

Лесоведение, 2020, № 2, стр. 162-174
Диагностика физиологического состояния хвойных деревьев по биоэлектрическим и температурным показателям
В. Н. Карасев a, М. А. Карасева a, Д. И. Мухортов a, *
a Поволжский государственный технологический университет
424000 Йошкар-Ола, пл. Ленина, 3, Россия
* E-mail: MuhortovDI@volgatech.net
Поступила в редакцию 24.01.2018
После доработки 05.04.2018
Принята к публикации 08.10.2019
Аннотация
Проводились исследования области применения биоэлектрических потенциалов (БЭП), импеданса прикамбиального комплекса и температурных параметров стволов деревьев для диагностики физиологического состояния хвойных древесных пород в природной среде. Установлено, что величины БЭП в пределах 180–210 мВ имеют лидирующие деревья, у ослабленных деревьев параметры БЭП уменьшаются до 60–70 мВ. Показатели импеданса прикамбиального комплекса тканей у здоровых деревьев имеют значения 20–30 кОм, при ухудшении жизненного состояния параметры возрастают в 2–3 раза. Установлена связь температуры стволов с физиологическим состоянием деревьев (r = –0.9). Предложена методика определения расхода воды деревом по линейной скорости водного тока и объему проходящей воды через единицу водопроводящей ксилемы. Установлена связь температуры стволов деревьев с расходом воды и их жизненным состоянием. Для здоровых деревьев температура стволов на высоте 1.3 м близка к температуре корнеобитаемых горизонтов почвы, ослабленные деревья имеют более высокие значения. Приведены примеры, показывающие целесообразность применения биоэлектрических показателей при отборе устойчивых высокопродуктивных биотипов и семенных деревьев и оценке жизненного состояния деревьев в рекреационных зонах. Рекомендована экспресс–диагностика физиологического состояния по биоэлектрическим и температурным параметрам.
В связи с глобальным изменением климата, антропогенным воздействием на лесные экосистемы ведение мониторинга физиологического состояния компонентов лесных фитоценозов является актуальной проблемой. Основными показателями устойчивости лесной экосистемы является интенсивность роста и продуктивность основных лесообразующих хвойных видов. В условиях усиливающего стрессового климатического и техногенного воздействия необходимы показатели, позволяющие проводить раннюю экспресс–диагностику физиологического состояния лесных фитоценозов на начальных этапах их ослабления.
Применяемая в практических целях Европейская шкала оценки состояния насаждений учитывает категории ослабления деревьев по внешнему виду листьев и состоянию кроны, но для древесных растений с многолетним циклом развития, эта шкала имеет малую прогнозную информативность (Алексеев, 1989; Критерии …, 1995). Индикаторами устойчивости растений к климатическим и техногенным стрессам являются морфометрические показатели (Иванов и др., 2015), параметры водного режима (Карасев, Карасева, 2004; Кудоярова и др., 2013), интенсивность фотосинтеза, содержание хлорофилла в хвое (Воробьев, 2013; Софронова и др., 2016), биохимические показатели, активность ферментов (Шевякова, 1983; Судачкова и др., 2003). При мониторинге состояния деревьев по биохимическим показателям отмечена значительная индивидуальная и эндогенная изменчивость параметров и необходимость учитывать специфичность тканей при выборе тест–объектов для диагностики.
Исследованиями установлена (Рутковский, 1975; Коловский, 1980; Карасев, 2001; Oyarce, Gurovich, 2011; Borges et al., 2014; Rnos–Rojasa et al., 2014) возможность оценки физиологического состояния растений по биоэлектрическим параметрам на уровне целостного организма. Представления о распространении электрических сигналов в растениях, их свойствах и функциональной роли изложены в работах В.А. Опритова с соавторами (1991) и С.С. Пятыгина (2008). Биоэлектрические показатели связаны с интенсивностью и характером обмена веществ, параметрами водного режима, интенсивностью роста и характеризуют адаптивные свойства растений при действии неблагоприятных факторов внешней среды. Известно, что физиологическое состояние растений, характер обмена веществ в тканях и величины полного электрического сопротивления прикамбиального комплекса тканей (импеданс ПКТ) взаимосвязаны. При снижении уровня жизнедеятельности импеданс ПКТ существенно увеличивается, а более низкие значения этого параметра свойственны деревьям лучшего состояния, но в ряде ситуаций требуется уточнение информативности показателей для диагностики состояния с учетом типа повреждения или ослабления растения.
Также при некоторых повреждениях показатели биоэлектрических потенциалов (БЭП), характеризующие интенсивность обменных процессов, не всегда позволяют правильно оценить жизненное состояние деревьев, в связи с этим необходимы дальнейшие исследования, уточняющие области применения электрофизиологических параметров для экспресс–диагностики деревьев.
Различные повреждения растений и нарушения их физиологических функций тесно связаны с интенсивностью водообмена, однако быстрое и точное получение количественных показателей скорости водного тока и расхода воды деревом являются методически трудной задачей. Более доступно определение температурных параметров прикамбиального комплекса тканей стволов деревьев, сопряженное с состоянием водного режима растений. Температура растений хорошо изучена, исследовано изменение температуры растений в различных климатических условиях, но недостаточно информации о связи температуры стволов деревьев с расходом воды деревом и их жизнеспособностью.
Цель работы – оценка информативности и особенностей применения БЭП, импеданса ПКТ и температурных показателей стволов деревьев для экспресс-диагностики физиологического состояния хвойных древесных пород на ранних этапах их ослабления при различных типах ослаблений и повреждений в природной среде.
ОБЪЕКТЫ И МЕТОДИКА
Изучение физиологического состояния основных лесообразующих хвойных видов проводили в лесных фитоценозах района хвойно-широколиственных лесов Среднего Поволжья. Объектами исследований являлись аборигенные Pinus sylvestris (L.), Picea abies (L.) Karst., и интродуцированные виды – Larix sibirica Ledeb., Pinus sibirica Du Tour, произрастающие в естественных насаждениях и производственных культурах Среднего Поволжья и на опытных стационарах Учебно-опытного лесхоза Поволжского государственного технологического университета (УОЛ ПГТУ).
Исследования интенсивности фотосинтеза, дыхания хвойных пород и БЭП проводили на опытных участках в одновозрастных несомкнувшихся культурах УОЛ ПГТУ, созданных в условиях свежей сурамени. Почвы участков лесных культур дерново-среднеподзолистые суглинистые на маломощных покровных суглинках, подстилаемых древнеаллювиальными песками. Содержание гумуса составляет 2.2%, кислотность почвы равна 4.4. По содержанию подвижных форм фосфора и калия по шкале Кирсанова почвы имеют среднюю и низкую степень обеспеченности. Плотность сложения почвы на глубине 0–25 см составляет 1.11 г см–3, на 25–50 см – 1.4 г см–3. Рельеф ровный.
Оценку индивидуальной изменчивости основных физиологических процессов древесных растений проводили на лесосеменных участках и плантациях Вятско-Полянского спецсемлехоза Кировской области. Тип лесорастительных условий на объекте – свежая суборь, содержание гумуса в корнеобитаемых горизонтах почвы 2.4%, возраст семенных деревьев 30 лет, средняя высота семенных деревьев 13.5 м, средний диаметр 16 см, протяженность кроны 8.8 м.
Оценку информативности температурных параметров как одного из физиологических показателей состояния древесных растений при длительном действии неблагоприятных факторов среды проводили в УОЛ ПГТУ в насаждениях сосны обыкновенной, ослабленных в результате периодического избыточного увлажнения. Объект исследования – сосновое насаждение естественного происхождения, подверженное временному периодическому избыточному увлажнению в весенне-летний период из-за нарушения поверхностного стока. Состав 9С1Е. Средняя высота 19 м, средний диаметр 24 см, возраст 60 лет. Тип леса – сосняк черничник. Рельеф ровный с микроповышениями высотой 0.5–0.7 м, которые в весенне-летний период не затапливались.
Район исследований характеризуется умеренно континентальным климатом, свежими слабоподзолистыми почвами, средним количеством осадков 481 мм в год и среднегодовой температурой воздуха 2.8°С. Продолжительность вегетационного периода составляет 120 дней. Лесные экосистемы Среднего Поволжья постоянно испытывают не только стрессовые климатические воздействия (периодически повторяющиеся засухи, избыточное увлажнение, сильные морозы), но и значительное рекреационное и техногенное воздействие, поэтому сохранение жизнеспособности и устойчивости лесных насаждений имеет важное экологическое значение.
Для выявления наиболее информативных диагностических параметров жизненного состояния деревьев проводили исследования интенсивности фотосинтеза, транспирации, определяли содержание общего хлорофилла, размеры хвои, измеряли БЭП, импеданс ПКТ стволов деревьев, определяли влажность хвои и побегов, скорость водного тока, расход воды деревом, температурные параметры стволов. Данные показатели изучали у аборигенных пород; сосны обыкновенной, ели европейской и перспективных интродуцентов; лиственницы сибирской, сосны кедровой сибирской. При оценке состояния по биоэлектрическим параметрам на каждой пробной площади проводили не менее 100 измерений с разделением на группы; лидирующие, средние, ослабленные.
При экспресс-оценке по температурным параметрам стволов жизненное состояние растений оценивали по методике В.А. Алексеева (1989) c некоторыми изменениями. Деревья по состоянию подразделяли на группы: 1 – здоровые; 2 – ослабленные; 3 – сильно ослабленные; 4 – усыхающие. Каждая группа состояния была представлена десятью деревьями. Состояние деревьев в пределах каждой группы оценивали комплексом признаков: массой сохранившейся хвои, ее цветом, скоростью водного тока, интенсивностью транспирации, расходом воды деревом, линейными и весовыми параметрами, влажностью древесины и хвои. После предварительного исследования физиологических параметров осуществляли распределение деревьев по группам и на этой основе выявляли информативную ценность методов экспресс–диагностики их жизненного состояния. При предварительном разделении деревьев на группы учитывали состояние кроны с учетом параметров, рекомендованных Европейской шкалой оценки состояния деревьев по классам повреждения (Критерии …, 1995). Определение массы сырой хвои проводили способом сплошного учета по пяти модельным деревьям каждой категории жизнеспособности.
Влажность прикамбиального комплекса тканей оценивали по образцам, взятым на высоте 1.3 м у деревьев с одинаковыми диаметрами каждой категории состояния.
Для определения расхода воды деревом применяли две различные методики: 1) расход воды рассчитывали по данным массы хвои на дереве и интенсивности транспирации; 2) определяли линейную скорость водного тока и содержание подвижной воды в единице объема водопроводящей ксилемы с учетом площади водопроводящей зоны ствола дерева. Показатели расхода воды деревом, полученные по описанным методикам, существенных различий не имели.
Расчет транспирации дерева проводился на отдельно стоящих деревьях с измерением интенсивности транспирации в верхней, средней и нижней частях кроны с учетом ее экспозиции и последующим определением массы хвои на всем дереве. Работа проводилась для сравнения с предлагаемой методикой определения расхода воды деревом по скорости водного тока и объема прохождения воды через единицу сечения водопроводящей ксилемы.
Для определения расхода воды деревом применяли разработанную нами методику определения линейной скорости водного тока и содержания подвижной воды в единице объема водопроводящей ксилемы с учетом площади водопроводящей зоны ствола дерева. Для определения линейной скорости водного тока использовали кондуктометрический экспресс-метод (Карасев, 2001). В ксилему водопроводящей зоны ствола, ветвей вводили растворы пробных веществ – электролитов и контролировали их продвижение вместе с водным током по изменению электрического сопротивления водопроводящей ксилемы. Возрастание электропроводимости ксилемы и падение электрического сопротивления по сравнению с естественным фоном происходит скачкообразно под влиянием транспортируемых электрически заряженных частиц – анионов и катионов пробного вещества. Линейную скорость водного тока по ксилеме определяли как отношение расстояния между линиями “старт” и “финиш” к времени, разделяющему возникновение скачкообразных падений электрического сопротивления водопроводящей ксилемы.
Измерение линейной скорости водного тока и количество подвижной воды в единице объема ксилемы производили в четырех точках средней части водопроводящей зоны ствола, ориентированных по сторонам света, на высоте 1.3 м.
Интенсивность фотосинтеза определяли кондуктометрическим способом (Карасев, 2001). Измерения проводили на пяти модельных деревьях каждого вида, в трехкратной повторности в средней части кроны на неотделенных побегах. Относительное световое довольствие растений оценивали в % от освещенности открытого места, измерялись биометрические показатели деревьев и параметры внешней среды, температура и влажность воздуха и почвы, скорость ветра. Данные, полученные в каждом вегетационном периоде, обрабатывались с применением методов математической статистики. Показатели, приведенные в табл. 1, получены на основе статистической обработки средних данных измерений за 10 вегетационных периодов в 11–12 ч дня в период полуденной депрессии фотосинтеза на одних и тех же объектах исследований, что позволяет провести сравнительную оценку адаптации аборигенных и интродуцированных хвойных видов к климатическим стрессам. Для получения сравнительных количественных показателей интенсивности фотосинтеза и дыхания проводили измерения также и в наиболее благоприятный для фотосинтеза период (9–10 ч утра). Определение содержания общего хлорофилла в хвое проводили фотоэлектроколориметрическим методом с применением прибора КФК-2 в красной области спектра. Расшифровка показателей осуществлялась по таблицам, составленным Т.Н. Годневым (1952). Для сравнительных исследований образцы хвои для установления содержания в них общего хлорофилла брали в средней части кроны южной экспозиции, с побегов, на которых определялась интенсивность фотосинтеза.
Таблица 1.
Интенсивность фотосинтеза, дыхания и содержание хлорофилла однолетней хвои хвойных древесных видов с мая по июль на 11–12 ч дня
| Вид | Интенсивность фотосинтеза, мг СО 2 (г ч)–1 на сухую массу хвои |
Дыхание, мг СО 2 (г ч)–1 на сухую массу хвои | Содержание общего хлорофилла, мг г–1 сух. хвои | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Х | Мx | V, % | Х | Мx | V, % | Х | Мx | V, % | |
| Сосна обыкновенная | 1.12 | 0.09 | 34.6 | 0.85 | 0.15 | 79.2 | 1.88 | 0.048 | 5.8 |
| Ель европейская | 1.09 | 0.18 | 67.0 | 0.75 | 0.09 | 61.2 | 3.91 | 0.060 | 3.1 |
| Сосна кедровая сибирская | 1.05 | 0.16 | 89.0 | 0.70 | 0.15 | 106.9 | 2.46 | 0.170 | 17.3 |
| Лиственница сибирская | 3.00 | 0.29 | 55.7 | 1.90 | 0.31 | 105.6 | 3.75 | 0.161 | 12.9 |
Примечание в табл. 1, 3–5 . X – среднее значение признака; Мx – ошибка среднего; V – коэффициент изменчивости.
Измерения БЭП производили милливольтметром постоянного тока с высоким входным сопротивлением (1–3 ГОм) с использованием игольчатых платиновых электродов (Экстра–999). Хотя рядом исследователей для отведения БЭП рекомендуется использование неполяризующихся электродов (Коловский, 1980), при ведении мониторинга физиологического состояния деревьев для сравнительных исследований более удобны и надежны в эксплуатации в полевых условиях поляризующие платиновые электроды. Абсолютная погрешность измерительного процесса в диапазоне измерений БЭП до 90 мВ составляла ±4.67 мВ, до 250 мВ – ±14 мВ.
Измерения электрического сопротивления ПКТ, проводили прибором Ц-4314 на частоте 500 Гц, использовали датчик от электронного влагомера древесины ЭВ–2К с двумя боковыми электродами, подключаемыми к прибору для измерения импеданса, центральный электрод удаляли. Электроды вводили с северной стороны стволов на высоте 1.3 м. Длина активной части электродов 10.0 мм, диаметр – 1.0 мм. Расстояние между электродами 20.0 мм. В диапазоне измерений до 100 кОм абсолютная суммарная погрешность параметров импеданса составляла ±4.96 кОм. Использование для измерений ПКТ, включающего луб, камбий и последние слои водопроводящей зоны, обусловлено тем, что они являются наиболее активными образовательными тканями, связанными с процессами жизнедеятельности растений. При электрофизиологических измерениях предпочтительнее биопотенциалы и импеданс отводить от этой зоны.
Измерение температуры стволов деревьев производилось на высоте 1.3 м в точках, обращенных к северу и защищенных от прямого воздействия солнечной радиации. Для измерения температуры стволов деревьев использовался цифровой датчик DS18B20 и цифровой микропроцессорный термометр ВМ8037.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Важное значение для получения информации о состоянии хвойных видов, степени адаптации их к воздействию неблагоприятных факторов внешней среды имеют комплексные эколого-физиологические исследования непосредственно в естественной среде их обитания. Фотосинтетическая активность и водный режим являются показателями, характеризующими интенсивность роста и устойчивость древесных растений к климатическим стрессам в лесорастительных условиях Среднего Поволжья, вызвавшими деградацию дубрав и массовое усыхание ельников.
Сравнительные средние, многолетние показатели, полученные за 10-летний период, характеризующие интенсивность фотосинтеза и дыхания хвойных пород в условиях Среднего Поволжья в 11–12 ч дня в середине вегетационного периода, приведены в табл. 1. В полуденные часы при высокой температуре воздуха и значительном водном дефиците в листьях наблюдается депрессия фотосинтеза у всех изученных видов, менее всего она выражена у лиственницы сибирской. Глубина депрессии фотосинтеза также во многом зависит и от физиологического состояния растений, от уровня их жизнеспособности. У деревьев, отставших в росте, этот показатель имеет очень низкие значения (0.05–0.8 мг СО2 г–1 сух. хвои ч–1), а в отдельные пасмурные дни не поддается учету.
В утренние часы при достаточной водообеспеченности и умеренных температурах воздуха наблюдается более высокая интенсивность фотосинтеза у всех изучаемых древесных пород. Интенсивность фотосинтеза здоровых деревьев лиственницы сибирской при относительном световом довольствии 70% и выше достигает в это время 14.0–15.0 мг СО2 г–1 сух. хвои ч–1. Значения нетто ассимиляции в утреннее время для двухлетней хвои сосны обыкновенной варьируют в пределах 7.0–9.0 мг СО2 г–1 сух. хвои ч–1, у ели европейской 5–7 мг СО2 г–1 сух. хвои ч–1. Хвоя лиственницы сибирской и сосны кедровой сибирской отличается высокой интенсивностью дыхания. Коэффициент изменчивости данного показателя очень высокий и составляет у лиственницы и кедра 105–106%. У сосны обыкновенной и ели европейской изменчивость интенсивности дыхания значительно меньше и равна соответственно 79% и 61%.
Содержание общего хлорофилла у лиственницы сибирской в Среднем Поволжье равно 3.75 мг г–1 сухой хвои и имеет показатели, близкие к параметрам, полученным для условий естественного ареала в Красноярском крае, которые составляют 3.27–3.77 мг г–1 сухой хвои (Милютин, 1974). В Московской области в интродукционных культурах лиственница имеет более высокое содержание общего хлорофилла (11.03–12.05 мг г–1 сухой хвои), что объясняется более благоприятными условиями почвенного питания (Надеждин, 1971).
Полученные нами данные согласуются с данными других исследователей, отмечающими высокую ассимиляционную способность лиственницы. Среди хвойных пород, отличающихся повышенным фотосинтезом, в условиях Белоруссии А.Ф. Иванов (1975) также выделяет лиственницу сибирскую, средняя интенсивность фотосинтеза которой в июне составляет 20.4 мг СО2 г–1 сух. хвои ч–1. В Красноярском крае, по данным Г.М. Голомазовой (1978), интенсивность фотосинтеза лиственницы сибирской варьировала в пределах от 1.5 до 4.5 мг СО2 г–1 сырой хвои ч–1, что может составлять от 4.5 до 13 мг СО2 г–1 сух. хвои ч–1. Существенная вариабельность фотосинтеза и дыхания свидетельствует о высокой адаптивности интродуцированных хвойных видов к климатическим условиям Среднего Поволжья.
У деревьев, растущих в одинаковых почвенных и световых условиях, установлена значительная индивидуальная изменчивость основных физиологических процессов. Так, на лесосеменной плантации деревья сосны обыкновенной с обильным семеношением отличаются более высокой интенсивностью фотосинтеза, содержанием хлорофилла в хвое и величинами биоэлектрических потенциалов по сравнению со слабоурожайными (табл. 2). По интенсивности транспирации и дыхания четкой связи с семенной продуктивностью не установлено.
Таблица 2.
Интенсивность основных физиологических процессов деревьев сосны обыкновенной различной репродуктивной способности
| № дерева | Интенсивность транспирации 2-летней хвои, г Н 2 О на 1 г абс. сух массы ч–1 | Интенсивность реального фотосинтеза, мг СО 2 на 1 г абс. сух массы ч–1 для однолетней хвои | Интенсивность дыхания однолетней хвои, мг СО 2 на 1 г абс. сух массы | Величина БЭП на участке бокового побега “верхушечная почка–первая мутовка”, мВ | Содержание общего хлорофилла, мг г–1 сух хвои |
|---|---|---|---|---|---|
| Обильносеменосящие | |||||
| 1 | 0.41 | 2.06 | 0.27 | 184.0 | 1.36 |
| 7 | 0.39 | 1.93 | 0.18 | 180.0 | 1.43 |
| 5 | 0.44 | 1.85 | 0.26 | 140.0 | 2.39 |
| Среднесеменосящие | |||||
| 4 | 0.37 | 0.45 | 0.24 | 64.0 | 1.61 |
| 10 | 1.15 | 2.31 | 0.28 | 178.0 | 1.61 |
| 13 | 0.68 | 1.38 | 0.19 | 80.0 | 0.71 |
| Слабосеменосящие | |||||
| 15 | 0.66 | 0.96 | 0.29 | 42.0 | 1.16 |
| 8 | 0.79 | 0.90 | 0.20 | 180.0 | 0.84 |
| 14 | 0.27 | 0.52 | 0.21 | 60.0 | 2.10 |
Приведенные нами данные свидетельствуют о взаимосвязи БЭП с интенсивностью обменных процессов растений, что отмечалось в работах многих исследователей (Коловский, 1980; Медведев, 1997; Пятыгин, 2008). Высокоурожайные деревья отличаются высокими величинами БЭП на всех участках отведения, большим содержанием хлорофилла и более высокими значениями реального фотосинтеза (Маторкин и др., 2011).
Имеются также малоурожайные семенные деревья, отличающиеся быстрым ростом и высокими значениями БЭП, что обусловлено их индивидуальными особенностями. У стадийно старых деревьев в возрасте 120–140 лет показатели БЭП снижаются до 40–45 мВ.
Подобные исследования по диагностике физиологического состояния деревьев сосны обыкновенной по БЭП были проведены в зеленой зоне Казани на опытных объектах Татарской лесной опытной станции (ныне Восточно-Европейская лесная опытная станция ВНИИЛМ). Объектами исследований являлись разновозрастные (70–150 лет) естественные насаждения сосны обыкновенной. В результате длительной рекреации и техногенного воздействия данные насаждения характеризуются почти полным отсутствием естественного возобновления главной породы, возрастанием доли ослабленных, фаутных и перестойных деревьев (Краснобаева и др., 1998). При измерении биоэлектрических показателей установлено, что в данном насаждении 70–80 летние деревья из категории “лучшие” имеют наиболее высокие значения БЭП (185–230 мВ), средние деревья имели параметры 143 ± 3.4 мВ, ослабленные – 100 ± 4.6 мВ и усыхающие – 70 ± 5.0 мВ. Деревья перестойные (140–150 лет), значительно ослабленные, стадийно-старые, фаутные и пораженные бугорчатым раком, отличаются низкими значениями БЭП (66–100 мВ). Импеданс ПКТ у лучших деревьев был равен 35 ± 0.8 кОм, средних 44 ± 3.8 кОм и перестойных и усыхающих варьировал в пределах 57–927 кОм. Все перестойные, физиологически ослабленные и селекционно-малоценные особи удалялись. Заключительная диагностика по величинам БЭП и импедансу ПКТ позволила выявить наиболее ценные в селекционном отношении экземпляры деревьев. Их семенное потомство, в пятилетнем возрасте имело высоту 72.5 ± 6.1 см, параметры БЭП 180.0 ± 5.0 мВ и импеданса ПКТ 37.0 ± 0.6 кОм, что свидетельствует о хорошем жизненном состоянии подроста и сохранении ценного адаптированного к местным условиям генофонда.
Установлена высокая взаимосвязь величин БЭП с интенсивностью ростовых процессов различных видов в природной среде. Наиболее адаптированные к климатическим условиям региона, формирующие устойчивые высокопродуктивные насаждения, быстрорастущие интродуценты (лиственница сибирская, сосна кедровая сибирская) имеют значения БЭП, близкие к аборигенным видам соответственно 211–186 мВ (табл. 3). Более высокие показатели БЭП имеют молодые хвойные растения в период активного роста. В возрасте 20–40 лет деревья–лидеры имеют значения БЭП почти в два раза выше, чем деревья средней категории состояния, что обусловлено высокой интенсивностью обменных процессов.
Таблица 3.
Показатели биоэлектрических потенциалов и импеданса ПКТ стволов деревьев Pinus sylvestris, Picea abies, и интродуцированных видов Larix sibirica, Pinus sibirica различного жизненного состояния (возраст 40 лет)
| Статистические показатели | Биоэлектрический потенциал деревьев, мВ | Импеданс ПКТ, кОм | ||||
|---|---|---|---|---|---|---|
| лидирующие | средние | ослабленные | лидирующие | средние | ослабленные | |
| Pinus sibirica | ||||||
| Х | 186.3 | 144.4 | 97.0 | 16.4 | 25.6 | 47.9 |
| Мx | 11.62 | 6.0 | 3.51 | 0.53 | 0.88 | 3.78 |
| V, % | 25.0 | 15.0 | 11.5 | 10.7 | 12.9 | 33.5 |
| Larix sibirica | ||||||
| Х | 211.5 | 149.4 | 66.8 | 15.4 | 28.6 | 149.4 |
| Мx | 10.60 | 11.80 | 5.90 | 0.30 | 0.90 | 11.80 |
| V, % | 15.2 | 50.1 | 30.9 | 15.2 | 19.9 | 50.1 |
| Pinus sylvestris | ||||||
| Х | 229.0 | 143.0 | 70.0 | 21.9 | 46.1 | 70.0 |
| Мx | 8.21 | 6.40 | 8.0 | 0.60 | 2.87 | 8.0 |
| V, % | 11.8 | 14.8 | 34.2 | 12.6 | 21.6 | 34.2 |
| Picea abies | ||||||
| Х | 196.0 | 134.0 | 60 | 21.3 | 38.6 | 52.3 |
| Мx | 6.0 | 8.1 | 0.6 | 0.4 | 2.7 | 2.04 |
| V, % | 15.0 | 20.0 | 35.0 | 5.7 | 13.4 | 9.6 |
Примечание. Обозначение см. табл. 1.
Лидирующие деревья всех изученных видов отличаются не только более высокими значениями БЭП по сравнению с ослабленными, но и более высокой оводненностью прикамбиального комплекса тканей, о чем свидетельствуют низкие величины импеданса ПКТ. Связь между влажностью растительных тканей и величинами импеданса ПКТ очень тесная и обратная (r = –0.85…–0.90). При нарушении деятельности корневых систем, водных стрессах значения параметров импеданса ПКТ увеличиваются. Коэффициент детерминации, характеризующий связь жизненного состояния деревьев и показателей импеданса ПКТ, равен 0.909. У здоровых растений значение импеданса ПКТ в 2−3 раза меньше, чем у ослабленных. Хвойные интродуценты хорошего состояния, произрастающие в идентичных условиях, имеют специфичные для вида величины импеданса ПКТ (рис. 1).
Рис. 1.
Распределение здоровых растений по ступеням импеданса ПКТ 1 – ель европейская; 2 – лиственница сибирская; 3 – сосна обыкновенная.
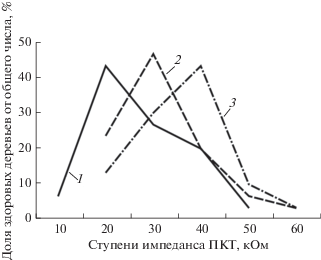
Для деревьев разного физиологического состояния наиболее существенные различия в показателях импеданса отмечены в период интенсивного роста и сохраняются на протяжении всего вегетационного периода, к осени величины импеданса ПКТ стволов постепенно увеличиваются, при подготовке к зимнему покою и в период зимнего покоя возрастают до 400 кОм. Наиболее высокая изменчивость импеданса ПКТ (13.2–21.0%) отмечена у побегов кроны, что обусловлено значительной изменчивостью влажности побегов. Влажность ПКТ стволов здоровых деревьев в условиях нормального водообеспечения довольно стабильна, изменчивость параметров импеданса ПКТ по высоте ствола не превышает 5–8%. Значительные различия в показателях импеданса по высоте стволов отмечены у семенных деревьев сосны обыкновенной, созданных прививками. Импеданс подвоя и привоя иногда имеет существенные различия в параметрах, что свидетельствует о нарушении водного режима растений и приводит к ослаблению роста таких деревьев. Средний прирост в высоту у них в два раза меньше, чем у растений, имеющих одинаковые параметры импеданса привоя и подвоя. Привитые перспективные деревья, имеют одинаковые параметры импеданса ПКТ привоя и подвоя, значения которых у семенников варьируют в пределах 25–30 кОм. Это свидетельствует о нормальной водообеспеченности и физиологической совместимости привоев с подвоями и позволяет проводить отбор перспективных, устойчивых к водным стрессам семенных деревьев (Маторкин и др., 2011; Карасев и др., 2016).
О целесообразности применения импедансметрии как индикатора физиологического состояния растений свидетельствуют исследования зарубежных авторов (Borges et al., 2014). Исследователи проводили измерения импеданса растений высотой 0.8 м и диаметром 1.0 см в ночное и дневное время в контролируемых условиях при различной интенсивности поливов, что позволило выявить изменение параметров импеданса при водных стрессах еще при отсутствии визуальных признаков.
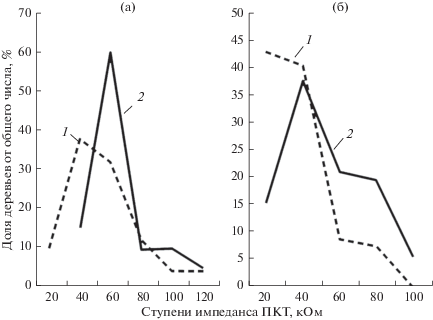
Неблагоприятными факторами, оказывающими негативное воздействие на жизненное состояние древостоев, является рекреационное воздействие, что приводит к ослаблению деятельности корневых систем и нарушению водного режима растений. Численные показатели, характеризующие данные изменения в состоянии древостоя, позволяет получить экспресс-диагностика по параметрам импеданса ПКТ, показатели которого являются индикатором состояния прикамбиального комплекса тканей и имеют более высокие значения при ослаблении (рис. 2). Число деревьев с параметрами импеданса ПКТ 60 кОм и более, что характерно для ослабленных деревьев, составило в рекреационной зоне у сосны обыкновенной 85.0%. У лиственницы, растущей в таких же условиях, это величина равнялась 46.5%, что свидетельствует об ее более высокой устойчивости к рекреационному воздействию.
Рис. 2.
Распределение деревьев сосны обыкновенной (a) и лиственницы сибирской (б) по ступеням импеданса ПКТ при различном рекреационном воздействии 1 – малопосещаемый лесной массив; 2 – рекреационная зона (площадь уплотненной поверхности почвы составляет 30% от общей площади).
При экстремальном повреждении или утрате части кроны и как следствие этого уменьшении расхода воды на транспирацию повышается оводненность стволов. Нарушение водного баланса приводит к уменьшению величин импеданса до очень низких значений, что необходимо учитывать при диагностике физиологического состояния деревьев по данному показателю и свидетельствует о целесообразности использования нескольких оценочных параметров. Измерения импеданса ПКТ и температурных параметров стволов проводили у деревьев лиственницы сибирской, поврежденных лиственничной чехликовой молью (Coleophora laricella), с различной степенью дефолиации крон (средняя степень дефолиации – 25–50% и сильная более – 50%) (рис. 3). При повреждении крон нарушается соотношение между поступлением воды и расходом ее на транспирацию. Первые измерения, проведенные 27 мая, показали, что изменение транспирирующей поверхности вследствие повреждения кроны вредителем приводит к повышению оводненности стволов и снижению их импеданса ПКТ, значения которого существенно меньше, чем у неповрежденных деревьев. Коэффициент детерминации импеданса ПКТ деревьев разных категорий состояния имел значения 0.497. При повторных измерениях через 14 дней поврежденные деревья имели параметры импеданса ПКТ, близкие к здоровым деревьям. Т.е. поддерживается оводненность прикамбиального комплекса на уровне, сохраняющем водный баланс, необходимый для жизнедеятельности растений. Информативность импеданса ПКТ при таком повреждении (η2 = = 0.026) низкая.
Рис. 3.
Показатели импеданса ПКТ (а) и температурные параметры (б) деревьев лиственницы сибирской при различной степени дефолиации крон: 1 – контроль (неповрежденная крона); 2 – средняя дефолиация крон (25–50%); 3 – сильная дефолиация крон (более 50%).
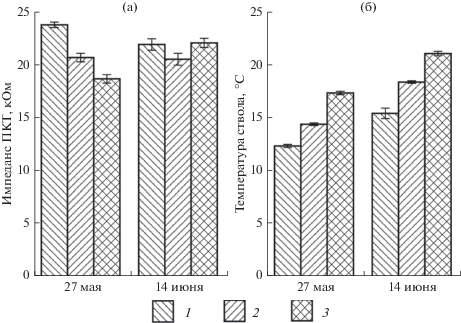
Интенсивность водообмена тесно связана с температурными параметрами деревьев, которые являются индикатором их физиологического состояния. Ослабленные деревья лиственницы сибирской с дефолиацией кроны вследствие слабого транспирационного тока отличаются более высокими температурными параметрами ствола по сравнению со здоровыми. Коэффициент детерминации, характеризующий связь температурных показателей и состояния, имеет высокие значения (η2 = 0.895). Более высокую информативность при водных стрессах имеют показатели температуры водопроводящей ксилемы стволов деревьев, характеризующие взаимосвязи показателей водного режима деревьев с их физиологическим состоянием.
В результате периодического, избыточного увлажнения почвы деревья сосны обыкновенной испытывают значительное ослабление. У ослабленных деревьев отмечен пониженный водный ток в результате нарушения корневой деятельности из-за недостатка кислорода (табл. 4).
Таблица 4.
Показатели водного режима деревьев сосны обыкновенной различных категорий состояния, ослабленных в результате периодического, избыточного увлажнения почвы
| Показатели | Статистические показатели деревьев разных категорий состояния | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| условно здоровые | средне- ослабленные |
сильно ослабленные |
усыхающие | |||||||||
| Х | Mx | V, % | Х | Mx | V, % | Х | Mx | V, % | Х | Mx | V, % | |
| Масса сырой хвои на дереве, кг | 47.2 | 2.6 | 18.0 | 20.0 | 1.4 | 24.0 | 12.3 | 0.57 | 24.6 | 4.70 | 0.46 | 23.0 |
| Интенсивность транспирации (на сырую массу), г кг–1 ч–1 | 134.6 | 11.8 | 28.4 | 122.3 | 8.6 | 22.6 | 70.2 | 7.0 | 22.6 | 48.9 | 4.3 | 23.2 |
| Скорость водного тока, м ч–1 | 0.34 | 0.01 | 12.2 | 0.15 | 0.01 | 16.9 | 0.15 | 0.004 | 16.8 | 0.04 | 0.002 | 18.2 |
| Расход воды деревом, кг ч–1 | 6.36 | 0.22 | 12.5 | 2.45 | 0.09 | 14.0 | 0.86 | 0.03 | 14.5 | 0.23 | 0.02 | 32.0 |
| Влажность водопроводящей ксилемы на высоте 1.3 м, % к абс. сух. массе | 114.7 | 4.2 | 11.9 | 109 | 5.4 | 15.9 | 89.6 | 5.0 | 16.2 | 56.0 | 2.9 | 16.9 |
Примечание. Обозначение см. табл. 1. Измерения проведены в 14 ч при температуре воздуха 28.0°С и почвы на глубине 0.7 м – 13.2°С, диаметр деревьев 28.0 см.
Установлено, что между расходом воды на транспирацию и массой хвои на дереве, а также между расходом воды и температурой ствола существует определенная связь. Так, для деревьев сосны обыкновенной диаметром 28.0 ± 1.0 см при суточной максимальной температуре воздуха 28°С связь между расходом воды на транспирацию и температурой ствола с северной стороны (на высоте 1.3 м, в средней части водопроводящей зоны) обратная, очень высокая (r = –0.985) и описывается уравнением: Y = 20.24 – 0.75 × X, где Y – расход воды деревом на транспирацию, кг ч–1; X – температура ствола в заданной точке, °С. Любые существенные нарушения водного тока в корнях, стволовой части либо в кроне немедленно сказываются на температуре ствола дерева, при этом ослабленные и усыхающие деревья характеризуются более высокими температурами. Скорость водного тока зависит от транспирационных потерь, определяемых интенсивностью транспирации и массой хвои на дереве. Вода, поступающая в ствол, имеет температуру, близкую к температуре корнеобитаемых горизонтов почвы. Ствол здорового дерева из-за высокого содержания воды, обладающей высокой теплоемкостью, большей скоростью водного тока противостоит нагревающему действию солнечной радиации окружающего воздуха намного эффективнее, чем стволы усыхающих деревьев. Разница температур между стволами здоровых и усыхающих деревьев составляет 10–14°С.
Температура стволов здоровых деревьев у корневой шейки и на высоте 1.3 м в период активного водного тока имеет показатели, близкие к температуре корнеобитаемых горизонтов почвы 13.2°С (табл. 5). Связь температуры стволов с жизнеспособностью деревьев высокая и обратная, коэффициент детерминации варьирует в пределах 0.965–0.985. Согласно проведенной оценке в 96.5% случаев температура ствола связана с состоянием дерева. При оценке по величинам БЭП такая связь составляет 75%, по величинам импеданса ПКТ – 90.9%. Экспресс-диагностика физиологического состояния деревьев по температуре стволов отличается высокой информативностью при нарушениях водного режима и малыми затратами времени на измерения.
Таблица 5.
Статистические показатели БЭП, импеданса ПКТ и температуры стволов деревьев сосны обыкновенной, в разной степени ослабленных в результате периодического избыточного увлажнения
| Категория жизнеспособности деревьев | Биоэлектрический потенциал деревьев, мВ | Импеданс ПКТ, кОм | Температура стволов деревьев на высоте 1.3 м, °С | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Х | Мx | V, % | Х | Мx | V, % | Х | Мx | V, % | |
| Здоровые | 120.0 | 4.0 | 10.7 | 57.6 | 5.48 | 30.1 | 18.7 | 0.13 | 2,3 |
| Незначительно ослабленные | 97.8 | 3.89 | 13.8 | 102.0 | 3.18 | 9.9 | 20.5 | 0.12 | 2.0 |
| Среднеослабленные | 76.4 | 6.14 | 27.9 | 127.3 | 3.12 | 7.7 | 23.8 | 0.15 | 2.4 |
| Сильноослабленные | 29.7 | 1.75 | 20.4 | 176.1 | 9.42 | 16.9 | 25.9 | 0.18 | 2,1 |
| Усыхающие | Отсутствует | 344.4 | 40.20 | 33.0 | 28.0 | 0.24 | 2.8 | ||
Примечание. Обозначение см. табл. 1.
Заключение. Большая изменчивость интенсивности фотосинтеза и дыхания хвойных в разные годы свидетельствует о высокой адаптивной способности аборигенных и интродуцированных хвойных видов в климатических условиях Среднего Поволжья. К числу пород, обладающих высокой фотосинтезирующей способностью и устойчивостью в климатических условиях региона, относится лиственница сибирская, что согласуется с исследованиями многих авторов и свидетельствует о целесообразности более широкого использования данной ценной породы при создании насаждений в рекреационных лесах данного района.
Исследования величин БЭП деревьев в лесных фитоценозах показали высокую связь параметров БЭП с физиологическим состоянием деревьев и целесообразность использования этого показателя при селекционной оценке семенных деревьев и отборе быстрорастущих биотипов.
Состояние водного режима растений диагностируется по показателям импеданса ПКТ. Показатели импеданса ПКТ имеют прямую обратную связь с влажностью прикамбиального комплекса тканей хвойных видов. Деревья сосны обыкновенной и лиственницы сибирской имеют более высокие значения импеданса ПКТ по сравнению с сосной кедровой сибирской и елью европейской.
Резкое изменение массы ассимиляционного аппарата при повреждении вредителями или другими факторами, ведущее к изменению баланса между подачей воды корневыми системами и расходом ее на транспирацию, приводит к повышению оводненности прикамбиального комплекса тканей и следовательно уменьшению значений импеданса ПКТ. Показатели импеданса ПКТ в данном случае низко информативны (η2 = 0.497).
Исследования состояния насаждений хвойных пород, испытывающих длительно действующие неблагоприятные факторы среды, показали целесообразность применения импеданса ПКТ для диагностики жизненного состояния на ранних этапах ослабления. Коэффициент детерминации, отражающий связь импеданса ПКТ с состоянием деревьев при таких воздействиях, имеет высокие значения (η2 = 0.757–0.909).
Установлено, что в 96.5% случаев температура ствола деревьев сосны обыкновенной различного жизненного состояния связана с состоянием дерева. Связь температуры стволов с физиологическим состоянием деревьев при нарушениях водного режима высокая и обратная, коэффициент детерминации варьирует в пределах 0.965–0.974. Выявленные взаимосвязи температурных параметров стволов деревьев с расходом воды деревом на транспирацию позволяют также получить информацию о расходе воды и массе хвои на дереве.
Комплексная экспресс-диагностика жизнеспособности деревьев хвойных пород по биоэлектрическим потенциалам, импедансу ПКТ и температурным параметрам позволяет контролировать изменение их физиологического состояния на ранних этапах ослабления, при этом необходимо учитывать особенности и типы повреждения и ослабления деревьев, календарные сроки реализации измерений и информативность используемых при этом показателей при ведении исследований.
Список литературы
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Воробьев Р.А. Содержание основных пигментов в хвое интродуцентов рода Ель в условиях южной тайги (на примере Нижегородской области) // Лесоведение. 2013. № 4. С. 22–31.
Годнев Т.Н. Строение хлорофилла и методы его количественного определения. Минск: Изд-во АН БССР, 1952. 327 с.
Голомазова Г.М. Фотосинтез и фотодыхание лиственницы сибирской при низких концентрациях СО2 // Лиственница и ее использование. Красноярск: Сибир. технол. ин-т, 1987. Вып. 9. С. 53–60.
Иванов А.Ф. Биология древесных растений. Минск: Наука и техника, 1975. 259 с.
Иванов В.П., Иванов Ю.В., Марченко С.И., Кузнецов В.В. Использование индексов флуктурирующей ассиметрии листа березы повислой для диагностики состояния фитоценозов в условиях техногенного загрязнения // Физиология растений. 2015. Т. 62. С. 368–377.
Карасев В.Н. Физиология растений. Йошкар-Ола: Марийский гос. тех. университет, 2001. 304 с.
Карасев В.Н., Карасева М.А. Эколого-физиологическая диагностика жизнеспособности деревьев хвойных пород // Лесной журн. 2004. № 4. С. 27–32.
Карасев В.Н., Карасева М.А., Панюшкина Н.В., Бродников С.Н. Способ формирования лесосеменных плантаций сосны обыкновенной. Патент № 2579798 (РФ) // Официальный бюллетень “Изобретения. Полезные модели”. 2016. № 10. 6 с.
С. Коловский Р.А. Биоэлектрические потенциалы древесных растений. Новосибирск: Наука, 1980. 176 с.
Краснобаева К.В., Мусин Х.Г., Карасев В Н. Отбор семенных деревьев при проведении рубок омоложения в сосновых лесах зеленой зоны г. Казани по их биоэлектрическим и тепловым параметрам // Современные проблемы учета и рационального использования лесных ресурсов: Матер. науч.-практич. конф., 27 января 1998 г. Йошкар-Ола, 1998. С. 127–128.
Критерии и индикаторы для сохранения устойчивости управления умеренных и бореальных лесов. М.: ВНИИЦлесресурс, 1995. 25 с.
Кудоярова Г.Р., Холодова В.П., Веселова Д.С. Современное состояние проблемы водного баланса растений при дефиците воды // Физиология растений. 2013. Т. 60. С. 150–160.
Маторкин А.А., Карасева М.А. Информативность импеданса прикамбиального комплекса тканей деревьев хвойных пород при диагностике их жизнеспособности // Современная физиология растений: от молекул до экосистем: Матер. доклад. Междунар. конф. Сыктывкар, 2007. Ч. 2. С. 265–266.
Медведев С.С. Электрофизиология растений. СПб.: Изд-во Санкт-Петербургского гос. университета, 1997. 86 с.
Милютин Л.И. О некоторых физиологических признаках лиственниц сибирской и даурской // Изменчивость древесных растений Сибири. Красноярск: Ин-т леса и древесины им. В.Н. Сукачева, 1974. С. 35–45.
Надеждин В.В. Влияние географического происхождения семян лиственницы на ее рост в подзоне хвойно-широколиственных лесов. М.: Наука, 1971. 131 с.
Опритов В.А., Пятыгин С.С., Ревитин В.Г. Биоэлектрогенез у высших растений. М.: Наука, 1991. 276 с.
Пятыгин С.С. Распространяющиеся электрические сигналы в растениях // Цитология. 2008. Т. 50. № 2. С. 154–159.
Рутковский И.В. Рекомендации по методике измерений электрофизиологических характеристик древесных растений с целью оценки их состояния и жизнеспособности. Пушкино: Всесоюз. НИИ лесн. механизации, 1975. 18 с.
Софронова В.Е., Дымова O.В., Головко Т.К., Чепалов В.А., Петров К.А. Адаптивные изменения пигментного комплекса хвои Pinus sylvestris L при закаливании к низкой температуре // Физиология растений. 2016. Т. 63. С. 461–471.
Судачкова Н.Е., Милютина И.Л., Семенова Г.П. Состав свободных аминокислот различных органов тканей Pinus sylvestris–Larix sibirica–Larix gmelina // Растительные ресурсы. 2003. Т. 39. С. 19–31.
Шевякова Н.И. Метаболизм и физиологическая роль пролина в растениях при водном и солевом стрессе // Физиология растений. 1983. Т. 30. С. 768–763.
Borges E., Sequeira M., Cortez A. F., Pereira H. Bioimpedance parameters as indicators of the Physiological States of Plants in situ // International J. Advances in Life Sciences. 2014. V. 6. P. 74–86.
Oyarce P., Gurovich L. Evidence for the transmission of information through electric potentials in injured avocado trees // Plant Physiology. 2011. V. 168. P. 103–108.
Rнos–Rojasa L., Franco T., Gurovich A.A. Electrophysiological assessment of water stress in fruit–bearing woody plants // Plant Physiology. 2014. V. 171. P. 799–806.
Дополнительные материалы отсутствуют.