

Лесоведение, 2019, № 5, стр. 341-356
Мультифункциональность и биоразнообразие лесных экосистем
Д. Н. Тебенькова a, *, Н. В. Лукина a, С. И. Чумаченко b, М. А. Данилова a, А. И. Кузнецова a, А. В. Горнов a, Н. Е. Шевченко a, А. Д. Катаев a, Ю. Н. Гагарин a
a Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32, стр. 14, Россия
b Мытищинский филиал Московского государственного технического университета имени Н.Э. Баумана
141005 Московская обл., Мытищи, ул.1-я Институтская, 1, Россия
* E-mail: tebenkova.dn@gmail.com
Поступила в редакцию 07.12.2018
После доработки 04.04.2019
Принята к публикации 05.06.2019
Аннотация
В статье дан анализ современных представлений о мультифункциональности лесных экосистем, связях “мультифункциональность–биоразнообразие” и о подходах к оценке компромиссов и синергии между экосистемными функциями и услугами лесов. Актуальность изучения экосистемных услуг обусловлена современными потребностями общества, сложившимися в результате роста мировой экономики и возрастающей численности населения Земли. Лесные экосистемы предоставляют множество услуг одновременно, т.е. им свойственна мультифункциональность. Однако на сегодняшний день большинство исследований ограничивается изучением связи между отдельными экосистемными услугами и биоразнообразием. В связи с этим влияние потери биоразнообразия на экосистемные услуги значительно недооценено, что связано с отсутствием методических основ и инструментария для оценки мультифункциональности экосистем. В статье приводится концептуальная схема взаимосвязей “биоразнообразие–множество функций–множество услуг” с учетом факторов, влияющих на эти связи. Для развития междисциплинарной концепции взаимосвязей между биоразнообразием и мультифункциональностью лесов и ее практического применения в системах поддержки принятия управленческих решений необходимо решить ряд задач. К основным из них относятся идентификация информативных индикаторов связей между биоразнообразием и мультифункциональностью лесов, оценка компромиссов и синергии между различными функциями и услугами. Решение этих вопросов позволит не только получить новые фундаментальные знания о процессах функционирования лесных экосистем, но и создать рынки ранее не оцениваемых экосистемных услуг и обеспечить непрерывное и устойчивое лесопользование.
Леса относятся к наиболее распространенным наземным экосистемам, выполняющим множество экосистемных функций (ЭФ) и предоставляющим множество экосистемных услуг (ЭУ) одновременно. Согласно международному отчету “Оценка экосистем на грани тысячелетий”, ЭУ понимаются как “выгоды для людей, получаемые от экосистем” (Millennium…, 2005). Поставщиком ЭФ и ЭУ лесов является биоразнообразие (The Economics…, 2010). Широкое осознание ведущей роли экосистем в поддержании экономического и социального благополучия существенно повысило интерес к изучению и оценкам ЭУ лесов. Такие исследования особенно актуальны для России, на долю которой приходится 20% всех лесных ресурсов планеты, в том числе более половины бореальных лесов (Стратегия…, 2018). Леса занимают половину земель Российской Федерации с общим запасом древесины 82.7 млрд куб. м. (Российский…, 2018).
Актуальность изучения ЭУ обусловлена современными проблемами, вызванными ростом мировой экономики и возрастающей численностью населения Земли. Прежде всего, это – деградация лесов и потеря биоразнообразия, поскольку они неизбежно приводят к нарушениям функционирования лесных экосистем и предоставления ЭУ. По данным Международного союза охраны природы IUCN из 96 тыс. видов, под угрозой исчезновения находится 27% (The International…, 2018). Главные причины исчезновения видов - деградация и потеря экосистем вследствие изменения землепользования, антропогенного влияния на биогеохимические циклы; распространение инвазивных видов; несбалансированное управление и нещадная эксплуатация природных ресурсов (Louman et al., 2011).
Лесные экосистемы предоставляют множество услуг одновременно, т.е. им свойственна мультифункциональность. Они должны обеспечивать пищей, биоэнергией, секвестрировать углерод, сохранять биоразнообразие и противостоять природным экстремумам (штормам, наводнениям, оползням и т.д.) (Manning et al., 2015). Однако на сегодняшний день большинство исследований ограничивается изучением связи между отдельными ЭУ и биоразнообразием, поэтому влияние потери биоразнообразия на ЭУ может быть значительно недооценено, что связано с отсутствием методических основ и инструментария для оценки мультифункциональности экосистем.
К тому же оценка мультифункциональности неразрывно связана с анализом сложных взаимодействий между ЭФ и ЭУ, которые выражаются в компромиссах или синергии. Поэтому возникает необходимость в лучшем понимании, где и как возникают синергия и конфликты/компромиссы между ЭФ и ЭУ, и как можно их усиливать/избегать и управлять ими. Это особенно актуально в настоящее время, поскольку в России, как и в других лесных странах, взят курс на интенсификацию лесного хозяйства (Основы…, 2013), что связано с возрастающими потребностями в лесных продуктах и услугах и с политически поддерживаемым в развитых странах трендом перехода к биоэкономике, основанной на возобновляемых биологических ресурсах. Удовлетворение растущей потребности в лесной биомассе требует решения вопросов, связанных в первую очередь с возникающими конфликтами при предоставлении других услуг (рекреационные услуги, сток углерода) и сохранении биоразнообразия.
Цель данной работы – проанализировать современные представления о (1) мультифункциональности лесных экосистем, (2) связях “биоразнообразие – мультифункциональность” лесов, (3) подходах к оценке компромиссов и синергии между ЭФ и ЭУ лесов.
Несмотря на то, что истоки современной концепции ЭУ можно проследить еще с 1970-х годов прошлого столетия (Gómez-Baggethun et al., 2010), до сих пор нет общепринятого определения и классификации ЭУ, поэтому необходимо обозначить основные понятия и классификацию, используемые в настоящей статье:
ЭУ – выгоды для людей, получаемые от экосистем (Millennium…, 2005);
ЭФ – совокупность физических, биологических, химических и иных экосистемных процессов, которые поддерживают целостность и сохранение экосистем (Ansink et al., 2008);
экосистемные процессы – взаимодействия между биотическими и абиотическими элементами экосистем, лежащие в основе потоков информации, энергии и вещества (Puydarieux, Beyou, 2017).
В зависимости от вида пользы, которую леса приносят человеку, выделяют четыре категории ЭУ: (а) обеспечивающие (англ. provisioning) – продукция, получаемая от экосистем (пища, пресная вода, древесина, волокна, генетические ресурсы, медикаменты); (б) регулирующие (англ. regulating) – выгоды, получаемые от регулирования экосистемными процессами (регулирование изменений климата, природная очистка воды и др.); (в) культурные (англ. cultural) – выгоды, получаемые от экосистем в форме духовного обогащения, интеллектуального развития, рекреации, эстетических ценностей; (г) поддерживающие (англ. supporting) – жизнеобеспечивающие ЭУ, необходимые для поддержания других ЭУ, от которых люди получают прямую экономическую выгоду (почвообразование, круговорот воды и питательных веществ, фотосинтез) (Millennium…, 2005).
Классификация ЭУ носит условный характер, потому что все услуги взаимосвязаны. Например, R. Haines-Young и M. Potschin (2010) проиллюстрировали связь между ЭУ с использованием модели “каскада услуг”, где выстроена линейная связь “биоразнообразие → поддерживающие ЭУ → регулирующие ЭУ → обеспечивающие ЭУ → культурные ЭУ”. Однако данная схема является частным случаем взаимодействия ЭУ и не раскрывает всей многогранности их отношений и вытекающей из этого мультифункциональности лесных экосистем.
МУЛЬТИФУНКЦИОНАЛЬНОСТЬ ЛЕСНЫХ ЭКОСИСТЕМ
Традиционные исследования функционирования экосистем обычно основаны на детальных экспериментах, нацеленных на оценку влияния биоразнообразия на отдельные ЭФ или ЭУ и связанные с ними факторы. Такая стандартизированная численная оценка позволяет сравнивать на основе этих отдельных ЭФ (например, потоки углерода, продукция биомассы) различные экосистемы, но одни и те же ландшафты выполняют все ЭУ одновременно.
В настоящее время разрабатывается новый общий подход, который определяет мультифункциональность на двух уровнях (Manning et al., 2018): (1) первый уровень – мультифункциональность ЭФ, на оценку которой направлены фундаментальные исследования биологических, геохимических и физических процессов, происходящих в экосистемах; (2) второй уровень – мультифункциональность ЭУ, которая определяется как совместное предоставление ряда ЭУ в ответ на запрос общества. На первом уровне все ЭФ одинаково важны и равноценны, на втором уровне приоритетность услуг определяется человеком. При этом встает вопрос разграничения функций и услуг. Часто одно и то же явление может рассматриваться и как функция, и как услуга. Например, формирование гидрологического режима представляет собой функцию, поскольку леса влияют на уровень грунтовых вод, вынос углерода и других биогенных элементов из почвы. В то же время эта функция может быть услугой с точки зрения обеспечения населения пресной водой.
Для предотвращения деградации лесов необходимы фундаментальные научные исследования, направленные на поиск связей между биоразнообразием – поставщиком всех ЭУ и мультифункциональностью лесных экосистем. Концепция мультифункциональности экосистем и связи “биоразнообразие – мультифункциональность” оценивалась в серии экспериментов. Показано, что для обеспечения мультифункциональности необходимо большее число видов, чем для единичных функций/услуг (Hector, Bagchi, 2007). В то же время уменьшение числа видов сильнее влияет на мультифункциональность, чем на отдельные ЭФ/ЭУ (Gamfeldt et al., 2008). При этом негативный эффект может сильнее проявляться на более высоких пространственных уровнях (Hautier et al., 2018).
Для оценок использовались дистанционные или наземные измерения (Ratcliffe et al., 2016), механистические модели (Mc Guire et al., 2001), косвенные методы, например, в тех случаях, когда предполагается, что некоторые типы мест обитания поддерживают определенные ЭФ (Maskell et al., 2013), или комбинация вышеназванных методов (Mouchet et al., 2017). Однако проведена оценка далеко не всех важных ЭУ, что ограничивает понимание мультифункциональности экосистемы, в первую очередь регулирующих ЭУ.
В настоящее время не существует единого подхода к оценкам связи между биоразнообразием и мультифункциональностью. Выделяют четыре основных подхода (Byrnes et al., 2014): первый подход – учет единичных ЭУ, который рассматривает набор функций и качественно отвечает на вопрос, достигают ли функции более высоких значений при увеличении уровня видового разнообразия. Анализ одномерных ответов дает информацию о взаимосвязях ЭУ с биоразнообразием, но не позволяет количественно оценить мультифункциональность. Второй подход – метод оборота (англ. turnover) (Hector, Bagchi, 2007). Сначала определяется набор видов, которые вносят вклад в каждую функцию. Затем оценивается избыточность вкладов видов путем оценки их перекрытия. Таким методом можно оценить взаимосвязь между числом функций и совокупным числом видов. Третий подход – усреднение или суммирование ЭФ, когда используется сумма стандартизированных значений каждой измеренной ЭФ (Maestre et al., 2012). Таким образом, положительная связь “биоразнообразие–мультифункциональность” может быть оценена на основе статистического усреднения влияния единичных ЭФ. Четвертый – пороговый подход, учитывает число ЭФ, которые перешли порог или диапазон порогов, обычно выражаемых как процент самого высокого уровня ЭФ, наблюдаемого в конкретном исследовании (Byrnes et al., 2014). При совместном использовании подхода усреднения и порогового подхода для оценки связей “биоразнообразие–мультифункциональность” всех доступных измерений ЭФ и ЭУ и смешивая показатели состояния и процессов исследователи придают равный вес всем переменным (Soliveres et al., 2016). Это не вызывает вопросов при рассмотрении естественных экосистем, развивающихся без прямого вмешательства человека, но такое смешивание неприемлемо при оценках ЭУ, среди которых человек устанавливает приоритетность. Существенное преимущество порогового подхода заключается в том, что он позволяет избегать предположений о возможности замещаемости ЭФ и ЭУ, в отличие от подхода усреднения. Однако этот подход не позволяет отражать значимость каждой отдельной ЭФ/ЭУ, поскольку устанавливается единый для всех ЭФ/ЭУ порог (Gamfeldt, Roger, 2017). Byrnes с соавт. (2014) предложили подход нескольких порогов, который обеспечивает измерение влияния биоразнообразия на несколько функций. Результаты анализа представлены в виде кривых связи между видовым богатством и числом функций на пороге или выше него, который является процентом от максимума наблюдаемых функций. Автором предложено определение трех основных порогов: (а) максимальный порог – верхний предел, при котором увеличение числа видов перестает влиять на мультифункциональность; (б) минимальный порог – нижний предел, при котором увеличение числа видов начинает влиять на мультифункциональность; (в) порог максимального эффекта разнообразия – значение порога, при котором увеличение числа видов обладает наибольшим положительным или отрицательным эффектом на мультифункциональность.
Концептуальные и математические достоинства вышеназванных подходов обсуждались и анализировались в различных работах, но их применимость в естественных экосистемах не исследовалась (Gamfeldt, Roger, 2017). Применение этих подходов требует учета факторов динамики экосистем, к которым, в том числе, относятся изменения климата и управленческие действия, особенно такие, как интенсификация лесопользования.
Большое значение имеют и пространственные масштабы оценки мультифункциональности. Большинство исследований проведено на локальном уровне. Экстраполяция таких локальных наблюдений на более крупные ландшафты могла бы улучшить понимание факторов, влияющих на функционирование экосистемы, и в результате обеспечить понимание их мультифункциональности (Van der Plas et al., 2018). В некоторых случаях оценки требуют небольших пространственных масштабов, но чаще всего управленческие решения принимаются на муниципальном, региональном и федеральном уровнях. В случае необходимости принятия управленческих решений, следует установить соотношение между спросом и предложением ЭУ. В этом случае можно представить мультифункциональность как общую экономическую оценку, но таких оценок для лесных экосистем пока выполнено не было.
Хотя исследования мультифункциональности на уровне ЭФ и ЭУ к настоящему времени продвинулись вперед, не решены очень важные вопросы. Среди них идентификация наиболее информативных индикаторов для каждой группы функций или оценка дифференциации выгод от ЭУ в пространстве (локализация выгод).
Одним из важнейших аспектов является и временная динамика, которая пока не оценивалась для мультифункциональности ЭУ лесов. Учет этого аспекта весьма важен для понимания стабильности и упругости экосистем на протяжении их развития, а также оценки долговременных выгод от ЭУ в целях повышения благосостояния населения. Данные по временным сериям обладают огромным потенциалом для расширения измерений мультифункциональности, поскольку объединяют измерения стабильности и мультифункциональности (Oliver et al., 2015), и позволяют выявить этапы развития экосистем, на которых ЭФ реализуются в максимальной степени.
Решение этих задач возможно только при сближении подходов к оценке связи “биоразнообразие–ЭФ–ЭУ” и одновременного обеспечения множеством ЭФ и ЭУ, т.е. мультифункциональности на обоих уровнях.
БИОРАЗНООБРАЗИЕ И ЭКОСИСТЕМНЫЕ УСЛУГИ ЛЕСОВ
Экосистемы – это комплексы, в которых взаимодействуют биотические и абиотические компоненты. Подобные взаимодействия определяют качество, количество, стабильность предоставления ЭУ и основываются на биоразнообразии во всех его проявлениях. В концепции услуг биоразнообразие может рассматриваться на трех уровнях: (1) основа фундаментальных экосистемных процессов; (2) фактор, который влияет на обеспечение ЭУ – к примеру, генетическое и видовое разнообразие напрямую влияет на некоторые товары и выгоды; (3) выгода – здесь само биоразнообразие является объектом, оцениваемым человеком (Mace et al., 2012).
Большинство исследований взаимосвязи “биоразнообразие–ЭФ–ЭУ” сфокусировано на изучении видового разнообразия. Однако возрастание видового биоразнобразия не всегда приводит к возросшему уровню выполнения функций и предоставления услуг (Hooper et al., 2005). На экосистемные процессы в большей степени влияет функциональное разнообразие (Díaz et al., 2007). Хотя функциональное разнообразие связано с видовым и теоретически может увеличиваться с ним, установлено, что изменение таксономических показателей (видовое богатство, альфа-, бета-, гамма-разнообразие, видовая выровненность) приводит к незначительному влиянию на ЭФ в сравнении с изменением функциональных признаков: функциональное богатство, функциональная выровненность и др. (Mokany et al., 2008). При этом нельзя исключить важность оценки видового разнообразия, которое в рамках концепции мультифункциональности может рассматриваться как буферный механизм функционирования экосистем, обеспечивающий надежность протекания экосистемных процессов в условиях изменения окружающей среды (Hooper et al., 2005). В отечественной геоботанике популярна концепция эколого-ценотических групп, которые могут в определенном смысле соответствовать функциональным группам растений, поскольку формируются по принципу сходства экологических и ценотических условий произрастания (Смирнова и др., 2004).
Практически не изучено влияние генетического и структурного биоразнообразия на ЭФ и ЭУ. Можно предположить, что характер связи генетического и видового разнообразия с ЭФ/ЭУ будет схож, потому что генетическое разнообразие определяет видовое. Структурное биоразнообразие, или аранжировка видов (Одум, 1986) представляет собой распределение элементов экосистемы в среде и его взаимодействие с ней, т.е. отражает мозаичность экосистем, которая является фундаментальным свойством биогеоценотического покрова. Причиной мозаичности покрова может быть влияние как абиотических (рельеф, почвообразующие породы и др.), так и биотических факторов. В настоящее время на лесных территориях наблюдается мозаика различных стадий дигрессионных и демутационных сукцессий, обусловленная действием природных (геоморфологическое положение, состав почвообразующих пород) и антропогенных (промышленное загрязнение, рубки, пожары) факторов. Широко известными базовыми концепциями мозаичности лесных биогеоценозов и их отдельных компонентов являются лесные элементарные почвенные ареалы В.М. Фридланда (1986), лесная парцелла Н.В. Дылиса (1969), ценобиотическая микрогруппировка Л.Г. Раменского (1938), тессера Ханса Йенни (Jenny, 1958), тессера Л.О. Карпачевского (1977). Предложена структурно-функциональная единица лесного биогеоценотического покрова, являющаяся элементарным пространственным провайдером экосистемных функций и услуг, таких как формирование плодородия почв, регулирование биогеохимических циклов – элементарный биогеоареал (Орлова, 2013).
Существуют значительные пробелы в знаниях в отношении комбинированного влияния компонентов биоразнообразия разных трофических уровней на мультифункциональность лесов (Van Der Plas et al., 2018). Недавние исследования в субтропических лесах показали, что функциональное разнообразие растений и разнообразие гетеротрофов, ускоряющих разложение и циклы элементов питания, оказывали более существенное влияние на отдельные экосистемные функции и мультифункциональность, чем видовое разнообразие древесных растений (Schuldt et al., 2018). Несмотря на очевидность взаимосвязей трофических уровней и их влияние на функционирование лесной экосистемы, для бореальных лесов и лесов умеренного пояса такие исследования отсутствуют. В широколиственных и хвойных лесах мощными агентами разложения растительного опада являются микроорганизмы, прежде всего, грибы, и мегасапрофаги – дождевые черви, двупарноногие многоножки, мокрицы, личинки двукрылых, почвообитающие моллюски, личинки жуков-щелкунов и др. Почвенные грибы разлагают органические остатки, преобразуя их в плодородную почву; энтомопатогенные грибы уничтожают многих фитопатогенных беспозвоночных; микоризные грибы положительно влияют на рост растений, активизируя поглощение элементов питания. Кроме того, грибы участвуют в фиксации азота, продуцировании гормонов, стабилизации органического вещества (Treseder, Lennon, 2015). Разнообразие и структура комплекса сапрофагов обеспечивают ряд важнейших ЭФ: определяют направление и динамику разложения подстилки, регулируют биогеохимические циклы и формируют почвенное плодородие (Стриганова, 1980; Yatso, Lilleskov, 2016; и др.). Грибы функционально тесно связаны с дождевыми червями, и они через экосистемные процессы совместно влияют на предоставление ЭУ.
Набор компонентов (гены, виды, признаки и др.) и атрибутов (количество, изменчивость, состав и др.) биоразнообразия, необходимых для успешного предоставления какой-либо конкретной услуги, зависит от степени и направления их влияния на ЭФ. Таким образом, оценка связей в системе “биоразнообразие–ЭФ–ЭУ” является двухступенчатой задачей, в которой необходимо оценить сначала связь “биоразнообразие–ЭФ”, а затем – связь “ЭФ–ЭУ” (Haines-Young, Potschin 2010; Duncan, 2015).
Первый этап оценки находится на более продвинутом уровне. На данной ступени используются общепринятые методы измерения экосистемных процессов. Масштабные международные проекты по оценке влияния биоразнообразия на функционирование экосистем приводят к выводу, что высокое биоразнообразие повышает интенсивность ЭФ. Показано влияние биоразнообразия древесных растений на производительность насаждений, опыление, распространение вредителей и болезней, регулирование пожаров, устойчивость к ветрам, депонирование углерода и др. (Brockerhoff et al., 2017).
О влиянии биоразнообразия на ЭФ выдвинуто несколько десятков теорий, но наиболее распространенными из них являются три. В основе первой – теории “страхования” – лежит принцип компенсации видов (Yachi, Loreau, 1999). Предполагается, что виды частично заменимы и их потеря может быть компенсирована другими видами. Поэтому при большом числе видов ЭФ мало меняются при изъятии/добавлении любого из них, но по мере сокращения числа видов эффект от этого действия становится все более сильным. Следующая теория – “соотношения масс”, согласно которой выполнение ЭФ определяется значением признаков доминирующей по биомассе растительности (Grime, 1998). Третья теория – “ключевых видов”, когда один или несколько видов оказывают существенное влияние на функционирование экосистем независимо от их распространения и биомассы. Выдвинут также принцип оптимального разнообразия. В его основе лежит предположение, что жизнеспособность и эффективность биосистем максимальны при некоторых оптимальных значениях их внутреннего разнообразия, к которым близки характеристики ненарушенных природных систем (Букварева, Алещенко, 2006).
Вторая ступень включает оценку связи “ЭФ–ЭУ”. На этой ступени важно численно понимать, насколько ЭФ близки к ЭУ (выгодам), какие ЭФ обеспечивают создание услуг и каковы взаимоотношения между различными ЭФ. Некоторые ЭФ, главным образом, обеспечивающие, являются одновременно ЭУ, но большинство ЭУ – это синтез множества одновременно действующих ЭФ (“пучок экосистемных функций” (Duncan et al., 2015). Для численной оценки ключевых ЭФ, лежащих в основе конечной ЭУ, предложена концепция “портфелей ЭФ” (Duncan et al., 2015), согласно которой учитывается коэффициент среднего относительного вклада ЭФ и коэффициент ее незаменимости. ЭФ возможно объединять в группы на основе сходства функциональных признаков, механизмов и трофических уровней биоразнообразия. Сравнение структуры портфелей различных ЭУ может быть использовано при оценке компромиссов и синергии между ними (см. ниже). Другим методом оценки связей ЭУ является сетевая теория оценки ЭУ (Dee et al., 2017). ЭУ представлены как социо- экономико-экологическая мета-сеть, в узлах которой расположены субъекты взаимодействия, например, запасы природного капитала в экологических сетях или отдельные организации в социально-экономических сетях, а линии, связывающие узлы, показывают взаимодействия между ними.
В последнее время был опубликован ряд обзоров, посвященных изучению связи “биоразнообразие – ЭУ” (Harrison, 2014; Evers et al., 2018; и др.). Большинством исследователей обнаружена положительная связь между биоразнообразием и экосистемными услугами, но она не всегда носила линейный характер. Например, выявлены нелинейные связи и смешанные варианты, или исследования вообще не выявили связи. К тому же, разные категории ЭУ по-разному реагируют на уровень биоразнообразия (Science…, 2015): высокий уровень биоразнообразия положительно влияет на регулирующие и культурные услуги. Однако высокий уровень обеспечивающих услуг может быть достигнут при низком уровне биоразнообразия. Примером могут служить лесные плантации с высоким запасом древесной биомассы, которые в меньшей степени выполняют функции по сохранению биоразнообразия и регулированию биогеохимических циклов (Тебенькова и др., 2017), чем естественные леса, и которые совершенно не представляют интереса для отдыха.
Несмотря на обилие исследований, открытым остается вопрос индикаторов биоразнообразия для оценки обеспечения ЭУ. Необходима идентификация индикаторов состояния, позволяющих оценивать уровень предоставления ЭУ, и индикаторов эффективности, определяющих уровень устойчивости использования ресурсов/услуг. Ранее были предложены потенциальные индикаторы для определения уровня использования экосистемных услуг (De Groot et al., 2010). Они включают показатели для всех групп ЭУ, определенных в докладе “Оценка экосистем на грани тысячелетий” (Millennium…, 2005). В работе показаны двадцать три ЭУ, представляющие их компоненты или явления, индикаторы состояния, показатель эффективности использования. Например, такая услуга, как обеспечение населения пищевой продукцией леса, характеризуется наличием съедобных растений и животных, объем которых оценивается через средний или общий запас; при этом максимальное изъятие ресурса рассчитывается на основании данных о приросте продукта. В Стратегии сохранения биоразнообразия ЕС до 2020 предложено 115 индикаторов оценки ЭУ леса (European…, 2011). Однако исследователи пришли к выводу, что легко измеримыми являются только индикаторы по обеспечению древесиной и регулированию климата, так как данные для них поступают в результате лесоустроительных и кадастровых работ. Индикаторы для регулирующих и рекреационных услуг имеют наименьшую степень измеримости (Maes et al., 2016). Это свидетельствует о недооценке реальной значимости ЭУ.
Таким образом, несмотря на многочисленные публикации в области ЭУ, выявляется недостаток эмпирических исследований, которые устанавливают численные и причинно-следственные связи между биоразнообразием лесов и множеством ЭУ, т.е. мультифункциональностью лесных экосистем (Brockerhoff et al., 2017).
КОНФЛИКТЫ И СИНЕРГИЯ
Изучение мультифункциональности лесных экосистем неразрывно связано с анализом взаимодействия между различными ЭФ/ЭУ в результате их одновременного выполнения. Эти взаимодействия могут возникнуть из-за одновременной реакции на действие одного и того же фактора (например, управленческая деятельность, изменение климата) или в результате прямого контакта между услугами (Bennett et al., 2009). Существует два основных типа отношений между ЭФ/ЭУ: конфликт и синергизм. Термины “конфликт” и, в меньшей степени, “синергизм” получили наибольшее внимание, несмотря на отсутствие общепринятого определения (Deng, Gibson, 2016). Конфликт между двумя услугами заключается в увеличении предоставления одной услуги в сочетании с одновременным уменьшением предоставления других услуг. Для разрешения конфликта требуется поиск компромиссов. Синергия подразумевает, что увеличение предоставления одной услуги положительно влияет на предоставление другой или не оказывает никакого влияния (Rodríguez et al., 2006; Raudsepp-Hearne et al., 2010). Эти взаимоотношения могут возникать между категориями услуг, в рамках услуг, а также во времени и пространстве (Rodríguez et al., 2006). В своем недавнем обзоре Lee H. и S. Lautenbach (2016) показали, что конфликты преобладают между регулирующими и обеспечивающими ЭУ, в то время как синергизм в основном наблюдается среди регулирующих и культурных ЭУ. Однако авторы также отмечают, что, хотя эти закономерности последовательно наблюдаются в их анализе, их причинность не может быть общепринятой.
Наиболее распространенные конфликты возникают между обеспечением ресурсами (главным образом, древесными) и другими ЭУ леса (Turner et al., 2014). Когда лесоуправление сосредоточено, в первую очередь, на получении выгод от древесной биомассы, снижается уровень предоставления других услуг. Например, увеличение заготовки древесины ведет к сокращению запасов углерода в биомассе и почвах (Frank et al., 2016), снижению способности сохранять биоразнообразие (Kuuluvainen et al., 2012), уменьшению урожайности недревесных продуктов леса (Kurttila et al., 2018). Однако некоторые исследования демонстрируют возможность синергетического отношения между изъятием древесины и отдыхом: так, в некоторых случаях люди предпочитают отдыхать в окультуренных лесопарках, а не в естественных лесах (Gundersen, Frivold, 2008). Существуют незначительные конфликты, а в некоторых случаях и синергизм, между сохранением биоразнообразия и обеспечением многих нерыночных экосистемных услуг (Hooper et al., 2005).
Большое значение имеет пространственный масштаб. При переходе на более высокие административные уровни сглаживается территориальная неоднородность предоставления ЭУ, поэтому некоторые конфликты между ЭУ можно обнаружить только на муниципальном уровне и ниже. К тому же, некоторые конфликты, выявленные на локальном уровне, меняются на синергию на более высоком пространственном масштабе (Roces-Díaz et al., 2018).
Значительному прогрессу способствовала публикация междисциплинарного методологического руководства по количественной оценке взаимоотношений между различными ЭУ (Mouchet et al., 2014). Сочетая экологические (предложение) и социально-экономические (спрос) аспекты ЭУ, было определено три типа взаимодействий внутри ЭУ: взаимодействия между предоставлением ЭУ (“предложение–предложение”), между предложением ЭУ и – запросом общества (“предложение–спрос”) и среди бенефициаров (“спрос–спрос”). Учитывая три основных этапа анализа (1) определение типа взаимоотношений ЭУ, (2) определение пучков ЭУ, (3) определение драйверов ЭУ) был определен широкий спектр численных методов по оценке компромиссов и синергии. На первом этапе анализа для определения пространственных отношений между ЭУ используются графические методы: сравнение карт распределения ЭУ (Anderson et al., 2009), кривые компромисса - для выявления тенденций (Viglizzo, Frank, 2006). Звездные диаграммы - для сравнения относительного предоставления ЭУ (Raudsepp- Hearne et al., 2010). Но ни один из них не дает численного определения силы взаимодействия между ЭУ. Наиболее популярные численные методы выявления взаимоотношений между ЭУ–корреляционный анализ и регрессионные методы (Bennett et al., 2009). Однако этот набор методов не дает динамическую оценку взаимоотношений между ЭУ. На втором этапе анализа однонаправленные в пространстве и времени ЭУ группируются в пучки ЭУ (Raudsepp- Hearne et al., 2010). Для их группировки используется кластерный анализ, в том числе и иерархический. На третьем этапе анализа для определения драйверов пучков ЭУ возможно использовать статистическую модель ANOVA (ANalysis of VAriance), канонические анализы, тесты Mantel и методы на основе расстояния: множественные регрессии на матрицах расстояний, MRM, сравнение между несколькими матрицами расстояний, CADM, анализ аналогии ANOSIM и др. Эмпирический сравнительный анализ трех наиболее встречаемых методов оценки связей ЭУ (статистические пространственные корреляции (англ. static spatial correlations), пространственные корреляции временных вариаций (англ. spatial correlations of temporal variations), границы возможности многомерного анализа (англ. тwo-dimension production possibility frontiers)) показал различия в уровне чувствительности при обнаружении связей между ЭУ. Также для большинства пар услуг методы приводили к различным интерпретациям, либо дополняющим друг друга либо противоречивым (Vallet et al., 2018). Таким образом, численная оценка компромиссов и синергии во многом зависит от выбранного метода. В этой связи встает вопрос разработки и принятия унифицированных методик для возможности сопоставления полученных результатов.
Важно подчеркнуть, что биоразнообразие, функции и услуги изменяются в процессе развития лесов, т.е. в ходе лесообразовательного процесса (Исаев, 2008). Леса представляют собой мозаику разных стадий сукцессий, вызванных комбинированным влиянием природных и антропогенных факторов, включая изменения климата и связанные с ними участившиеся пожары, загрязнение воздуха, лесоуправление. В настоящее время закономерности динамики конфликтов и синергии между ЭФ и ЭУ лесов в ходе их сукцессий не исследованы. Поскольку сукцессии лесов характеризуются значительной длительностью (Мониторинг…, 2008), для их оценки возможно использовать метод хронопоследовательностей, т.е. выстраивания пространственно-временных рядов. С использованием этого подхода демонстрируется аккумуляция углерода в почвах в ходе постпирогенной сукцессии лесов при отсутствии нарушений в период от 50 до 5000 лет на островах в Швеции (Clemmensen et al., 2013). Для прогноза сукцессионных изменений возможно использовать методы математического анализа. А.С. Исаевым с соавт. (2005, 2008) предложена бистабильная феноменологическая модель роста лесных насаждений и выполнения ими продукционной функции, связанной с предоставлением такой ЭУ лесов, как обеспечение древесиной. Она позволяет рассматривать закономерности формирования лесных экосистем на различных этапах лесообразовательного процесса и объяснять экологические эффекты, связанные с динамикой лесного покрова в бореальной зоне. Это достигается построением, структуризацией и анализом фазовых портретов, на которых отражаются динамические процессы возникновения, развития и смены лесных ассоциаций (Мониторинг…, 2008).
Понимание компромиссов и синергии между экосистемными услугами для различных видов лесопользования, их численная оценка, прогноз последствий лесопользования имеют первостепенное значение для интеграции концепции ЭУ в процесс принятия управленческих решений, особенно в свете спроса на предоставление множества ЭУ (Bennett et al., 2009). Без этих знаний существует риск нежелательного отказа от некоторых услуг и потенциальной деградации экосистем. Вместе с целями, поставленными перед лесопользованием, эти знания определяют выбор альтернативных и конкретных управленческих действий (Filyushkina, 2016). Решение этих задач невозможно без использования имитационных моделей. В настоящее время разработан обширный инструментарий математических моделей. Для примера, синергия и компромиссы между обеспечением древесиной и другими различными ЭУ на уровне страны и регионов могут быть оценены с использованием российской модели РОБУЛ (Замолодчиков и др., 2011) и канадской модели CBM-CFS3 (Kurz et al., 2009), на уровне ландшафтов – с применением российских моделей FORRUS-S (Chumachenko et al., 2000) и ILHM, ILLM (Кондратьев, 2007), австрийской iLand (Seidl et al.,, 2012), немецкой 4C (Reyer et al., 2014), шведской Heureka (Wikström, 2011); на уровне отдельных сайтов – на основе интеграции российских моделей EFIMOD (Komarov et al., 2003), ROMUL (Chertov et al., 2001) и BioCalc (Khanina et al., 2007). Однако в настоящее время эти инструменты сфокусированы на оценке отдельных ЭУ, без связи друг с другом. Поэтому актуальной проблемой является создание интегрированной платформы, объединяющей математические модели, оценивающие и прогнозирующие различные ЭФ и ЭУ на разных пространственных уровнях.
Заключение. Обобщая вышесказанное, мультифункциональность лесных экосистем – это явление, в основе которого лежит биоразнообразие во всех его проявлениях (рис. 1). Живые организмы во взаимодействии с окружающей средой формируют экосистемные процессы, которые обеспечивают выполнение лесами ЭФ, лежащих в основе ЭУ. Человек определяет ценность ЭУ, объемы, сроки, место их использования в зависимости от запросов общества, тем самым влияя на биоразнообразие. Таким образом, образуются многочисленные обратные связи между биоразнообразием и ЭУ. В некоторых случаях биоразнообразие напрямую влияет на спрос на ЭУ. К примеру, уровень биоразнообразия определяет рекреационную привлекательность леса и побуждает к принятию тех или иных защитных мер.
Рис. 1.
Концептуальная схема взаимосвязей “биоразнообразие – множество функций – множество услуг” и факторов, влияющих на них.
*В случае рекреации биоразнообразие может напрямую влиять на обеспечение экосистемными услугами, через формирование привлекательности леса.
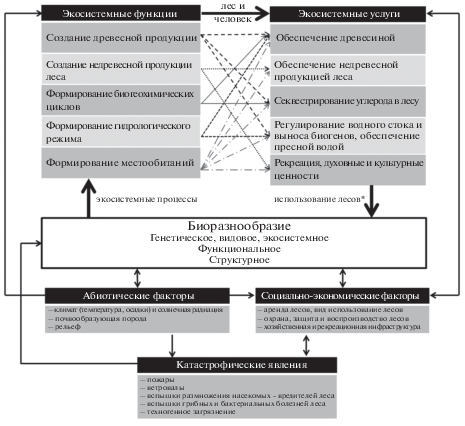
Как упоминалось выше, каждая ЭФ участвует в создании пучка ЭУ, т.е. одна функция участвует в формировании нескольких ЭУ, т.е. одна услуга является результатом действия нескольких ЭФ. На рисунке представлены примеры пучков ЭФ и ЭУ. Сложность оценки мультифункциональности лесных экосистем состоит в одновременном выполнении лесами всех функций и предоставлении услуг, между которыми возникают конфликты и синергия. В связи с этим остро встает вопрос разработки методологических решений, помогающих избегать конфликтов и усиливать синергию между услугами, поскольку невозможно максимизировать все ЭФ/ЭУ одновременно. Такие методологические решения должны предусматривать экологические лимиты потребления ЭУ для удовлетворения потребностей общества в лесных благах. Очевидно, что одним из таких лимитов может стать уровень биоразнообразия экосистем, индикаторы которого еще предстоит обосновать.
Лесные экосистемы формируются под воздействием ряда факторов, как природных, так и антропогенных, влияющих на мультифунцкиональность лесов. К природным абиотическим факторам относятся климат и солнечная радиация, почвообразующая порода, рельеф и др. Под социально-экономическими факторами понимают совокупность человеческой деятельности в лесу, направленной на удовлетворение социальных потребностей и/или получение экономической выгоды. К этой группе факторов относится строительство инфраструктуры, мероприятия по охране, защите и воспроизводству лесов, вид использования лесов и др. Последняя группа факторов объединяет катастрофические события, возникшие в результате природной или техногенной чрезвычайной ситуации и повлекшие за собой гибель лесной экосистемы или ущерб в значительных размерах. Например, ежегодно в России происходят десятки тысяч лесных пожаров, уничтожающих миллионы гектаров леса. Ослабленные после воздействия огня деревья активно заселяются насекомыми-ксилофагами, что приводит к массовому усыханию древостоев (Isaev et al., 2017).
Таким образом, концепция мультифункциональности лесных экосистем находится в процессе становления в качестве нового направления экологических исследований, основными вызовами которой на сегодняшний день являются: (1) разграничение понятий “функции” и “услуги” лесов; (2) поиск связей между биоразнообразием и мультифункциональностью, их причинно-следственный анализ и разработка единого подхода к оценкам этих связей; (3) разработка методических основ оценки компромиссов и синергии между различными функциями и услугами; (4) обоснование наиболее информативных индикаторов биоразнообразия для оценки устойчивости обеспечения ЭУ и лимитов потребления ЭУ; (5) разработка интегрированных платформ математических моделей, оценивающих и прогнозирующих динамику различных экосистемных функций и услуг на разных пространственных уровнях; (6) разработка систем поддержки принятия решений, направленных на соблюдение баланса синергии и компромиссов между различными ЭУ. Решение этих вопросов позволит не только получить новые фундаментальные знания о процессах функционирования лесных экосистем, но и создать рынки ранее не оцениваемых экосистемных услуг, а также обеспечить непрерывное и устойчивое лесопользование.
Список литературы
Букварева Е.Н., Алещенко Г.М. Принцип оптимального разнообразия биосистем и стратегия управления биоресурсами // Государственное управление в XXI веке: традиции и инновации. М.:РОССПЭН, 2006. С. 204–210.
Дылис Н.В. Структура лесного биогеоценоза // Комаровские чтения. М.: Наука, 1969. Т. 21. 28 с.
Исаев А.С., Суховольский В.Г., Овчинникова Т.М. Феноменологические модели роста лесных насаждений // Журн. общей биологии. 2008. Т. 69. № 1. С. 3–9.
Исаев А.С., Суховольский В.Г., Бузыкин А.И., Овчинникова Т.М. Сукцессионные процессы в лесных сообществах: модели фазовых переходов // Хвойные леса бореальной зоны. 2008. Т. 25. № 1–2. С. 9–15
Исаев А.С., Суховольский В.Г., Хлебопрос Р.Г., Бузыкин А.И., Овчинникова Т.М. Моделирование лесообразовательного процесса: феноменологический подход // Лесоведение. 2005. № 1. С. 3–11.
Замолодчиков Д.Г., Грабовский В.И., Краев Г.Н. Динамика бюджета углерода лесов России за два последних десятилетия // Лесоведение. 2011. № 6. С. 16–28.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Московского Университета, 1977. 312 с.
Кондратьев С.А. Формирование внешней нагрузки на водоемы: проблемы моделирования. СПб.: Наука, 2007. 255 с.
Мониторинг биологического разнообразия лесов России: методология и методы/ Под ред. А.С. Исаева. М.: Наука, 2008. 453 с.
Национальная Стратегия сохранения биоразнообразия России. М.: Министерство природных ресурсов, 2001. 76 с.
Одум Ю. Экология. М.: Рипол Классик, 1986. 740 с.
Орлова М.А. Элементарная единица лесного биогеоценотического покрова для оценки экосистемных функций лесов // Труды Карельского НЦ РАН 2013. № 6. С. 126–132.
Основы государственной политики в области обеспечения химической и биологической безопасности Российской Федерации на период до 2025 года и дальнейшую перспективу. Утверждено Указом Президента Российской Федерации от 11 марта 2019 г. № 97.
Раменский Л.Г. Введение в комплексное почвенно-геоботаническое исследование земель. Л.: Сельхозгиз, 1938. 620 с.
Российский статистический ежегодник. 2018: Стат.сб./Росстат. – Р76. М., 2018. 694 с.
Смирнова О.В., Ханина Л.Г., Смирнов В.Э. Эколого-ценотические группы в растительном покрове лесного пояса Восточной Европы // Восточно-Европейские леса (история в голоцене и современность) М.: Наука, 2004. Кн. 1. С. 165–175.
Стратегия развития лесного комплекса Российской Федерации до 2030 года. Утверждено распоряжением Председателя Правительства Российской Федерации 20 сентября 2018 г. № 1989-р // Дата обновления: 24.09.2018. Доступ из системы ГАРАНТ.
Стриганова Б.Р. Питание почвенных сапрофагов. М.: Наука, 1980. 243 с.
Тебенькова Д.Н., Лукина Н.В. Фомич А.Ф., Орлова М.А., Гагарин Ю.Н. Почвы лесов и лесных культур: сравнение кислотности и плодородия // Бореальные леса: состояние, динамика, экосистемные услуги: Тезисы докладов Всерос. науч. конф. с междунар. участием, посвященной 60-летию Института леса Карельского НЦ РАН (Петрозаводск, 11–15 сентября 2017 г.). Петрозаводск: Изд-во Карельского НЦ РАН, 2017. С. 292.
Фридланд В.М. Проблемы географии генезиса и классификации почв. М.: Наука, 1986. 243 с.
Anderson B.J. Armsworth P.R., Eigenbrod F., Thomas C.D., Gillings S., Heinemeyer A., Roy D.B., Gaston K.J. Spatial covariance between biodiversity and other ecosystem service priorities // J. Applied Ecology. 2009. V. 46. № 4. P. 888–896.
Ansink E., Hein L., Hasund K.P. To value functions or services? An analysis of ecosystem valuation approaches // Environmental Values. 2008. V. 17. № 4. P. 489–503.
Bennett E.M., Peterson G.D., Gordon L.J. Understanding relationships among multiple ecosystem services // Ecology letters. 2009. V. 12. № 12. P. 1394–1404.
Brockerhoff E.G., Barbaro L., Castagneyrol B., Forrester D.I., Gardiner B., González-Olabarria J.R., Lyver P.O., Meurisse N., Oxbrough A., Taki H., Thompson I.D., van der Plas F., Jactel H. Forest biodiversity, ecosystem functioning and the provision of ecosystem services // Biodiversity and Conservation. 2017. V. 26. P. 3005–3035.
Byrnes J.E., Gamfeldt L., Isbell F., Lefcheck J.S., Griffin J.N., Hector A. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions // Methods in Ecology & Evolution. 2014. V. 5. № 2. P. 111–124.
Chertov O.G., Komarov A.S., Nadporozhskaya M.A., Bykhovets S.S., Zudin S.L. ROMUL – a model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modelling // Ecological Modelling. 2001. V. 138. P. 289–308.
Chumachenko S.I., Syssouev V.V., Palyonova M.M., Bredikhin M.A., Korotkov V.N. Simulation of heterogeneous uneven-aged stands dynamics under human-induced disturbance // USDA. Forest Service. NC General Technical Report. St. Paul. 2000. № 209. P. 104–112.
Clemmensen K.E., Bahr A., Ovaskainen O., Dahlberg A., E-kblad A., Wallander H., Lindahl B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest // Science. 2013. V. 339. № 6127. P. 1615–1618.
Dee L.E., Allesina S., Bonn A., Eklöf A., Gaines S.D., Hines J., Jacob U., McDonald-Madden E., Possingham H., Schröter M., Thompson R.M. Operationalizing network theory for ecosystem service assessments // Trends in Ecology & Evolution. 2017. V. 32. № 2. P. 118–130.
De Groot R.S., Alkemade R., Braat L., Hein L., Willemen L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making // Ecological Complexity. 2010. V. 7. № 3. P. 260–272
Deng X., Li Z., Gibson J. A review on trade-off analysis of ecosystem services for sustainable land-use management // J. Geographical Sciences. 2016. V. 26. № 7. P. 953–968.
Díaz S., Lavorel S., de Bello F., Quétier F., Grigulis K., Robson T.M. Incorporating plant functional diversity effects in ecosystem service assessments // Proceedings of the National Academy of Sciences. 2007. V. 104. № 52. P. 20 684–20 689.
Duncan C., Thompson J.R., Pettorelli N. The quest for a mechanistic understanding of biodiversity–ecosystem services relationships // Proceedings of the Royal Society B: Biological Sciences. 2015. V. 282. № 1817. P. 201 51 348.
European Commission. Our life insurance, Our Natural Capital: An EU Biodiversity Strategy to 2020. European Commission: Brussels. 2011. P. 16
Evers C.R. Wardropper C.B., Branoff B., Granek E.F., Hirsch S.L., Link T.E., Olivero-Lora S., Wilson C. The ecosystem services and biodiversity of novel ecosystems: A literature review // Global Ecology & Conservation. 2018. V. 13. P. e00362.
Filyushkina A. Ecosystem services and forest Management in the nordic countries. 2016. V. 2016. № 53. P. 1652–6880.
Frank S., Böttcher H., Gusti M., Havlík P., Klaassen G., Kindermann G., Obersteiner M. Dynamics of the land use, land use change, and forestry sink in the European Union: the impacts of energy and climate targets for 2030 // Climatic Change. 2016. V. 138. № 1–2. P. 253–266.
Gamfeldt L., Roger F. Revisiting the biodiversity–ecosystem multifunctionality relationship // Nature Ecology & Evolution. 2017. V. 1. № 7. P. 0168.
Gamfeldt L., Hillebrand H., Jonsson P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning // Ecology. 2008. V. 89. № 5. P. 1223–1231.
Gómez-Baggethun E., De Groot R., Lomas P.L., Montes C. The history of ecosystem services in economic theory and practice: from early notions to markets and payment schemes // Ecological Economics. 2010. V. 69. № 6. P. 1209–1218.
Grime J.P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects // Journal of Ecology. 1998. V. 86. № 6. P. 902–910.
Gundersen V.S., Frivold L.H. Public preferences for forest structures: a review of quantitative surveys from Finland, Norway and Sweden // Urban Forestry & Urban Greening. 2008. V. 7. № 4. P. 241–258.
Haines-Young R., Potschin M. The links between biodiversity, ecosystem services and human well-being // Ecosystem Ecology: a new synthesis. 2010. V. 1. P. 110–139.
Harrison P.A., Berry P.M., Simpson G., Haslett J.R., Blicharska M., Bucur M., Dunford R., Egoh B., Garcia-Llorente M., Geamănă N., Geertsema W., Lommelen E., Meiresonne L., Turkelboom F. Linkages between biodiversity attributes and ecosystem services: a systematic review // Ecosystem Services. 2014. V. 9. P. 191–203.
Hautier Y., Isbell F., Borer E.T., Seabloom E.W., Harpole W.S., Lind E.M., MacDougall A.S., Stevens C.J., Adler P.B., Alberti J., Bakker J.D., Brudvig L.A., Buckley Y.M., Cadotte M., Caldeira M.C., Chaneton E.J., Chu C., Daleo P., Dickman C.R., Dwyer J.M., Eskelinen A., Fay P.A., Firn J., Hagenah N., Hillebrand H., Iribarne O., Kirkman K.P., Knops J.M.H., La Pierre K.J., McCulley R.L., Morgan J.W., Pärtel M., Pascual J., Price J.N., Prober S.M., Risch A.C., Sankaran M., Schuetz M., Standish R.J., Virtanen R., Wardle G.M., Yahdjian L. Hector A. Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality // Nature ecology & evolution. 2018. V. 2. № 1. P. 50–56.
Hector A., Bagchi R. Biodiversity and ecosystem multifunctionality // Nature. 2007. V. 448. № 7150. P. 188–190.
Hooper D.U., Chapin F.S., Ewel J.J., Hector A., Inchausti P., Lavorel S., Lawton J.H., Lodge D.M., Loreau M., Naeem S., Schmid B., Setälä H., Symstad A.J., Vandermeer J., Wardle D.A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge // Ecological Monographs. 2005. V. 75. № 1. P. 3–35.
Isaev A.S., Sukhovolsky V.G., Tarasova O.V., Palnikova E.N., Kovalev A.V. Forest insect population dynamics, outbreaks and global warming effects. John Wiley & Sons. 2017. P. 304.
Jenny H. Role of the plant factor in the pedogenic functions // Ecology. 1958. V. 39. № 1. P. 5–16.
Khanina L.G., Bobrovsky M.V., Komarov A.S., Mikhajlov A.V. Modeling dynamics of forest ground vegetation diversity under different forest management regimes // Forest Ecology & Management. 2007. V. 248. P. 80–94.
Komarov A.S., Chertov O.G., Zudin S.L., Nadporozhskaya M.A., Mikhailov A.V., Bykhovets S.S., Zudina E.V., Zoubkova E.V. EFIMOD 2 – A model of growth and elements cycling of boreal forest ecosystems // Ecological Modelling. 2003. V. 170. P. 373–392.
Kurttila M., Pukkala T., Miina J. Synergies and trade-offs in the production of NWFPs predicted in boreal forests // Forests. 2018. V. 9. № 7. P. 400–417.
Kurz W.A., Dymond C.C., White T.M., Stinson G., Shaw C.H., Rampley G.J., Smyth C.E., Simpson B.N., Neilson E.T., Trofymow J.A., Metsaranta J.M., Apps M.J. CBM-CFS3: a model of carbon-dynamics in forestry and land-use change implementing IPCC standards // Ecological Modelling. 2009. V. 220. № 4. P. 480–504.
Kuuluvainen T., Tahvonen O., Aakala T. Even-aged and uneven-aged forest management in boreal Fennoscandia: a review // Ambio. 2012. V. 41. № 7. P. 720–737.
Lee H., Lautenbach S. A quantitative review of relationships between ecosystem services // Ecological Indicators. 2016. V. 66. P. 340–351.
Louman B., Cifuentes M., Chacón M. REDD+, RFM, development, and carbon markets // Forests. 2011. V. 2. № 1. P. 357–372.
Mace G.M., Norris K., Fitter A.H. Biodiversity and ecosystem services: a multilayered relationship // Trends in Ecology & Evolution. 2012. V. 27. № 1. P. 19–26.
Maes J., Liquete C., Teller A., Erhard M., Paracchini M.L., Barredo J.I., Meiner A. An indicator framework for assessing ecosystem services in support of the EU Biodiversity Strategy to 2020 // Ecosystem Services. 2016. V. 17. P. 14–23.
Maestre F.T., Quero J.L., Gotelli N.J., Escudero A., Ochoa V., Delgado-Baquerizo M., García-Palacios P. Plant species richness and ecosystem multifunctionality in global drylands // Science. 2012. V. 335. № 6065. P. 214–218.
Manning P., Plas F., Soliveres S., Allan E., Maestre F.T., Mace G., Whittingham M.J., Fischer M. Redefining ecosystem multifunctionality // Nature Ecology & Evolution. 2018. V. 2. № 3. P. 427–436.
Manning P., Taylor G., Hanley M.E. Bioenergy, food production and biodiversity–an unlikely alliance? // Global Change Biology Bioenergy. 2015. V. 7. № 4. P. 570–576.
Maskell L.C., Crowe A., Dunbar M.J., Emmett B., Henrys P., Keith A.M., Norton L.R., Scholefield P., Clark D.B., Simpson I.C., Smart S.M. Exploring the ecological constraints to multiple ecosystem service delivery and biodiversity // J. Applied Ecology. 2013. V. 50. № 3. P. 561–571.
McGuire A.D., Sitch S., Clein J.S., Dargaville R., Esser G., Foley J., Heimann M., Joos F., Kaplan J., Kicklighter D.W., Meier R.A., Melillo J.M., Moore B., Prentice I.C., Ramankutty N., Reichenau T., Schloss A., Tian H., Williams L.J., Wittenberg U. Carbon balance of the terrestrial biosphere in the twentieth century: Analyses of CO2, climate and land use effects with four process-based ecosystem models // Global Biogeochemical Cycles. 2001. V. 15. № 1. P. 183–206.
Millennium Ecosystem Assessment. Ecosystems and Human Wellbeing: Synthesis. USA, Washington: Island Press, 2005. (URL: http://www.millenniumassessment.org/en/Reports. aspx#)
Mokany K., Ash J., Roxburgh S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland // Journal of Ecology. 2008. V. 96. № 5. P. 884–893.
Mouchet M., Paracchini M.L., Schulp C.J.E., Stürck J., Verkerk P.J., Verburg P.H., Lavorel S. Bundles of ecosystem (dis) services and multifunctionality across European landscapes // Ecological Indicators. 2017. V. 73. P. 23–28.
Mouchet M.A., Lamarque P., Martín-López B., Crouzat E., Gos P., Byczek C., Lavorel S. An interdisciplinary methodological guide for quantifying associations between ecosystem services // Global Environmental Change. 2014. V. 28. P. 298–308.
Oliver T.H., Heard M.S., Isaac N.J., Roy D.B., Procter D., Eigenbrod F., Proença V. Biodiversity and resilience of ecosystem functions // Trends in Ecology & Evolution. 2015. V. 30. № 11. P. 673–684.
Puydarieux P., Beyou W. Evaluation française des écosystèmes et des services écosystémique–cadre conceptuel. Ministère de l’Environnement & Fondation pour la Recherche sur la Biodiversité. 2017. URL: https://www. ecologique-solidaire.gouv.fr/sites/default/files/Thema%20-% 20Efese%20-%20Le%20cadre%20conceptuel.pdf (дата обращения 3.12.2018)
Ratcliffe S., Liebergesell M., Ruiz-Benito P., Madrigal González J., Muñoz Castañeda J.M., Kändler G., Lehtonen A., Dahlgren J., Kattge J., Peñuelas M.J., Zavala A., Wirth C. Modes of functional biodiversity control on tree productivity across the European continent // Global Ecology and Biogeography. 2016. V. 25. № 3. P. 251–262.
Raudsepp-Hearne C., Peterson G.D., Bennett E.M. Ecosystem service bundles for analyzing tradeoffs in diverse landscapes // Proceedings of the National Academy of Sciences. 2010. V. 107. № 11. P. 5242–5247.
Reyer C., Lasch-Born P., Suckow F., Gutsch M., Murawski A., Pilz T. Projections of regional changes in forest net primary productivity for different tree species in Europe driven by climate change and carbon dioxide // Annals of forest science. 2014. V. 71. № 2. P. 211–225.
Roces-Díaz J.V., Vayreda J., Banqué-Casanovas M., Díaz-Varela E., Bonet J.A., Brotons L., de-Miguel S., Herrando S., Martínez-Vilalta J. The spatial level of analysis affects the patterns of forest ecosystem services supply and their relationships // Science of The Total Environment. 2018. V. 626. P. 1270–1283.
Rodríguez J.P. Beard Jr.T.D., Bennett E.M., Cumming G.S., Cork S.J., Agard J., Dobson A.P., Peterson G.D. Trade-offs across space, time, and ecosystem services // Ecology and Society. 2006. V. 11. № 1. P. 28.
Schuldt A., Assmann T., Brezzi M., Buscot F., Eichenberg D., Gutknecht J., Härdtle W., He J.-S., Klein A.-M., Kühn P., Liu X., Ma K., Niklaus A.P., Pietsch K.A., Purahong W., Scherer-Lorenzen M., Schmid B., Scholten Th., Staab M., Tang Z., Trogisch S., von Oheimb G., Wirth C., Wubet T., Zhu C.-D., Bruelheide H. Biodiversity across trophic levels drives multifunctionality in highly diverse forests // Nature Communications. 2018. V. 9. № 1. P. 2989–2999.
Science for Environment Policy. Ecosystem Services and Biodiversity. In-depth Report 11 produced for the European Commission. DG Environment by the Science Communication Unit. UWE: Bristol, 2015. P. 32.
Seidl R. Rammer W., Scheller R.M., Spies T.A. An individual-based process model to simulate landscape-scale forest ecosystem dynamics // Ecological Modelling. 2012. V. 231. P. 87–100.
Soliveres S., Van Der Plas F., Manning P., Prati D., Gossner M.M., Renner S.C., Birkhofer K. Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality // Nature. 2016. V. 536. № 7617. P. 456–459.
The Economics of Ecosystems and Biodiversity: Mainstreaming the Economics of Nature. A synthesis of the approach, conclusions and recommendations of TEEB. Malta: Progress Press. 2010. 49 p.
The International Union for Conservation of Nature’s Red List of Threatened Species. URL: https://www.iucnredlist. org/ (дата обращения 03.12.2018).
Treseder K.K., Lennon J.T. Fungal traits that drive ecosystem dynamics on land // Microbiology and Molecular Biology Reviews. 2015. V. 79. P. 243–262.
Turner K.G. Odgaard M.V., Bocher P.K., Dalgaard T., Svenning J.C. Bundling ecosystem services in Denmark: Trade-offs and synergies in a cultural landscape // Landscape and Urban Planning. 2014. V. 125. P. 89–104.
Vallet A. Locatelli B., Levrel H., Wunder S., Seppelt R., Scholes R.J., Oszwald J. Relationships between ecosystem services: Comparing methods for assessing tradeoffs and synergies // Ecological Economics. 2018. V. 150. P. 96–106.
Van der Plas F., Ratcliffe S., Ruiz-Benito P., Scherer-Lorenzen M., Verheyen K., Wirth C., Zavala M.A., Ampoorter E., Baeten L., Barbaro L., Bastias C., Bauhus J., Benavides R., Benneter A., Bonal D., Bouriaud O., Bruelheide H., Bussotti F., Carnol M., Castagneyrol B., Charbonnier Y., Cornelissen J.H.C., Dahlgren J., Checko E., Coppi A. Dawud S.M., Deconchat M., Smedt P., Wandeler H., Domisch T., Finér L., Fotelli M., Gessler A., Granier A., Grossiord C., Guyot V., Haase J., Hättenschwiler St., Jactel H., Jaroszewicz B., Joly Fs-X., Jucker T., Kambach S., Kaendler G., Kattge J., Koricheva J., Kunstler G., Lehtonen A., Liebergesell M., Manning P., Milligan H., Müller S., Muys B., Nguyen D., Nock C., Ohse B., Paquette A., Peñuelas J., Pollastrini M., Radoglou K., Raulund R., Fabian R., Rupert S., Federico S., Jan S., Fernando V., Karsten K., Keer J., Vesterdal L., Fischer M., Gamfeldt L., Allan E. Continental mapping of forest ecosystem functions reveals a high but unrealised potential for forest multifunctionality // Ecology Letters. 2018. V. 21. № 1. P. 31–42.
Viglizzo E.F., Frank F.C. Land-use options for Del Plata Basin in South America: Tradeoffs analysis based on ecosystem service provision // Ecological Economics. 2006. V. 57. № 1. P. 140–151.
Wikström P., Edenius L., Elfving B., Eriksson L.O., Lämås T., Sonesson J., Karin Ö., Wallerman J., Waller C., Klintebäck F. The Heureka forestry decision support system: an overview. // International J. Mathematical and Computational Forestry & Natural-Resource Sciences. 2011. V. 3. № 2. P. 87–95.
Yachi S., Loreau M. Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis // Proceedings of the National Academy of Sciences. 1999. V. 96. № 4. P. 1463–1468.
Yatso K.N., Lilleskov E.A. Effects of tree leaf litter, deer fecal pellets, and soil properties on growth of an introduced earthworm (Lumbricus terrestris): implications for invasion dynamics // Soil Biology & Biochemistry. 2016. V. 94. P. 181–190.
Дополнительные материалы отсутствуют.