

Лесоведение, 2020, № 1, стр. 17-34
Вероятностная оценка распределения ключевых биотопов в лесах национального парка “Смоленское Поозерье” на основе спутниковых и топографических данных
Д. В. Ершов a, *, Е. А. Гаврилюк a, Е. В. Тихонова a, Т. Ю. Браславская a, Н. В. Королева a, И. М. Бавшин b, Д. Г. Груммо c, А. В. Судник c, А. В. Титовец b
a Центр по проблемам экологии и продуктивности лесов
117997 Москва, ул. Профсоюзная, 84/32, стр. 14, Россия
b Национальный парк “Смоленское Поозерье”
216270 Демидовский район, Смоленская область, пос. Пржевальское, ул. Гуревича, 19, Россия
c Институт экспериментальной ботаники им. В.Ф. Купревича НАН Беларуси
220072 Минск, ул. Академическая, 27, Республика Беларусь
* E-mail: ershov@ifi.rssi.ru
Поступила в редакцию 19.03.2019
После доработки 29.04.2019
Принята к публикации 08.10.2019
Аннотация
Изучена возможность вероятностной оценки пространственного распределения ключевых биотопов в лесах национального парка “Смоленское Поозерье” с использованием спектральных и топографических характеристик местности. Из представленных на территории парка 19 типов лесных биотопов (классификация EUNIS) для исследований были выбраны следующие 4 ключевых: бореально-неморальные черничные еловые таежные леса, сосновые и елово-сосновые брусничные таежные леса, дубово-липовые леса и подтаежные заболоченные ольховые леса. Все прочие типы лесных биотопов рассматривались совместно, как второстепенные относительно выбранных ключевых. Мы использовали разносезонные мультиспектральные спутниковые изображения Sentinel-2 совместно с топографическими показателями, рассчитанными на основе цифровой модели рельефа и гидрографической сети, в качестве исходных данных для геопространственного моделирования распространения биотопов по территории. Априорная информация о лесах парка была получена из материалов лесоустройства, которые использовались для обучения вероятностной модели, и результатов геоботанических обследований, примененных для независимой оценки полученных результатов. Тематическая классификация проводилась методом случайных лесов. Точность полученных вероятностных оценок распространения ключевых биотопов по территории парка, рассчитанная по показателю площади под ROC-кривой, варьировала от 88 до 96%, а значения показателя Бриера изменялись от 0.03 до 0.07, что характеризует полученную нами модель как достаточно надежную. Общая площадь, занимаемая четырьмя ключевыми биотопами, согласно нашим данным, составила 14.6 тыс. га, или около 10% от площади национального парка. Сравнение полученных вероятностных оценок с материалами геоботанических описаний дало довольно противоречивые результаты: степень соответствия варьирует от 30 до 95% в зависимости от биотопа. Это может являться следствием особенностей планирования и проведения наземных обследований, вошедших в контрольную выборку, которые изначально не предназначались для обучения и оценки качества геопространственных моделей.
Деградация и потеря местообитаний в результате изменения землепользовании в условиях меняющегося климата остается важным фактором сокращения биоразнообразия (Hansen et al., 2004). Получение количественных оценок распространения и состояния местообитаний является фундаментальной задачей для разработки общей политики по поддержанию и улучшению биоразнообразия в Европе (Mücher et al., 2009). Для обеспечения требований международных процессов, связанных с мониторингом и отчетами по биоразнообразию, в том числе Бернской конвенции, Конвенции по сохранению биоразнообразия “Цели-2020”, была разработана единая европейская классификация всех типов наземных и водных местообитаний EUNIS (Davies et al., 2004). Классификация EUNIS имеет иерархическую структуру, в разных ее разделах число выделяемых уровней составляет от пяти до восьми (EUNIS …, 2019). На высших (обобщающих) уровнях выделение категорий проводится по ландшафтным, экологическим, антропогенным и ботаническим критериям, а на низшем (базовом) – чаще всего на основе географического (регионального) подхода или же по критериям важной роли местообитаний в жизнедеятельности одного или нескольких редких и уязвимых видов (Браславская и др., 2018).
В научной литературе и природоохранной практике используется несколько определений понятия “биотоп” (англ. biotope), которое подразумевает сочетание факторов физической среды (например, типа почв и климата) с сообществом растений и животных. Близким ему по смыслу является широко употребляемое определение понятия “местообитание” (англ. habitat), содержащееся в The European Nature Information System (EUNIS) habitats classification (Раздел 3.2.1): “Место, где растения или животные обычно живут, характеризуемое в первую очередь по параметрам физической среды (топография, почвенные характеристики, климат, качество воды и т.д.) и, во-вторых, видами растений и животных, которые там обитают”. Для практического применения этого понятия существенно предложение включать в него те участки, площадь которых составляет не менее 100 м2 (Davies et al., 2004). В рамках нашего исследования эти два понятия употребляются как синонимы. Закон Республики Беларусь от 26 ноября 1992 г. № 1982-XII “Об охране окружающей среды” определяет биотоп как “природный объект (участок территории или акватории) с однородными экологическими условиями, являющийся местом обитания сообщества тех или иных видов диких животных и произрастания дикорастущих растений”, а ключевой биотоп – как обладающий какими-либо уникальными (отличными от окружения) свойствами, благодаря которым он имеет повышенное значение для сохранения биоразнообразия для данной территории (Редкие …, 2013).
В России необходимость классифицировать местообитания и выделять их ключевые типы возникает, прежде всего, у биологов и экологов, участвующих в международных проектах по разработке трансграничных способов охраны редких и уязвимых видов, биоразнообразия в целом (Крышень и др., 2009; Браславская, 2016). Также выявление участков приоритетных (ключевых) типов местообитаний актуально в связи с участием России в формировании Изумрудной сети (the Emerald Network) в рамках Конвенции о сохранении европейской дикой природы и естественной среды обитания (Бернская конвенция) (Изумрудная книга …, 2011–2013).
В качестве критериев выделения биотопов используются признаки растительности и экотопа (Крышень и др., 2009; Королева, 2011). Флористический состав растительных сообществ, особенно в естественных и малонарушенных условиях среды, в значительной мере определяется абиотическими факторами, поэтому синтаксоны (типы растительных сообществ) могут служить индикаторами местообитаний наземных экосистем (Terrestrial …, 2014). Было установлено, что около 60% местообитаний имеют четкую связь с одним или несколькими синтаксонами флористической классификации (Evans, 2010). Выявленные связи местообитаний с синтаксонами растительности позволили определить флористический состав лесных типов местообитаний EUNIS (Schaminée et al., 2013). Однако применение синтаксономического подхода для выделения типов биотопов в средней полосе Европейской России затруднено из-за очень ограниченного предшествующего опыта использования классификации EUNIS (Изумрудная сеть …, 2015; Браславская, 2016, 2017), а также фрагментарности геоботанических данных и недостатка экспертных знаний о степени соответствия категорий системы EUNIS существующим классификациям растительности.
Исследованию возможностей применения спутниковых данных в комбинации с другой тематической информацией для картографирования биотопов посвящено немало научных работ (Nagendra et al., 2013; Corbane et al., 2015). В зависимости от охвата территории эта задача решается с помощью разных источников пространственных данных. При пространственном моделировании биотопов из классификации EUNIS на континентальном уровне с пространственным разрешением 1 км применяются два метода: “снизу вверх”: пространственное моделирование с использованием данных геоботанических описаний и “сверху вниз”: согласование карты земного покрова с пространственными данными о биотопах. В первом подходе используются спутниковые тематические карты растительного покрова наземных экосистем и данные наземных наблюдений на площадках из европейского архива растительности EVA (Schaminée et al., 2014), по которым определяются условия местопроизрастания и устанавливаются связи с параметрами окружающей среды (почва, климат и др.). Затем осуществляется пространственное моделирование вероятности расположения биотопа методом максимальной энтропии MaxEnt (Berger et al., 1996; Philips et al., 2006). В качестве индикаторов окружающей среды для биотопа используются характеристики потенциальной эвапотранспирации и прямой солнечной радиации, уровень кислотности верхнего горизонта почв, сезонной динамики температур воздуха и осадков, максимума нормализованного вегетационного индекса и удаленности от водных объектов (река, озеро и море). Для уточнения местоположения биотопов применяется второй подход (“сверху вниз”), который предусматривает использование спутниковых карт породной структуры лесов и сомкнутости древостоя высокого разрешения (20 м) в сочетании с результатами наземных обследований, собранных на регулярных сетях ICP-Forests и национальных систем инвентаризации стран Европы. В качестве предикторов состояния окружающей среды рассматриваются данные карт почвы, температуры и осадков. С помощью ГИС-анализа наземные данные регулярных сетей интерполируются до пространственного разрешения спутниковых карт, и затем с помощью MaxEnt моделируется вероятность местоположения биотопа на территории Европы (Mücher et al., 2015).
Для построения карт биотопов небольших локальных территорий многие авторы используют такие пространственные индикаторы, или предикторы биотопов, как данные рельефа (абсолютные высоты, крутизна склона и его ориентация) и его индексы (TWI, TPI и др.), величину падающей солнечной радиации, значения яркостных коэффициентов отражения земного покрова в спектральных диапазонах, данные лидарной съемки, сезонные климатические измерения температуры воздуха и осадков, почвенные и ландшафтные карты (Крицук, 2012; Díaz Varela et al., 2008; Kushwah et al., 2012; Tagil, 2015; Álvarez-Martínez et al., 2018).
Целью нашей работы является получение вероятностных оценок пространственного распространения ключевых лесных биотопов EUNIS на основе комбинации данных наземных обследований, серии спутниковых изображений и топографических характеристик местности, включая определение оптимального состава исходных индикаторов (признаков) и степени надежности получаемых результатов. Для проведения исследований была выбрана особо охраняемая природная территория федерального значения – национальный парк “Смоленское Поозерье”.
ОБЪЕКТЫ И МЕТОДИКА
Национальный парк “Смоленское Поозерье” создан в 1992 г. на землях Демидовского и Духовщинского районов Смоленской области для сохранения природных комплексов в рекреационных, просветительских, научных и культурных целях (Постановление …, 1992). Его общая площадь составляет 146 237 га, в том числе 114 444 га (78%) – это площадь земель особо охраняемых территорий и объектов (Лесохозяйственный …, 2015). К лесным землям (включая покрытые лесом) относятся 108 327 га (94.6%); среди нелесных земель преобладают болота и водоемы (4%).
Национальный парк расположен в границах трех существенно различающихся геоморфологических районов (Шкаликов, 1995; Шкаликов и др., 2005): большая часть территории относится к Слободской холмисто-моренной возвышенности и Аржатско-Ельшанской озерно-ледниково-зандровой низине, а самая восточная и юго-восточная части – к Духовщинской моренно-эрозионной возвышенности. Практически вся его территория находится в пределах области валдайского оледенения, и только ландшафты Духовщинской возвышенности сформировались в эпоху днепровского оледенения. В районе Слободской холмисто-моренной возвышенности наибольшее распространение имеют мелкохолмистые пологоволнистые моренные равнины. Рельеф центральной части этого района устроен особенно сложно, для него характерны крутые конечно-моренные холмы, комплексы озовых и озово-камовых образований. Южнее конечно-моренных образований распространены плоские и слабоволнистые зандровые равнины, которые занимают относительно большие площади. В районе Аржатско-Ельшанской озерно-ледниково-зандровой низины, расположенном восточнее р. Ельша, преимущественное распространение имеет водно-ледниковый рельеф, представленный плоскими и слабоволнистыми зандровыми и озерно-ледниковыми равнинами. Заметно меньшие площади занимают пологоволнистые и мелкохолмистые моренные равнины. В целом территория этого района на 10–20 м ниже первого, поэтому она заметно менее дренирована, здесь намного больше избыточно увлажненных земель и торфяников (рис. 1).
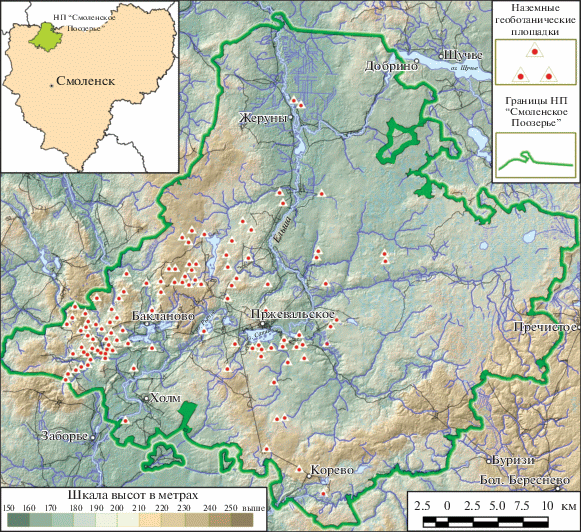
Леса территорий разнообразны по составу древесных видов, среди которых наибольшее представительство по площади имеют береза (41.2%), ель (13.8%), сосна (12.5%), осина (14%), ольха серая (9.3%), дуб (6.5%) и липа (2.4%). Для лесных биогеоценозов характерны подзолистые и дерново-подзолистые почвы разной степени оподзоливания и оглеения под лесами; кроме того, значительную часть территории национального парка занимают торфяные почвы (Научные …, 2003; Растительность …, 2003). Подзолистые почвы широко распространены под хвойными, преимущественно еловыми лесами, обеднены гумусом, азотом и элементами минерального питания. Дерново-подзолистые почвы распространены под хвойно-лиственными и лиственными (дубово-липовыми, березовыми, осиновыми) лесами. Для сосновых лесов, формирующихся на песчаных отложениях, характерно развитие подзолов и дерново-подзолов – кислых почв, бедных обменными основаниями, элементами минерального питания, азотом и гумусом. Химический состав торфяных почв определяется в основном видовым составом растений-торфообразователей: торфяные почвы верховых болот отличаются низкой зольностью, максимальное содержание зольных элементов характерно для торфяных почв низинных болот, из которых многие покрыты черноольховыми лесами.
В 2006–2018 гг. в национальном парке “Смоленское Поозерье” проводились геоботанические исследования лесов с геопривязкой заложенных временных учетных площадок (400 м2) при помощи GPS-навигатора, что позволяет использовать собранные данные в тематической обработке спутниковых изображений. В ходе выполнения геоботанических описаний характеризовали местоположения и ярусную структуру лесных сообществ, составляли полные списки видов сосудистых и моховидных растений, в ряде случаев выборочно определяли возраст деревьев при помощи бурава Пресслера (Методические …, 2010).
На основе классификации EUNIS составлен предварительный список типов биотопов (местообитаний), представленных на рассматриваемой территории. Для этого были проанализированы классификационные категории и их описательные характеристики, начиная с самого нижнего уровня, для которого наиболее детально и точно описаны диагностические признаки, в т. ч. географическая локализация типов биотопов. Используя материалы собственных геоботанических исследований (183 описания), а также фондовых и опубликованных источников, определяли принадлежность выявленных растительных сообществ к типологическим категориям системы EUNIS с помощью анализа флористических списков. Затем устанавливали, какие из них рассматриваются как ключевые в современной природоохранной практике (Annex …, 2007; Revised Annex …, 2010-2014). После этого на основе лесоустроительных материалов оценили наличие всех и ключевых биотопов в лесах национального парка. В результате применения изложенного выше подхода составлен перечень типов наземных биотопов, который включает 19 лесных, 2 кустарниковых, 8 болотных и 10 травяных (в т. ч. 7 луговых, 1 опушечный, 2 рудеральных). Среди них к ключевым относятся 12 лесных, 7 болотных и 1 луговой тип. Для исследований выбраны четыре типа ключевых лесных биотопов (табл. 1).
Таблица 1.
Краткая характеристика типов ключевых биотопов, выбранных для исследования возможности моделирования их пространственного распространения в лесах национального парка “Смоленское Поозерье"
| №№ п/п | Код EUNIS | Название
по классификации EUNIS |
Краткая характеристика |
|---|---|---|---|
| 1 | T3-F14 | Бореально-неморальные черничные еловые таежные леса | Леса с доминированием ели (Picea spp.) и участием сосны (Pinus sylvestris) и листопадных деревьев: дуба (Quercus spp.), липы мелколистной (Tilia cordata), клена остролистного (Acer platanoides), вяза (Ulmus spp.), осины (Populus tremula); в напочвенном покрове доминирует Vaccinium myrtillus |
| 2 | T3-G2 | Сосновые и елово-сосновые брусничные таежные леса | Хвойные леса с доминированием сосны и часто с участием ели (которая может доминировать или со-доминировать), с напочвенным покровом, в котором доминируют кустарнички (в основном Vaccinium vitis-idaea и/или Empetrum spp.), мхи (Dicranum spp., Hylocomium splendens, Pleurozium schreberi) и лишайники (Cladonia spp.). Видовой состав также включает: в древостое – березу пушистую (Betula pubescens), в напочвенном покрове – кустарнички Calluna vulgaris, Ledum palustre, Vaccinium myrtillus, V. uliginosum, травы Pyrola chlorantha, Goodyera repens, Deschampsia flexuosa, Lycopodium complanatum |
| 3 | T1-E16A | Дубово-липовые леса на севере широколиственной и в подтаежной зоне | Леса из дуба черешчатого (Quercus robur) и липы мелколистной (Tilia cordata), но без участия граба обыкновенного (Carpinus betulus); в составе древостоя также могут быть Acer platanoides, Populus tremula, Picea abies. Подлесок формируют Corylus avellana, Sorbus aucuparia, Euonymus verrucosa, Daphne mezereum, напочвенный покров – Galium odoratum, Anemone nemorosa и бореальные травы |
| 4 | T1-5114 | Подтаежные заболоченные ольховые леса | Мезотрофные и мезо-евтрофные заболоченные леса, сформированные ольхой черной (Alnus glutinosa) или серой (A. incana), при подкислении грунтовых вод – с примесью березы пушистой; в напочвенном покрове обычны гигрофильные травы (Calamagrostis canescens, Athyrium filix-femina, Cardamine amara, Filipendula ulmaria, Lysimachia thyrsiflora, Carex elongata, C. remota) и мхи (Brachythecium rivulare, Calliergon cordifolium, Climacium dendroides, Thuidium tamariscinum) |
Для моделирования распределения ключевых биотопов в лесах национального парка использовалось два типа геопространственных данных: мультиспектральные спутниковые изображения Sentinel-2 (ESA Sentinel-2, 2019) и базовые топографические показатели, рассчитанные с использованием цифровой модели рельефа и гидрографической сети местности. Было проанализировано более ста сцен Sentinel-2 из онлайн-архива Европейского космического агентства (ESA) за период с 2015 по 2018 г. и отобрано для тематической обработки шесть безоблачных разносезонных изображений (продукты уровня L1C) за следующие даты: 23 сентября 2017 г., 11 апреля, 11 мая, 26 мая, 24 августа и 18 октября 2018 г. Исходные значения спектральных яркостей на уровне верхних слоев атмосферы в каналах отобранных изображений были пересчитаны в соответствующие значения на уровне земной поверхности с использованием алгоритма MAJA (Hagolle et al., 2015). Цифровая модель рельефа местности была построена на основе горизонталей, оцифрованных по крупномасштабной топографической карте масштаба 1 : 25 000 (материалы обрабатывались сотрудниками национального парка). Прочие элементы географической основы, включая гидрографическую сеть, были сформированы из открытых данных проекта OpenStreetMap (Open …, 2019). Полный список использованных в работе спектральных и топографических признаков с пояснениями содержится в табл. 2. Для предварительной стратификации территории на покрытые и не покрытые лесом участки использовались актуальные (по состоянию на 2017 г.) материалы проекта GFC – Global Forest Change (Hansen et al., 2013), содержащие оценки сомкнутости и нарушений лесного покрова в глобальном масштабе, полученные на основе спутниковых данных Landsat. В качестве априорной информации о лесах парка использовались материалы лесоустройства 2014–2015 гг., включающие географически привязанную сеть выделов с полными таксационными описаниями.
Таблица 2.
Исходные спектральные и топографические признаки и комментарии по их использованию для вероятностной оценки пространственного распределения ключевых лесных биотопов
| Признак | Описание | Пространственное разрешение, м | Участие в анализе | Комментарий |
|---|---|---|---|---|
| Спектральные каналы Sentinel-2 (для каждой из 6 дат съемки) | ||||
| BLUE | Синий (0.439–0.535 мкм) | 10 | Нет | Сигнал зашумлен, коррелирован с GREEN и RED |
| GREEN | Зеленый (0.537–0.582 мкм) | 10 | Да | – |
| RED | Красный (0.646–0.685 мкм) | 10 | Да | – |
| VRE1 | Красный край (0.694–0.714 мкм) | 20 | Нет | Коррелирован с RED |
| VRE2 | Красный край (0.731–0.749 мкм) | 20 | Нет | Коррелирован с VRE1 и VRE3 |
| VRE3 | Красный край (0.768–0.796 мкм) | 20 | Нет | Коррелирован с NIR1 |
| NIR1 | Ближний инфракрасный (0.767–0.908 мкм) | 10 | Да | – |
| NIR2 | Ближний инфракрасный (0.848–0.881 мкм) | 20 | Нет | Коррелирован с NIR1, ниже разрешение |
| SWIR1 | Средний инфракрасный (1.539–1.681 мкм) | 20 | Нет | Коррелирован с RED и SWIR2 |
| SWIR2 | Средний инфракрасный (2.072–2.312 мкм) | 20 | Да | – |
| Спектральные индексы на основе каналов Sentinel-2 (для каждой из 6 дат съемки) | ||||
| NDVI | (NIR1 – RED1)/(NIR1 + RED1) | 10 | Да | (Rouse et al., 1974) |
| SWVI | (NIR2 –SWIR1)/(NIR2 +SWIR1) | 20 | Да | (Gao, 1996) |
| NDRE | (VRE3 – VRE1)/(VRE3 + VRE1) | 20 | Да | (Sims, Gamon, 2002) |
| Топографические признаки | ||||
| DEM | Высота над уровнем моря, м | 10 | Да | – |
| Slope | Крутизна склона, ° | 10 | Да | (Travis et al., 1975) |
| Aspect | Экспозиция склона, ° | 10 | Да | |
| GenCurv | Общая кривизна склона, м–1 | 10 | Да | (Shary, 1995) |
| TPI | Индекс топографического положения, м | 10 | Да | (Guisan et al., 1999) |
| TWI | Топографический индекс влажности | 10 | Да | (Beven, Kirkby, 1979) |
| DTW | Индекс глубины залегания грунтовых вод | 10 | Да | (Murphy et al., 2009) |
| TDI | Прямая инсоляция склона, Вт м-2 | 10 | Да | (Boehner, Antonic, 2009) |
| EDtW | Евклидово расстояние до ближайшего водного объекта, м | 10 | Да | – |
| CoordX | Координата пикселя по оси X (c запада на восток), м | 10 | Да | – |
| CoordY | Координата пикселя по оси Y (c севера на юг), м | 10 | Да | – |
Выборка эталонных участков для моделирования пространственного распределения ключевых и всех прочих лесных биотопов парка была сформирована на основе пространственного пересечения сети лесоустроительных выделов с результатами автоматической сегментации спутниковых изображений Sentinel-2 алгоритмом Full Lambda Schedule (Redding et al., 1999). Для сегментации использовались только каналы видимого и ближнего инфракрасного диапазонов с пространственным разрешением 10 м. Участки, образованные пересечением выделов и сегментов, отбирались для дальнейшего анализа по нескольким критериям: площадь (не менее 1 га), доля взаимного перекрытия (не менее 50%) и сомкнутость древостоев (не менее 50%). Затем они объединялись на основе таксационных характеристик в комплексные тематические группы, которые в дальнейшем рассматривались как прототипы для ключевых и прочих лесных биотопов. Для этого каждому участку присваивался восьмизначный составной код, первые пять цифр которого соответствовали долям ели, сосны, широколиственных пород, ольхи черной и всех прочих мелколиственных пород в составе, следующие две цифры отображали значения трофности и влажности условий местопроизрастания (по П.С. Погребняку), а последняя цифра идентифицировала возраст древостоев. При этом доли породного состава объединялись в три условных градации – преобладание (6 и более единиц в породной формуле), присутствие (1–5 единиц) и отсутствие той или иной группы пород. Показатели влажности были преобразованы в две категории – сухие или нормальные (1–3) и влажные (4–5) условия произрастания. Значения возраста также были сведены к двум группам – молодые (1–2 класс возраста) и взрослые (3 и более) древостои. По уникальным значениям составного кода было выделено 88 наиболее представленных на территории парка прототипов биотопов, которые суммарно покрывают более 95% от площади лесного фонда. Общий размер эталонной выборки, полученной на данном этапе, составил 1399 участков, при этом четыре прототипа, соответствующие ключевым биотопам, вошли в нее целиком, а для всех остальных групп случайным образом формировались подвыборки в объеме 20% от общего числа входящих в них участков, которые затем объединялись в один комплексный тематический класс “все прочие биотопы”. Такой подход позволил сформировать относительно сбалансированную по составу обучающую выборку – от 110 до 401 наблюдения в каждом из пяти классов, для дальнейшей вероятностной классификации.
Мы использовали метод случайных лесов (Breiman, 2001) для анализа возможностей распознавания ключевых биотопов и получения вероятностных оценок их пространственного распределения по территории исследования. В контексте случайных лесов вероятность отнесения объекта к конкретному тематическому классу – это доля деревьев решений от общего числа, в результате классификации по которым этот объект был отнесен к данному классу. При этом оценки вероятностей, полученные этим методом, как правило, являются достаточно объективными без дополнительной калибровки (Kruppa et al., 2014). В качестве переменных при обучении классификационной модели использовались медианные значения спектральных и топографических признаков, рассчитанные в пределах отобранных эталонных участков. Степень информативности каждой из переменных, а также оптимальный состав признаков, позволяющий достигнуть наилучших результатов при классификации, оценивались в процессе автоматизированной процедуры рекурсивного исключения признаков на основе критерия MDA – Mean Decrease in Accuracy (Liaw, Wiener, 2002). Качество классификационной модели, выстроенной с использованием фиксированного набора признаков, оценивалось стандартным для случайных лесов методом out-of-bag (OOB), когда случайная выборка для построения дерева решений генерируется с помощью бутстрепа (т.е. с возвращением элементов), а та часть измерений, которая не попала в эту выборку, используется в качестве контрольных данных. Мы использовали значения OOB-каппы в качестве основного критерия для оптимизации набора переменных, а также для автоматического подбора основных входных параметров для алгоритма классификации. Состав обучающей выборки корректировался с помощью показателей отступа (разность долей от общего числа деревьев, по которым элемент выборки классифицировался правильно и неправильно), определяемых в процессе обучения модели. Элементы исходной выборки со значениями отступа меньше –0.1 были отброшены. В результате объем итоговой обучающей выборки составил 1046 наблюдений.
Вероятностные оценки пространственного распределения ключевых биотопов по территории парка рассчитывались попиксельно на основе оптимального набора наиболее информативных признаков, приведенных к пространственному разрешению 10 метров. Точность результатов вероятностной классификации оценивалась на основе показателя Бриера (Brier, 1950) и величины площади под ROC-кривой (Bradley, 1997), рассчитанных индивидуально для каждого из пяти тематических классов относительно исходных прототипов биотопов, полученных из лесоустроительных материалов. Оптимальные пороговые значения вероятностей для однозначного отнесения пикселей изображения к ключевым биотопам определялись автоматически по критерию минимума суммарного числа ошибок распознавания I и II рода.
Для сравнения результатов классификации с наземными геоботаническими описаниями вычислялись средние значения вероятностей отнесения пикселей к тому или иному биотопу в радиусе 15 м от центра пробной площади (т.е. на площадке 3 × 3 пикселя), после чего весь участок относился к биотопу с максимальным значением средней вероятности.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
По результатам процедуры рекурсивного исключения признаков (рис. 2) мы определили набор из 30 переменных, использование которых позволяет достичь наилучших результатов классификации (рис. 3а). При этом сравнительный анализ информативности не выявил конкретных групп признаков, однозначно дискриминирующих ключевые биотопы, как между собой, так и от всех прочих биотопов на территории исследования.
Рис. 2.
Зависимость формальных показателей точности классификационной модели случайных лесов от числа переменных, использованных в ходе обучения. 1 – OOВ-точность; 2 – ООВ-каппа.
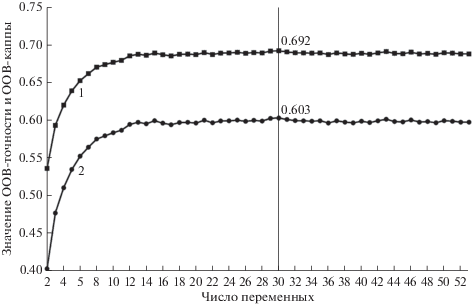
Рис. 3
Информативность спектральных и топографических переменных в процессе обучения классификационной модели случайных лесов (“больше – лучше”). (а) MDA всех переменных (пунктирной линией отделены 30 оптимальных); (б) MDA, осредненное по спектральным признакам; (в) MDA, осредненное по датам съемки; (г) MDA топографических признаков. Обозначения признаков см. табл. 2; Topo* – осредненное MDA всех топографических признаков.
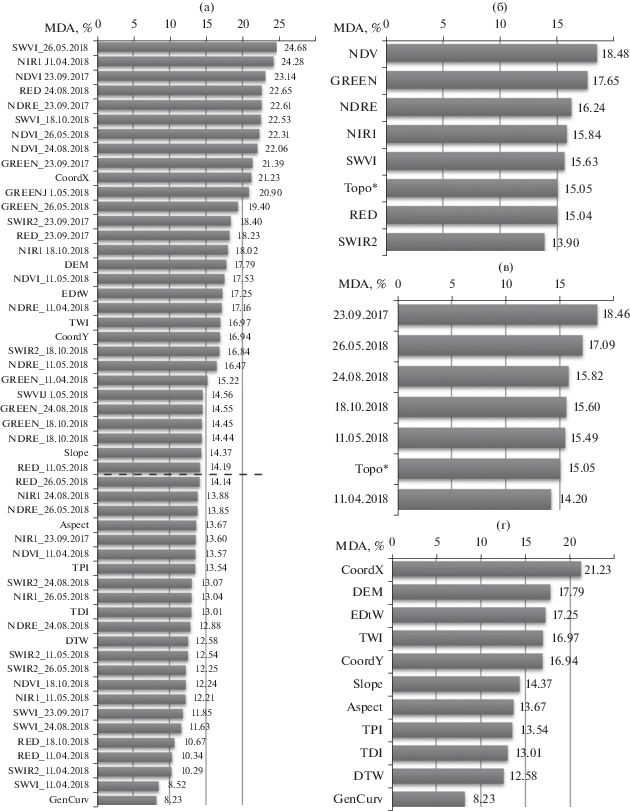
Формально среди спектральных признаков наибольшую информативность продемонстрировали индекс NDVI и значения зеленого канала Sentinel-2, наименьшую – значения второго среднего инфракрасного канала (рис. 3б). Наиболее значимыми периодами съемки для распознавания биотопов оказались конец сентября и конец мая, наименее значимым – начало апреля (рис. 3в). Разница в значениях MDA по группам признаков и по датам съемки относительно невелика (не более 4.6%), что не позволяет единовременно отказаться от всех переменных одного типа или одного периода без потерь в точности классификационной модели.
Из одиннадцати исходных топографических признаков только шесть самых информативных вошли в оптимизированный набор переменных для классификации биотопов – это координаты пикселей по осям X и Y, высота над уровнем моря, расстояние до ближайших водных объектов, топографический индекс влажности и крутизна склонов (рис. 3г). В целом можно отметить явное (но не абсолютное) преимущество в информативности спектральных признаков над топографическими.
На основе оптимизированного набора переменных и скорректированной в процессе обучения эталонной выборки мы получили итоговую модель для вероятностной классификации ключевых биотопов. Общая OOB-точность модели составила 93% при величине OOB-каппы, равной 0.91. Точность распознавания отдельных классов варьирует в пределах от 89 до 96% (табл. 3).
Таблица 3.
OOB-матрица ошибок, полученная в процессе обучения классификационной модели случайных лесов
| №№ п/п |
Лесной биотоп | 1 | 2 | 3 | 4 | 5 | Всего | U_Acc, % | F1, % |
|---|---|---|---|---|---|---|---|---|---|
| 1 | T3-F14 – бореально-неморальные черничные еловые таежные леса | 261 | 3 | 0 | 0 | 11 | 275 | 94.9 | 95.1 |
| 2 | T3-G2 – сосновые и елово-сосновые брусничные таежные леса | 6 | 216 | 0 | 0 | 6 | 228 | 94.7 | 95.6 |
| 3 | T1-E16A – дубово-липовые леса на севере широколиственной и в подтаежной зоне | 0 | 0 | 73 | 1 | 8 | 82 | 89.0 | 90.7 |
| 4 | T1-5114 – подтаежные заболоченные ольховые леса | 1 | 0 | 2 | 187 | 9 | 199 | 94.0 | 94.2 |
| 5 | Все прочие лесные биотопы | 6 | 5 | 4 | 10 | 237 | 262 | 90.5 | 88.9 |
| 6 | Всего | 274 | 224 | 79 | 198 | 271 | 1046 | 92.6 | – |
| 7 | P_Acc, % | 95.3 | 96.4 | 92.4 | 94.4 | 87.5 | 93.2 | 93.1 | – |
Примечания. U_Acc – User’s Accuracy (или Recall) – точность пользователя карты, характеризует величину ошибок I рода; F1 – среднее гармоническое между U_Acc и P_Acc, интегральная мера точности классификации для каждого из тематических классов; P_Acc –Producer’s Accuracy (или Precision) – точность производителя карты, характеризует величину ошибок II рода. Жирным шрифтом отмечена главная диагональ матрицы, показывающая количество случаев совпадения классов производителя и пользователя карты (правильная классификация). Сумма значений диагональных элементов показывает общее количество правильно распознанных случаев (пересечение столбца и строки “Всего”), а их отношение к общему количеству случаев показывает общую точность классификации в процентах.
По результатам классификации были получены четыре тематических изображения территории национального парка и его окрестностей с пространственным разрешением 10 м, характеризующие вероятность отнесения каждого пикселя к одному из четырех типов ключевых лесных биотопов (рис. 4).
Рис. 4.
Вероятностные оценки пространственного распределения ключевых типов лесных биотопов на территории национального парка “Смоленское Поозерье”: бореально-неморальных черничных еловых лесов (T3-F14) (а), сосновых и елово-сосновых брусничных лесов (T3-G2) (б), дубово-липовых лесов (T1-E16A) (в), заболоченных ольховых лесов (T1-5114) (г).
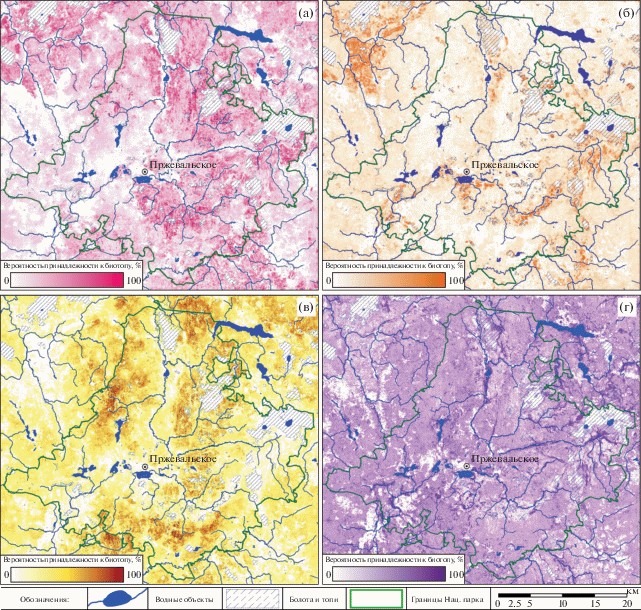
Значения площади под ROC-кривой, определяющие точность вероятностных оценок, для ключевых биотопов находятся в пределах от 88 до 96%, а показатели Бриера не превосходят величины 0.1, что можно интерпретировать как достаточно надежный результат. С использованием оптимальных пороговых значений вероятностей и показателей сомкнутости древостоев по данным GFC мы оценили потенциальные площади, занимаемые ключевыми лесными биотопами модельной территории (табл. 4).
Таблица 4.
Основные характеристики полученных вероятностных оценок пространственного распределения ключевых лесных биотопов на территории национального парка “Смоленское Поозерье"
| №№ п/п |
Лесной биотоп | AUC, % | Brier | Порог вероятности | Площадь, га | Доля от площади лесов национального парка, % | Доля от общей площади национального парка, % |
|---|---|---|---|---|---|---|---|
| 1 | T3-F14 – бореально-неморальные черничные еловые таежные леса | 90.2 | 0.07 | 0.73 | 4555.4 | 4.2 | 3.1 |
| 2 | T3-G2 – сосновые и елово-сосновые брусничные таежные леса | 95.8 | 0.04 | 0.60 | 2094.9 | 1.9 | 1.4 |
| 3 | T1-E16A – дубово-липовые леса на севере широколиственной и в подтаежной зоне | 94.6 | 0.03 | 0.64 | 3726.4 | 3.4 | 2.5 |
| 4 | T1-5114 – подтаежные заболоченные ольховые леса | 88.2 | 0.06 | 0.71 | 4216.5 | 3.9 | 2.8 |
| 5 | Все прочие лесные биотопы | 82.3 | 0.25 | 0.19 | 93 591.4 | 86.6 | 62.9 |
| 6 | Всего | – | – | – | 108 184.6 | 100 | 72.7 |
Примечания. AUC – Area under ROC-curve – доля площади по ROC-кривой (больше – лучше); Brier – показатель Бриера, изменяется от 0 до 1 (меньше – лучше); Порог вероятности – оптимальное значение вероятности, выше которого пиксели изображения были однозначно отнесены к соответствующему ключевому биотопу.
Согласно нашим оценкам, четыре рассматриваемых типа ключевых лесных биотопов суммарно занимают чуть менее 10% от общей площади модельной территории (14.6 тыс. га), или немногим более 13% от покрытой лесом площади. Из них наиболее распространенными являются бореально-неморальные черничные еловые леса, которые занимают 3.1% от площади парка и более характерны для Аржатско-Ельшанской озерно-ледниково-зандровой низины (на восток от р. Ельша), где часто примыкают к крупным болотным массивам. Заболоченные ольховые леса (2.8% территории) приурочены, главным образом, к долинам рек и ручьев, а также бессточным депрессиям на водоразделах, они формируются в условиях застойного увлажнения на торфяных почвах. Дубово-липовые леса занимают 2.5% территории национального парка, их крупные массивы сохранились в пределах Слободской холмисто-моренной возвышенности на западе территории, а также на севере и юге, где они приурочены к возвышенным моренным равнинам с хорошим дренажом и богатыми суглинистыми почвами. Наименее представлены на территории исследования сосновые и елово-сосновые брусничные леса (1.4%), которые приурочены к комплексам озово-камовых образований и повышенным участкам волнистых зандровых равнин.
Сравнение полученных вероятностных оценок с данными наземных обследований (типами биотопов, которые были диагностированы для площадок с геоботаническими описаниями) дало довольно противоречивые результаты (табл. 5). Наилучшая сходимость получена для дубово-липовых лесов (T1-E16A) – лишь одна из десяти площадок была ошибочно отнесена к “прочим биотопам”. В то же время, бореально-неморальные черничные еловые (T3-F14) и сосновые брусничные леса (T3-G2) продемонстрировали довольно низкие показатели сходимости (30 и 40% соответственно), главным образом, за счет ошибочного отнесения к близким к ним по породному составу “прочим биотопам” – мелкотравным (T3-F34) и мезотрофным заболоченным (T3-K) еловым лесам, а также разнотравным (T3-5211) и олиготрофным заболоченным (T3-J) сосновым лесам. Заболоченные ольховые леса, в свою очередь, были корректно распознаны лишь в половине случаев, также из-за перепутывания с “прочими биотопами” – мезофитными мелколиственными лесами (T1-G23, T1-C24, T1-C15).
Таблица 5.
Матрица ошибок, полученная при сравнении результатов тематической классификации с наземными геоботаническими обследованиями
| №№ п/п |
Тип лесных биотопов | T3-F14 | T3-G2 | T1-E16A | T1-5114 | Все прочие | Всего | U_Acc, % | F1, % |
|---|---|---|---|---|---|---|---|---|---|
| 1 | T3-F14– бореально-неморальные черничные еловые таежные леса | 5 | 3 | 0 | 0 | 1 | 9 | 55.6 | 30.3 |
| 2 | T3-G2 – сосновые и елово-сосновые брусничные таежные леса | 0 | 6 | 0 | 0 | 0 | 6 | 100.0 | 40.0 |
| 3 | T1-E16A – дубово-липовые леса на севере широколиственной и в подтаежной зоне | 0 | 0 | 9 | 0 | 1 | 10 | 90.0 | 94.7 |
| 4 | T1-5114–подтаежные заболоченные ольховые леса | 0 | 0 | 0 | 4 | 4 | 8 | 50.0 | 50.0 |
| 5 | Все прочие лесные биотопы, в том числе: |
19 | 15 | 0 | 4 | 112 | 150 | 74.7 | 83.6 |
| 6 | T1-G23 – мезофитные ольховые леса | 0 | 0 | 0 | 2 | 32 | 34 | 94.1 | – |
| 7 | T1-C24 – мезофитные осиновые леса | 0 | 0 | 0 | 1 | 11 | 12 | 91.7 | – |
| 8 | T1-61–мезотрофные заболоченные березовые леса | 0 | 0 | 0 | 0 | 4 | 4 | 100.0 | – |
| 9 | T1-C15 – мезофитные березовые леса | 1 | 0 | 0 | 1 | 45 | 47 | 95.7 | – |
| 10 | T3-5211 – таежные разнотравные сосновые леса | 1 | 10 | 0 | 0 | 11 | 22 | 50.0 | – |
| 11 | T3-J–олиготрофные заболоченные сосновые леса | 0 | 3 | 0 | 0 | 2 | 5 | 40.0 | – |
| 12 | T3-F34 – таежные мелкотравные еловые леса | 14 | 2 | 0 | 0 | 6 | 22 | 27.3 | – |
| 13 | T3-K – мезотрофные заболоченные еловые леса | 3 | 0 | 0 | 0 | 1 | 4 | 25.0 | – |
| 14 | Всего | 24 | 24 | 9 | 8 | 118 | 183 | 74.1 | – |
| 15 | P_Acc, % | 20.8 | 25.0 | 100.0 | 50.0 | 94.9 | 58.1 | 74.3 | – |
Примечание. U_Acc – User’s Accuracy (или Recall) – точность пользователя карты, характеризует величину ошибок I рода; F1 – среднее гармоническое между U_Acc и P_Acc, интегральная мера точности классификации для каждого из тематических классов; P_Acc –Producer’s Accuracy (или Precision) – точность производителя карты, характеризует величину ошибок II рода. Жирным шрифтом отмечена главная диагональ матрицы, показывающая количество случаев совпадения классов производителя и пользователя карты (правильная классификация). Сумма значений диагональных элементов показывает общее количество правильно распознанных случаев (пересечение столбца и строки “Всего”), а их отношение к общему количеству случаев показывает общую точность классификации в процентах.
Возможными причинами выявленных несоответствий могут являться следующие факторы:
использованные спектральные и топографические признаки недостаточны для надежного распознавания лесных биотопов близких по породному составу древостоев; необходимо исследовать возможность применения почвенных характеристик (тип почвы, плодородия и увлажнения) для повышения уровня разделимости;
критерии применяемой классификации EUNIS допускают смешанный (разновидовой) состав древостоя в ключевых лесных биотопах, выбранных для анализа (табл. 1), и при этом для многих лесных сообществ модельной территории именно такой состав очень характерен в результате многовековой хозяйственной деятельности, которая была особенно интенсивной в ХХ в., когда сформировалось подавляющее большинство современных древостоев; для картографирования биотопов на территории с такой сложной историей требуется больше данных, полученных в ходе наземных геоботанических исследований;
большинство геоботанических площадок было заложено для решения различных научно-исследовательских задач, в число которых, тем не менее, изначально не входила валидация результатов геопространственного моделирования распределения биотопов на основе спутниковых и топографических данных; для целей нашего исследования при планировании геоботанического обследования территории необходимо учитывать пространственное разрешение (масштаб) используемых спутниковых изображений и картографических материалов.
Заключение. Использованный в работе подход, основанный на совместном анализе спектральных и топографических признаков, оправдал себя лишь отчасти. Нам удалось с помощью разносезонных спутниковых изображений добиться высокой точности распознавания ключевых лесных биотопов друг относительно друга благодаря их принципиальным различиям в породном составе древостоев. При распознавании ключевых биотопов относительно прочих, близких к ним по породному составу, но различных по условиям местопроизрастания и соответственно типам напочвенного покрова, получить надежный результат нам не удалось. Теоретически дискриминацию по условиям местопроизрастания должны были обеспечить топографические признаки, но, судя по всему, состав и/или детальность исходных данных, использованных нами в работе, оказались недостаточными для решения поставленной задачи. Наилучшие результаты при обучении классификационной модели случайных лесов были достигнуты при одновременном использовании 30 переменных, 80% из которых приходится на спектральные признаки. Кроме того, нам не удалось обнаружить как единственно необходимых, так и абсолютно бесполезных спектральных каналов и индексов, а также периодов съемки среди исходного набора переменных. Это свидетельствует как об относительно низкой информативности исходных топографических данных, так и об очевидном преимуществе временных серий мультиспектральных спутниковых изображений над одномоментными наблюдениями.
Анализ точности полученных нами геопространственных оценок приводит к противоречивым выводам. С одной стороны, мы использовали данные лесоустройства как наиболее полного источника информации о лесах модельной территории, для обучения классификационной модели. Формальные показатели точности распознавания отдельных классов, а также объективности полученных вероятностных оценок показали достаточно высокие результаты. С другой стороны, значительная по объему контрольная выборка из наземных обследований в целом продемонстрировала невысокую сходимость с вероятностными оценками. Оставив за скобками достоверность обоих источников априорной информации, можно заключить что, возможной причиной данной ситуации является недостаточная представительность имеющейся сети площадок геоботанических описаний в силу специфики их исходного назначения и пространственного распределения по территории парка. Очевидно, что для достоверной оценки точности геопространственного моделирования сеть контрольных пробных площадей должна быть спроектирована с учетом особенностей исходных данных (прежде всего, это пространственное разрешение) и полученных результатов (распределения вероятностей тематических классов по территории).
Таким образом, наши дальнейшие исследования будут направлены, во-первых, на выявление дополнительных признаков, которые могут повысить точность распознавания близких по породному составу древостоев лесных биотопов, и, во-вторых, на разработку методики проектирования сети наземных обследований, способной дать объективную оценку точности тематических продуктов, получаемых на основе спутниковых, топографических и других геопространственных данных.
***
Авторы статьи благодарят сотрудников национального парка “Смоленское Поозерье” – директора Александра Семёновича Кочергина и начальника отдела инвентаризации и мониторинга природных комплексов Владимира Рафаэльевича Хохрякова за оказанную помощь и предоставление необходимого материала для выполнения научных исследований в рамках представленной тематики статьи.
Список литературы
Браславская Т.Ю. Широколиственные и черноольховые леса Европейской России как объекты в составе экологической сети // Географические основы формирования экологических сетей в Северной Евразии. Т. 6. Матер. 6-й междунар. науч. конф. (г. Тверь, 8–10 ноября 2016 г). М.: Институт географии РАН, 2016. С. 18–23.
Браславская Т.Ю. О природоохранной ценности восточноевропейских широколиственных лесов // Вестник Тверского гос. университета. Серия “Биология и экология”. 2017. № 2. С. 278–286.
Браславская Т.Ю., Тихонова Е.В., Гаврилюк Е.А., Бавшин И.М., Ершов Д.В. Опыт применения классификации местообитаний EUNIS в средней полосе Европейской России на примере национального парка “Смоленское Поозерье” // Научные основы устойчивого управления лесами: Матер. III Всеросс. (с междунар. участием) науч. конф. “Научные основы устойчивого управления лесами” (г. Москва, 30 октября–1 ноября 2018 г.). М.: ЦЭПЛ РАН, 2018. С. 24–26.
Изумрудная книга Российской Федерации. Территории особого природоохранного значения Европейской России. Предложения по выявлению. М.: Институт географии РАН, 2011–2013. Ч. 1. 308 с.
Изумрудная сеть территорий особого природоохранного значения. Руководство для органов государственной власти субъектов Российской Федерации, дирекций особо охраняемых природных территорий и органов местного самоуправления / Сост. Н.А. Соболев, Н.М. Алексеева, Е.С. Пушай. М.; СПб: Институт географии РАН, 2015. 48 с.
Королева Н.Е. Основные биотопы северо-таежных лесов и березовых криволесий Мурманской области: ландшафтное и ботаническое разнообразие, основания для охраны // Вестник Мурманского гос. технич. университета. 2011. Т. 14. № 4. С. 819–832.
Крицук С.Г. Картирование бореальных лесов на основе спутниковых данных (на примере особо охраняемых природных территорий Ленинградской области) // Современные проблемы дистанционного зондирования Земли из космоса. 2012. Т. 9. № 4. С. 255–264.
Крышень А.М., Полевой А.В., Гнатюк Е.П., Кравченко А.В., Кузнецов О.Л. База данных местообитаний (биотопов) Карелии // Труды Карельского научного центра РАН. 2009. № 4. Сер. Биогеография. Вып. 9. С. 3–10.
Лесохозяйственный регламент национального парка “Смоленское Поозерье”. М.: Филиал ФГУП “Рослесинфорг” “Заплеспроект”, 2015. 190 с.
Методические подходы к экологической оценке лесного покрова в бассейне малой реки / Отв. ред. Заугольнова Л.Б. и Браславская Т.Ю. М.: Товарищество научных изданий КМК, 2010. 240 с.
Научные исследования в национальном парке “Смоленское Поозерье” / Под ред. Волкова С.М. М.: НИА-Природа, 2003. Вып. 1. 295 с.
Постановление Правительства Российской Федерации от 15 апреля 1992 г. № 247 “О создании национального парка "Смоленское Поозерье” Министерства экологии и природных ресурсов Российской Федерации в Смоленской области". [Электронный ресурс]. URL: https://base.garant.ru/70252316/ (дата обращения 12.03.2019).
Пугачевский А.В., Вершицкая И.Н., Ермохин М.В., Ермохин М.В., Степанович И.М., Созинов О.В., Сакович А.А., Рудаковский И.А., Кулак А.В., Журавлев Д.В. Редкие биотопы Беларуси. Минск: Альтиора–Живые краски, 2013. 236 с.
Растительность и почвы национального парка “Смоленское Поозерье” / Под ред. Копцик Г.Н., Березиной Н.А. М.: НИА-Природа, 2003. 307 с.
Шкаликов В.А. Характеристика рельефа // Изучение состояния природы Смоленского Поозерья в целях охраны и рационального использования на момент организации в этом регионе национального природного парка (итоговый отчет за 1991-1995 гг.). Смоленск. 1995. С. 49–53.
Шкаликов В.А., Ерашов М.А., Борисовская И.А. Особо охраняемые природные территории Смоленской области. Смоленск: Универсум, 2005. 464 с.
Álvarez-Martínez J.M., Jiménez-Alfaro B., Barquín J., Ondiviela B., Recio M., Silió-Calzada A., Juanes J.A. Modelling the area of occupancy of habitat types with remote sensing // Methods in Ecology & Evolution. 2018. V. 9(3). P. 580–593.
Annex I: natural habitat types of community interest whose conservation requires the designation of special areas of conservation // Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora – consolidated version 01/01/2007. [Электронный ресурс]. URL::http://eur-lex.europa.eu/LexUriServ/ LexUriServ.do?uri=CONSLEG:1992L0043:20070101:EN: PDF (дата обращения 12.03.2019).
Berger A.L., Pietra S.A.D., Pietra V.J.D. A maximum entropy approach to natural language processing. Computational Linguistics. 1996. № 22. P. 39–71.
Beven K.J., Kirkby M.J. A physically-based variable contributing area model of basin hydrology // Hydrology Science Bulletin. 1979. V. 24. № 1. P. 43–69.
Boehner J., Antonic O. Land Surface Parameters Specific to Topo-Climatology // Developments in Soil Science. 2009. V. 33. P. 195–226.
Bradley A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms // Pattern Recognition. 1997. V. 30. № 7. P. 1145–1159.
Breiman L. Random forests // Machine Learning. 2001. V. 45. № 1. P. 5–32.
Brier G.W. Verification of forecasts expressed in terms of probability // Monthly Weather Review. 1950. V. 78. № 1. P. 1–3.
Corbane C., Lang S., Pipkins K., Alleaume S., Deshayes M., García Millán V.E., Strasser T., Vanden Borre J., Toon S., Förster M. Remote sensing for mapping natural habitats and their conservation status – New opportunities and challenges // International Journal of Applied Earth Observation & Geoinformation. 2015. V. 37. P. 7–16.
Davies C.E., Moss D., Hill M.O. EUNIS Habitat Classification revised 2004. Report to the European Topic Centre on Nature Protection and Biodiversity, European Environment Agency. Huntingdon: Institute of Terrestrial Ecology, 2004. 307 p.
Díaz Varela R.A., Ramil Rego P., Calvo Iglesias S., Muñoz Sobrino C. Automatic habitat classification methods based on satellite images: A practical assessment in the NW Iberia coastal mountains // Environmental Monitoring & Assessment. 2008. № 144. P. 229–250. https://doi.org/10.1007/s10661-007-9981-y
ESA Sentinel-2 [Электронный ресурс]. URL: http:// www.esa.int/Our_Activities/Observing_the_Earth/Copernicus/Sentinel-2 (дата обращения 12.03.2019).
EUNIS habitat type hierarchical view [Электронный ресурс]. URL: http://eunis.eea.europa.eu/habitats-code-browser.jsp (дата обращения 12.03.2019).
Evans D. Interpreting the habitats of Annex I – Past, present and future // Acta Botanica Gallica. 2010. V. 157. № 4. P. 677–686.
Gao B.C. NDWI – A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water from Space // Remote Sensing of Environment. 1996. V. 58. № 3. P. 257–266.
Guisan A., Weiss S.B., Weiss A.D. GLM versus CCA spatial modeling of plant species distribution // Plant Ecology. 1999. V. 143. № 1. P. 107–122.
Hagolle O., Huc M., Villa Pascual D., Dedieu G. A multi-temporal and multi-spectral method to estimate aerosol optical thickness over land, for the atmospheric correction of FORMOSAT-2, LANDSAT, VENS and SENTINEL-2 images // Remote Sensing. 2015. V. 7. № 3. P. 2668–2691.
Hansen A.J., De Fries R., Turner W. Land use change and biodiversity: a synthesis of rates and consequences during the period of satellite imagery // Land change science: Observing, monitoring, and understanding trajectories of change on the earth’s surface. N.Y.: Kluwer Academic Publishers, 2004. P. 277–299.
Hansen M.C., Potapov P.V., Moore R., Hancher M., Turubanova S.A., Tyukavina A., Thau D., Stehman S.V., Goetz S.J., Loveland T.R., Kommareddy A., Egorov A., Chini L., Justice C.O., Townshend J.R.G. High-Resolution Global Maps of 21st-Century Forest Cover Change // Science. 2013. V. 342. P. 850–853.
Kruppa J., Liu Y., Biau G., Kohler M., König I.R., Malley J.D., Ziegler A. Probability estimation with machine learning methods for dichotomous and multicategory outcome: theory // Biometrical J. 2014. V. 56. № 4. P. 534–563.
Kushwah K., Rao R.J., Phurailatpam B. Geo-Spatial Modelling of Habitat Suitability of wildlife species of Kuno Wildlife Sanctuary // IOSR J. Environmental Science, Toxicology & Food Technology. V. 1. Is. 4. P. 12–19.
Liaw A., Wiener M. Classification and Regression by random forest // R News. 2002. V. 2. № 3. P. 18–22.
Mücher C.A., Hennekens S.M., Bunce R.G.H., Schaminée J.H.J., Schaepman M.E. Modelling the spatial distribution of Natura 2000 habitats across Europe // Landscape & Urban Planning. 2009. V. 92. P. 148–159.
Mücher C.A., Hennekens S.M., Schaminée J., Halada L., Halabuk A. Modelling the spatial distribution of EUNIS forest habitats based on vegetation relevés and Copernicus HRL. ETC/BD report to the EEA. 2015. 48 p.
Murphy P., Ogilvie J., Arp P. Topographic modelling of soil moisture conditions: a comparison and verification of two models // European Journal of Soil Science. 2009. V. 60(1). P. 94–109.
Nagendra H., Lucas R., Honrado J.P., Jongman R.H., Tarantino C., Adamo M. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats // Ecological Indicators. 2013. V. 33. P. 45–59.
Open Street Map [Электронный ресурс]. URL: https:// www.openstreetmap.org (дата обращения 12.03.2019).
Phillips S.J., Anderson R.P., Schapire R.E. Maximum entropy modelling of species geographic distributions // Ecological Modelling. 2006. № 190. P. 231–259.
Redding N.J., Crisp D.J., Tang D., Newsam G.N. An efficient algorithm for Mumford-Shah segmentation and its application to SAR imagery // Proc. Conf. “Digital Image Computing: Techniques & Applications” (DICTA-99), Perth, Australia, 1999. P. 35–41.
Revised Annex I of Resolution 4 (1996) of the Bern Convention on endangered natural habitats types using the EUNI-S habitat classification (year of revision 2014). [Электронный ресурс]. Council of Europe, 2010-2014. URL: https://wcd.coe.int/ViewDoc.jsp?Ref=T-PVS/PA (2010)10&Language=lanEnglish&Ver=rev&Site=DG4-Nature&BackColorInternet=DBDCF2&BackColorIntranet= FDC864&BackColorLogged=FDC864 (дата обращения 12.03.2019).
Rouse J.R., Haas J.S., Deering D. Monitoring Vegetation Systems in the Great Plains with ERTS // Goddard Space Flight Center 3d ERTS-1 Symp., NASA, 1974. V. 1. Sect. A. P. 309–317.
Schaminée J.H.J., Chytrý M., Hennekens S.M., Jiménez-Alfaro B., Mucina L., Rodwell J.S., Tichý L. Review of EUNIS forest habitat classification. Report to the European Environmental Agency (EEA/NSV/13/005). Alterra WUR, Wageningen, the Netherlands. 2013. 111 p.
Schaminée J.H.J., Chytrý M., Hennekens S.M., Janssen J.A.M., Jiménez-Alfaro B., Knollová I., Mucina L., Rodwell J.S., Tichý L. Vegetation analysis and distribution maps for EUNIS habitats. Report to the European Environmental Agency (EEA/NSV/14/006). Alterra WUR, Wageningen, the Netherlands. 2014. 175 p.
Shary P.A. Land surface in gravity points classification by complete system of curvatures // Mathematical Geology. 1995. V. 27. № 3. P. 373–390.
Sims D., Gamon J. Relationships Between Leaf Pigment Content and Spectral Reflectance Across a Wide Range of Species, Leaf Structures and Developmental Stages // Remote Sensing of Environment. 2002. V. 81. P. 337–354.
Tagil S. Effect of Topographic Habitat Characteristics on the Spatial Distribution of Landuse-Landcover in the Kapidag Peninsula, Turkey // J. Applied Sciences. 2015. V. 15(6). P. 850–861.
Terrestrial habitat mapping in Europe: an overview // Luxembourg: Publications Office of the European Union, 2014. 152 p.
Travis M.R., Elsner G.H., Iverson W.D., Johnson C.G. VIEWIT: computation of seen areas, slope, and aspect for land-use planning / Gen. Tech. Rep. PSW-GTR-11. Berkeley, CA: Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture. 1975. 70 p.
Дополнительные материалы отсутствуют.