

Лесоведение, 2019, № 6, стр. 556-566
Сопротивление движению влаги в проводящей системе сосны обыкновенной
Т. А. Сазонова a, *, В. К. Болондинский a, В. Б. Придача a, *
a Институт леса КарНЦ РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: sazonova@krc.karelia.ru
* E-mail: sazonova@krc.karelia.ru
Поступила в редакцию 13.12.2017
После доработки 10.08.2018
Принята к публикации 05.06.2019
Аннотация
Вода является ведущим фактором среды, лимитирующим продуктивность растений. Водоснабжение растения обусловлено гидравлической проводимостью всех его тканей и органов от корней до листьев. Для выявления количественной оценки сопротивлений движению влаги в разных звеньях водопроводящей системы древесного растения нами проведен анализ данных многолетних наблюдений скорости водного потока по ксилеме ствола, абсолютной влажности ксилемы, водных потенциалов корней и охвоенных побегов сосны обыкновенной (Pinus sylvestris L.) в условиях сосняка черничного свежего европейской части средней тайги (Южная Карелия). На основе полученных данных рассчитаны сопротивления движению влаги в звеньях “почва – корень” и “корень – побег”, равные 2.2 × 106 и 5.5 × 106 МПа с м–3 соответственно. Установлен диапазон изменчивости удельного сопротивления в звене “корень – побег” в течение вегетации, который составил 0.39 × 104… 0.84 × 104 МПа с м–2 при изменении абсолютной влажности ксилемы от ее максимальных до минимальных значений.
Проблема устойчивости гидрологического цикла в лесных экосистемах является актуальной в контексте проблемы глобального изменения климата (Katul et al., 2012; Olchev et al., 2013; Fatichi et al., 2015). Отклик растений на эти изменения может проявляться, в частности, через изменение транспирации, основная регуляция которой происходит посредством изменения устьичной проводимости (Berry et al., 2010). При этом изменения в устьичной проводимости и транспирации тесно связаны с изменением общей гидравлической проводимости в системе “почва – лист” (Cruiziat et al., 2002; Brodribb, Cochard, 2009; Sperry et al., 2016).
Обычно для расчета проводимости или ее обратной величины – сопротивления движению влаги в разных звеньях системы используют аналог закона Ома. Известно, что из всей воды, проходящей через растение, только 1% идет на синтез растительной массы, часть воды тратится на возмещение испарившейся и поддержание тургора и флоэмного транспорта, остальная вода расходуется в процессе транспирации (Kramer, Boyer, 1995). Учитывая незначительность расхода воды на биохимические превращения и испарение с поверхности ствола и ветвей, считают, что практически вся влага, поглощенная за вегетационный период корнями, проходя через водопроводящую систему дерева, испаряется в атмосферу. Данное условие сохранения массы потока воды обеспечивает возможность применения аналога закона Ома для описания движения водного потока в системе “почва–растение–атмосфера” (ПРА). Согласно термодинамическим представлениям градиенты водного потенциала как показатели различных энергетических состояний создают движущую силу для перемещения воды по системе ПРА. Интенсивность перемещения (или скорость водного потока) прямо пропорциональна градиенту водного потенциала и обратно пропорциональна сопротивлению в каждом звене системы.
Идея использования аналога с электрическим током (закон Ома) для описания движения воды по дереву или в системе ПРА достаточно стара (Huber, 1924; Van den Honert, 1948), и ее развитие приходится на вторую половину прошлого века. При этом некоторые исследователи указывали на необходимость учета способности отдельных частей дерева выступать в роли источника или, напротив, резервуара влаги, а также принимать во внимание однородность и стационарность потока воды во всех звеньях системы (Jarvis, 1975; Hinckley et al., 1978). Однако эти замечания справедливы при исследованиях в короткие интервалы времени, когда расход и приход влаги в ксилему не сбалансированы. В длительный интервал времени, например, вегетационный период, когда поглощение влаги из почвы и транспирационный расход практически равны, аналог закона Ома справедлив для потока на участке ствола (Кайбияйнен, 2003). В формулировке Т.Н. Van den Honert (1948) аналогия с законом Ома для разных звеньев транспортной цепи “почва (п)–корень (к), корень (к)–хвоя (хв), хвоя (хв)–атмосфера (атм)”, представлена как
(1)
${{U}_{{{\text{ПРА}}}}} = \frac{{\Delta {{\Psi }_{{{\text{п}} - {\text{к}}}}}}}{{{{R}_{{{\text{п}} - {\text{к}}}}}}} = \frac{{\Delta {{\Psi }_{{{\text{к}} - {\text{хв}}}}}}}{{{{R}_{{{\text{к}} - {\text{хв}}}}}}} = \frac{{\Delta {{\Psi }_{{{\text{хв}} - {\text{атм}}}}}}}{{{{R}_{{{\text{хв}} - {\text{атм}}}}}}},$Используя уравнение (1), можно рассчитать сопротивление (R) движению воды как во всей системе, так и в разных ее звеньях. Однако определение R в каждом из звеньев является непростой задачей. Основная трудность связана, прежде всего, с определением скорости водных потоков в разных частях системы ПРА. Особую сложность представляет определение гидравлического сопротивления (или проводимости) ризосферы. Эта величина существенным образом зависит от механического состава почвы, наличия в ней влаги. Насыщенная влагой почва определенного механического состава имеет постоянную величину проводимости, но в процессе понижения влажности проводимость может уменьшиться на несколько порядков (Hinckley et al., 1978; Whitehead, Jarvis, 1981). Поэтому обычно определяют Rп–к звена “почва–корень”. Однако и на эту величину оказывают влияние многие факторы, в частности сопротивление почвы, частичная потеря контакта между почвой и корнем во время засухи (North, Nobel, 1991; Irvine et al., 1998), обеспеченность почвы макроэлементами (Trubat et al., 2006), архитектура и возраст корней (Draye et al., 2010; Carminati, 2013), микоризные отношения (Nardini et al., 2000; Smith et al., 2010), что приводит к существенным различиям в величинах Rп–к.
В работах по исследованию гидравлического сопротивления растений можно выделить два подхода. В первом случае проводимость ствола, ветвей и корней измеряют на отрезках древесины путем деаэрации прокачиваемой воды и полного насыщения образца (Whitehead, Jarvis, 1981; Sellin, 1993, 2001). Одним из существенных недостатков данного метода являются завышенные значения характеристик водообмена, полученных в лабораторных условиях, поскольку в естественной среде у древесных растений полное насыщение ксилемы ствола наблюдается редко. Во втором случае исследования проводят в естественных условиях среды на ненарушенных растениях (Granier et al., 1989; Lu et al., 1996; Irvine et al., 1998). Также отмечают значительную видовую изменчивость гидравлического сопротивления ксилемы ствола разных древесных растений (Whitehead, Jarvis, 1981; Tyree, Ewers, 1991; Cruiziat et al., 2002; Martínez-Vilalta et al., 2004; Sack, Holbrook, 2006; McCulloh et al., 2011; Bouche et al., 2014). Вместе с тем, для установления закономерностей транспирационного расхода воды отдельным деревом и древостоем в целом в ответ на влияние средообразующих факторов необходима оценка величины сопротивлений движению влаги в разных частях системы “почва–лист” и анализ причин ее временно́й динамики.
Цель нашей работы – провести оценку сопротивлений движению влаги в звеньях “почва–корень” (Rп–к) и “корень–охвоенный побег” (Rк–поб) у сосны обыкновенной (Pinus sylvestris L.), а также пределов изменчивости Rк–поб в течение вегетации для сосняка черничного свежего.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в сосняке черничном свежем европейской части средней тайги (Южная Карелия, 61°13′ с. ш., 34°10′ в. д.). Состав древостоя 9С1Б, где С – сосна, Б – береза, II–III класс бонитета – возник после беглого пожара. Средний возраст – 60 лет, диаметр – 18.0 см, высота деревьев – 15.9 м. Почва – песчаный иллювиально-гумусово-железистый подзол. Основная масса корней сосредоточена в почвенной толще 2–20 см. Грунтовые воды залегают на глубине 1.5–1.7 м (Сазонова и др., 2011).
Объектами исследования послужили деревья сосны обыкновенной (Pinus sylvestris L.) 50–70–летнего возраста. Для изучения показателей водного режима в течение ряда вегетационных периодов (май–сентябрь), различающихся погодными условиями (рис. 1), определяли водные потенциалы охвоенных побегов (Ψпоб, МПа) и скорости водного потока по ксилеме ствола (U, г см–2 ч–1) в суточной динамике и путем однократных замеров в разные фенофазы развития сосны. Для изучения суточной динамики измерения проводили через 0.5–2 ч в течение суток по 5–10 раз в мес. Следует отметить, что сравнительный анализ климатических норм средней годовой температуры воздуха в Карелии за два стандартных климатических периода XX в. (1931–1960 и 1961–1990 г.) показал незначимое (0.1–0.2°С) изменение климатических норм, в то время как для периода с 1991 по 2011 г. установлено превышение средних многолетних значений годовой температуры воздуха на 0.9–1.2°С (Назарова, 2014). Вместе с тем, сопоставление многолетних данных по режиму осадков на территории Карелии в течение второй половины ХХ и начале ХХI в. также выявило рост сумм атмосферных осадков во все сезоны года (Назарова, 2015); превышение средних многолетних значений климатической нормы за 1991–2013 гг. составило 20–70 мм.
Рис. 1.
Динамика среднемесячных многолетних величин атмосферных осадков (а) и температуры воздуха (б) в сосняке черничном свежем.
Годы исследования: 1 – 1977; 2 – 1978; 3 – 1979; 4 – 1992; 5 – 2011; 6 – 2012; 7 – 2013.
Для определения Ψпоб с каждого дерева отбирали по 3 побега со средней части кроны; наблюдения проводили для 5 деревьев. Водные потенциалы корней (Ψк, МПа) определяли в ночное и дневное время суток по 3–4 раза в месяц. Измерения Ψ охвоенных побегов и корней проводили с помощью камеры давления (Сазонова и др., 2011) с точностью определения 0.05 МПа, в исследованиях последних лет – камеры давления Plant Moisture Vessel SKPM 1400 (Skye Instruments Ltd., Великобритания) с точностью определения 0.01 МПа. Сравнение результатов определения Ψ с помощью приборов старого и нового поколения показало хорошую сходимость. Водные потенциалы почвы (Ψп, МПа) измеряли с помощью тензиометра (Соловьев, 1971). Микрометеорологические наблюдения проводили по стандартным методикам (Романов, 1961).
Датчики для измерения U устанавливали на 3 деревьях. Термопары внедряли на глубину 1.5 см, нагреватель – 3 см от поверхности ствола, при этом верхняя и нижняя термопары находились на расстоянии соответственно 1 и 0.5 см от нагревателя. Для измерения скорости водного потока использовали тепловой импульсный метод (Marshall, 1958; Swanson, 1972), согласно которому количество воды, протекающее через единицу поперечного сечения ксилемы в единицу времени (U), определяется из соотношения:
где U – объемная скорость ксилемного водного потока, г см–2 ч–1; γдр – объемная плотность древесины в абсолютно сухом состоянии, г см–3; Wa – абсолютная влажность древесины в месте измерения, %; V – линейная скорость водного потока, см ч–1.
В свою очередь V рассчитывали по формуле:
где x1 и x2 – расстояния от места внедрения в ствол дерева верхней и нижней термопар до нагревателя; t0 – время, через которое устанавливается равенство температур верхней и нижней термопар после подачи теплового импульса.Для определения абсолютной влажности древесины использовали модифицированный Л.К. Кайбияйненом с соавт. (1981) тепловой импульсный метод. Было показано, что между временем достижения максимальной разности температур между точками x1 и x2 (tmax), линейной скоростью водного потока (V) и температуропроводностью среды (а), входящей в решение уравнения теплопроводности, существует функциональная связь, аппроксимируемая соотношением:
Далее функциональная зависимость между а и Wa для случая двухфазной системы “древесинное вещество–вода” была найдена аналитически (Кайбияйнен и др., 1981):
(5)
$\begin{gathered} a = ({{\gamma }_{{{\text{др}}}}}{{W}_{{\text{a}}}} - 0.46)({{\lambda }_{{\text{в}}}} - {{\lambda }_{{{\text{др}}}}}) \\ + \,\,(0.54{{W}_{{\text{a}}}} + 0.7){{\lambda }_{{{\text{др}}}}}{{[0.5({{W}_{{\text{a}}}} + 0.33)]}^{{--1}}}, \\ \end{gathered} $Таким образом, определив параметры t0 и tm из кривой распространения теплового импульса и дальнейших расчетов посредством приведенных уравнений, определяли U и Wа. В качестве системы измерения и регистрации применяли усилитель постоянного тока с самописцем (Кайбияйнен и др., 1981).
Сопротивления в звеньях “почва–корень” (Rп–к, МПа с м–3), “корень–охвоенный побег” (Rк–поб, МПа с м–3) рассчитывали по формулам, полученным из уравнения (1):
(6)
${{R}_{{{\text{п}} - {\text{к}}}}} = ({{\Psi }_{{\text{п}}}} - {{\Psi }_{{\text{к}}}}){{({{\bar {U}}_{{{\text{уд}}}}}{{S}_{с}})}^{{ - 1}}},$(7)
${{R}_{{{\text{к}} - {\text{поб}}}}} = ({{\Psi }_{{\text{к}}}} - {{\Psi }_{{{\text{поб}}}}}){{({{\bar {U}}_{{{\text{уд}}}}}{{S}_{с}})}^{{ - 1}}},$Удельное сопротивление (Rуд, МПа с м–2), характеризующее сопротивление движению влаги единицы поперечного сечения (м2) и единицы длины (м) ксилемы от корней до охвоенных побегов, рассчитывали по формуле:
где ∇Ψ = (Ψк – Ψпоб)h–1, h – расстояние от земли до части кроны, в которой определяются Ψпоб.Для обработки результатов использовали методы вариационной статистики (Ивантер, Коросов, 2003; и др.). Проверку гипотез и оценку существенных различий между средними величинами осуществляли при 5%-ом уровне значимости.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ данных многолетних наблюдений показателей водного режима Pinus sylvestris в условиях сосняка черничного свежего показал, что средние за вегетационный период минимальные за день значения водных потенциалов почвы (Ψп), корней (Ψк) и охвоенных побегов (Ψпоб), а также скорости водного потока по ксилеме ствола (${{\bar {U}}_{{{\text{уд}}}}}$) обладают малой вариабельностью в межгодичной динамике по сравнению с показателями внешней среды (табл. 1), и их величины равны –0.03, –0.32 ± 0.02, –1.06 ± ± 0.02 МПа и 3.2 ± 0.1 г см–2 ч–1 (8.9 × 10–6 м3 м–2 с–1) соответственно. Используя эти величины, по формулам (6) и (7) мы рассчитали сопротивления движению влаги в звеньях “почва–корень” (Rп–к) и “корень–охвоенный побег” (Rк–поб), которые составили 2.2 × 106 и 5.5 × 106 МПа с м–3 соответственно. При этом установили, что величина Rк–поб в 2.5 раза превосходит Rп-к, что свидетельствует об увеличении сопротивления и, напротив, снижении проводимости ксилемы вдоль транспортного пути. Полученный результат хорошо согласуется с известными данными других исследователей, которые показали, что у Picea abies (Roberts, 1978) и Picea sitchensis (Hellkvist et al., 1974) большее сопротивление также приходится на ксилему ствола и ветвей по сравнению с корнем.
Таблица 1.
Средние многолетние за вегетационный период (май–сентябрь) величины показателей водного обмена Pinus sylvestris и гидротермических условий в сосняке черничном свежем
| Показатель | x ± mx | Диапазон за вегетацию | C.V., % |
|---|---|---|---|
| Ψmax, МПа | –0.44 ± 0.01 | –0.40…–0.48 | 6 |
| Ψmin, МПа | –1.06 ± 0.02 | –0.85…–1.19 | 9 |
| U, г см–2 ч–1 | 3.2 ± 0.1 | 2.9–3.6 | 7 |
| Р, мм | 357 ± 13 | 300–392 | 10 |
| RH, % | 74.6 ± 1.5 | 56–90 | 12 |
| Tair, оС | 12.8 ± 0.6 | 7.0–20.5 | 26 |
| Tair_min, °С | 6.6 ± 0.6 | –1.4–11.9 | 54 |
| Tair_max, °С | 19.9 ± 0.9 | 9.8–30.2 | 27 |
| Tsoil, оС | 9.2 ± 0.6 | 1.1–14.9 | 35 |
| Wsoil, мм | 44.7 ± 1.8 | 26–65 | 24 |
Примечание. Ψmax – предрассветный водный потенциал побега; Ψmin – дневной водный потенциал побега; U – скорость водного потока; Р – суммарное количество атмосферных осадков; RH – относительная влажность воздуха; Tair – температура воздуха; Tair min – минимальная температура воздуха; Tairmax – максимальная температура воздуха; Tsoil – температура почвы на глубине 20 см; Wsoil – запасы влаги в горизонте почвы 0–50 см; x ± mx – среднее значение и стандартная ошибка; C.V. – коэффициент вариации.
Следует отметить, что величину Rп-к составляет как сопротивление самого корня растения, так и сопротивление переходного слоя “почва – корень”, которое может сильно зависеть от сопротивления почвы, ее влажности (North, Nobel, 1991; Irvine et al., 1998), качества контакта корня с почвой (Doussan et al., 2006; Draye et al., 2010), микоризных отношений (Nardini et al., 2000; Smith et al., 2010) и плодородия почвы (Tyree et al., 1998; Trubat et al., 2006). Однако в большей степени величина Rп–к зависит от влажности почвы. Так, например, у Pinus sylvestris (Roberts, 1977) и Pinus contorta (Running, 1980) в условиях дефицита почвенной влаги были получены более высокие значения Rп–к по сравнению со значениями Rк–поб. Наше исследование проходило в условиях сосняка черничного, для которого характерен высокий уровень почвенной влагообеспеченности (Сазонова и др., 2017), о чем свидетельствует высокое значение Ψп в течение всех лет исследований. Кроме того, исследования гидравлической проводимости ксилемы ствола, ветвей и корней, проведенные на отрезках древесины Picea abies (Sellin, 1993, 2001), показали, что она уменьшается в направлении от корней к стволу и далее к ветвям. При этом проводимость корней в 5 раз превышает проводимость ствола. Следовательно, чем удаленней орган от почвы – источника воды, тем меньше его проводимость и, напротив, больше сопротивление движению влаги. Отмечают также адаптивную роль высокой водопроводимости ксилемы корней древесных растений. Низкая величина Rк создает условия для лучшего транспорта воды из корневой системы, обеспечивая более стабильное водоснабжение надземных органов дерева в изменяющихся экологических условиях, поскольку доступность почвенной воды для растений может существенно варьировать.
Полученная нами для Pinus sylvestris величина Rк–поб хорошо согласуется с известными данными других авторов (Roberts, 1977; Whitehead et al., 1984) с учетом различий в объектах и методиках исследований. Поскольку Rк–поб зависит от размеров и возраста дерева, то как для внутривидового, так и для межвидового сравнения величин R более предпочтительной является величина удельного сопротивления Rуд. Следует отметить, что рассчитанные нами величины сопротивлений являются усредненными за вегетацию значениями, поскольку рассчитаны по средним за вегетационные периоды показателям. Результаты наших многолетних наблюдений (Сазонова и др., 2011) показали, что величины Uуд и Ψпоб закономерно изменяются в течение суток и вегетационного периода. Поэтому следует ожидать таких же изменений и величин сопротивления.
Для выявления диапазона вариабельности R мы проанализировали взаимосвязь Uуд – ∇Ψ, где ∇Ψ = (Ψк – Ψпоб)h–1. Представленные зависимости (рис. 2) являются результатом одновременного определения величин Uуд и Ψпоб в суточной и сезонной динамике. При расчете ∇Ψ величинуΨк считали постоянной, поскольку она значительно меньше варьировала в течение суток и вегетации по сравнению с Ψпоб. При этом нами было отмечено, что при одном и том же значении ∇Ψ в стволе могут регистрироваться водные потоки с разной скоростью. Кроме того, при уменьшении влажности ксилемы ствола (Wа) было показано снижение скорости водного потока при постоянстве ∇Ψ. В этой связи мы условно разбили поле точек на три области, соответствующие значениям Wа, равным 125–115, 115–105 и 105–95% (кривые 1–3 на рис. 2). Следует отметить, что точки, расположенные в верхней части поля (кривая 1), были получены в холодные и влажные дни весны и осени или в ранние утренние часы летом, а также после обильных дождей. Напротив, данным, полученным в сухие жаркие периоды лета, соответствуют точки, расположенные в нижней части поля (кривая 3). При этом если в первом случае (кривая 1) значения Wа максимальные, то во втором (кривая 3) – меньшие. Следовательно, чем выше значения Wа, тем ниже гидравлическое сопротивление (выше проводимость), и, напротив, уменьшение Wа приводит к увеличению сопротивления (уменьшению проводимости). Поэтому, несмотря на постоянство ∇Ψ, значения Uуд могут существенно варьировать. В то же время в выделенных по величине Wа областях (кривые 1–3) зависимости u = f(∇Ψ) хорошо аппроксимируются линейной функцией (R2 = 0.91…0.96). Этот факт свидетельствует о возможности использования аналога закона Ома для определения величины R без существенных ошибок.
Рис. 2.
Зависимость удельной скорости потока влаги в ксилеме ствола (Uуд) от градиента водного потенциала (∇Ψ) при разной абсолютной влажности древесины (Wа): 125–115 (1), 115–105 (2) и 105–95% (3).
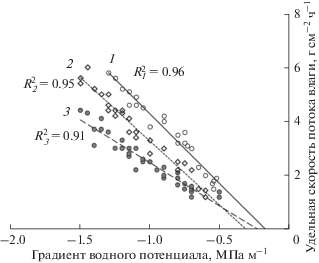
Проведенный по формуле (8) расчет удельных сопротивлений движению влаги по стволу Pinus sylvestris показал, что при изменении Wа от ее максимальных до минимальных значений диапазон Rуд составил 0.39 × 104…0.84 × 104 МПа с м–2. Сравнение полученных величин с известными данными для других видов хвойных растений обнаружило их близость. Так, RудPinus contorta составило 0.5 × 104 МПа с м–2 (Running, 1980). Более высокие значения Rуд ксилемы были выявлены у Picea sitchensis и составили 1.0 × 104…1.3 × 104 МПа с м–2 (Hellkvist et al., 1974). В то же время отмечают меньшие различия Rуд у различных хвойных растений по сравнению с лиственными видами, существенно различающимися строением проводящих воду путей (Cruiziat et al., 2002; Martínez-Vilalta et al., 2004; Bouche et al., 2014; и др.). Так, например, Rуд хвойных растений превышает Rуд рассеяннососудистых и кольцесосудистых лиственных пород в 5 и 10 раз соответственно.
Поскольку сопротивление движению влаги по ксилеме существенно зависит от влажности ксилемы, представляет интерес анализ динамики Wа. Такие исследования для Pinus sylvestris были проведены нами ранее (Кайбияйнен, Сазонова, 1993). Они показали, что наибольшие изменения Wа в течение суток, в частности в полуденные часы, наблюдаемые в дни с интенсивной транспирацией, составляли 15%. И это соответствовало изменению запасов в заболони ствола, объем которой составлял 100 дм3, на 7.3 л. Наряду с суточными изменениями запасов влаги в заболони ствола были обнаружены и сезонные ритмы Wа. Так, отмечалось уменьшение влажности ствола с мая по июль с дальнейшим ее повышением в сентябре. Для Pinus sylvestris максимальная и минимальная величины Wа были равны 140 и 85%, что соответствовало влагозапасам 75 и 65 л соответственно (Кайбияйнен, Сазонова, 1993). Очевидно, что жизнедеятельность древесного растения сопровождается непрерывным и закономерно изменяющимся в течение суток и вегетационного периода процессом дегидратации – регидратации ствола (Köcher et al., 2013). Это обусловливает и закономерные изменения R ксилемы, происходящие непрерывно в течение всего транспирационного периода. Результаты автоматической регистрации суточных и сезонных ритмов изменений диаметра ствола Pinus sylvestris, проведенной ранее на тех же объектах (Балыков, 1979), также отражают динамику величины R ксилемы в процессе регидратации – дегидратации ствола. Так, в ходе исследований колебаний диаметра ксилемы ствола были выявлены изменения в течение вегетационного периода, составившие за сезон 100 мкм. Для дерева, диаметр ксилемы которого и объем заболони составил 16 см и 100 дм3 соответственно, подобное изменение диаметра ксилемы ствола соответствует изменению объема заболони на 3%. В то же время, если в суточной динамике в процессе транспирации Wa ксилемы может уменьшиться на 15%, что соответствует снижению запасов влаги в стволе на 7.3 л, то в вегетационной динамике ствол может израсходовать на транспирацию 26.8 л (Кайбияйнен, Сазонова, 1993). Тогда в случае эластичности стенок водопроводящих сосудов подобный расход влаги привел бы к уменьшению объема заболони на 7.3 и 26.8% соответственно, что превышает наблюдаемое сокращение (3%) объема трахеид.
Отмеченное несоответствие, вероятно, обусловлено тем, что освобождающееся пространство замещается водяными парами. Процесс замещения в ксилеме расходуемой на транспирацию воды газом в результате кавитации общеизвестен (Tyree, Ewers, 1991; Cruiziat et al., 2002; и др.). Образовавшиеся пузырьки блокируют поры трахеид, разрывают водные нити, что приводит к нарушению транспорта влаги в определенной доле поперечного сечения ксилемы. В случае сильного обезвоживания ксилемы вода в результате кавитации замещается газом, что приводит к закупорке трахеид (Zimmermann, 1983; Cochard, 1992; Brodersen et al., 2013) и, как следствие, к уменьшению проводимости ксилемы и, напротив, увеличению сопротивления движения влаги. Вместе с тем, при сильном уменьшении проводимости ксилемы вдоль транспортного пути для поддержания интенсивности тока воды необходимо повышение ∇Ψ. Для сохранения притока воды из корней происходит уменьшение водного потенциала хвои (Ψхв), негативным последствием которого является усиление дегидратации ксилемы. Увеличение степени водного стресса повышает вероятность эмболии проводящих элементов, необратимое нарушение транспортной функции которых вызывает катастрофическую дисфункцию ксилемы ствола и гибель растения.
Однако адаптивные механизмы растения в большинстве случаев позволяют избежать катастрофической эмболии (Wullschleger et al., 1998; Cruiziat et al., 2002; Franks et al., 2007; Nardini et al., 2018). При определенной величине Ψхв начинается устьичное ограничение транспирации, что снижает интенсивность дегидратации ксилемы. Степень и интенсивность такой гидравлической регуляции варьируют среди видов и зависят от размера, структуры и функционального типа растения (Buckley, 2005; Domec et al., 2010; Meinzer, McCulloh, 2013; Martín-Gómez et al., 2017). Кроме того, значимый вклад в регуляцию водного транспорта вносят радиальные потоки воды между ксилемой, флоэмой и живыми клетками паренхимы, гидравлически их связывая и обеспечивая, таким образом, растение некоторой буферной емкостью для смягчения колебаний между транспирацией листа и поступлением воды в корень (Гамалей, 2004; Sevanto et al., 2011; Stroock et al., 2014). Большое внимание уделяется также гидравлическим характеристикам листа, в частности, гидравлической проводимости живых тканей листа за пределами ксилемы, включая устьица, сосудистую паренхиму и мезофилл листа (Sack et al., 2016; Scoffoni et al., 2017). У исследуемого нами хвойного растения Pinus sylvestris ограничение транспирационных расходов влаги посредством закрытия устьиц начинается при значениях Ψхв –1.0…–1.1 МПа (Сазонова и др., 2005). Отмеченная “пороговая” величина видоспецифична и имеет большой физиологический смысл, так как указывает на взаимную связь регуляторных механизмов потоков влаги в ксилеме и транспирации. Для разных видов и родов хвойных пород диапазон пороговых значений Ψхв составляет –1.1…–2.5 МПа (Running et al., 1975; Jarvis, 1980; и др.). В ряде исследований критический порог потери влаги также связывают с величиной водного потенциала, индуцирующего потерю гидравлической проводимости ксилемы ствола хвойных (Brodribb, Cochard, 2009) и лиственных древесных растений (Urli et al., 2013) на 50 и 80% соответственно. Следует также отметить, что чем ниже значения Ψхв, тем сильнее и продолжительнее ограничение транспирации. Подобное явление депрессии транспирации у Pinus sylvestris наблюдается вплоть до снижения значений Ψхв до –1.5 МПа. В данных пределах изменения Ψхв процесс замещения воды газом в ксилеме в результате кавитации имеет обратимый характер. Таким образом, изменяющееся устьичное и ксилемное сопротивление, посредством которых растение удерживает величину водного дефицита, в частности Ψ в определенном диапазоне, является адаптивным механизмом регуляции водного гомеостаза.
Заключение. Результаты наших многолетних исследований показателей водного обмена (Ψ и U) Pinus sylvestris на фоне изменяющихся в течение суток и вегетационного периода гидротермических условий позволили оценить межгодичную динамику этих величин. Нами показано постоянство усредненных за вегетационные периоды среднесуточных значений водного потенциала охвоенных побегов и скорости водного потока по стволам от года к году. Использование этих данным для расчета сопротивления движению влаги в звеньях “почва–корень” и “корень–побег” позволило выявить количественные характеристики процесса водного обмена Pinus sylvestris, не зависящие от погодных условий конкретного полевого сезона. Этот результат представляет особую ценность при прогнозировании возможного отклика как отдельного вида, так и лесной экосистемы в целом на изменения природной среды и климата. Наряду с этим в результате нашего исследования показана тесная взаимосвязь между сопротивлением ксилемы и ее оводненностью, а также рассчитаны пределы изменчивости R в связи с изменением Wa. Проведенное исследование взаимосвязи функциональных показателей водообмена Pinus sylvestris в условиях таежной зоны позволило нам выявить особенности механизма эндогенной регуляции этого процесса, а также дать количественную оценку пороговых величин гидравлической дисфункции древесного растения. Такая оценка важна для понимания динамики структуры и видового состава растительного сообщества в ответ на изменение условий внешней среды.
Список литературы
Балыков Н.Г. Методы и устройства автоматической регистрации процессов роста древесных растений // Биофизические методы исследований в экофизиологии древесных растений. Л.: Наука, 1979. с. 18–34.
Гамалей Ю.В. Транспортная система сосудистых растений. СПб: Изд-во Санкт-Петербургского государственного университета, 2004. 424 с.
Ивантер Э.В., Коросов А.В. Введение в количественную биологию. Петрозаводск: Изд-во Петрозаводского государственного университета, 2003. 304 с.
Кайбияйнен Л.К. Эколого-физиологические исследования сосны и сосновых древостоев // Труды Карельского НЦ РАН. 2003. Вып. 5. С. 65–73.
Кайбияйнен Л.К., Сазонова Т.А. Запасы воды в древесине сосны обыкновенной // Лесоведение. 1993. № 6. С. 58–64.
Кайбияйнен Л.К., Сазонова Т.А., Тихов П.В. Транспирационные потоки в ксилеме сосны и динамика потребления влаги // Лесоведение. 1981. № 2. С. 27–34.
Назарова Л.Е. Климат республики Карелия (Россия): температура воздуха, изменчивость и изменения // Геополитика и экогеодинамика регионов. 2014. № 1. С. 746−749.
Назарова Л.Е. Атмосферные осадки в Карелии // Труды Карельского НЦ РАН. 2015. № 9. С. 114−120.
Романов А.А. О климате Карелии. Петрозаводск: Карелия, 1961. 140 с.
Сазонова Т.А., Болондинский В.К., Придача В.Б. Эколого-физиологическая характеристика сосны обыкновенной. Петрозаводск: Verso, 2011. 207 с.
Сазонова Т.А., Болондинский В.К., Придача В.Б. Влияние водного дефицита хвои сосны обыкновенной на фотосинтез в условиях достаточного почвенного увлажнения // Лесоведение. 2017. № 4. С. 311–318.
Сазонова Т.А., Кайбияйнен Л.К., Колосова С.В. Диагностика водного режима Pinus sylvestris (Pinaceae) // Ботанический журн. 2005. Т. 90. № 7. С. 1012−1022.
Соловьев С.А. Применение тензиометров в экологических исследованиях // Экология. 1971. № 6. С. 93–95.
Berry J.A., Beerling D.J., Franks P.J. Stomata: key players in the earth system, past and present // Current Opinion in Plant Biology. 2010. V. 13. P. 232–239.
Bouche P.S., Larter M., Domec J.C., Burlett R., Gasson P., Jansen S., Delzon S. A broad survey of hydraulic and mechanical safety in the xylem of conifers // Journal of Experimental Botany. 2014. V. 65. P. 4419–4431.
Brodersen C.R., McElrone A.J., Choat B., Lee E.F., Shackel K.A., Matthews M.A. In vivo visualizations of drought-induced embolism spread in Vitis vinifera // Plant Physiology. 2013. V. 161. P. 1820–1829.
Brodribb T.J., Cochard H. Hydraulic failure defines the recovery and point of death in water-stressed conifers // Plant Physiology. 2009. V. 149. P. 575–584.
Buckley T.N. The control of stomata by water balance // New Phytologist. 2005. V. 168. P. 275–292.
Carminati A. Rhizosphere wettability decreases with root age: a problem or a strategy to increase water uptake of young roots? // Frontiers in Plant Science. 2013. V. 4. P. 1–9.
Cochard H. Vulnerability of several conifers to air embolism // Tree Physiology. 1992. V. 11. P. 73–83.
Cruiziat P., Cochard H., Améglio T. Hydraulic architecture of trees: main concepts and results // Annals of Forest Science. 2002. V. 59. P. 723–752.
Domec J.C., Schäfer K., Oren R., Kim H.S., McCarthy H.R. Variable conductivity and embolism in roots and branches of four contrasting tree species and their impacts on whole-plant hydraulic performance under future atmospheric CO2 concentration // Tree Physiology. 2010. V. 30. P. 1001–1015.
Doussan C., Pierret A., Garrigues E., Pagès L. Water uptake by plant roots: II – modelling of water transfer in the soil root-system with explicit account of flow within the root system – comparison with experiments // Plant & Soil. 2006. V. 283. P. 99–117.
Draye X., Kim Y., Lobet G., Javaux M. Model-assisted integration of physiological and environmental constraints affecting the dynamic and spatial patterns of root water uptake from soils // Journal of Experimental Botany. 2010. V. 61. P. 2145–2155.
Fatichi S., Pappas C., Ivanov V. Modeling plant–water interactions: an ecohydrological overview from the cell to the global scale // WIREs: Water. 2015. P. 1–42.
Franks P.J., Drake P.L., Froend R.H. Anisohydric but isohydrodynamic: seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance // Plant, Cell & Environment. 2007. V. 30. P. 19–30.
Granier A., Breda N., Claustres J.P., Colin F. Variation of hydraulic conductance of some adult conifers under natural conditions // Annals of Forest Science. 1989. V. 46. P. 357–360.
Hellkvist J., Richards G., Jarvis P.G. Vertical gradients of water potential and tissue relations in Sitka spruce trees measured with the pressure chamber // J. Applied Ecology. 1974. V. 11. P. 637–667.
Hinckley T.M., Lassoie J.R., Running S.W. Temporal and spatial variations in the water status of forest trees // Forest Science. 1978. V. 24. P. 186–194.
Huber B. Die beurteilung des wasserhaushaltes der pflanze // Jb. Wiss. Botanik. 1924. V. 64. S. 1–20.
Irvine J., Perks M.P., Magnani F., Grace J. The response of Pinus sylvestris to drought: stomatal control of transpiration and hydraulic conductance // Tree Physiology. 1998. V. 18. P. 393–402.
Jarvis P.G. Water transfer in plants // Heat and mass transfer in the plant environment. Washington: Scripta, 1975. P. 369–394.
Jarvis P.G. Stomatal response to water stress in conifers // Adaptation of plants to water and high temperature stress. N. Y.: Academic Press, 1980. P. 105–121.
Katul G.G., Oren R., Manzoni S., Higgins C., Parlange M.B. Evapotranspiration: a process driving mass transport and energy exchange in the soil-plant-atmosphere-climate system // Reviews of Geophysics. 2012. V. 50. P. 1–25.
Köcher P., Horna V., Leuschner C. Stem water storage in five coexisting temperate broad-leaved tree species: significance, temporal dynamics and dependence on tree functional traits // Tree Physiology. 2013. V. 33. P. 817–832.
Kramer P.J., Boyer J.S. Water relations of plants and soil. N. Y.: Academic Press, 1995. 495 p.
Lu P., Biron P., Granier A., Cochard H. Water relations of adult Norway spruce (Picea abies (L) Karst.) under soil drought in the Vosges mountains: whole-tree hydraulic conductance, xylem embolism and water loss regulation // Annals of Forest Science. 1996. V. 53. P. 113–121.
Marshall D.C. Measurement of sap flow in conifers by heat transport // Plant Physiology. 1958. V. 39. P. 385–396.
Martín-Gómez P., Aguilera M., Pemán J., Gil-Pelegrín E., Ferrio J.P. Contrasting ecophysiological strategies related to drought: the case of a mixed stand of Scots pine (Pinus sylvestris) and a submediterranean oak (Quercus subpyrenaica) // Tree Physiology. 2017. V. 37. P. 1478–1492.
Martínez-Vilalta J., Sala A., Piñol J. The hydraulic architecture of Pinaceae – a review // Plant Ecology. 2004. V. 171. P. 3–13.
McCulloh K.A., Meinzer F.C., Sperry J.S., Lachenbruch B., Voelker S.L., Woodruff D.R., Domec J.C. Comparative hydraulic architecture of tropical tree species representing a range of successional stages and wood density // Oecologia. 2011. V. 167. P. 27–37.
Meinzer F.C., McCulloh K.A. Xylem recovery from drought-induced embolism: where is the hydraulic point of no return? // Tree Physiology. 2013. V. 33. P. 331–334.
Nardini A., Salleo S., Tyree M.T., Vertovec M. Influence of the ectomycorrhizas formed by Tuber melanosporum Vitt. on hydraulic conductance and water relations of Quercus ilex L. seedlings // Annals of Forest Science. 2000. V. 57. P. 305–312.
Nardini A., Savi T., Trifilò P., Gullo M.A. Drought stress and the recovery from xylem embolism in woody plants // Progress in Botany. 2018. V. 79. P. 197–232.
North G.B., Nobel P.S. Changes in hydraulic conductivity and anatomy caused by drying and rewetting roots of Agave deserti (Agavaceae) // American Journal of Botany. 1991. V. 78. P. 906–915.
Olchev A.V., Deshcherevskaya O.A., Kurbatova Y.A., Molchanov A.G., Novenko E.Y., Pridacha V.B., Sazonova T.A. CO2 and H2O exchange in the forest ecosystems of southern taiga under climate change // Doklady Biological Sciences. 2013. V. 450. P. 173−176.
Roberts J. The use of tree-cutting techniques in the study of the water relations of mature Pinus sylvestris L. I. The technique and survey of the results // Journal of Experimental Botany. 1977. V. 28. P. 751–767.
Roberts J. The use of the ‘tree cutting’ technique in the study of the water relations of Norway spruce, Picea abies (L.) Karst. // Journal of Experimental Botany. 1978. V. 29. P. 465–471.
Running S.W. Environmental and physiological control of water flux through Pinus contorta // Canadian J. Forest Research. 1980. V. 10. P 82–91.
Running S.W., Waring R.H., Rydell R.A. Physiological control of water flux in conifers // Oecologia. 1975. V. 18. P. 1–16.
Sack L., Holbrook N.M. Leaf hydraulics // Annual Review of Plant Biology. 2006. V. 57. P. 361–381.
Sack L., Buckley T.N., Scoffoni C. Why are leaves hydraulically vulnerable? // Journal of Experimental Botany. 2016. V. 67. P. 4917–4919.
Scoffoni C., Albuquerque C., Brodersen C., Townes S.V., John G.P., Bartlett M.K., Buckley T.N., McElrone A.J., Sack L. Outside-xylem vulnerability, not xylem embolism, controls leaf hydraulic decline during dehydration // Plant Physiology. 2017. V. 173. P. 1197–1210.
Sellin A. Resistance to water flow in xylem of Picea abies (L.) Karst. trees grown under contrasting light conditions // Trees. 1993. V. 7. P. 220–226.
Sellin A. Hydraulic and stomatal adjustment of Norway spruce trees to environmental stress // Tree Physiology. 2001. V. 21. P. 879–888.
Sevanto S., Hölttä T., Holbrook N.M. Effects of the hydraulic coupling between xylem and phloem on diurnal phloem diameter variation // Plant, Cell & Environment. 2011. V. 34. P. 690–703.
Smith S.E., Facelli E., Pope S., Smith F.A. Plant performance in stressful environments: interpreting new and established knowledge of the roles of arbuscular mycorrhizas // Plant & Soil. 2010. V. 326. P. 3–20.
Sperry J.S., Wang Y., Wolfe B.T., Mackay D.S., Anderegg W.R., McDowell N.G., Pockman W.T. Pragmatic hydraulic theory predicts stomatal responses to climatic water deficits // New Phytologist. 2016. V. 212. P. 577–589.
Stroock A.D., Pagay V.V., Zwieniecki M.A., Holbrook N.M. The physicochemical hydrodynamics of vascular plants // Annual Review of Fluid Mechanics. 2014. V. 46. P. 615–642.
Swanson R.H. Water transpired by trees is indicated by heat pulse velocity // Agricultural Meteorology. 1972. V. 10. P. 277–281.
Trubat R., Cortina J., Vilagrosa A. Plant morphology and root hydraulics are altered by nutrient deficiency in Pistacia lentiscus (L.) // Trees. 2006. V. 20. P. 334–339.
Tyree M.T., Ewers F.W. The hydraulic architecture of trees and other woody plants // New Phytologist. 1991. V. 119. P. 345–360.
Tyree M.T., Velez V., Dalling J.W. Growth dynamics of root and shoot hydraulic conductance in seedlings of five neotropical tree species: scaling to show possible adaptation to differing light regimes // Oecologia. 1998. V. 114. P. 293–298.
Urli M., Porté A.J., Cochard H., Guengant Y., Burlett R., Delzon S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees // Tree Physiology. 2013. V. 33. P. 672–83.
Van den Honert T.H. Water transport in plants as a catenary process // Interaction of water and porous materials. Discussions of the Faraday Society. 1948. V. 3. P. 146–153.
Whitehead D., Jarvis P.G. Coniferous forest and plantations // Water deficits and plant growth. N. Y.: Academic Press, 1981. V. 6. P. 50−151.
Whitehead D., Jarvis P.G., Waring R.H. Stomatal conductance, transpiration, and resistance to water uptake in a Pinus sylvestris spacing experiment // Canadian Journal of Forest Research. 1984. V. 14. P. 692–700.
Wullschleger S.D., Meinzer F.C., Vertessy R.A. A review of whole-plant water use studies in tree // Tree Physiology. 1998. V. 18. P. 499–512.
Zimmermann M.H. Xylem Structure and the aspect of sap. N. Y.: Springer-Verlag, 1983. 143 p.
Дополнительные материалы отсутствуют.