

Лесоведение, 2019, № 3, стр. 163-176
Сукцессионная динамика растительности и запасы почвенного углерода в хвойно-широколиственных лесах Северо-Западного Кавказa
Н. Е. Шевченко a, *, А. И. Кузнецова a, Д. Н. Тебенькова a, В. Э. Смирнов a, b, А. П. Гераськина a, А. В. Горнов a, Е. А. Грабенко c, Е. В. Тихонова a, Н. В. Лукина a
a Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32, стр. 14, Россия
b Институт математических проблем биологии РАН – филиал Федерального исследовательского центра “Института прикладной математики им. М.В. Келдыша РАН”
142290 Пущино, ул. Профессора Виткевича, 1, Россия
c Майкопский государственный технологический университет
385000 Республика Адыгея, г. Майкоп, ул. Первомайская, 191, Россия
* E-mail: neshevchenko@gmail.com
Поступила в редакцию 06.04.2018
После доработки 21.05.2018
Принята к публикации 04.02.2019
Аннотация
В статье дана оценка динамики растительности и изменений запасов почвенного углерода в хвойно-широколиственных лесах Северо-Западного Кавказа в ходе послерубочной сукцессии. Эта проблема актуальна в связи с изменением климата и вкладом лесных экосистем в сток углерода. Объекты исследования – хвойно-широколиственные леса Северо-Западного Кавказа (Кавказский биосферный заповедник и Апшеронское лесничество в верховьях р. Белой и р. Пшехи) на разных стадиях восстановительной сукцессии пихтово-буковых лесов. На основе эколого-ценотической классификации выделены и описаны три стадии: ранняя – осиново-грабовые жимолостно-мелкотравные сообщества, переходная – буково-пихтово-грабовые мелкотравные сообщества, поздняя – пихтово-буковые мертвопокровные сообщества. Дана оценка условий местообитаний стадий восстановительной сукцессии по результатам многомерного анализа геоботанических описаний с использованием экологической шкалы Г. Элленберга. Проведен анализ кислотности, содержания углерода, азота и запасов углерода в почвах с применением современных методов. Выявлено, что на поздней стадии сукцессии, представленной буково-пихтовыми лесами, происходит возрастание запасов почвенного углерода в подстилке и их снижение в гумусовом горизонте. Эти разнонаправленные изменения в верхних горизонтах почв обусловлены доминированием древесных растений, формирующих опад низкого качества. Дополнительным объяснением снижения запасов почвенного углерода является отсутствие трав на поздней стадии сукцессии. Отношение C : N в гумусовом горизонте регулируется не только качеством опада, но и биомассой дождевых червей. Выявлено сходство трендов, установленных на основе количественной оценки почвенных показателей и с применением экологической шкалы Г. Элленберга, в изменении характеристик местообитаний.
Лесной растительный и почвенный покров формируются в тесных взаимосвязях, оценка которых имеет огромную прогностическую ценность. Эта проблема приобрела особую актуальность в настоящее время в связи с изменениями климата и значительным вкладом лесов в сток углерода. Ряд работ демонстрирует накопление углерода на поздних стадиях сукцессий в старовозрастных лесах (Luyssaert et al., 2008; Lewis et al., 2009), в том числе в почвах (Zhou et al., 2006; Framstad et al., 2013). Около 73% почвенного углерода аккумулируется именно в лесах (Vashum et al., 2012), и изменение содержания углерода в почвах связано с сукцессионной динамикой лесов. Виды доминирующих растений, составляющих сообщества разных стадий сукцессии лесов, влияют на характеристики местообитаний, такие как освещенность, влажность, температура, кислотность, богатство почв азотом и другими элементами питания, что оказывает воздействие на способность почв аккумулировать углерод.
На территории России леса представлены мозаикой различных сукцессионных стадий, однако результатов исследований, на основании которых можно сделать определенные заключения о связи между сукцессионным статусом лесов разных типов и закономерностями накопления почвенного углерода в них, явно недостаточно. Горные леса занимают около 40% покрытой лесом площади России (Луганский и др., 2010), и оценка связи между уровнем аккумуляции углерода в почвах этих лесов и их сукцессионным статусов является важной фундаментальной и прикладной задачей.
Цель данной работы – оценить динамику растительности, содержания и запасов углерода в почвах хвойно-широколиственных лесов Северо-Западного Кавказа в ходе послерубочной сукцессии.
Для достижения поставленной цели решались следующие задачи:
дать оценку динамики растительности в ходе послерубочной сукцессии;
оценить условия местообитаний на разных стадиях сукцессий с применением метода экологических шкал;
дать оценку динамики кислотности, содержания азота и углерода в почве в ходе сукцессии;
дать оценку динамики запасов углерода в почвах в ходе сукцессии лесов.
ОБЪЕКТЫ И МЕТОДИКА
Объекты исследований расположены на территории Кавказского биосферного заповедника и Апшеронского лесничества в верховьях рр. Белой и Пшехи. Высота над уровнем моря 650–700 м. Среднегодовая температура составляет 9.3°С (по данным метеостанции в пос. Гузерипль с 2009–2018 гг.), годовое количество осадков – 1200 мм. В геолого-геоморфологическом отношении район исследования входит в область северной сланцевой депрессии (Гузерипльский прогиб), характеризующейся широким распространением глинистых сланцев юрского возраста, алевритов и песчаников (Геология …, 1968).
Для выявления влияния сукцессионного статуса лесов на свойства почв выбран пространственно-временной ряд демутационной сукцессии буково-темнохвойных лесов среднегорного пояса Северо-Западного Кавказа на почвообразующих породах сходного валового и гранулометрического состава (Горнов и др., 2018).
По эколого-флористической классификации леса района исследования относятся к ассоциации Ilici colchicae-Abietetum nordmannianae Korotkov et Belonovskaja, 1987 союза Vaccinio-Fagion orientalis (Zohary, 1973) Passarge, 1981 класса Querco-Fagetea Br.-Bl. et Vlieger, 1937 (Биота экосистем …, 1990). Изученные сообщества пространственно-временного ряда хвойно-широколиственных лесов нами были условно разделены на три стадии восстановительной сукцессии – раннюю, переходную и позднюю. На ранней стадии демутационной сукцессии в древесном пологе доминирует Carpinus betulus с примесью Populus tremula, Quercus petraea, Cerasus avium, травяной ярус представлен мелкотравьем. В почвенном покрове распространены буроземы (World …, 2014). Подстилка маломощная, деструктивного типа. В древостое переходной стадии в верхний подъярус выходят позднесукцессионные виды – Fagus orientalis и Abies nordmanniana. Нижний подъярус формируют раннесукцессионные виды деревьев – Carpinus betulus, Populus tremula, Quercus petraea и Betula pubescens, в напочвенном покрове происходит развитие неморального мелкотравья. В почвенном покрове распространены буроземы, встречаются буроземы глееватые (Cambisols Dystric, по WRB, 2014). На поздней стадии древесный ярус состоит из Fagus orientalis и Abies nordmanniana, напочвенный ярус мертвопокровный. В почвенном покрове распространены буроземы глеевые. Наблюдаемое оглеение типично для почв района исследования и определяется избыточным поверхностным переувлажнением (Зонн, 1950). Признаки оглеения выражены более ярко на третьей стадии сукцессии с преобладанием бука, отличающегося низкой интенсивностью транспирации (Кrämer, 2009; Gebauer, 2010). Почвы всех трех стадий сукцессии развиваются на элювии глинистых сланцев (Голгофская и др., 1967).
В летне-осенний период 2016 г. было заложено 9 пробных площадей (ПП), размером 50 × 50 м, по 3 в лесах каждой стадии сукцессии. На каждой пробной площадке выполнялось сплошное измерение деревьев выше 1.5 м. Оценивалась высота (лазерный высотомер Nikon Forestry Pro), абсолютный возраст (возрастной бурав) и окружность стволов (рулетка). На основе полученных данных определен запас стволовой древесины всех видов деревьев на основе Справочника лесотаксационных нормативов для Северного Кавказа (1995).
Для оценки внутрибиогеоценотической неоднородности почв на каждой пробной площади закладывались опорные разрезы, откуда отбирались образцы подстилки (с разделением ее на подгоризонты L, F, H) и минеральных горизонтов почв (каждые 10 см). Кроме того, каждую пробную площадь размером 0.25 га делили на 25 квадратов (0.01 га) и с помощью почвенного бура отбирали по 3 усредненных образца по горизонтам (L, FH, A, АВ, B) путем смешивания 8 единичных проб из каждого горизонта. Для подготовки усредненных образцов не использовался только квадрат, в котором закладывался опорный разрез. Для пересчета запасов углерода на каждой пробной площади отобраны образцы подстилки с использованием рамки размером 0.25 × 0.25 м в трехкратной повторности. При расчете запасов углерода пользовались методическими указаниями по количественному определению объема поглощения парниковых газов (Распоряжение …, 2017).
Определение плотности почв проводилось с помощью отбора образцов с использованием бюкса. В лабораторных условиях образцы почв и подстилки, отобранные для пересчета запасов, высушивали до воздушно-сухого состояния и взвешивали. Почвенные образцы, отобранные для определения физико-химических свойств почв, в лабораторных условиях высушивали, просеивали через сито 2 мм. Актуальную кислотность (рН) измеряли потенциометрически в водной вытяжке. Обменную кислотность определяли в вытяжках однонормального раствора KCl титрованием до pH = 7.8 (Теория …, 2006). Оценка содержания органического углерода и общего азота проводилась на CHN-анализаторе (ЕА 1110 (CHNS-O)). Для определения содержания доступных для биоты соединений элементов питания образцы почв обрабатывали одномолярным раствором CH3COONH4 (pH = = 4.65) (Halonen et al., 1983; Christensen et al., 1989). Содержание металлов определяли методом атомно-абсорбционной спектрофотометрии (AAnalyst 800 spectrometer).
Сбор и обработка геоботанических материалов, полученных в ходе исследований, проведены по общепринятым методикам (Методические подходы …, 2010). Сукцессионный статус лесов оценивали по индикаторам, которые разработаны отечественными популяционными биологами (Смирнова и др., 1988, 2006; Евстигнеев и др., 1992). К основным индикаторам относятся полночленность видового набора деревьев (как основных ценозообразователей) и онтогенетический состав их популяций (Горнов и др., 2018).
На каждой пробной площади проведены учеты численности дождевых червей по стандартной методике (Гиляров, 1987). Первоначально обследовалась подстилка, из которой выбирали все найденные черви, затем проведена послойная раскопка почвы до глубины встречаемости червей (на Кавказе, как правило, не более 40 см).
Экологические характеристики местообитаний растительных сообществ (N – богатство почвы азотом; R – почвенная кислотность; L – освещенность; F – увлажнение почвы; T – температура) получены как средневзвешенные баллы по соответствующим характеристикам видов согласно экологической шкале Г. Элленберга (Ellenberg, 1996).
Латинские названия сосудистых растений приведены по С.К. Черепанову (1995), мхов – в соответствии со списком (Ignatov et al., 2006). Сообщества типизированы на основе работы “Определитель типов леса Европейской России” (www.cepl.rssi.ru/bio/forest/). Для выявления особенностей сообществ разных стадий демутационной сукцессии исследованных лесов Северо-Западного Кавказа проведено сопоставление их эколого-ценотического состава. Выделено шесть эколого-ценотических групп: Br – бореальные виды; Nm – неморальные; MFr – лугово-опушечные; Nt – нитрофильные; Qx – байрачные; Rock – скальные.
На каждой пробной площади заложены по 3 геоботанические площадки размером 20 × 20 м, всего выполнено 27 описаний. На геоботанических площадках учтены все виды сосудистых растений и мхов, определено общее покрытие по ярусам. Балльная оценка покрытия–обилия выполнена с использованием шкалы Браун-Бланке. Ординацию геоботанических описаний проводили с помощью многомерного неметрического шкалирования в среде статистического программирования R (www.R-project.org). Использовали матрицу расстояний Брея–Кертиса – перед ординацией данные преобразовали следующим образом: баллы покрытия для древесных пород заменили площадями поперечных сечений (м2 га–1), затем обилие всех видов стандартизировали путем деления на максимальные значения. Всего в ординационом анализе участвовало 74 вида сосудистых растений, мхов и лишайников (виды, встреченные менее чем на 3 площадках, из анализа исключили). Для интерпретации ординационной диаграммы строили корреляционные векторы экологических шкал, видового богатства по ярусам, высоты и возраста древесных пород.
Для оценки влияния стадии сукцессии на почвенные характеристики применяли v-критерий (Husson, 2017; Лукина и др., 2018).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Растительность
Анализ сообществ идентифицированных стадий восстановительной сукцессии по экологическим шкалам Г. Элленберга (Ellenberg, 1996) позволил определить наиболее значимые факторы для функционирования растительности на объектах исследований: освещенность, увлажнение, кислотность/щелочность и богатство почвы азотом. На рис. 1 изображены векторы экологических факторов, рассчитанные по шкалам Г. Элленберга, которые наиболее четко отражают экологическую дифференциацию стадий сукцессии. Длина и направление векторов отражают степень корреляции факторов с осями, но не являются регрессионными прямыми.
Рис. 1.
Ординация 27 геоботанических описаний методом неметрического шкалирования. Стадии сукцессии хвойно-широколиственных лесов: I – ранняя, II – переходная, III – поздняя; 1–9 – номера пробных площадей. Векторы Шкалы Г. Элленберга: F – почвенное увлажнение, L – освещенность, N – богатства почвы азотом, R – почвенная кислотность, T – температура; возраст – средний возраст деревьев по пробным площадям; высота – средняя высота деревьев, SR.A, SR.B, SR.C, SR.D – число видов в ярусах A, B, C, D, соответственно.
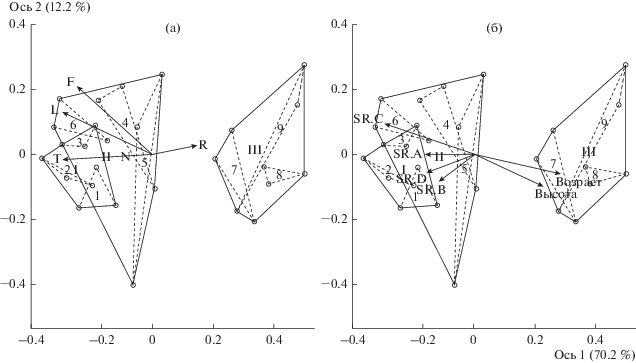
Многомерный анализ геоботанических описаний разных стадий демутационной сукцессии по экологическим шкалам Г. Элленберга (рис. 1) выявил приуроченность раннесукцесионных сообществ (осиново-грабовые жимолостно-мелкотравные) к участкам с повышенной освещенностью, низкой кислотностью, высоким увлажнением и богатством почвы азотом по сравнению с сообществами переходной (буково-пихтово-грабовые мелкотравные) и поздней (пихтово-буковые мертвопокровные) стадиями. Во флоре раннесукцесионных сообществ выше доля светолюбивых, влаголюбивых, нитрофильных и теплолюбивых средиземноморских и субсредиземноморских видов по сравнению с флорой сообществ переходной и поздней стадий. На ранней стадии проективное покрытие травяно-кустарничкового яруса достигает 20–25(35)%.
Важно отметить, что сообщества переходной стадии по всем экологическим факторам занимают промежуточное положение между сравниваемыми сообществами. На данной стадии происходит распад яруса первого послерубочного поколения деревьев Carpinus betulus и выход в верхний древесный ярус позднесукцессионных видов – Abies nordmanniana и Fagus orientalis. Этот процесс сопровождается постепенным вытеснением на поздней стадии демутационной сукцессии светолюбивых раннесукцесионных видов теневыносливыми позднесукцессионными, поэтому флора буково-пихтово-грабовых мелкотравных сообществ отличается наибольшим видовым богатством (71 вид), включая виды растений ранней и поздней стадий. Проективное покрытие травяно-кустарничкового яруса – 35–50(80)%.
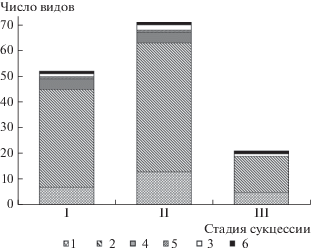
Сравнивая эколого-ценотическую структуру разных стадий изучаемого сукцессионного ряда хвойно-широколиственных лесов, следует отметить преобладание на всех стадиях сукцессии бореальных и неморальных видов (86–89%), формирующих основное ядро лесной флоры этой территории. Анализ эколого-ценотической структуры растительных сообществ на разных стадиях сукцессии позволил установить, что в горных лесах Северо-Западного Кавказа на поздней стадии сукцессии эколого-ценотическая структура упрощается, что приводит к формированию олигодоминантных мертвопокровных сообществ. На ранней и переходной стадиях выше доля неморальных, нитрофильных и лугово-опушечных видов, отмечено присутствие байрачных видов, на поздней – значительно возрастает доля бореальных видов, резко снижается доля лугово-опушечных видов, полностью отсутствуют байрачные и нитрофильные виды.
Пихтово-буковые мертвопокровные леса поздней стадии оцениваются как самые маловидовые (22 вида) олигодоминантные сообщества. По числу видов в составе растительных сообществ на поздней стадии преобладают деревья (8 видов), из которых Abies nordmanniana и Fagus orientalis являются основными средообразователями. Остальные виды деревьев встречаются в подлеске единично. Переход к заключительной стадии происходит после полного выпадения из древесного яруса всех светолюбивых видов. На поздней стадии высота древесного полога увеличивается до 60 м. Данный процесс сопровождается дифференциацией древостоя на два хорошо различимых подъяруса: А2 – высота 20–30 м и А1 – 50–60 м. Верхний подъярус формирует Fagus orientalis, а нижний – Abies nordmanniana, что приводит к сильному затенению и формированию метровопокровных сообществ. Мелкотравные и высокотравные виды встречаются единично в редких окнах древесного яруса. Проективное покрытие травяно-кустарничкового яруса составляет 3–5(8)%.
Важным показателем для оценки восстановительной сукцессии растительных сообществ и динамики запаса углерода в условиях Северо-Западного Кавказа является абсолютный возраст деревьев и общий запас стволовой древесины на разных стадиях развития древостоя (рис. 1). В осиново-грабовых жимолостно-мелкотравных сообществах ранней стадии максимальный возраст деревьев отмечен для Populus tremula (65 лет), Carpinus betulus (54 лет), Quercus petraea (55 лет), Fagus orientalis (45 лет) и Cerasus avium (43 года) при средней высоте древесного яруса 22 м. Абсолютный возраст доминантов древесного яруса сопоставим, поэтому можно утверждать, что эти раннесукцессионные сообщества представляют собой первое поколение деревьев демутационной сукцессии. Средний запас стволовой древесины составляет 293 ± 34 м3 га–1, из которых около 75% составляет запас Carpinus betulus и около 20% – Populus tremula; на долю остальных видов приходится менее 5%.
Полная смена осиново-грабовых жимолостно-мелкотравных сообществ ранней стадии на буково-пихтово-грабовые мелкотравные сообщества переходной стадии происходит ориентировочно в возрасте 80–110 лет, когда средняя высота древостоя достигает 30–35 м. В сообществах этой стадии происходит постепенное выпадение раннесукцессионных видов, поэтому максимальный абсолютный возраст деревьев имеют позднесукцессионные виды Abies nordmanniana – 94 г и Fagus orientalis – 91 г. Максимальный возраст раннесукцессионных видов деревьев составляет 60–90 лет – Populus tremula (88 лет), Carpinus betulus (80 лет), Cerasus avium (55 г). Все раннесукцессионные виды деревьев имеют пониженную жизненность, их онтогенетические спектры прерывистые и инвазионно-регрессивные, эти деревья постепенно отмирают и выпадают из состава сообщества (Горнов и др., 2018). Средний запас стволовой древесины составляет 319 ± 87 м3 га–1, при этом на долю Carpinus betulus приходится от 40 до 50% всего запаса древесины, Abies nordmanniana – 20–30 и Fagus orientalis – 20–25%.
Рис. 2.
Эколого-ценотические группы растений на разных стадиях сукцессии. Стадии сукцессии хвойно-широколиственных лесов: I – ранняя, II – переходная, III – поздняя. Эколого-ценотические группы: 1 – бореальные; 2 – неморальные; 3 – лугово-опушечные; 4 – нитрофильные; 5 – байрачные; 6 – скальные виды.
В пихтово-буковых мертвопокровных сообществах поздней стадии полночленный онтогенетический спектр имеется лишь у Abies nordmanniana. У Fagus orientalis спектр прерывистый (Горнов и др., 2018), что можно объяснить способностью подроста бука в условиях крайне низкой освещенности долго находиться в имматурном состоянии, не переходя в виргинильное (Helliwell, 2012). При увеличении уровня освещенности в условиях оконной динамики подрост бука быстро выходит в верхний древесный ярус, обгоняя в росте пихту (Ammer et al., 2008). Максимальный абсолютный возраст Abies nordmanniana достигает 327 лет (на соседних участках – до 450 лет), а Fagus orientalis – 186 лет. На поздней стадии резко увеличивается средний запас стволовой древесины (рис. 1) – до 1097 ± 265 м3 га–1за счет существенно увеличения высоты древесного полога – до 60 м и увеличения диаметра деревьев. При этом в буково-пихтовых мертвопокровных сообществах поздней стадии на долю Fagus orientalis приходится от 66 до 82% всего запаса стволовой древесины, а Abies nordmanniana – от 16 до 32%. Таким образом, в позднесукцессионных лесах Северо-Западного Кавказа при переходе от средней к поздней стадии демутационной сукцессии в возрасте 150–180 лет происходит значительное депонирование углерода за счет резкого увеличения общего запаса стволовой древесины.
Кислотность, содержание азота и углерода в почвах
Результаты оценки влияния стадий сукцессий на характеристики кислотности почв с использованием v-критерия демонстрируют, что самым высоким значением рН подгоризонта L подстилки отличались леса ранней стадии сукцессии (осиново-грабовые жимолостно-мелкотравные): (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ = 6.3), а самым низким – леса переходной стадии (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ = 5.1). Леса поздней стадии занимали промежуточное положение (табл. 1). Подгоризонт опада буково-пихтово-грабовых лесов (переходная стадия) отличается самой высокой актуальной и обменной кислотностью. Природу высокой кислотности подгоризонта L переходной стадии можно объяснить комбинированным влиянием опада пихты, граба и бука. Известно, что уровень рН буковой подстилки низкий (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ = 4.5 (Anderson, 1973); опад бука характеризуется высоким содержанием вторичных метаболитов (танинов и др.) и низкой скоростью разложения. На примере таежных лесов показано, что молодые деревья ели (40–50 лет) оказывают подкисляющее воздействие на подстилку. Старовозрастные деревья оказывают обратное воздействие, что определяется (1) накоплением в высоковозрастной опадающей хвое старых деревьев не способного к ретранслокации кальция внутри дерева, а также (2) формированием ими плотного полога, слабо пропускающего осадки, что предотвращает вымывание оснований из почв (Лукина и др., 2008). Поскольку пихта, как и ель, является позднесукцессионным видом и имеет сходные с елью биологические особенности, можно предположить, что в данном случае также наблюдается подкисляющее влияние опада обильного подроста пихты возраста 5–10 лет. Существенный вклад в повышенное содержание обменного алюминия в органогенных горизонтах вносит граб, листья которого накапливают алюминий в два раза больше, чем листья бука (Kooijman et al., 2018). Верхний подгоризонт подстилки на начальной стадии отличается высокой степенью насыщенности основаниями, которая постепенно снижается к поздней стадии.
Таблица 1.
Кислотность, содержание углерода и азота в почвах на разных сукцессионных этапах хвойно-широколиственных лесов
| Показатель | v-критерий | Среднее по горизонту | Стандартное отклонение по горизонту | Общее среднее | Общее стан-дартное откло-нение | р-value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | I | II | III | I | II | III | |||
| Подгоризонт L (nI = 9, nII = 9, nIII = 15) | ||||||||||||||
| Собщ, % | 1.0 | 1.3 | –2.1 | 38.7 | 39.0 | 35.0 | 3.4 | 3.1 | 6.2 | 37.1 | 5.2 | 0.30 | 0.20 | 0.04 |
| Nобщ, % | 2.8 | 0.2 | –2.7 | 1.75 | 1.58 | 1.44 | 0.21 | 0.14 | 0.21 | 1.56 | 0.23 | 0.00 | 0.83 | 0.01 |
| C : N | –2.1 | 1.2 | 0.8 | 22 | 25 | 24 | 1 | 2 | 3 | 24 | 3 | 0.03 | 0.22 | 0.42 |
| ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | 3.5 | –3.2 | –0.4 | 6.3 | 5.1 | 5.6 | 0.6 | 0.2 | 0.1 | 5.6 | 0.6 | 0.00 | 0.00 | 0.70 |
| ОК, смоль(+) кг–1 | –3.0 | 4.2 | –1.8 | 4.2 | 7.3 | 5.0 | 0.4 | 0.7 | 0.8 | 5.5 | 1.5 | 0.00 | 0.00 | 0.07 |
| Обм. Аl, смоль(+) кг–1 | –1.4 | 3.6 | –3.2 | 1.0 | 2.2 | 0.8 | 0.7 | 0.7 | 0.2 | 1.4 | 0.8 | 0.16 | 0.00 | 0.00 |
| BS, % | 4.4 | –2.8 | –2.4 | 95 | 90 | 91 | 1 | 1 | 1 | 92 | 2 | 0.00 | 0.01 | 0.02 |
| Подгоризонт FН (nI = 9, nII = 9, nIII = 15) | ||||||||||||||
| Собщ, % | 1.6 | 1.9 | –3.1 | 27.4 | 28.1 | 20.3 | 7.3 | 5.7 | 4.1 | 24.4 | 6.7 | 0.11 | 0.06 | 0.00 |
| Nобщ, % | 2.5 | 1.9 | –4.0 | 1.45 | 1.39 | 0.90 | 0.34 | 0.25 | 0.20 | 1.18 | 0.36 | 0.01 | 0.06 | 0.00 |
| C : N | –3.0 | –0.8 | 3.4 | 19 | 20 | 23 | 1 | 2 | 2 | 21 | 3 | 0.00 | 0.44 | 0.00 |
| ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | 2.8 | 1.9 | –6.9 | 6.8 | 6.7 | 6.1 | 0.1 | 0.0 | 0.1 | 6.5 | 0.3 | 0.01 | 0.06 | 0.00 |
| ОК, смоль(+) кг–1 | –3.5 | 3.3 | 0.3 | 1.4 | 3.3 | 2.4 | 0.4 | 0.9 | 0.3 | 2.4 | 1.0 | 0.00 | 0.00 | 0.73 |
| Обм. Аl, смоль(+) кг–1 | –2.6 | 4.0 | –2.1 | 0.6 | 1.7 | 0.8 | 0.4 | 0.4 | 0.1 | 1.0 | 0.6 | 0.01 | 0.00 | 0.04 |
| BS, % | 4.1 | –1.2 | –4.3 | 98 | 95 | 95 | 1 | 1 | 0 | 96 | 1 | 0.00 | 0.25 | 0.00 |
| Горизонт А (nI = 15, nII = 15, nIII = 15) | ||||||||||||||
| Собщ, % | 2.0 | 2.8 | –4.8 | 4.8 | 5.0 | 2.8 | 1.1 | 0.9 | 0.9 | 4.2 | 1.4 | 0.04 | 0.00 | 0.00 |
| Nобщ, % | 2.3 | 2.1 | –4.3 | 0.42 | 0.41 | 0.26 | 0.10 | 0.07 | 0.07 | 0.36 | 0.11 | 0.02 | 0.04 | 0.00 |
| C : N | 0.7 | 3.3 | –4.0 | 12 | 12 | 10 | 1 | 1 | 1 | 11 | 1 | 0.50 | 0.00 | 0.00 |
| ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | –0.4 | 1.3 | –0.9 | 5.0 | 5.1 | 5.0 | 0.3 | 0.2 | 0.2 | 5.1 | 0.2 | 0.70 | 0.20 | 0.36 |
| ОК, смоль(+) кг–1 | 0.3 | –1.1 | 0.7 | 2.9 | 2.5 | 3.0 | 0.9 | 1.3 | 1.9 | 2.8 | 1.4 | 0.73 | 0.28 | 0.45 |
| Обм. Аl, смоль(+) кг–1 | 0.3 | –1.1 | 0.8 | 2.5 | 2.1 | 2.7 | 0.9 | 1.3 | 1.8 | 2.4 | 1.4 | 0.79 | 0.29 | 0.43 |
| BS,% | 0.0 | 1.8 | –1.8 | 65 | 73 | 58 | 13 | 14 | 23 | 66 | 18 | 0.98 | 0.06 | 0.07 |
| Горизонт В (nI = 6, nII = 15, nIII = 9) | ||||||||||||||
| Собщ, % | 0.6 | 0.1 | –0.8 | 0.6 | 0.6 | 0.5 | 0.2 | 0.2 | 0.2 | 0.6 | 0.2 | 0.53 | 0.89 | 0.44 |
| Nобщ, % | –2.6 | 1.8 | 0.8 | 0.08 | 0.10 | 0.09 | 0.01 | 0.01 | 0.02 | 0.09 | 0.02 | 0.01 | 0.07 | 0.44 |
| C : N | 3.0 | –1.0 | –2.0 | 8 | 6 | 5 | 1 | 1 | 2 | 6 | 2 | 0.00 | 0.31 | 0.04 |
| ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | 2.4 | –1.3 | –1.1 | 5.6 | 5.3 | 5.3 | 0.3 | 0.1 | 0.2 | 5.4 | 0.3 | 0.02 | 0.20 | 0.27 |
| ОК, смоль(+) кг–1 | 0.3 | 3.5 | –3.8 | 4.8 | 6.1 | 3.1 | 0.6 | 1.1 | 1.1 | 4.7 | 1.6 | 0.78 | 0.00 | 0.00 |
| Обм. Аl, смоль(+) кг–1 | 0.3 | 3.5 | –3.9 | 4.6 | 5.9 | 2.9 | 0.6 | 1.1 | 1.1 | 4.5 | 1.5 | 0.76 | 0.00 | 0.00 |
| BS, % | –0.4 | –2.1 | 2.5 | 48 | 43 | 58 | 6 | 12 | 12 | 50 | 12 | 0.65 | 0.04 | 0.01 |
Примечание. Стадия сукцессии: I – ранняя; II – переходная; III – поздняя; n – объем выборки; Собщ, % – содержание углерода; Nобщ, % – содержание азота; C : N – отношение углерода к азоту; ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ – актуальная кислотность; ОК, смоль(+) кг–1 – обменная кислотность; Обм. Аl, смоль(+) кг–1 – обменный алюминий; BS,% – насыщенность основаниями.
Подгоризонты FH на поздней стадии с доминированием бука и пихты, формирующих трудноразлагаемый опад, отличались самым низким значением рН при сопоставимых уровнях на ранней и переходной стадиях. При этом содержание обменного алюминия, так же как и в верхнем L-подгоризонте, достигает самого высокого уровня на переходной стадии, где вклад граба в формирование подстилки достигает максимума, что обусловливает и самую высокую обменную кислотность.
Актуальная кислотность минеральных горизонтов А и В между стадиями сукцессии достоверно не различалась. При этом следует отметить, что содержание обменного алюминия в минеральном профиле почв на начальной и переходной стадиях достигало максимума в иллювиальном горизонте, а на поздней стадии относительно равномерно распределялось по профилю, что связано с более ярко выраженными процессами оглеения. В иллювиальном горизонте содержание обменного алюминия и обменная кислотность оказались достоверно выше на переходной стадии сукцессии и ниже на поздней.
Содержание азота в L-подгоризонте на ранней стадии оказалось достоверно выше, а углерода сопоставимо по сравнению с другими стадиями. Отношение C : N существенно уже на ранней стадии и не отличалось между промежуточной и поздней стадиями. Аналогичные закономерности для отношения C : N наблюдаются и в подгоризонте FH.
Выявленные различия объясняются качеством опада: на начальной стадии опад граба, жимолости, мелкотравья богат азотом, а на поздней стадии опад бука и пихты содержит меньше азота и больше вторичных метаболитов. Опад низкого качества разлагается медленнее, накопление углерода происходит за счет накопления в подстилках вторичных метаболитов.
В гумусовом горизонте А на поздней стадии происходит снижение содержания и углерода, и азота, но при этом происходит и сужение отношения C : N: 10 против 12 на переходной и ранней стадиях. Снижение содержания биогенов на поздней стадии в горизонте А объясняется низкой скоростью разложения опада низкого качества, отсутствием нижнего травяно-кустарничкового яруса и, соответственно, его корневого опада. Сходные тенденции снижения содержания углерода в горизонте А в буковых лесах наблюдали и в других исследованиях (Aubert et al., 2006; Kramer, 2009; Langenbruch, 2012).
Относительное обогащение гумуса азотом можно объяснить также деятельностью почвенной мезофауны. Так, согласно нашим данным (табл. 2), на поздней стадии биомасса норных дождевых червей (Dendrobaena mariupoliensis) возрастает почти в восемь раз по сравнению с предыдущими стадиями. Как известно, норные черви на поверхности почвы оставляют копролиты с повышенным содержанием азота (Битюцкий, 2007), их деятельность приводит к относительному обогащению гумуса азотом. Также на этой стадии значимо выше численность и биомасса собственно почвенных дождевых червей (главным образом, Dendrobaena shmidti shmidti), которые как вторичные гумусопотребители могут способствовать снижению содержания почвенного углерода в гумусовом горизонте и обогащению органического вещества азотом. В горизонте В наблюдается более широкое отношение C : N на ранней и переходной стадиях по сравнению с поздней. Благодаря преобладанию на ранней и переходной стадиях легкоразлагаемого лиственного и травяного опада, а также развитой сети корней мелкотравья, происходит более интенсивная минерализация органического вещества подстилки. Углерод, высвобожденный в результате разложения в подстилках, мигрирует и накапливается в минеральных горизонтах, тогда как азот активно поглощается растениями.
Таблица 2.
Численность и биомасса (Х ± SE) основных видов дождевых червей (сем. Lumbricidae) в почвах на разных сукцессионных этапах хвойно-широколиственных лесов
| Вид | Стадия | |||||
|---|---|---|---|---|---|---|
| I стадия | II стадия | III стадия | ||||
| численность, экз. м–2 | биомасса, г м–2 | численность, экз. м-2 | биомасса, г м–2 | численность, экз. м–2 | биомасса, г м–2 | |
| Dendrobaena octaedra | 1.3 ± 0.17 | 0.02 ± 0.01 | 7.0 ± 1.6 | 0.44 ± 0.21 | 1.67 ± 0.33 | 0.69 ± 0.01 |
| D. shmidti shmidti | 17.7 ± 2.9 | 3.92 ± 0.96 | 22.0 ± 2.5 | 2.94 ± 0.29 | 22.63 ± 8.62 | 6.81 ± 1.72 |
| D. mariupoliensis | + | + | 2.0 ± 0.2 | 0.8 ± 0.14 | 6.7 ± 3.3 | 7.22 ± 4.1 |
Примечание. “+” – вид найден только в древесине валежа. I–III – см. табл. 1.
Изменения запасов углерода в ходе сукцессионного развития лесов
В ходе сукцессионного развития лесов происходит закономерное изменение запасов углерода, что объясняется изменением и содержания углерода в горизонтах почв, и запасов подстилки, а также плотностью и мощностью минеральных горизонтов почв. Общие запасы углерода в профиле почв LFH-A-AB-B на ранней и промежуточной стадиях сукцессий не различались и составляли 74 и 78 т/га соответственно. На поздней стадии происходит существенное снижение запасов до 61 т/га (табл. 3).
Таблица 3.
Характеристика запасов углерода (т/га) почв на разных сукцессионных этапах хвойно-широколиственных лесов
| Запасы углерода (т га–1) | v-критерий | Среднее | Стандартное отклонение | Общее среднее | Общее стандартное отклонение | р value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | I | II | III | I | II | III | |||
| Запасы C подгоризонта L | –2.1 | –2.7 | 4.3 | 1.4 | 1.3 | 2.8 | 0.1 | 0.2 | 0.8 | 2 | 0.9 | 0.035 | 0.006 | 0 |
| Запасы C подгоризонта FH | –0.3 | 0.4 | –0.1 | 1.4 | 1.5 | 1.4 | 0.4 | 0.4 | 0.2 | 1.4 | 0.3 | 0.78 | 0.673 | 0.899 |
| Запасы C горизонта А | 2.1 | 2.5 | –4.6 | 42.5 | 43.2 | 24.5 | 10.3 | 8.9 | 7.6 | 36.6 | 12.5 | 0.035 | 0.013 | 0 |
| Запасы C горизонта АВ | –2.2 | 1.7 | 0.5 | 9.0 | 15.9 | 13.8 | 4.1 | 11.7 | 5 | 13 | 8.3 | 0.031 | 0.096 | 0.652 |
| Запасы C горизонта В | 1.1 | –1.5 | 0.3 | 19.3 | 15.7 | 18.3 | 6.1 | 1.4 | 6 | 17.8 | 5.3 | 0.272 | 0.134 | 0.739 |
| Запасы C в 10 см горизонта ВС | 0.4 | 1.4 | –1.6 | 4.7 | 5.2 | 3.5 | 0.1 | 4.1 | 1.5 | 4.3 | 2.8 | 0.681 | 0.176 | 0.101 |
| Запасы С подстилки (LFH) | –1.6 | –1.5 | 3.1 | 2.7 | 2.8 | 4.7 | 0.4 | 0.6 | 1.2 | 3.4 | 1.2 | 0.105 | 0.132 | 0.002 |
| Запасы C в слое 0–30 см | 0.8 | 1.9 | –2.8 | 57.5 | 62.1 | 42.9 | 7.8 | 4.2 | 12.2 | 54.2 | 11.9 | 0.416 | 0.052 | 0.006 |
| Запасы C в слое 0–50 см | 0.8 | 1.5 | –2.3 | 68.7 | 71.9 | 55.5 | 8.9 | 4.4 | 15.6 | 65.3 | 12.9 | 0.446 | 0.135 | 0.024 |
| Запасы C в слое 50–100 см | 1.9 | –0.2 | –1.7 | 23.7 | 19.7 | 16.9 | 5 | 0.6 | 6.2 | 20.1 | 5.4 | 0.051 | 0.812 | 0.087 |
| Запасы C в слое 0–100 см | 1.2 | 1.1 | –2.3 | 92.4 | 91.6 | 72.3 | 12.1 | 4.7 | 20.7 | 85.5 | 16.9 | 0.229 | 0.287 | 0.023 |
| Запасы C в слое LFH–100 см | 1.1 | 1 | –2.1 | 95.2 | 94.4 | 77.0 | 12.1 | 4.7 | 21 | 88.9 | 16.5 | 0.267 | 0.328 | 0.037 |
Примечание. I–III – см. табл. 1.
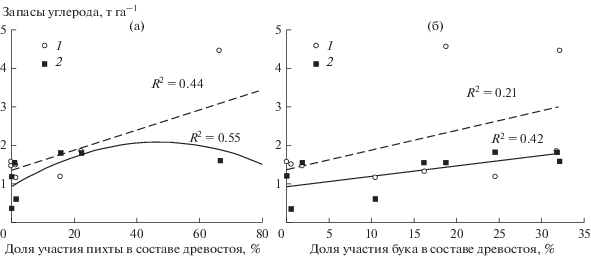
Согласно полученным результатам, на поздней стадии сукцессии в подстилках происходило существенное накопление углерода как за счет увеличения концентрации углерода, так и вследствие возрастания массы подстилки. Выявлена положительная зависимость запасов углерода в подстилке от вклада в состав древостоя бука и пихты: с увеличением доли этих деревьев в составе древостоя увеличиваются запасы углерода в подстилке. Это обусловлено тем, что опад бука и пихты отличается низким качеством (высокое содержание лигнинов и танина, высокая кислотность) (рис. 3), обеспечивающее снижение скорости разложения органического вещества. На ранней и переходной стадиях запасы углерода в подстилке варьировали от 1.8 до 3.7 т га–1, на поздней стадии они составляли в среднем 4.7 т га–1. При этом вклад подстилки в общие запасы углерода возрастал от 4.2 и 3.9% на начальной и переходной стадиях до 10% на поздней стадии, где в подгоризонте L содержалось до 70% запасов подстилки.
Рис. 3.
Зависимость запасов углерода подстилки от доли участия пихты (а) и бука (б) в составе древостоя в лесах Северо-Западного Кавказа. 1 – запасы углерода в подгоризонте L; 2 – запасы углерода в подгоризонте FH.
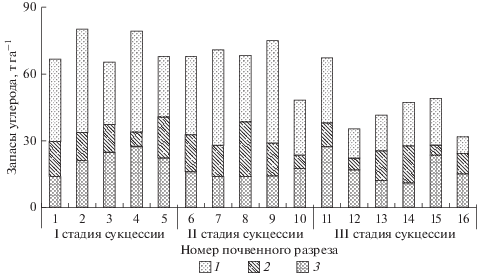
Запасы углерода в минеральных горизонтах почв в почвенном профиле на поздней стадии сукцессии существенно снижаются (рис. 4): если на ранней и переходной стадиях запасы углерода в них варьировали от 49 до 80 т га–1, то на поздней стадии в большинстве случаев они составляли от 32 до 49 т га–1. Исключением является одна пробная площадь, на которой запасы углерода в гумусовом горизонте достигают значений первых двух стадий сукцессии, что приводит к увеличению общих запасов углерода до 68 т га–1. Это может объясняться наличием трав, которые определяют повышение массы корневого опада – источника углерода в горизонте А. Наличие трав обусловлено более высоким уровнем освещенности данной пробной площади. Следует отметить, что такие участки на поздней стадии – редкое явление. В горизонтах АВ и В варьирование запасов углерода существенно, но при этом закономерных изменений не обнаруживается. Максимальный вклад в запасы углерода почвы вносит горизонт А. Так, доля этого горизонта в общем запасе углерода изменяется в среднем от 57–59% на ранней стадии до 40% на поздней. Велика также доля переходного горизонта АВ, варьирующая от 16 до 22%, тогда как доля горизонта В достигает 23 и 22% на начальной и переходной стадиях, соответственно, и возрастает до 30% на поздней.
Рис. 4.
Запас почвенного углерода на разных стадиях сукцессии, т га–1. I–III – см. рис. 1. 1 – гумусовый горизонт A; 2 – переходный горизонт AB; 3 – нижний минеральный горизонт B.
Вклад горизонта ВС, отличающегося от других минеральных существенно более высокой плотностью, в общие запасы углерода незначителен и составляет 5–6%, не различаясь (как и содержание углерода и азота) между стадиями сукцессии. При этом, как и в горизонте В, отношение C/N достоверно выше на ранней стадии и ниже на поздней стадии сукцессии.
Таким образом, снижение общих запасов углерода в почвах на поздней стадии сукцессии в лесах Северо-Западного Кавказа связано со снижением запасов в гумусовом горизонте почв, тогда как запасы в подстилке, напротив, возрастали.
Сравнение запасов углерода с учетом фиксированного слоя минерального профиля мощностью 0–100 см демонстрирует, что меньше всего почвенного углерода (72 т га–1) аккумулировано на поздней стадии сукцессии, тогда как на начальной и промежуточной стадиях запасы существенно не различались, составляя соответственно около 92 т га–1. При этом запасы углерода в подстилке были значительно выше на поздней стадии: 4.7 т га–1 против 2.7 и 2.8 на начальных и промежуточных стадиях.
Заключение. 1. Раннесукцессионными сообществами послерубочной сукцессии хвойно-широколиственных лесов Северо-Западного Кавказа являются осиново-грабовые жимолостно-мелкотравные сообщества с преобладанием видов неморального мелкотравья и высокотравья; переходная стадия представлена буково-пихтово-грабовым мелкотравьем с преобладанием неморальных мелких трав и кустарников; поздняя стадия – олигодоминантными пихтово-буковыми сообществами с неразвитым напочвенным покровом.
2. Многомерный анализ геоботанических описаний разных стадий послерубочной сукцессии по экологическим шкалам Г. Элленберга выявил приуроченность раннесукцесионных сообществ к участкам с повышенной освещенностью, низкой кислотностью, высоким увлажнением и богатством почвы азотом по сравнению с сообществами переходной и поздней стадий. Сообщества поздней стадии приурочены к кислым и бедным азотом почвам с преобладанием теневыносливых и холодостойких видов растений. Сообщества переходной стадии занимают промежуточное положение между сообществами ранней и поздней стадий по экологическим шкалам.
3. На поздней стадии сукцессии хвойно-широколиственных лесов происходит возрастание запасов углерода в подстилке и их снижение в гумусовом горизонте А – основном аккумуляторе углерода в почвах. Изменения запасов углерода в этих горизонтах почв обусловлены сменой доминирования древесных растений, формирующих опад разного качества, т.е. с разным содержанием азота и вторичных метаболитов. Отношение C : N в горизонте А регулируется как качеством опада, так биомассой и численностью почвенных червей.
4. Показано, что без учета вклада нижних горизонтов почв В и ВС, залегающих на глубине от 20 до 80 см и ниже, достоверная оценка запасов почвенного углерода невозможна, поскольку вклад этих горизонтов в общие запасы углерода в почвах может превышать 20%.
5. Количественная оценка характеристик почв, таких как кислотность, богатство азотом, демонстрирует сходство обнаруженных трендов в ходе сукцессионного развития почв при сравнении с изменениями растительности, выявленными при применении экологических шкал Г. Элленберга.
Список литературы
Битюцкий Н.П., Соловьева А.Н., Лукина Е.И., Олейник А.С., Завгородняя Ю.А., Демин В.В., Бызов Б.А. Экскреты дождевых червей – стимулятор минерализации соединений азота в почве // Почвоведение. 2007. № 4. P. 468–473.
Биота экосистем Большого Кавказа / Под ред. Зиминой Р.П., Злотина Р.И. М.: Наука, 1990. 221 с.
Геология СССР. Геологическое описание. Северный Кавказ. / Под ред. В.Л. Андрущука, А.Я. Дубинского, В.Е. Хаина. М.: Недра, 1968. Т. 9. Ч. 1. 760 с.
Гиляров М.С. Учет крупных беспозвоночных (мезофауна) // Количественные методы в почвенной зоологии / Под ред. М.С. Гилярова, Б.Р. Стригановой. М.: Наука, 1987. С. 9–26.
Голгофская К.Ю., Горчарук Л.Г., Егорова С.В. К изучению взаимоотношений некоторых компонентов горно-лесных биогеоценозов Кавказского заповедника // Труды Кавказского заповедника. 1967. Вып. 9. С. 59–118.
Горнов А.В., Горнова М.В., Тихонова Е.В., Шевченко Н.Е., Казакова А.И., Ручинская Е.В., Тебенькова Д.Н. Оценка сукцессионного статуса хвойно-широколиственных лесов европейской части России на основе популяционного подхода // Лесоведение. 2018. № 6. С. 16–30.
Евстигнеев О.И., Коротков В.Н., Бакалина Л.В. Популяционная организация грабовых лесов Каневского заповедника // Бюллетень Московского общества испытателей природы. Отдел биологический. 1992. Т. 97. № 2. С. 81–89.
Методические подходы к экологической оценке лесного покрова в бассейне малой реки / Под ред. Заугольновой Л.Б., Браславской Т.Ю. М.: Товарищество научных изданий КМК, 2010. 383 с.
Зонн С.В. Горно-лесные почвы северо-западного Кавказа. Л.: Изд-во АН СССР. 1950. 334 с.
Луганский Н.А., Залесов С.В., Луганский В.Н. Лесоведение. Екатеринбрг: Уральский гос. лесотех. университет, 2010. 432 с.
Лукина Н.В., Полянская Л.М., Орлова М.А. Питательный режим почв северотаежных лесов. М.: Наука, 2008. 341 с.
Лукина Н.В., Орлова М.А., Бахмет О.Н., Тихонова Е.В., Тебенькова Д.Н., Казакова А.И., Крышень А.М., Горнов А.В., Смирнов В.Э., Шашков М.П., Ершов В.В., Князева С.В. Влияние растительности на характеристики лесных почв Республики Карелия // Почвоведение. 2019. № 5. (в печати).
О методических указаниях по количественному определению объема поглощения парниковых газов: Распоряжение Министерства природных ресурсов и экологии РФ от 30 июня 2017 г.
Смирнова О.В., Попадюк Р.В., Чистякова А.А. Популяционные методы определения минимальной площади лесного ценоза // Ботанический журн. 1988. Т. 73. № 10. С. 1423–1434.
Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов Европейской России // Успехи современной биологии. 2006. Т. 126. № 1. С. 26–48.
Приказ Рослесхоза № 10: Справочник лесотаксационных нормативов для Северного Кавказа. Утверждено 19.01.1995.
Теория и практика химического анализа почв / Под ред. Л.А. Воробьевой. М.: ГЕОС, 2006. 400 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). Cambridge: Cambridge university press, 1995, 516 p.
Ammer C., Stimm B., Mosandl R. Ontogenetic variation in the relative influence of light and belowground resources on European beech seedling growth // Tree Physiology. 2008. V. 28. № 5. P. 721–728.
Anderson J.M. The breakdown and decomposition of sweet chestnut (Castanea sativa Mill.) and beech (Fagus sylvatica L.) leaf litter in two deciduous woodland soils // Oecologia. 1973. V. 12. № 3. P. 275–288.
Aubert M., Margerie P., Ernoult A., Decaëns T., Bureau F. Variability and heterogeneity of humus forms at stand level: comparison between pure beech and mixed beech-hornbeam forest // Annals of Forest Science. 2006. V. 63. № 2. P. 177–188.
Christensen N., Kismul V., Sisula H., Giege B. Methods for integrated monitoring in the Nordic countries. Copenhagen: Nordic Council of Ministers, 1989. 280 p.
Ellenberg H. Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht. 5. Stuttgart: Ulmer Eugen, 1996. 1096 s.
Framstad E., Wit H., Mäkipää R., Larjavaara M., Vesterdal L., Karltun E. Biodiversity, carbon storage and dynamics of old northern forests. Copenhagen: Nordic Council of Ministers, 2013. 130 p.
Gebauer T. Water turnover in species-rich and species-poor deciduous forests: xylem sap flow and canopy transpiration: Dissertation zur Erlangung des Doktorgrades. Göttingen: GAU, 2010. 128 p.
Halonen O., Tulkki H., Derome J. Nutrient analysis methods // Metsäntutkimuslaitoksen tiedonantoja. 1983. V. 121. P. 1–28.
Helliwell D.R. Plant growth and daylight in woodland; the role of diffuse and direct sunlight // Quarterly J. Forestry. 2012. V. 106. № 1. P. 37–42.
Husson F., Le S., Pages J. Exploratory multivariate analysis by example using R. Boca Raton: CRC Press, 2017. 248 p.
Ignatov M.S., Afonina O.M., Ignatova E.A., Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A.G., Boychuk M.A., Cherdantseva V.Y., Czernyadjeva I.V., Doroshina G.Y., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Kharzinov Z.K., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K., Manakyan V.A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Y., Pisarenko O.Y., Popova N.N., Rykovsky G.F., Tubanova D.Y., Zheleznova G.V., Zolotov V.I. Checklist of mosses of East Europe and North Asia // Arctoa. 2006. V. 15. P. 1–130.
Kooijman A.M., Smit A. Relationships between forest vegetation, parent material and soil development in the Luxembourg cuesta landscape / The Luxembourg Gutland Landscape. Cham: Springer, 2018. P. 153–176.
Krämer I. Rainfall partitioning and soil water dynamics along a tree species diversity gradient in a deciduous old-growth forest in Central Germany. Dissertation zur Erlangung des Doktorgrades. Göttingen: GAU, 2009, 162 p.
Langenbruch C. Effects of nutrient cycling through litter of different broadleaved deciduous tree species on soil biochemical properties and the dynamics of carbon and nitrogen in soil. Dissertation zur Erlangung des Doktorgrades. Göttingen: GAU, 2012, 93 p.
Lewis S.L., Lopez-Gonzalez G., Sonké B., Affum-Baffoe K., Baker T.R., Ojo L.O., Phillips O.L., Reitsma J.M., White L., Comiskey J.A., K M.-N.D., Ewango C.E.N., Feldpausch T.R., Hamilton A.C., Gloor M., Hart T., Hladik A., Lloyd J., Lovett J.C., Makana J.-R., Malhi Y., Mbago F.M., Ndangalasi H.J., Peacock J., Peh K.S.H., Sheil D., Sunderland T., Swaine M.D., Taplin J., Taylor D., Thomas S.C., Votere R., Wöll H. Increasing carbon storage in intact African tropical forests // Nature. 2009. V. 457. № 7232. P. 1003–1006.
Luyssaert S., Schulze E.-D., Börner A., Knohl A., Hessenmöller D., Law B., Ciais P., Grace J. Old-growth forests as global carbon sinks // Nature. 2008. V. 455. № 7210. P. 213–215.
Vashum K.T., Jayakumar S. Methods to estimate above-ground biomass and carbon stock in natural forests – a review // J. Ecosystem & Ecography. 2012. V. 2. № 4. Article 116.
World reference base for soil resources 2014. International soil classification system for naming soils and creating legends for soil maps. Rome: FAO, 2014. 181 p.
www.cepl.rssi.ru/bio/forest/ (дата обращения: 08.11.2017).
www.R-project.org (дата обращения: 01.04.2018).
Zhou G.-Q., Zhang Y., Ferguson D.J.P., Chen S., Rasmuson-Lestander Å., Campbell F.C., Watt S.M. The drosophila ortholog of the endolysosomal membrane protein, endolyn, regulates cell proliferation // J. Cellular Biochemistry. 2006. V. 99. № 5. P. 1380–1396.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение