

Лесоведение, 2020, № 3, стр. 195-204
Влияние контролируемого выжигания на содержание стрессовых белков в хвое сосны обыкновенной в условиях Красноярской лесостепи
И. Г. Гетте a, Н. Е. Коротаева b, *, И. В. Косов c, Н. В. Пахарькова a, Г. Б. Боровский b
a Сибирский федеральный университет
660041 Красноярск, просп. Свободный, 79, Россия
b Сибирский институт физиологии и биохимии растений СО РАН
664033 Иркутск, ул. Лермонтова, 132, Россия
c Институт леса им. В.Н. Сукачева СО РА
660036 Красноярск, ул. Академгородок, 50/28Н, Россия
* E-mail: knev73@yandex.ru
Поступила в редакцию 28.04.2018
После доработки 23.08.2018
Принята к публикации 29.01.2020
Аннотация
Для сопоставления величины накопления белков теплового шока HSP (Heat Shock Proteins) с интенсивностью и длительностью температурного воздействия исследовано влияние экспериментального контролируемого выжигания, имитирующего низовой пожар слабой силы (с регистрацией продолжительности и температуры огневого воздействия), на содержание HSP в хвое сосны обыкновенной (Pinus sуlvestris L.) в насаждениях I класса возраста в лесостепной зоне Южной Сибири, как сразу после него, так и в краткосрочный послепожарный период (2 и 5 сут). Установлено, что в момент контролируемого выжигания в течение 5.5–11.8 мин. на крону оказывали влияние температуры в диапазоне от 48 до 76°С. Через 30 мин. после высокотемпературного воздействия содержание HSP в хвое значимо не отличалось от контрольного уровня. Наиболее высокое содержание HSP 101 и HSP 17.6, было отмечено через 2 сут после горения. На 5-е сут содержание HSP 17.6 вернулось к первоначальному уровню, содержание HSP 101 несколько снизилось по сравнению со значениями на 2-е сут. Полученные данные позволяют предположить, что в послепожарный период в клетках хвои происходит синтез специфических HSP, возможно, обусловленный их участием в восстановлении поврежденных огневым воздействием тканей.
В процессе индивидуального развития растительные организмы в естественных условиях периодически подвергаются действию различных стрессовых факторов. В лесных биогеоценозах существенное значение имеет пирологический фактор, влияющий на продуктивность и устойчивость насаждений (Швиденко, Щепащенко, 2013; Kukavskaya et al., 2014; Цветков, Буряк, 2014). Многими авторами получены данные по многоплановому влиянию лесных пожаров на ствол и крону деревьев, как через последующее изменение условий среды, так и через непосредственное воздействие высоких температур во время горения (Горшков и др., 2004; Varner et al., 2009; Судачкова и др., 2016). При этом исследователями отмечается как отрицательное, так и положительное действие пожара на динамику и продуктивность лесных экосистем (Бузыкин и др., 1978; Дымов и др., 2015; Иванова и др., 2017). Так, пожары низкой или средней интенсивности могут приводить к увеличению прироста и диаметра древостоя, изменению в темпах фотосинтеза, увеличению доступности воды и питательных элементов (Fleck et al., 1996; Верховец, 2000; Фуряев, 2008; Renninger et al., 2013).
Степень пирогенной трансформации и последующее восстановление растительности зависят от вида и характера повреждения, которые определяются длительностью и интенсивностью пожара. Краткосрочное действие высоких температур при прохождении низового пожара оказывает существенное влияние на ряд физиолого-биохимических процессов и показателей растительных организмов (Rieske, 2002; Adams, Rieske, 2003; Креславский и др., 2007; Масягина и др., 2007; Судачкова и др., 2015). Эти изменения вызывают особый интерес, так как могут быть индикаторами стрессового состояния. Особое место среди защитно-приспособительных механизмов растений к действию высоких температур занимает изменение экспрессии генов и рост синтеза стрессовых белков (Кулаева, 1997; Wang et al., 2004). Стрессовые белки (или белки теплового шока, heat shock proteins, HSP) синтезируются в клетках живых организмов в ответ на различные стрессовые факторы и играют ключевую роль в адаптивных реакциях, обеспечивающих выживание, адаптацию и восстановление при неблагоприятных условиях (Кулаева, 1997; Timperio et al., 2008; Al-Whaibi, 2011).
Большинство HSP действуют как внутриклеточные шапероны, отвечающие в оптимальных и неблагоприятных условиях за формирование третичной конформации полипептидов, формирование четвертичных белковых комплексов, транспорт через мембраны и утилизацию белков. Шапероны нековалентно связываются с утратившими нативную конформацию белками и, не являясь частью конечного продукта после его восстановления, способствуют стабилизации и репарации структуры поврежденных белков в условиях стресса (Wang et al., 2004). В зависимости от молекулярной массы описано несколько классов этих белков, одними из которых являются класс высокомолекулярных белков HSP 100 и класс низкомолекулярных белков (small, sm) HSP (Mr от 15 до 45 кД). Защитный эффект шаперонов в клетках растений объясняется согласованной работой представителей нескольких классов этих белков, когда smHSP связываются с частично поврежденными белками, предотвращая их агрегацию и формируя пул субстратов для последующего восстановления шаперонами семейств HSP 70/HSP 100 (Smŷkal et al., 2000; Lee, Vierling, 2000; Mogk et al., 2003). В настоящее время признано, что HSP 101 накапливается под действием теплового шока и необходим для формирования индуцированной термотолерантности (Queitsch et al., 2000); smHSP обнаружены практически во всех компартментах клетки. В хлоропластах smHSP поддерживают активность фотосинтетического транспорта электронов в условиях теплового и окислительного стрессов (Downs et al., 1998; Neta-Sharir et al., 2005), защищают термолабильную фотосистему II и, следовательно, электронный транспорт во время теплового воздействия (Heckathorn et al., 1998). Таким образом, эти стрессовые белки, действуя сообща, могут быть компонентом клеточного ответа на огневое воздействие.
Продолжительность и интенсивность влияния высоких температур и вызванная ими степень стрессового состояния деревьев могут варьировать. В условиях естественного пожара проследить точечные изменения температуры в кроне дерева и связанные с ними первичные реакции фотосинтетического аппарата хвои достаточно сложно. В то же время, контролируемое выжигание позволяет получить характеристики температурного воздействия и проследить стрессовую реакцию дерева как сразу, так и в период восстановления. Таким образом, цель исследования заключалась в моделировании воздействия низового пожара и выявлении изменений в содержании белков теплового шока в хвое сосны обыкновенной, ранее не подвергавшейся пирогенному стрессу.
ОБЪЕКТЫ И МЕТОДИКА
Район исследования расположен в 5 км к северо-востоку от пос. Устюг Емельяновского района Красноярского края и представляет собой участок островной лесостепи на дерново-подзолистой почве с насаждением сосняка разнотравной группы типов леса I класса возраста, сформировавшегося от материнского древостоя и ранее не подвергавшегося воздействию пожаров. Экспериментальный участок был заложен в 2017 г. для проведения контролируемого горения методом отжига от одной стороны участка, имитирующего кромку низового пожара слабой силы (Курбатский, 1962). Экспериментальное выжигание проводилось 5 августа 2017 г. в 16 ч, на участке площадью 0.04 га. Температура воздуха составляла 31°С, относительная влажность воздуха – 39%, скорость ветра – 0–1 м с–1.
Во время эксперимента оценивались высота пламени, скорость продвижения кромки пожара, глубина прогорания лесных горючих материалов, характеристики упавших древесных горючих материалов. Температура конвективного потока в кроне модельных деревьев на высоте 3 м фиксировалась датчиками температуры, которые крепились на модельные деревья в том месте веток, где в дальнейшем отбирались пробы хвои для полуколичественного анализа содержания HSP. Считывание показания датчиков на персональный компьютер производилось через USB-интерфейс.
На экспериментальном участке проводился сплошной учет древостоя с замером основных таксационных показателей, определялся запас лесных горючих материалов. Средний диаметр стволов на высоте 1.3 м составлял 8.5 ± 3.3 м, высота древостоя – 7.0 ± 1.3 м. Для оценки запаса горючих материалов использовали методику Н.П. Курбатского (1970). Учет запаса упавших древесных горючих материалов проводили по методике пересеченных линий (McRae et al., 1979). Лесотаксационные работы, определение жизненного состояния проводились в соответствии с общепринятыми рекомендациями и методиками (Побединский, 1966; Курбатский, 1970; Сукачев, 1972; Алексеев, 1989).
Температуру конвективного потока в кроне измеряли 4-я автономными регистраторами температуры EClerk (“РЭЛСИБ”, Россия) с разрешающей способностью в 1 с. Измеряющим элементом служили хромель-алюминиевые термопары заводского изготовления (“РЭЛСИБ”, Россия). Также во время эксперимента с помощью тепловизора FLIR E5 (FLIR, США) фиксировались комбинированные снимки объекта исследования для полного анализа температурного профиля.
Для выявления содержания стрессовых белков 2-летнюю хвою отбирали с двух веток средней части кроны (с разных сторон ствола) у трех экспериментальных деревьев в равных пропорциях. Таким образом, для анализа использовали объединенные пробы с трех деревьев. Отбор хвои проводили перед выжиганием (контроль), через 30 мин. и спустя 2 и 5 сут после выжигания, в трех повторностях. Отобранная хвоя сразу фиксировалась в жидком азоте, транспортировалась и хранилась в кельвинаторе при температуре –80°С до начала экстракции общего белка.
Общий белок выделяли по ранее опубликованной методике (Korotaeva et al., 2012). Концентрацию белка определяли с помощью реактива Брэдфорда (“Bio-Rad”, США). Перед электрофорезом количество наносимого белка было выровнено, на гель наносили по 5 мкг белка каждой пробы. После разделения белков с помощью Na-ДДС-электрофореза в 12%-м ПААГ и WesternBlot в системе mini–Protean III (“BioRad”, США) проводили инкубирование полученных нитроцеллюлозных мембран в растворе антител против стрессовых белков HSP 101 (AS07 253, “Agrisera”, Швеция) или HSP 17.6 (класс I, AS07 254, “Agrisera”, Швеция). Антитела визуализировали с помощью вторичных антител, конъюгированных с щелочной фосфатазой (“Sigma”, США), в присутствии BCIP и NBT (“Gerbu”, Германия). Интенсивность окрашивания белкового пятна на мембране определяли после сканирования в цифровой формат денситометрически с помощью программы GelAnalysis (Россия) в условных единицах, которые соответствовали числу окрашенных пикселей на изображении. HSP в каждом образце выражали в услов. единицах относительно контрольного образца, содержание соответствующего белка в котором было принято за единицу.
Эксперименты по полуколичественному определению содержания HSP проводились минимум в двукратной повторности для каждой обобщенной пробы. Для статистической оценки содержания белков и формирования статистической диаграммы с помощью программы SigmaPlot 12.0 использовали не менее трех повторностей, вычисляли медиану и 25- и 75-й процентили. Значимость различий оценивали с помощью непараметрического критерия Манна-Уитни (p < 0.05) в программе STATISTICA 8.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
При низовых пожарах в горении участвуют лесные горючие материалы, масса которых составила 0.133 т га–1 и была представлена живым напочвенным покровом (0.009 т га–1), опадом текущего года, подстилкой (0.119 т га–1), упавшими древесными горючими атериалами (0.005 т га–1), куда входят ветви и валеж. В данном эксперименте опад отдельно не отбирался, его масса учтена в составе подстилки.
В силу возраста и состава древостоя напочвенный покров имел слабую степень разложения, в подстилке хвоя 3–4 летней давности сохраняла свою форму, находясь на различной стадии ферментации. В подстилке на экспериментальном участке до выжигания можно выделить два слоя, один из которых представлен слаборазложившимся опадом растений, а второй является слоем ферментации, где опад потерял форму, структуру и частично гумифицировался (Богатырев, 1990). Состав подстилки был представлен отмершими травами и кустарничками (0.015 т га–1), хвоей (0.014 т га–1), корой (0.001 т га–1), шишками (0.002 т га–1), остальная часть подстилки (0.087 т га–1).
С пирологической точки зрения основной неразложившейся частью подстилки являлись проводники горения, такие как отмершие травы и кустарнички, хвоя, кора и шишки (Курбатский, 1970). Вместе с упавшими древесными горючими материалами они составили 0.124 т га–1, что соответствовало 93% общего запаса горючих материалов на участке.
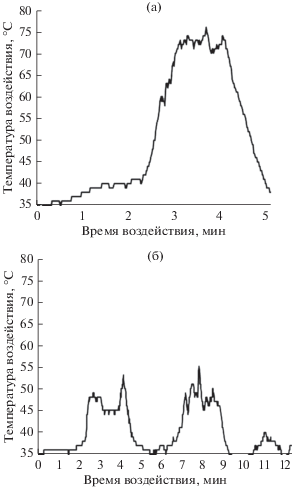
Во время проведения экспериментального выжигания горение протекало в различных фазах, принимая ускоренную или устойчивую форму (рис. 1). Колебания температур во время горения связаны с периодическими возмущениями воздуха в зоне эксперимента, в том числе при штиле. Интенсивное и быстрое сгорание лесных горючих материалов (рис. 1а) вызвано подтоком воздуха и на графике имеет один пик в одной точке кроны, с меньшей продолжительностью, но более высокой температурой. Устойчивое горение (рис. 1б) без ускорения движения воздуха может сопровождаться двумя и более пиками температур и более продолжительным воздействием температуры в одной точке кроны. Стоит отметить, что движение воздуха (поток) к кромке пожара над землей присутствует даже при полном штиле, что связано с вертикальным движением горячей воздушной массы при горении (Валендик, 1968).
Рис. 1.
Кривые изменения температуры конвективного потока от кромки пожара для модельных деревьев 1 (а) и 2 (б).
Хотя сосна обыкновенная относится к группе ксерофитов, для нее оптимум протекания физиологических процессов, в частности, фотосинтеза, лежит в пределах 10–30°С в разные сезоны года (Суворова, 2009), поэтому даже кратковременное повышение температуры до 45–50°С в ходе низового пожара может стать для них значимым стрессовым фактором.
В проведенном эксперименте скорость распространения огня и глубина прогорания подстилки составили 0.25 м мин–1 и 3.5 ± 0.9 см соответственно, средние температуры, превысившие физиологический оптимум в кроне модельных деревьев, составили 46 ± 7°С. Температуры выше 30°С оказывали влияние в диапазоне от 5.5 до 11.8 мин, их максимальные значения варьировали от 48 до 76°С. Результаты, полученные в ходе этого эксперимента, согласуются с данными других исследований о температурных характеристиках пожара, полученных при контролируемых выжиганиях (Валендик, Косов, 2008; Иванова и др., 2014; Carlo et al., 2016).
Снимки, отражающие изменение конвективного потока по кроне, полученные с помощью тепловизора FLIR E5, представлены на рис. 2. Как видно, температура в средней части кроны модельного дерева в момент горения может изменяться в пределах от 40 до 80°С, что соответствует температурам конвективного потока, замеренным автономными регистраторами температуры.
Рис. 2.
Внешний вид древостоя в момент проведения контролируемого выжигания а – Инфракрасный снимок древостоя, курсор наведен на модельное дерево, в среднюю часть кроны; б – цифровое изображение древостоя. Изображения получены одномоментно с помощью тепловизора FLIR Е5.

Таким образом, температура в кроне во время низового пожара превышает максимальные значения температуры естественной среды обитания древостоя. В ответ на эти воздействия в растении должна развиваться стрессовая реакция, приводящая к физиологическим изменениям в нем.
Одним из проявлений защитных механизмов в фотосинтезирующих органах растений является накопление белков теплового шока, действующих как молекулярные шапероны (Валиуллина и др., 2008; Timperio et al., 2008; Xu et al., 2011). Из литературных источников известно, что HSP обнаруживаются в клетках уже через 15 мин после повышения температуры на 10–15°С от оптимальной (Кулаева, 1997). В наших экспериментах образцы свежей хвои отбирались со средней части кроны спустя 30 мин, через 2 и 5 сут после действия огня. На рис. 3 и 4 представлены данные, характеризующие изменения в содержании высокомолекулярного HSP 101 и низкомолекулярного HSP 17.6 в послепожарной хвое по отношению к контрольной.
Рис. 3.
Изменение содержания HSP 101 (а) и HSP 17.6 (б) в хвое сосны обыкновенной в контрольном образце и после искусственного выжигания. Приведено изображение типичной мембраны после визуализации антител.
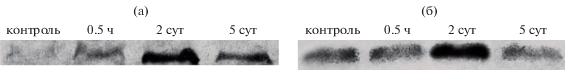
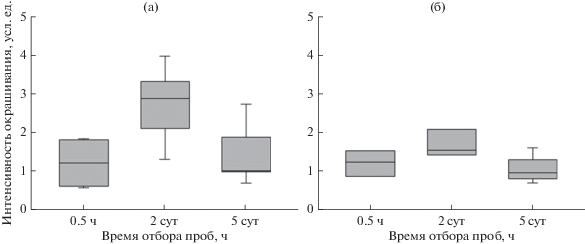
Рис. 4.
Денситометрическая оценка интенсивности окрашивания пятен белков HSP 101 (а) и HSP 17.6 (б) в хвое сосны обыкновенной в контрольном образце и после искусственного выжигания. На рис. 4 звездочкой обозначено значение, которое значимо отличаются от остальных образцов при p < 0.05.
Сведений о содержании HSP в хвое голосеменных крайне мало. Однако имеются данные, что с небольшими количественными изменениями HSP 101 присутствует в клетке постоянно, а HSP 17.6 накапливается преимущественно в теплое время года (Korotaeva et al., 2012). Это согласуется с полученными нами результатами для контрольных образцов (рис. 3, 4).
Как видно на рис. 4, через 30 мин накопление HSP 17.6 было довольно слабым, хотя согласно литературным данным, воздействия температурами от 40 до 70°C должно быть достаточно для накопления HSP в растительной ткани. Такое слабое накопление может говорить либо о кратковременности воздействия, недостаточной для активации защитных механизмов на уровне белкового синтеза, либо о сильных барьерных свойствах наружных структур хвои. Сосна относится к группе ксерофитов, хвоя которой имеет ксероморфное строение и отличается усиленным слоем наружных защитных тканей (кутикула, гиподерма) (Загирова, 1999), которые хорошо предохраняют мезофилл от внешних воздействий. С другой стороны, в работе Н.Е. Судачковой с соавт. (2015) по оценке влияния контролируемого выжигания, и, как следствие, теплового шока на древостой приведены экспериментальные данные о проявлении окислительного стресса в прикамбиальной зоне деревьев. В качестве первичной реакции на пирогенный стресс было отмечено повышение содержания пероксида водорода, снижение содержания низкомолекулярных углеводов и активности большинства ферментов антиоксидантной системы. Так, авторами приводятся результаты, указывающие, что содержание Н2О2 в участках прикамбиальной зоны модельных деревьев сосны с различными признаками повреждения оказалось выше от 2 до 8 раз относительно содержания в тканях неповрежденных деревьев через 3 ч после пожара. Такое смещение в прооксидантно-антиоксидантном равновесии может характеризовать одно из ключевых звеньев начала стрессовой реакции, которое приводит к запуску защитных механизмов, в том числе, синтезу специфических белков теплового шока. Таким образом, возможно, выбранный в данной работе интервал для отбора проб после горения (30 мин) оказался слишком коротким, чтобы зафиксировать развитие стрессового ответа в виде накопления HSP.
Наибольший рост содержания стрессовых белков относительно контроля отмечен в пробах, взятых через 2 сут после воздействия. При этом для HSP 101 медиана выборки значений содержания этого белка относительно контроля, принятого за 1, была на уровне 3 и значимо отличалась от значений содержания белка через 0.5 ч и 5 сут; для HSP 17.6 на вторые сутки она равнялась 1.5; для обоих белков 100% значений оказались выше 1. Подобное накопление могло быть следствием продолжительного тления подстилающей поверхности. В результате происходил прогрев поверхностного горизонта почвы и корней, а также живых клеток камбия, что отразилось на состоянии организма в целом. В этом случае экспрессия генов стрессовых белков может быть не результатом воздействия непосредственно на хвою, а следствием изменения физиологического статуса дерева в целом, которое берет начало от изменений, происходящих в стволе и корнях. С другой стороны, причиной позднего накопления HSP могла быть отсроченная по времени экспрессия генов стрессовых белков, возможно, связанная с особенностями объекта исследования, например, высоким содержанием защитных тканей хвои сосны. Для выяснения причины максимального накопления HSP не сразу, а через некоторое время после воздействия необходимы дальнейшие исследования.
По результатам наших экспериментов содержание НSP на 5-е сут существенно не отличалось от его содержания спустя 30 мин после выжигания (для HSP 101) и от содержания в контрольном образце (для HSP 17.6). Сокращение количества стрессовых белков на 5-е сут связано, по-видимому, с прекращением или снижением стрессового давления. Такой результат согласуется с данными о том, что уровень Н2О2 снижался во всех участках тканей побуревших деревьев через 5 дней после пожара (Судачкова и др., 2015). С другой стороны, сохранение на 5-е сут относительного количества HSP 101 на уровне выше контрольного может говорить в пользу сохранения стрессирующего эффекта либо продолжения восстановительных процессов, для которых наличие стрессового белка может быть необходимо.
Ранее (Коротаева и др., 2017) нами было проведено исследование оценки содержания стрессовых белков в хвое сосны обыкновенной, сформировавшейся в период восстановления после экспериментального выжигания. Проведенные исследования выявили различия в содержании белков теплового шока. Так, содержание стрессового белка HSP 101 в хвое с древостоя, пережившего пожар несколько лет назад, превышало содержание белка в контрольной хвое. Однако причиной такого накопления не могло быть термическое влияние пожара на хвою, поскольку она сформировалась после огневого воздействия. Подобные изменения отражали физиологический отклик поврежденного древостоя в постпирогенный период. Результаты экспериментального выжигания 2017 г., изложенные в настоящей статье, определенно говорят в пользу накопления в хвое стрессовых белков в ответ на огневое воздействие.
Заключение. В ходе проведенного контролируемого выжигания, имитирующего низовой пожар слабой силы, установлено, что температуры, превышающие физиологический оптимум произрастания сосны обыкновенной (выше 30°С), оказывали в течение нескольких минут термическое воздействие, приводящее к обгоранию кроны и стволов деревьев, тлению лесной подстилки. Продолжительность влияния температур с максимальными значениями от 48 до 76°С составляла от 5.5 до 11.8 мин, их средние значения равнялись 46 ± 7°С. Содержание стрессовых белков HSP 101 и HSP 17.6 значимо не отличалось от контрольного уровня через 30 мин и на 5-е сут после выжигания и достигало максимальных значений на 2-е сут после выжигания, при этом оно в 0.5–3 раза превышало контрольные значения. Таким образом, проведенное исследование впервые выявило признаки стрессового состояния древостоя в виде накопления HSP в краткосрочном послепожарном периоде и позволило сопоставить температурный диапазон огневого воздействия с динамикой и степенью накопления стрессовых белков в хвое.
Список литературы
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Бузыкин А.И., Попова Ф.П., Пшеничникова Л.С. Продуктивность сосновых лесов. М.: Наука, 1978. 230 с.
Валендик Э.Н. Ветер и лесной пожар. М.: Наука, 1968. 118 с.
Валендик Э.Н., Косов И.В. Влияние теплового излучения лесного пожара на окружающую среду // Сибирский экологический журн. 2008. № 4. С. 517–523.
Валиуллина Р.Н., Рябовол В.В., Хохлова Л.П. Изменение экспрессии генов белков теплового шока в связи с разной устойчивостью растений к повышенной температуре // Доклады академии наук. 2008. Т. 422. № 6. С. 845–847.
Верховец С.В. Влияние контролируемых выжиганий на пожароопасность и лесовосстановление на сплошных вырубках: Дисс. … канд. с-х. наук (спец. 06.03.03). Красноярск: Сибирский гос. техн. университе, 2000. 200 с.
Горшков В.В., Ставрова Н.И., Тарасова В.Н. Повреждение деревьев сосны обыкновенной и древесного яруса сосновых лесов европейского севера в результате пожаров // Лесоведение. 2004. № 5. С. 10–19
Дымов А.А., Дубровский Ю.А., Габов Д.Н., Жангуров Е.В., Низовцев Н.А. Влияние пожара в северотаежном ельнике на органическое вещество почвы // Лесоведение. 2015. № 1. С. 52–62.
Загирова С.В. Структура ассимиляционного аппарата и СО2-газообмен у хвойных. Екатеринбург: Изд-во УрО РАН, 1999. 108 с.
Иванова Г.А., Конард С.Г., Макрае Д.Д. Воздействие пожаров на компоненты экосистемы среднетаежных сосняков Сибири. Новосибирск: Наука, 2014. 232 с.
Иванова Г.А., Иванов В.А., Ковалева Н.М., Конард С., Жила С.В., Тарасов П.А. Сукцессия растительности после высокоинтенсивного пожара в сосняке лишайниковом // Сибирский экологический журн. 2017. Т. 24. № 1. С. 61–71.
Коротаева Н.Е., Гетте И.Г., Косов И.В., Пахарькова Н.В., Боровский Г.Б. Белки теплового шока и фотосинтетическая активность хвои сосны обыкновенной в постпирогенный период // Вестник КрасГАУ. 2017. № 10. С. 79–87.
Креславский В.Д., Карпентиер Р., Климов В.В., Мурата Н., Аллахвердиев С.И. Молекулярные механизмы устойчивости фотосинтетического аппарата к стрессу // Биологические мембраны. 2007. Т. 24. № 3. С. 195–217.
Кулаева О.Н. Белки теплового шока и устойчивость растений к стрессу // Соросовский образовательный журн. 1997. № 2. С. 5–13.
Курбатский Н.П. Техника и тактика тушения лесных пожаров. М.: Гослесбумиздадт, 1962. 154 с.
Курбатский Н.П. Исследование количества и свойств лесных горючих материалов // Вопросы лесной пирологии: Сб. статей, Красноярск, 1970. С. 5–58.
Масягина О.В., Прокушкин С.Г., Иванова Г.А. Влияние пожаров на интенсивность дыхания ствола сосны обыкновенной (Pinus sylvestris L.) // Хвойные бореальной зоны. 2007. Т. 24. № 1. С. 82–91.
Побединский А.В. Изучение лесовосстановительных процессов. М.: Наука, 1966. 60 с.
Суворова Г.Г. Фотосинтез хвойных деревьев в условиях Сибири. Новосибирск: ГЕО, 2009. 195 с.
Судачкова Н.Е., Милютина И.Л., Романова Л.И., Косов И.В., Собачкин Д.С. Воздействие низовых пожаров на жизнеспособность и антиоксидантную защиту молодняков сосны обыкновенной в красноярской лесостепи // Лесоведение. 2015. № 2. С. 16–25.
Судачкова Н.Е., Романова Л.И., Астраханцева Н.В., Новоселова М.В., Косов И.В. Стрессовые реакции деревьев сосны обыкновенной наповреждение низовым пожаром // Сибирский экологический журн. 2016. № 5. С. 739–749.
Сукачев В.Н. Руководство к исследованию типов леса // Избранные труды. 1972. Т. 1. С. 15–141.
Фуряев В.В., Фуряев Е.А. Пироэкологические свойства сосны обыкновенной в Средней Сибири // Хвойные бореальной зоны. 2008. № 1–2. С. 103–108.
Цветков П.А., Буряк Л.В. Исследование природы пожаров в лесах Сибири // Сибирский лесной журн. 2014. № 3. С. 25–42.
Швиденко А.З., Щепащенко Д.Г. Климатические изменения и лесные пожары в России // Лесоведение. 2013. № 5. С. 50–61.
Adams A.S., Rieske L.K. Prescribed fire affects white oak seedling phytochemistry: implications for insect herbivory // Forest Ecology &Management. 2003. V. 176. № 1–3. P. 37–47.
Al-Whaibi M.H. Plant heat-shock proteins: A mini review // J. King Saud University – Science. 2011. V. 23. P. 139–150.
Carlo N.J., Renninger H.J., Clark K.L., Schäfer K.V. Impacts of prescribed fire on Pinus rigida Mill. in upland forests of the Atlantic Coastal Plain // Tree Physiology. 2016. V. 36. P. 967–982.
Downs C.A., Heckathorn S.A., Bryan J.K., Coleman J.S. The methionine-rich low-molecular-weight chloroplast heat-shock protein: evolutionary conservation and accumulation in relation to thermotolerance // American J. Botany. 1998. V. 85. P. 175–183.
Fleck I., Grau M.D., Sanjose D. Vidal Influence of fire and tree-fell on physiological parameters in Quercus ilex resprout // Annals of Forest Science. 1996. V. 53. № 2–3. P. 337–348.
Heckathorn S.A., Downs C.A., Sharkey T.D., Coleman J.S. The small, methioninerich chloroplast heat-shock protein protects photosystem II electron transport during heat stress // Plant Physiology. 1998. V. 116. P. 439–444.
Korotaeva N.E., Oskorbina M.V., Kopytova L.D., Suvorova G.G., Borovskii G.B., Voinikov V.K. Variations in the content of stress proteins in the needles of common pine (Pinus sylvestris L.) within an annual cycle // J. Forest Research. 2012. V. 17. № 1. P. 89–97.
Kukavskaya E.A., Ivanova G.A., Conard S.G., McRae D.J., Ivanov V.A. Biomass dynamics of central Siberian Scots pine forests following surface fires of varying severity // International J. Wildland Fire. 2014. V. 23. № 6. P. 872–876.
Lee G.J., Vierling E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein // Plant Physiology. 2000. V. 122. P. 189–197.
McRae D.J., Alexander M.E., Stocks B.J. Measurement and description of fuels and fire behavior on prescribed burns. Ontario: Canadian Forest Service Publications, 1979. 44 p.
Mogk A., Schlieker C., Friedrich K.L., Schonfeld H.J., Vierling E., Bukau B. Refolding of substrates bound to small HSPs relies on a disaggregation reaction mediated most efficiently by ClpB/DnaK // The J. Biological Chemistry. 2003. V. 278. P. 31 033–31 042.
Neta-Sharir I., Isaacson T., Lurie S., Weissa D. Dual role for tomato heat shock protein 21: protecting photosystem II from oxidative stress and promoting color changes during fruit maturation // Plant Cell. 2005. V. 17. P. 1829–1838.
Queitsch C., Hong S.W., Vierling E., Lindquist S. Heat shock protein 101 plays a crucial role in thermotolerance in Arabidopsis // Plant Cell. 2000. V. 12(4). P. 479–492.
Renninger H.J., Clark K.L., Skowronski N., Schäfer K.V.R. Effects of a prescribed fire on water use and photosynthetic capacity of pitch pines // Trees. 2013. № 27. P. 1115–1127.
Rieske L.K. Wildfire alters oak growth, foliar chemistry, and herbivory // Forest Ecology & Management. 2002. V. 168. № 1–3. P. 91–99.
Sm$\hat {y}$kal P., Malin J., Hrdy I., Konopasek I., Zarsky V. Chaperone activity of tobacco HSP18, a small heat-shock protein, is inhibited by ATP // Plant J. 2000. V. 23. P. 703–713.
Timperio A.M., Egidi M.G., Zolla L. Proteomics applied on plant abiotic stresses: role of heat shock proteins (HSP) // J. Proteomics. 2008. № 71. P. 391–411.
Varner J.M., Putz F.E., O’Brien J J., Hiers J.K., Mitchell R.J., Gordon D.R. Post-fire tree stress and growth following smoldering duff fires // Forest Ecology & Management. 2009. № 258. P. 2467–2474.
Wang W., Vinocur B., Shoseyov O., Altman A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response // Trends in Plant Science. 2004. V. 9(5). P. 244–252.
Xu Y., Zhan C., Huang B. Heat Shock Proteins in Association with Heat Tolerance in Grasses // International J. Proteomics. 2011. V. 2011. P. 1–11.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение