. > 20%) (табл. 1). Уровни эндогенной вариации параметров шишек довольно близки к индивидуальным и связаны с нормой реакции генотипа на изменение внешних условий.
Таблица 1.
Изменчивость морфоструктурных признаков шишек в разных популяциях ели
| Показатель | L | M | Cp |
|---|---|---|---|
| Усть-Двинская* (64°–65° с.ш., 40°–41° в.д.) | |||
| Min | 49.6 | 49.6 | 46.7 |
| Max | 78.7 | 78.7 | 72.0 |
| С.V., % | 8.6 | 8.6 | 8.1 |
| Пинежская* (64°–65° с.ш., 43°–44° в.д.) | |||
| Min | 38.4 | 1.6 | 55.3 |
| Max | 75.6 | 8.9 | 71.2 |
| С.V., % | 11.6 | 29.5 | 5.6 |
| Плесецкая* (63° с.ш., 40° в.д.) | |||
| Min | 56.7 | 2.9 | 49.4 |
| Max | 84.5 | 13.3 | 67.9 |
| С.V., % | 10.3 | 27.9 | 6.8 |
| Онежская* (64° с.ш., 38° в.д.) | |||
| Min | 52.3 | 4.2 | 59.9 |
| Max | 81.6 | 10.5 | 71.7 |
| С.V., % | 9.3 | 20.7 | 3.9 |
В табл. 1 и 3: * n = 50. ** n = 39. Примечание. L – длина шишки, мм; М – масса шишки в сухом состоянии, г; Cp – показатель вытянутости семенных чешуй, %; С.V. – коэффициент вариации; n – число деревьев в выборке.
В усть-двинской популяции у форм с разным типом семенных чешуй (“сибирского” и “промежуточного” типов) не наблюдается достоверных различий в размерах ствола и кроны, численности боковых побегов и почек, предельной продолжительности жизни хвои, длине и массе зрелых шишек (t-критерий, p < 0.05) (рис. 3). Это подтверждается результатами однофакторного дисперсионного анализа в отношении параметров охвоенных побегов (F < F0.05). Размерам и численности боковых побегов, длине хвои у деревьев разных форм присуща существенная изменчивость по годам, обусловленная как возрастным фактором, так и влиянием внешних условий (климатических и других факторов) (табл. 2). Об этом свидетельствуют результаты однофакторного (F = 2.17–117.1; F0.05 = 1.90–1.92) и двухфакторного (F = 5.76–109.0; F0.05 = 2.03) дисперсионного анализа Установлены существенные различия форм ели “сибирского” и “промежуточного” типов по коэффициенту вытянутости семенных чешуй (t-критерий, р < 0.05) (рис. 3). Выявлено достоверное влияние фактора “тип семенных чешуй” на этот показатель методом однофакторного дисперсионного анализа (F = 8.62; F0.05 = 4.04). Форма “промежуточного” типа имеет более вытянутые семенные чешуи по сравнению с формой “сибирского” типа, что подтверждает объективность визуального выделения групп деревьев с разным типом шишек по классификации Л.Ф. Правдина (1975). Коэффициенты вариации длины и массы шишки, показателя вытянутости семенных чешуй внутри разных выборок деревьев близки (табл. 3).
Рис. 3.
Морфоструктурные признаки (среднее значение с ошибкой) деревьев ели с разным типом семенных чешуй (средний возраст 103–105 лет). 1 – “cибирский”, 2 – “гибридный”.
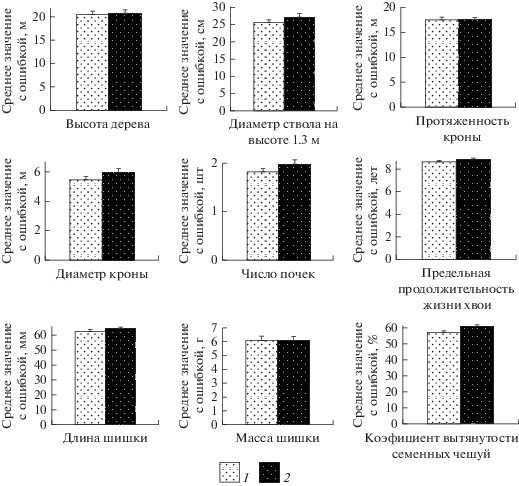
Таблица 2.
Возрастная изменчивость морфоструктурных признаков боковых побегов у форм ели с разным типом семенных чешуй (усть-двинская популяция)
| Год | “Сибирский” тип* | “Промежуточный” тип** | ||||||
|---|---|---|---|---|---|---|---|---|
| L, см | D, мм | N | l, мм | L, см | D, мм | N | l, мм | |
| 2011 | $\frac{{4.0}}{{\begin{array}{*{20}{c}} {0.5} \end{array}}}$ | $\frac{{1.2}}{{0.0}}$ | $\frac{{1.2}}{{0.2}}$ | $\frac{{14.6}}{{2.9}}$ | $\frac{{4.1}}{{0.6}}$ | $\frac{{1.4}}{{0.0}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{15.2}}{{3.0}}$ |
| 2010 | $\frac{{4.1}}{{0.4}}$ | $\frac{{1.4}}{{0.1}}$ | $\frac{{1.4}}{{0.1}}$ | $\frac{{14.7}}{{2.3}}$ | $\frac{{4.0}}{{0.4}}$ | $\frac{{1.6}}{{0.1}}$ | $\frac{{1.5}}{{0.1}}$ | $\frac{{15.2}}{{4.0}}$ |
| 2009 | $\frac{{4.3}}{{0.8}}$ | $\frac{{1.7}}{{0.1}}$ | $\frac{{1.6}}{{0.1}}$ | $\frac{{15.2}}{{2.2}}$ | $\frac{{4.2}}{{0.5}}$ | $\frac{{1.8}}{{0.1}}$ | $\frac{{1.6}}{{0.1}}$ | $\frac{{15.7}}{{4.5}}$ |
| 2008 | $\frac{{5.0}}{{1.0}}$ | $\frac{{1.9}}{{0.2}}$ | $\frac{{1.5}}{{0.2}}$ | $\frac{{15.9}}{{2.5}}$ | $\frac{{4.8}}{{0.8}}$ | $\frac{{2.1}}{{0.1}}$ | $\frac{{1.6}}{{0.2}}$ | $\frac{{16.4}}{{3.5}}$ |
| 2007 | $\frac{{4.2}}{{0.5}}$ | $\frac{{2.2}}{{0.2}}$ | $\frac{{1.4}}{{0.1}}$ | $\frac{{16.3}}{{2.9}}$ | $\frac{{4.2}}{{0.5}}$ | $\frac{{2.3}}{{0.2}}$ | $\frac{{1.5}}{{0.1}}$ | $\frac{{16.7}}{{3.4}}$ |
| 2006 | $\frac{{4.2}}{{0.5}}$ | $\frac{{2.5}}{{0.3}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{16.3}}{{2.7}}$ | $\frac{{4.2}}{{0.6}}$ | $\frac{{2.6}}{{0.2}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{16.7}}{{2.9}}$ |
| 2005 | $\frac{{4.1}}{{0.4}}$ | $\frac{{2.7}}{{0.3}}$ | $\frac{{1.4}}{{0.1}}$ | $\frac{{15.7}}{{3.0}}$ | $\frac{{4.0}}{{0.4}}$ | $\frac{{2.8}}{{0.2}}$ | $\frac{{1.4}}{{0.1}}$ | $\frac{{16.1}}{{2.6}}$ |
| 2004 | $\frac{{4.0}}{{0.4}}$ | $\frac{{2.8}}{{0.3}}$ | $\frac{{1.3}}{{0.1}}$ | $\frac{{15.3}}{{3.1}}$ | $\frac{{3.9}}{{0.4}}$ | $\frac{{3.0}}{{0.2}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{16.1}}{{3.6}}$ |
| 2003 | $\frac{{3.8}}{{0.4}}$ | $\frac{{3.0}}{{0.3}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{15.0}}{{3.8}}$ | $\frac{{3.7}}{{0.3}}$ | $\frac{{3.1}}{{0.2}}$ | $\frac{{1.4}}{{0.2}}$ | $\frac{{14.7}}{{2.5}}$ |
| 2002 | $\frac{{3.8}}{{0.4}}$ | $\frac{{3.2}}{{0.4}}$ | $\frac{{1.3}}{{0.2}}$ | $\frac{{14.5}}{{4.0}}$ | $\frac{{3.5}}{{0.4}}$ | $\frac{{3.2}}{{0.2}}$ | $\frac{{1.3}}{{0.1}}$ | $\frac{{14.5}}{{4.4}}$ |
В табл. 2 и 3: * с признаками типичной ели сибирской; ** промежуточные формы (с преобладанием признаков ели сибирской или ели европейской). Примечание. В числителе – среднее значение признака, в знаменателе – дисперсия; L – длина бокового побега, D – диаметр бокового побега, N – число боковых побегов в “мутовке”, l – длина хвои на боковом побеге.
Таблица 3.
Изменчивость морфоструктурных признаков шишек у форм ели с разным типом семенных чешуй (усть-двинская популяция)
| Показатель | L | М | Cp |
|---|---|---|---|
| “Сибирский” тип | |||
| Min | 53.6 | 3.9 | 46.7 |
| Max | 78.7 | 11.5 | 67.8 |
| С.V., % | 8.5 | 26.8 | 7.3 |
| “Промежуточный” тип | |||
| Min | 49.6 | 4.3 | 54.2 |
| Max | 77.2 | 9.9 | 72.0 |
| С.V., % | 8.5 | 23.7 | 7.7 |
Примечание. Обозначения см. табл. 1.
Анализ сопряженной изменчивости морфоструктурных признаков шишек дает возможность рассматривать женскую генеративную сферу в популяциях ели как единое целое. Корреляционный анализ не выявил линейных связей (r < 0.3) между коэффициентом вытянутости семенных чешуй и параметрами зрелых шишек (длиной и массой) внутри популяции, однако последние тесно коррелируют между собой (r = 0.73 ± 0.07 – 0.85 ± 0.04). Это согласуется с полученными ранее результатами об отсутствии корреляций и показателей длины зрелых шишек и формы семенных чешуй в пределах популяции (Мамаев, Попов, 1989; Попов, 2012).
Заключение. В ельниках черничных свежих в бассейне рек Северной Двины, Онеги и Пинеги на севере Архангельской области высокая частота встречаемости присуща деревьям с типом шишек ели сибирской и близкой к ней промежуточной формы (70–95%). В усть-двинской популяции формы ели с разным типом семенных чешуй в спелом возрасте существенно не различаются по размерам ствола, кроны, численности боковых побегов и почек, продолжительности жизни хвои, длине и массе зрелых шишек. Вместе с тем, параметрам охвоенных побегов присущи значительные колебания по годам, обусловленные их возрастной изменчивостью в процессе онтогенеза деревьев, а также влиянием факторов внешней среды. Выявлены достоверные различия форм ели с разным типом семенных чешуй по показателю вытянутости. Формы “промежуточного” типа имеют более вытянутые семенные чешуи по сравнению с формой “сибирского” типа, что подтверждает объективность их визуального выделения по классификации Л.Ф. Правдина (1975). Более северные и восточные (усть-двинская и пинежская) популяции значительно уступают более южным и западным (плесецкой и онежской) по длине и сухой массе зрелых шишек. Внутри популяций ели морфометрические параметры зрелых шишек не коррелируют с коэффициентом вытянутости семенных чешуй.
Список литературы
Абатурова М.П. Исследование элементарных морфологических признаков ели обыкновенной // Научные основы селекции хвойных древесных пород. М.: Наука, 1978. С. 87–98.
Бобров Е.Г. История и систематика рода Picea A. Dietr. // Новости систематики высших растений. 1971. Т. 7. С. 5–40.
Бобров Е.Г. Интрогрессивная гибридизация, формообразование и смены растительного покрова // Ботанический журн. 1972. Т. 57. № 8. С. 865–879.
Бобров Е.Г. Лесообразующие хвойные СССР. Л.: Наука, 1978. 189 с.
Грант В. Видообразование у растений. М.: Мир, 1984. 528 с.
Коропачинский И.Ю., Потемкин О.Н., Рудиковский А.В., Кузнецова Е.В. Полиморфизм и структура популяций ели сибирской (Picea obovata Ledeb.) на северном пределе распространения вида // Сибирский экологический журнал. 2012. Т. 19. № 2. С. 175–184.
Мамаев С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). М.: Наука, 1972. 284 с.
Мамаев С.А., Попов П.П. Ель сибирская на Урале (внутривидовая изменчивость и структура популяций). М.: Наука, 1989. 104 с.
Молчанов А.А., Преображенский И.Ф. Леса и лесное хозяйство Архангельской области. М.: Изд-во АН СССР, 1957. 238 с.
Попов П.П. Географическая изменчивость формы семенных чешуй ели в Восточной Европе и Западной Сибири // Лесоведение. 1999. № 1. С. 68–73.
Попов П.П. Фенотипическое разнообразие семенных чешуй северодвинских популяций ели и их таксономическая интерпретация // Ботанический журн. 2010. Т. 95. № 5. С. 647–656.
Попов П.П. Формовая структура и географическая дифференциация популяций ели на северо-западе России // Экология. 2010. № 5. С. 336–343.
Попов П.П. Популяционно-географическая изменчивость шишек ели европейской и сибирской // Лесоведение. 2011. № 5. С. 54–60.
Попов П.П. Соотношение показателей длины шишек и формы семенных чешуй в популяциях Piceae abies и P. obovata (Pinaceae) // Ботанический журнал. 2012. Т. 97. № 6. С. 729–734.
Правдин Л.Ф. Ель европейская и ель сибирская в СССР. М.: Наука. 1975. 176 с.
Anderson E. Introqressive hybridization. New York: John Wiley and Sons, 1949. 109 p.
Дополнительные материалы отсутствуют.