

Лесоведение, 2019, № 3, стр. 228-240
Субстрат-специфичность эпиксильных видов в старовозрастных ельниках заповедника “Кивач”
Е. В. Кушневская a, *, Е. А. Боровичев b, Е. В. Шорохова c, d
a Санкт-Петербургский государственный университет
199034 Санкт Петербург, Университетская наб., 7–9, Россия
b Институт проблем промышленной экологии Севера Кольского научного центра РАН
184209 Апатиты, Академгородок, ул. Ферсмана, 14А, Россия
c Институт леса Карельского научного центра РАН
185910 Петрозаводск, Пушкинская ул., 11, Россия
d Санкт-Петербургский государственный лесотехнический университет
194021 Санкт Петербург, Институтский пер., 5, Россия
* E-mail: e.kushnevskaya@spbu.ru
Поступила в редакцию 07.11.2017
После доработки 15.12.2017
Принята к публикации 04.02.2019
Аннотация
Проанализирована субстратная специализация видов эпиксильных сообществ в среднетаежных старовозрастных ельниках черничных заповедника “Кивач” в 2015–2016 гг. Субстратную специализацию видов и их систематических групп на основе встречаемости анализировали с использованием нормализованного индекса специализации d'. В качестве факторов, влияющих на встречаемость видов в эпиксильных сообществах на валеже, рассматривали древесную породу и возраст валежа (давность отпада дерева), степень разложения древесины, диаметр и положение ствола относительно земной поверхности, количество опада на стволе, площадь, покрытая корой, влажность и pH коры. Представители различных групп, составляющих эпиксильную растительность (лишайники, печеночники, мхи, сосудистые растения), были чувствительны к разным факторам. Наиболее важным фактором, определяющим состав эпиксильных сообществ для всех групп, помимо сосудистых растений, оказалась древесная порода валежа. Следующими по значимости были такие факторы, как количество опада, степень разложения древесины и pH коры. Субстратная специализация лишайников и растений различалась. Микроместообитания с наиболее высоким числом видов характеризовались наименьшей специализацией. Наибольшее число видов растений отмечено на валеже ели, наименьшее – на валеже сосны, на которой отмечено наивысшее видовое богатство лишайников. Отдельно проанализирована субстратная приуроченность видов - индикаторов старовозрастных лесов. Бóльшая часть редких и охраняемых видов была встречена на валеже хвойных пород и осины.
В таежных лесах многие редкие и нуждающиеся в охране виды встречаются на крупных древесных остатках (КДО) разных древесных пород и степени разложения, недостаточно представленных в лесах с интенсивной лесохозяйственной деятельностью (Stokland et al., 2012). Часть этих видов является эпиксильными, т.е. обитает на поверхности КДО. Без глубокого понимания их экологии, в особенности требований к качеству субстрата, невозможно сохранение биологического разнообразия, которое, в свою очередь, является неотъемлемой частью устойчивого лесопользования (Карпачевский и др., 2009).
Данных, позволяющих охарактеризовать экологические ниши эпиксильных видов в таежных лесах и разработать стратегии охраны эпиксильного разнообразия, недостаточно (Seibold et al., 2015). Для прогнозирования и моделирования биологического разнообразия особый интерес представляют сведения о сукцессионном статусе видов в масштабе микроместообитания. Сукцессионный статус можно оценивать, как в “абсолютном”, так и в “относительном” времени, т.е. по их приуроченности к стволам определенных давности отмирания (Brūmelis et al., 2017) или стадии разложения (Söderström, 1988). Для таежных лесов России получены фрагментарные оценки “относительного” сукцессионного статуса отдельных эпиксильных видов (Kushnevskaya et al., 2007; Кушневская, 2012). Необходимы количественные оценки их субстратной приуроченности, т.е. оценки их специализации по отношению к древесной породе, размеру и положению ствола, а также “абсолютному” и “относительному” времени эпиксильной сукцессии.
В качестве критериев выделения центров биологического разнообразия – ключевых биотопов и лесов высокой природоохранной ценности используют индикаторные виды (Конечная и др., 2009). Индикаторы, или “суррогаты” биологического разнообразия характеризуют разнообразие группы организмов или всей биоты (Rodrigues, Brooks, 2007; Šálek et al., 2015). Их выделение основано на региональных экспертных оценках встречаемости и уязвимости. Однако для обоснования индикаторной роли видов необходимы количественные оценки их субстратной приуроченности (Halme et al., 2017).
Цель данного исследования – оценить приуроченность эпиксильных видов к субстратам с различными параметрами и выделить факторы, специфичные для редких и нуждающихся в охране видов в условиях средней тайги.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в 2015–2016 гг. в заповеднике “Кивач” (62°28' N, 33°95' E). Территория заповедника находится в юго-восточной части Балтийского кристаллического щита на северо-западном побережье Онежского озера. Среднегодовая температура в районе заповедника составляет +2.4°C, продолжительность вегетационного периода – 90 дней, среднегодовое количество осадков – 625 мм (Скороходова, 2008).
Эпиксильные сообщества изучали на постоянных пробных площадях, первая из них была заложена в старовозрастном смешанном ельнике – Piceetum oxalidosum и Piceetum oxalidoso-myrtillosum с фрагментами Piceetum fontinale и Piceetum oxalidoso-myrtillosum (Федорчук и др., 2005). Почва определена, как элювиально-поверхностно-глееватая глинистая на ленточных глинах и элювиально-поверхностно-глееватая глинистая на ленточных глинах с фрагментами торфянисто-перегнойно-глеевой почвы (Федорец и др., 2006). В древостое преобладала ель (60%, 90–160 лет). 20% запаса приходилось на березу (40–90 лет), 20% – на осину (80 лет). Присутствовала также сосна (+, 90–120 лет). Общий запас древостоя составил 347 м3 га–1. Запас КДО составил 52 м3 га–1. Вторая пробная площадь была заложена в ельнике со значительной примесью сосны, асс. Piceetum oxalidoso-myrtillosum (Федорчук и др., 2005) на поверхностно-подзолистой супесчаной почве. Общий запас древостоя в 369 м3 га–1 был распределен между породами следующим образом: 40% ели (120–160 лет), 30% сосны (120–160 лет), 20% березы (50–60 лет) и 10% осины (70 лет). Запас КДО составил 43 м3 га–1.
В ходе комплексного эксперимента по изучению влияния физических и химических параметров коры и древесины на разнообразие эпиксильной и ксилотрофной биоты было выбрано 83 валежных ствола, диаметром от 16 до 80 см на расстоянии 1.3 м от комля, основных лесообразующих пород: ели (Picea abies H. Karst.), березы пушистой (Betula pubescens Ehrh.) и повислой (B. pendula Roth), осины (Populus tremula L.) и сосны (P-inus sylvestris L.). Стволы датировали (определяли время, пошедшее с момента отмирания дерева) с использованием дендрохронологических методов кросс-датировки, увеличения радиального прироста и механического повреждения соседних деревьев (Dynesius, Jonsson, 1991) за период до 66 лет.
В рамках этого эксперимента были проведены исследования по выявлению основных факторов, определяющих состав и динамику эпиксильной растительности, а также оценена субстратная приуроченность отдельных видов. Использованы флористический и геоботанический подходы к ее изучению. На стволах, отобранных для комплексного эксперимента, заложены круговые площадки 10 см в диаметре на верхней стороне лежащего ствола от комля до вершины через каждые 2 м. Всего было сделано 416 описаний. На этих же стволах обследовали все доступные микроместообитания флористическим методом. Также при отборе образцов коры для физико-химических исследований получены данные о встречаемости эпиксильных видов. Образцы коры отбирали, не отделяя флоэму, но тщательно очищая поверхность от эпиксильной растительности. Охарактеризованы следующие параметры субстрата, которые могут влиять на формирование криптогамной эпиксильной растительности: древесная порода, диаметр ствола, давность отпада, степень разложения субстрата (глубина проникновения ножа, см), процент покрытия валежа корой, проективное покрытие опада на валеже (%), положение относительно земной поверхности (высота, см), рН коры (определяли для экстракта коры в дистиллированной воде из расчета 1 : 25 рН-метром HANNA (Германия)), влажность коры (определяли весовым методом (Shorohova et al., 2016)).
Субстратную приуроченность видов и систематических групп на основе встречаемости анализировали с использованием нормализованного индекса специализации d ', предложенного в качестве одного из показателей для характеристики экологических сетей, преимущественно мутуалистических взаимодействий между организмами различных групп, например, растений и их опылителей (Olesen et al., 2002; Ollerton, Cranmer, 2002). В ее основу положена стандартизированная форма расстояния Кульбака–Лейблера, взятая из теории информации:
где(2)
${{d}_{i}} = \sum\limits_{j - 1}^c {\left( {p_{{ij}}^{'}\ln \frac{{p_{{ij}}^{'}}}{{{{q}_{j}}}}} \right)} ,$Расчет d ' проводили отдельно для лишайников, мхов, печеночников и сосудистых растений, а также для эпиксильного сообщества (синузии) в целом. В настоящей работе мы понимаем синузию в соответствии с трактовкой В.С. Ипатова и Л.А. Кириковой (1999) как ценоэлемент, объединяющий виды близких жизненных форм, находящихся в конкурентных взаимоотношениях, относительно автономно занимающие одно микроместообитание, а, следовательно, связанные совместным произрастанием в определенных условиях среды. В этом же смысле используются термины “эпиксильные группировки” и “эпиксильные сообщества”. Вычисление среднего арифметического d' фактора для видов каждой группы позволяет охарактеризовать роль этих факторов для изучаемых видов, а также сравнить среднюю специализацию для систематических групп. При расчетах использовали следующие градации факторов: древесная порода (ель, береза, осина, сосна); диаметр (≤40 см, >41 см); давность отпада дерева (возраст валежа): лет (0–5, 6–25, 26–40, >41); стадия разложения древесины, выраженная через глубину проникновения ножа, (<1, 1–3, 3.1–10, >10 см); покрытие коры на стволе (<10, 10–90, >91%); влажность коры (0–100, 101–200, 201–300%); опад (ПП) (0–19, 20–39, >40% покрытия); рН коры (4–4.7, 4.8–5.3, 5.4–6.3); положение относительно земной поверхности (0–3, 3–20, >20 см).
Результаты расчета d ' для каждого фактора ранжированы, равным значениям приданы средние ранги. Для каждого вида растений или лишайников посчитан суммарный ранг по результатам расчета всех d' по всем факторам, который также ранжирован. Результат представлен в таблице.
Также для оценки специализации эпиксильной синузии в целом к древесной породе валежа была рассчитана двумерная энтропия Шеннона (Blüthgen et al., 2006):
(3)
$H_{2}^{'} = - \mathop \sum \limits_{i = 1}^r \mathop \sum \limits_{j = 1}^c ({{p}_{{ij}}}{\text{ln}}{{p}_{{ij}}})$Названия видов сосудистых растений даны по сводке С.К. Черепанова (1995) с учетом новейших таксономических изменений. Названия лишайников приведены согласно работе А. Nordin et al. (2011). Названия видов мхов даны в соответствии со сводкой М.С. Игнатова с соавторами (Ignatov et al., 2006); названия печеночников, в основном, приведены по мировому списку печеночников (Söderström et al., 2016), с некоторыми поправками (Konstantinova et al., 2009).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Специфичность субстратов, специализация видов и их систематических групп
В результате расчетов получены данные о специализации каждого вида к различным факторам или характеристикам валежного субстрата. В таблице приведены значения d' и результирующий ранг для каждого вида. Средние d' для групп видов позволили оценить значение различных микроместообитаний на уровне систематических групп. Общие закономерности специализации различных групп отображены на рис. 1–4.
Рис. 1.
Видовое богатство и специализация видов лишайников. а – индекс специализации d' по каждой градации фактора; б – индекс специализации d' средний для всех видов группы; в – общее число видов, отмеченных в каждой градации фактора; г – связь числа видов в каждой градации фактора и уровня специализации (d') этой градации. Древесные породы: Е – ель, Б – береза, С – сосна, О – осина. рН коры: 1 – 4–4.7, 2 – 4.8–5.3, 3 – 5.4–6.3. Положение относительно поверхности земли, см: 1 – 0–3, 2 – 3–20, 3 – >20. Диаметр, см: 1 – ≤40, 2 – >41. Степень покрытия коры на стволе в %: 1 – <10, 2 – 11–90, 3 – >91. Время – давность отпада дерева, лет: 1 – 0–5, 2 – 6–25, 3 – 26–45 и 4 – > 46. Проективное покрытие опада на стволе, %: 1 – 0–19, 2 – 20–39, 3 – >40. Влажность коры в %: 1 – 0–100, 2 – 101–200, 3 – 201–300. Стадия разложения древесины, выраженная через глубину проникновения ножа, см: 1 – <1, 2 – 1–3, 3 – 3.1–10, 4 – >10.
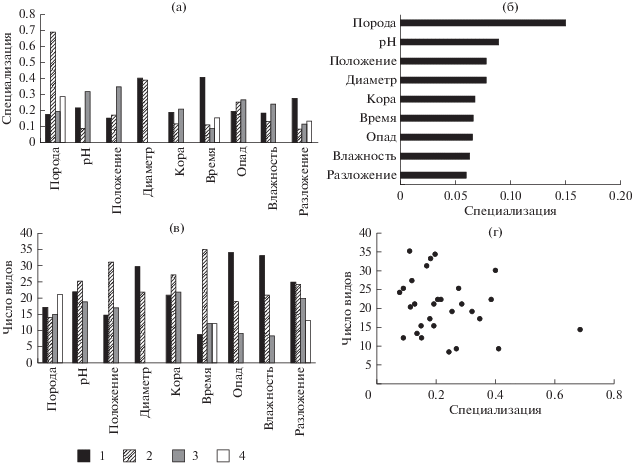
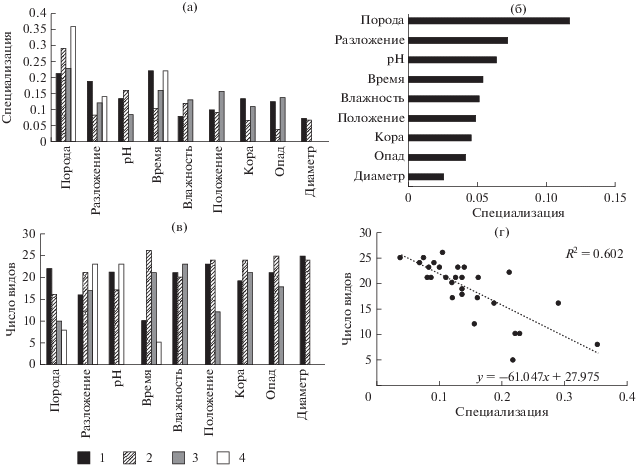
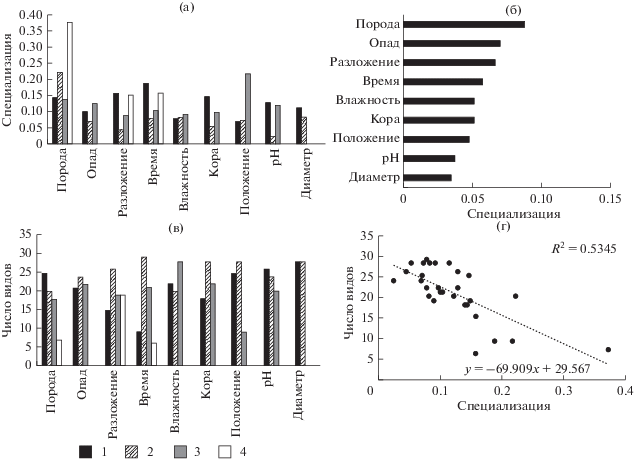
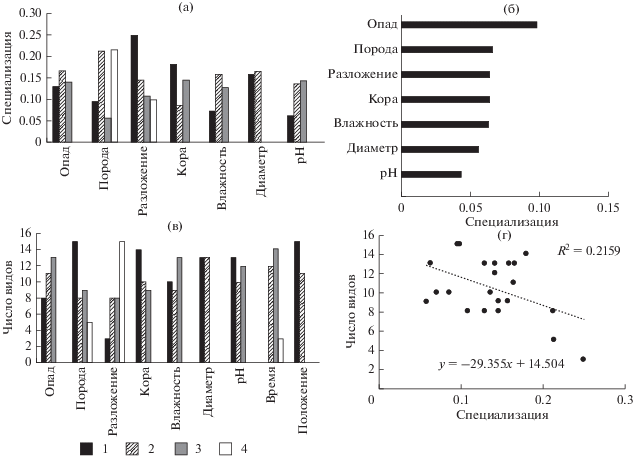
Индекс общей специализации эпиксильной синузии по древесным породам валежа, рассчитанный для всех систематических групп, составил $H_{2}^{'}$ = 0.27 (H2 = 5.11). Это совпадает со значениями, приведенными в работе J. Müller at al. (2015), где общая специализация составила $H_{2}^{'}$ = 0.27 (H2 = 3.74), что указывает на схожий уровень энтропии в эпиксильных сообществах. При расчете общего уровня специализации отдельно для систематических групп были получены следующие результаты: $H_{2}^{'}$ лишайников = 0.33, $H_{2}^{'}$ печеночников = 0.26, $H_{2}^{'}$ мхов = 0.21, $H_{2}^{'}$ сосудистых = 0.14. N. Blüthgen et al. (2006) указывают, что значения $H_{2}^{'}$ уменьшаются по мере увеличения специализации, следовательно распределение сосудистых растений более структурировано, по сравнению, с распределениями криптогамных видов. Возможно это объясняется приуроченностью сосудистых растений к более разложившимся, покрытым опадом стволам.
Ассиметричная оценка специализации градаций фактора дала возможность оценить степень отличий для групп видов и связать этот показатель с числом видов, встреченных на различных вариантах субстрата. Общая закономерность, выявленная нами для трех из четырех систематических групп, выглядит следующим образом: чем выше значения d ' для определенной субстратной категории, тем меньше видов на ней было зарегистрировано (рис. 2г, 3г, 4г). Таким образом, для мхов потенциально наиболее богатым субстратом является еловый валеж средних стадий разложения с фрагментированной корой (рис 3а, 3в), для печеночников – еловый и березовый валеж средних стадий разложения, но со значениями pH, превышающими средние для данных пород (рис. 2а, 2в). Для сосудистых растений вышеописанная тенденция менее выражена, для них наиболее важно высокое количество опада (рис 4а, 4в). Отсутствие подобной закономерности у лишайников, возможно, объясняется тем, что лишайники, в основном, представлены эпифитными или напочвенными видами, для которых эпиксильные сообщества – неоптимальное местообитание.
Лишайники, мхи и печеночники оказались наиболее специализированными к древесной породе валежа. Это отчасти объясняется наличием в эпиксильных сообществах на ранних стадиях зарастания эпифитных видов с высокой специализацией к определенной породе, как, например, лишайника Lobaria pulmonaria, мха Orthotrichum speciosum или печеночника Radula complanata, которые были приурочены исключительно к осине. Однако высокая значимость породы характерна и для эпиксильных печеночников и, что несколько неожиданно, для мхов с очень широкой субстратной амплитудой. Например, Sciuro-hypnum starkei значительно чаще встречался на ели, а Plagiomnium cuspidatum – на осине, хотя оба отмечены на трех древесных породах.
Лишайники наиболее специализированы к осине, на которой встречено небольшое число видов, но большинство из них (10 из 14) не растут на других породах. На сосне встретилось наибольшее число видов лишайников при относительно высоком среднем значении d ' (0.29). Несмотря на то, что видов, встреченных исключительно на сосне, оказалось немного (6 из 21), некоторые из них, отмеченные также на ели или березе, на сосне встречались намного чаще и характеризовались довольно высокими значениями d '. Например, эпифитные Platismatia glauca, Parmeliopsis ambigua на валежной сосне встречались чаще, чем на ели. Отчасти это можно объяснить значительным участием сосны в фитоценозе и хорошей освещенностью на второй пробной площади, на которой обследовали сосновый валеж.
Для числа видов мхов и печеночников и их специализации по породам валежа обнаружены сходные закономерности. На сосне отмечено наименьшее разнообразие мхов и печеночников – распространенных видов с широкой экологической амплитудой. Только Dicranum fuscescens и Crossocalyx hellerianus встретились на сосне чаще, чем на других породах. На ели встречено больше всего видов в обеих вышеупомянутых систематических группах. Соотношение видов, встреченных только на ели, к общему числу видов составило 9 : 25 и 7 : 22 для мхов и печеночников, соответственно, но набор видов сильно отличался. Мхи, отмеченные на ели, характеризовались преимущественно широкой субстратной амплитудой, а среди печеночников оказалось много специализированных эпиксильных видов (Cephalozia macounii, Neoorthocaulis attenuatus, Schistochilopsis incisa, Syzygiella autumnalis, Tritomaria exsecta). Осина – это порода со вторыми по величине d ′ и числом видов за счет эпифитов. Только на осине среди лишайников отмечены Bacidia subincompta, Catinaria atropurpurea, Leptogium saturninum, Lobaria pulmonaria, Mycobilimbia carneoalbida, Peltigera canina, P. praetextata, Pertusaria amara, Phlyctis argena; среди мхов – Brachythecium erythrorrhizon, Eurhynchiastrum pulchellum, Orthotrichum speciosum, Pylaisia polyantha и два вида среди печеночников – Plagiochila porelloides и Radula complanata. На березе не встретилось ни одного уникального вида. Все встретившиеся на березе виды имеют широкую субстратную амплитуду.
Больше всего видов сосудистых растений отмечено на ели, меньше всего – на сосне. Возможно, в чуть более сухих условиях на сосновых стволах, при меньшем количестве опада (см. ниже) сосудистым растениям сложнее заселять валеж.
Сосудистые растения оказались наиболее приуроченными к высокому обилию опада: чем его больше, тем больше видов сосудистых встречено на валеже. С одной стороны, это может быть объяснено тем, что чем дольше лежит ствол, тем больше на нем может накопиться опада. С другой стороны, обилие опада, возможно, благоприятно для разрастания корневых систем. Наиболее специализированным субстратом являлись стволы с покрытием опада 20–40% (вторая градация), где зафиксировано наибольшее число встреч сосудистых растений. На стволах с наименьшим количеством опада разрастались преимущественно O-xalis acetosella, Linnaea borealis и др. Наиболее чувствительными к данному фактору оказались Rubus saxatilis, Maianthemum bifolium. Число видов мхов при разном количестве опада почти не отличалось. Различия показателей d ' между градациями субстрата по количеству опада невелики. Второе по величине среднее значение фактора для видов объясняется чувствительностью эпифитных видов, чаще произрастающих при проективном покрытии (ПП) опада <20% (например, Pylaisia polyantha, Orthotrichum speciosum), или чувствительностью напочвенных видов, чаще встречающихся при ПП опада >40% (например, Dicranum majus, Plagiomnium ellipticum). Для лишайников и печеночников ПП опада было одним из самых малозначимых факторов, но реагировали они на него по-разному. Максимальное число видов лишайников отмечено на стволах с низким ПП опада. При этом такие стволы характеризовались наименьшей специализацией. При большом количестве опада встречены только представители рода Cladonia spp. и Peltigera spp. – эпигейные виды или генералисты. Даже они предпочитали стволы с невысоким ПП опада (в градации 20–40% они встречались чаще). У печеночников максимальное число видов наблюдалось на стволах с 20–40% опада. Ptilidium ciliare и Cephalozia macounii встретились только на площадках с низким ПП опада. Ptilidium ciliare – нехарактерный для эпиксильной растительности вид - был встречен только однажды на каменистых грядах менее чем в 50 м от пробной площади; изредка вид встречается на сухих голых стволах, но на этом субстрате растет плохо и типичных куртин не образует. Cephalozia macounii – очень мелкий печеночник, облигатный эпиксил, растет на влажных, затененных валежных стволах, очень слабоконкурентен. В странах Скандинавии – это очень редкий вид, ограниченный в своем распространении старовозрастными хвойными лесами с давностью хозяйственной деятельности 80–100 лет (Damsholt, 2002). Не отмечено видов печеночников, растущих при ПП опада >40%. При увеличении количества опада снижалось как общее число видов, так и суммарное число встреч. Возможно, тот факт, что большинство видов печеночников приурочены к стволам со средним количеством опада, объясняет их максимальное обилие на средних стадиях эпиксильной сукцессии.
Стадия разложения древесины – важный фактор, и по шкале значимости занимающий 2–3-ю позицию для печеночников, мхов и сосудистых растений; наравне со временем, прошедшим с момента отмирания дерева, влияет на смену видов в процессе сукцессии. Стадию разложения можно рассматривать, как фактор “относительного времени”, влияющий на ход эпиксильной сукцессии. Его влияние на специализацию отдельных видов оказалось сильнее влияния “абсолютного времени”, поскольку влажность и другие параметры стволов, упавших в одно и то же время, могут различаться в зависимости от положения над земной поверхностью и характеристик парцелл, в которых они находятся.
Среднее значение d ' по стадии разложения для видов лишайников было минимальным. Интересно, что валеж первой стадии разложения оказался и наиболее специализированным, и наиболее богатым видами субстратом, что объясняется значительным числом эпифитных видов, в изобилии встречающихся на твердой древесине, но уступающих место мохообразным по мере разложении субстрата.
Динамика d ' по отношению к стадии разложения и давности отпада у мхов имела сходный характер, а у печеночников различалась. У печеночников и мхов первая стадия разложения характеризовалась максимальной специализацией – на самой твердой древесине (глубина проникновения ножа менее 1 см) свежего отпада – оказалось меньше всего видов. Среди видов, характеризующихся высокой специализацией к валежу этой стадии разложения, отмечены типичный эпифитный печеночник Radula complanata, длительное время сохраняющийся на валеже при благоприятных условиях (Brūmelis et al., 2017), и три нетипичных для эпиксильных сообществ вида – Ptilidium ciliare, Rhytidiadelphus subpinnatus и Rhizomnium pseudopunctatum, встретившиеся только по одному разу. Значительное число видов мохообразных отмечено на наименее специализированной категории – древесине второй стадии разложения (глубина проникновения ножа 1–3 см) и времени отпада до 25 лет. Вероятно, эта стадия разложения древесины соответствует той стадии эпиксильной сукцессии, когда открытый субстрат заселяется большим количеством различных видов; разрастаются типичные эпиксильные и эпигейные виды, а также виды генералисты, но еще могут сохраняться некоторые эпифиты, хотя их встречаемость снижается. Большинство видов печеночников отмечены на стволах второй и четвертой стадий разложения. На валеже второй-третьей стадий разница по специализации была незначительной; у мхов число видов немного снижалось. На валеже третьей–четвертой стадий разложения разрастались крупные напочвенные мхи, исчезали эпифиты (Orthotrichum obtusifolium, Pylaisia polyantha) и некоторые генералисты (например, Brachythecium salebrosum). Мхов, приуроченных к поздним стадиям разложения древесины, немного, например, Tetraphis pellucida (собственные неопубликованные данные; Brūmelis et al., 2017). Для печеночников четвертая стадия – самая богатая по числу видов и довольно высокоспециализированная. Причиной этого является то, что большинство видов, появляясь, не исчезают, а продолжают расти отдельными побегами среди куртин крупных мхов. Но есть виды, которые встречаются только на сильно разложившейся древесине, например, Schistochilopsis incisa.
Наибольшее число видов сосудистых растений зафиксировано на самой разложившейся древесине (>10 см). Между валежом 2 и 3-й стадий разложения практически не было разницы ни по числу видов, ни по видовому составу. Заселение субстрата осуществляется в основном вегетативно подвижными видами, наиболее активный из которых – Fragaria vesca. Проростки ели, начиная со второй стадии разложения, отмечены почти в равном количестве.
Оценивая субстрат-специфичность мохообразных, следует заметить, что в нашем исследовании преобладали виды с относительно низкой специализацией, т.е. встречающиеся на большом градиенте субстрата по разложению или накоплению опада. Это отличается от выявленных ранее более высоких значениях специализации отдельных видов (Brūmelis et al., 2017, Műller et al., 2015) в лесах, нарушенных лесохозяйственной деятельностью. Возможно, в условиях старовозрастных лесов с постоянным уровнем атмосферной влажности, постоянно значительным количеством подходящего для заселения субстрата эпиксильные виды демонстрируют более широкую экологическую амплитуду.
Вторым по важности фактором для лишайников и третьим для печеночников являлась pH коры. Мхи и сосудистые растения, напротив, оказались наименее специализированными к рН коры и диаметру ствола. У лишайников наименее специализированным был центральный класс pH коры (pH 4.8–5.3). В этом диапазоне встречались виды лишайников и приуроченных к более кислой коре (pH 4–4.7), и группы предпочитающих слабокислую, близкую к нейтральной (pH 5.3–6.3) среды. Группа видов, более обильная на коре первого класса кислотности, – это виды рода Cladonia spp. и широко распространенные эпифитные виды, например, Parmeliopsis ambigua, Platismatia glauca. При повышенной pH коры встречались виды, характерные для осины (Lobaria pulmonaria, Nephroma parile и др.), поскольку ее кора в норме имеет практически самую высокую щелочную реакцию среди лиственных древесных пород (Werth et al., 2005). Однако приуроченность рассматриваемых нами видов к определенной кислотности коры была выражена слабее, чем к древесной породе, что, возможно, объясняется изменением pH при разложении коры.
Влияние pH коры на поселение и рост печеночников оказалось менее очевидным. Максимальное число видов отмечено на коре с pH 5.3–6.3, однако максимальное число встреч – на стволах с pH коры 4–4.7. Реакция печеночников на изменение pH выражалась, в основном, в числе встреч; подавляющее большинство видов встретилось на стволах, относящихся ко всем трем классам кислотности коры.
Средние значения d ' для видов по остальным факторам оказались довольно низкими, что объясняется широкой экологической амплитудой видов к данным типам микроместообитаний в районе исследования.
Индикаторные и охраняемые виды и их субстрат-специфичность
На обследованных стволах выявлено 14 индикаторных и специализированных к старовозрастным лесам видов (Конечная и др., 2009): три вида лишайников (Cladonia norvegica, Leptogium saturninum и Nephroma parile), три вида мхов (Eurhynchiastrum pulchellum, Hylocomiastrum umbratum и Orthotrichum obtusifolium), семь видов печеночников (Crossocalyx hellerianus, Lophozia ascendens, Syzygiella autumnalis, Neoorthocaulis attenuatus, Riccardia latifrons, R. palmata, Scapania apiculata) и один вид цветковых растений (Galium triflorum).
Cladonia norvegica приурочена к слегка зависшим сосновым стволам средних стадий разложения. Leptogium saturninum, Nephroma parile и Orthotrichum obtusifolium приурочены к валежу осины. При этом Leptogium saturninum отмечен лишь однажды и приурочен только к самым ранним стадиям эпиксильных сукцессий, поскольку быстро отмирает при падении дерева. Nephroma parile и Orthotrichum obtusifolium значительно дольше сохраняются на валеже, они избегают только самых низких значений pH коры и стволов без коры.
Syzygiella autumnalis, Neoorthocaulis attenuatus, Scapania apiculata показали высокую специализацию к породе и чаще встречались на ели, а C. hellerianus – чаще на сосне, чем на ели. Crossocalyx hellerianus и Scapania apiculata, показывая значительную специализацию ко времени, прошедшему с момента отпада дерева, чаще встречались на временном отрезке 6–25 лет, а Neoorthocaulis a-ttenuatus приурочен к более старому валежу возраста 25–40 лет с мягкой древесиной. Riccardia latifrons получила очень низкий ранг и не показала специализации к какому-либо фактору, в данных условиях вид заселяет самые разнообразные доступные субстраты.
Согласно методике выявления и обследования биологически ценных лесов на Северо-западе Европейской части России (Конечная и др., 2009), часть найденных видов маркирует старовозрастные леса, в основном, еловые: Cladonia norvegica, Crossocalyx hellerianus, Galium triflorum, Hylocomiastrum umbratum, Neoorthocaulis attenuatus, Scapania apiculata. Другая часть является индикаторами лесных местообитаний с постоянным высоким уровнем атмосферной влажности: Syzygiella autumnalis Riccardia latifrons, R. palmatа и, наконец, третья группа – индикаторы лиственных лесов с участием широколиственных видов деревьев: Eurhynchiastrum pulchellum, Leptogium saturninum, Nephroma parile, Orthotrichum obtusifolium.
Анализ рангов специализации показал, что большинство индикаторных и специализированных видов, за редким исключением, имели средние ранги специализации. Зачастую наиболее высокие ранги специализации отмечались у видов, нехарактерных для эпиксильных сообществ, – эпифитов или эпигеидов. Наиболее низкие ранги получили, в большинстве случаев виды с широкой экологической амплитудой, для которых весь спектр возможных условий на валеже входит в реализованную экологическую нишу. Основываясь на результатах настоящего исследования, можно рекомендовать в группу индикаторов старовозрастных лесов в условиях подзоны средней тайги еще 3 вида: Cephalozia macounii, Schistochilopsis -incisa, Tritomaria exsecta.
Из встреченных нами видов в Красную книгу Республики Карелии (2007) внесены четыре печеночника: Cephalozia macounii, Lophozia ascendens, Syzygiella autumnalis, Scapania apiculata и один лишайник: Lobaria pulmonaria. Последний также включен в Красную книгу России (2008). Среди охраняемых в регионе, наиболее редким является печеночник Cephalozia macouni. Это второе достоверное указание для Карелии после находки в Муезерском районе (Потемкин, 2005); известное ранее местонахождение из Кемского района (Junninen et al., 1996), возможно, является сомнительным (Бакалин, 1999). Tritomaria exsecta впервые зарегистрирована для флоры печеночников Карелии в рамках выполнения настоящего проекта (Ellis et al., 2016). Находка в заповеднике “Кивач” – единственная изолированная от ближайших местонахождений популяция на северной границе распространения вида. Рекомендуется включить вид в третье издание Красной книги Карелии с категорией 1 – “Critically endangered”.
Заключение. Изучение субстратной приуроченности видов эпиксильных сообществ показало, что важнейшим фактором, влияющим на их состав, является древесная порода валежа. Наиболее богатые группировки формируются на еловом валеже, наиболее специфичные – на осиновом. Валеж березы с нефрагментированной медленно разлагающейся корой является наименее специфичным и наименее привлекательным для специализированных эпиксильных видов. Полученные закономерности следует учитывать, как при оценке, так и при разработке стратегии сохранения биоразнообразия в лесах таежной зоны. Максимально полночленные эпиксильные сообщества могут формироваться только в лесах с валежом хвойных и лиственных пород. При наличии только валежа березы, что характерно для определенных стадий лесовосстановительных сукцессий, будут возникать обедненные группировки, без специализированных и редких эпиксильных видов. Вторым важным фактором формирования полночленных эпиксильных сообществ в фитоценозе является наличие валежа разных стадий разложения, необходимые для поддержания постоянного пула различных микроэкониш. При сочетании этих двух факторов гниющая древесина является важным субстратом для большого числа редких и охраняемых видов лишайников и мохообразных.
***
Мы благодарим сотрудников заповедника “Кивач” за организацию полевых работ; Алексея Полевого, Екатерину Капица, Анастасию Мамай, Ивана Ромашкина и Анну Руоколайнен – за помощь в подборе пробных площадей и датировке объектов, Наталию Галибину и Ксению Никерову – за лабораторные анализы образцов коры. Выражаем слова благодарности рецензентам статьи за вопросы и замечания, позволившие улучшить рукопись.
Список литературы
Бакалин В.А. Печеночники Карелии // Arctoa. 1999. Т. 8. С. 17–26.
Ипатов В.С. Кирикова Л.А. Фитоценология. СПб.: Изд-во Санкт-Петербургского университета. 1999. 316 с.
Карпачевский М.Л., Тепляков В.К., Яницкая Т.О., Ярошенко А.Ю. Основы устойчивого лесоуправления: Учеб. пособие для вузов. М.: WWF, 2009. 143 с.
Конечная Г.Ю., Курбатова Л.Е., Потемкин А.Д., Гимельбрант Д.Е., Кузнецова Е.С., Змитрович И.В., Коткова В.М., Малышева В.Ф., Морозова О.В., Попов Е.С., Яковлев Е.Б., Andersson L., Кияшко П.В., Skujienė G. Выявление и обследование биологически ценных лесов на Северо-Западе Европейской части России. Т. 2. Пособие по определению видов, используемых при обследовании на уровне выделов. СПб.: Победа, 2009. 258 с.
Красная книга Республики Карелия. Петрозаводск: Карелия, 2007. 368 с.
Красная книга Российской Федерации (растения и грибы). М.: Товарищество научных изданий КМК, 2008. 855 с.
Кушневская Е.В. Эпиксильные сукцессии в ельниках Ленинградской области // Ботанический журн. 2012. Т. 97. №. 7. С. 917–939.
Потемкин А.Д. К флоре печеночных мхов Муезерского района Республики Карелия // Новости систематики низших растений. 2005. Т. 39. С. 263–269.
Скороходова С.Б. О климате заповедника “Кивач” // Труды государственного природного заповедника “Кивач”. Вып. 4. Петрозаводск: Изд-во Петрозаводского государственного университета, 2008. С. 3–34.
Федорец Н.Г., Морозова Р.М., Бахмет О.Н., Солодовников А.Н. Почвы и почвенный покров заповедника Кивач” // Труды Карельского НЦ РАН. 2006. Т. 10. С. 131–152.
Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. Лесные экосистемы северо-западных районов России: типология, динамика, хозяйственные особенности. СПб.: СПбНИИ лесного хозяйства, ЗАО “Хромис”, 2005. 382 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
Blüthgen N., Menzel F., Blüthgen N. Measuring specialization in species interaction networks // BMC Ecology. 2006. V. 6. P. 9. doi .https://doi.org/10.1186/1472-6785-6-9
Brūmelis G., Oļehnoviča E., Šūba U., Treimane A., Inne S., Zviedre E., Elferts D., Dakša M., Tjarve D. Bryophyte and polypore richness and indicators in relation to type, age and decay stage of coarse woody debris of Picea abies // Environmental & Experimental Biology. 2017. V. 15. P. 95–103.
Damsholt K. Illustrated flora of Nordic liverworts and hornworts. Lund: Nord. Bryol. Soc., 2002. 840 p.
Dynesius M., Jonsson B.G. Dating uprooted trees: comparison and application of eight methods in a boreal forest // Canadian J. Forest Research. 1991. V. 21. P. 655–665.
Ellis L.T., Alatas M., Asthana A.K., Rawat K.K., Sahu V., Srivastava A., Bakalin V.A., Batan N., Bednarek-Ochyra H., Bester S.P., Borovichev E.A., Beer D.De, Enroth J., Erzberger P., Fedosov V.E., Feuillet-Hurtado C., Gradstein S.R., M. Gremmen N.J., Hedenas L., Katagiri T., Yamaguchi T., Lebouvier M., Maity D., Mesterhazy A., Műller F., Natcheva R., Nemeth Cs., Opisso J., Ozdemir T., Erata H., Parnikoza I., Plasek V., Sabovljevic S., Sabovljevic A.D., Saha P., Aziz Md Nehal, Schroder W., Vana J., Rooy J. van, Wang J., Yoon Y.–J., Kim J.H. New national and regional bryophyte records, 47 // J. Bryology. 2016. V. 38. № 2. P. 151–168.
Halme P., Holec J., Heilmann-Clausen J. The history and future of fungi as biodiversity surrogates in forests // Fungal Ecology. 2017. V. 27. P. B. P. 193–201.
Ignatov M.S., Afonina O.M., Ignatova E.A., Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A G., Boychuk M.A., Cherdantseva V.Ya., Czernyadjeva I.V., Doroshina G.Ya., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Kharzinov Z.Kh., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K., Manakyan V. A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Ya., Pisarenko O.Yu., Popova N.N., Rykovsky G.F., Tubanova D.Ya., Zheleznova G.V., Zolotov V.I. Check-list of mosses of East Europe and North Asia // Arctoa. 2006. V. 15. P. 1–130.
Junninen K., Lindgren M., Nykänen R., Tikkanen O.P. Survey in Russian Karelian Natural Forests in Vienansalo. Helsinki: WWW Finland, 1996. 33 p.
Konstantinova N.A., Bakalin V.A., Andreeva E.N., Bezgodov A.G., Borovichev E.A., Dulin M.V., Mamontov Yu.S. Checklist of liverworts (Marchantiophyta) of Russia // Arctoa. 2009. V. 18. P. 1–63.
Kushnevskaya H., Mirin D., Shorohova E. Patterns of epixylic vegetation on spruce logs in late-successional boreal forests // Forest Ecology and Management2007. V. 250. P. 25–33.
Müller J., Boch S., Blaser S., Fischer M., Prati D. Effects of forest management on bryophyte communities on deadwood // Nova Hedwigia. 2015. V. 100. P. 423–438.
Nordin A., Moberg R., Tønsberg T., Vitikainen O., Dalsätt Å., Myrdal M., Snitting D., Ekman S. Santesson’s Checklist of Fennoscandian Lichen-forming and Lichenicolous Fungi. Ver. April 29, 2011 – http://130.238.83.220/santesson/ home.php (9 May 2016).
Olesen J.M., Jordano P. Geographic patterns in plant-pollinator mutualistic networks // Ecology. 2002. V. 83. P. 2416–2424.
Ollerton J., Cranmer L. Latitudinal trends in plant-pollinator interactions: are tropical plants more specialized? // O-ikos. 2002. V. 98. P. 340–350.
Rodrigues A.S.L., Brooks T.M. Shortcuts for biodiversity conservation planning: the effectiveness of surrogates // Annual Review of Ecology, Evolution, and Systematics. 2007. V. 38. P. 713–737.
Šálek, M., Kučera, T., Zimmermann, K., Bartůšková I., Plátek M., Grill S., Konvička M. Edges within farmland: management implications of taxon specific species richness correlates // Basic and Applied Ecology. 2015. V. 16. P. 714–725.
Seibold S., Bässler C., Brandl R., Gossner M., Thorn S., Ulyshen M., Müller J. Experimental studies of dead-wood biodiversity – A review identifying global gaps in knowledge // Biological Conservation. 2015. V. 191. P. 139–149.
Shorohova E., Kapitsa E., Kazartsev I., Romashkin I., Polevoi A., Kushnevskaya H. Tree species traits are the predominant control on the decomposition rate of tree log bark in a mesic old-growth boreal forest // Forest Ecology and Management. 2016. V. 377. P. 36–45.
Söderström L. Sequence of bryophytes and lichens in relation to substrate variables of decaying coniferous wood in Northern Sweden // Nordic J. Botany.1988. V. 8. № 1. P. 89–97.
Söderström L., Hagborg A., von Konrat M., Bartholomew-Began S., Bell D., Briscoe L., Brown E., Cargill D.C., Costa D.P., Crandall-Stotler B.J., Cooper E.D., Dauphin G., Engel J.J., Feldberg K., Glenny D., Gradstein S.R., He X., Heinrichs J., Hentschel J., Ilkiu-Borges A.L., Katagiri T., Konstantinova N.A., Larraín J., Long D.G., Nebel M., Pócs T., Felisa Puche F., Reiner-Drehwald E., Renner M.A.M., Sass-Gyarmati A., Schäfer-Verwimp A., Moragues J.G.S., Stotler R.E., Sukkharak P., Thiers B.M., Uribe J., Váňa J., Villarreal J.C., Wigginton M., Zhang L., Zhu R.-L. World checklist of hornworts and liverworts // PhytoKeys. 2016. V. 59. P. 1–828.
Stokland J.N., Siitonen J., Jonsson B.G. Biodiversity in Dead Wood. Cambrigde: Cambrigde University Press, 2012. 509 p.
Werth S., Tømmervik H., Elvebakk A., Økland R.H. Epiphytic macrolichen communities along regional gradients in northern Norway // J. Vegetation Science. 2005. V. 16. № 2. P. 199–208.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение