

Лесоведение, 2020, № 3, стр. 239-249
Сезонная изменчивость эмиссии диоксида углерода при разложении елового валежа южной тайги Валдая
М. Л. Гитарский a, *, Д. Г. Замолодчиков b, В. А. Мухин c, Д. К. Диярова c, В. А. Грабар d, Д. В. Карелин b, e, А. И. Иващенко f, А. С. Марунич g
a Филиал “Угреша” Государственного университета “Дубна”
140090 Московская обл., Дзержинский, ул. Академика Жукова, д. 24, Россия
b Центр экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, д. 84/32, стр. 14, Россия
c Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, д. 202, Россия
d Институт глобального климата и экологии им. акад. Ю.А. Израэля
107258 Москва, ул. Глебовская, д. 20Б, Россия
e Институт географии РАН
119017 Москва, Старомонетный переулок, д. 29, стр. 4, Россия
f Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, д. 1, Россия
g Валдайский филиал Государственного гидрологического института
175400 Валдайский р-н, Новгородская обл., Валдай, ул. Победы, д. 2, Россия
* E-mail: m.gitarskiy@gmail.com
Поступила в редакцию 05.09.2019
После доработки 08.11.2019
Принята к публикации 29.01.2020
Аннотация
Сезонная изменчивость эмиссии диоксида углерода (СО2) при микогенном разложении елового валежа исследована в старовозрастном ельнике и окне распада елового древостоя. Потоки СО2 с поверхности валежа измеряли камерным методом. Видовой состав, встречаемость и относительное обилие дереворазрушающих грибов определяли по данным маршрутных учетов. В комплексах дереворазрушающих грибов доминируют Fomitopsis pinicola (Sw.) P. Karst., Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden и Fomitopsis rosea (Alb. & Schwein.) P. Karst. Они ответственны за разложение 81% елового валежа и сухостоя. Эмиссионая активность от древесного валежа характеризуется температурно обусловленным годичным циклом: в обоих биотопах ее начало соответствует марту–апрелю, максимальная величина наблюдается в мае, а затем происходит снижение до минимальных значений в октябре–ноябре. При диапазоне среднемесячных температур 3.5–16.7°C, минимум измеренного эмиссионного потока от древесного валежа 22.9 г C-СО2 м–2 мес.–1 установлен в ельнике в октябре, а максимум – 80.4 г C-СО2 м–2 мес.–1 – в окне распада в мае. Период эмиссионной активности с марта по ноябрь можно приближенно принять за продолжительность годовой эмиссии СО2 в южнотаежной зоне. Полученные результаты повышают точность оценок потока СО2 при разложении древесного валежа.
Мертвая древесина (древесный дебрис) является значимым элементом глобального биогеохимического цикла углерода и играет определяющую роль в обмене органического вещества и энергии в лесных экосистемах (Harmon et al., 1986; Кудеяров и др., 2007; Мухортова, Ведрова, 2012; Стороженко, 2012). Древесный дебрис представляет собой огромный резервуар биогенного углерода, величина которого составляет 73 ± 6 Pg С, или около 8% запаса углерода в лесах мира (Pan et al., 2011). В спелых и перестойных лесах его запас составляет от 10 до 20% (Turner et al., 1995; Brown, 2002), что позволяет считать мертвую древесину вторым по величине, после лесной фитомассы, наземным резервуаром углерода, обеспечивающим его накопление и длительное хранение в лесах бореальной зоны (Кудеяров и др., 2007).
При этом древесный дебрис является весьма лабильным резервуаром углерода, открытым как для поступления, так и для потерь в результате выщелачивания, механического разрушения, дыхания и биохимической трансформации органического вещества. Газообразные потери в форме СО2 в результате дыхания являются основной расходной частью пула углерода мертвой древесины и премущественно регулируются деятельностью дереворазрушающих грибов–ксилотрофов (Harmon et al., 1986; Moore et al., 2004; Zhou et al., 2007). Согласно ряду оценок, в бореальных лесах ксилотрофные грибы ответственны за более 75% потока диоксида углерода, выделяющегося в атмосферу при микогенном разложении древесного дебриса (Бурова, 1986; Мухин, Воронин, 2007; Мухин и др., 2010). Эмиссионная активность дереворазрушающих грибов зависит от условий окружающей среды – температуры и влажности воздуха, физико-химических свойств субстрата, определяемых древесной породой, а также от стадии ее разложения (Progar et al., 2000; Herrmann, Bauhus, 2013; Мухин и др., 2015; Kahl et al., 2015; Гитарский и др., 2017). В литературе содержатся сведения о многократном увеличении интенсивности эмиссии СО2 с ростом температуры окружающей среды на каждые 10°С (Chen et al., 2000; Herrmann, Bauhus, 2013; Kahl et al., 2015). В большинстве случаев подобные зависимости были установлены в ходе лабораторных экспериментов (Chen et al., 2000; Wang et al., 2002; Herrmann, Bauhus, 2013).
Между тем исследования эмиссионной активности крупных древесных остатков в естественных условиях умеренной климатической зоны сравнительно немногочисленны (см., например, Молчанов и др., 2011; Сафонов и др., 2012; Kahl et al., 2015; Гитарский и др., 2017; Иванов и др., 2018). В свою очередь, быстрый рост или падение температуры воздуха на 10°С и более в естественной среде – явление редкое и маловероятное. Зимой, в период низких температур и залегания устойчивого снегового покрова, дыхательная активность ксилотрофных грибов и, соответственно, эмиссионный поток диоксида углерода от древесного дебриса в бореальных лесах практически полностью прекращаются (Бурова, 1986; Мухин, Диярова, 2012), что позволяет говорить об его сезонной цикличности, которая до настоящего времени исследована недостаточно. Для восполнения наметившегося пробела мы провели исследование сезонной изменчивости СО2-эмиссионной активности крупного древесного дебриса в естественных условиях южной тайги.
ОБЪЕКТЫ И МЕТОДИКА
Исследования выполнены в национальном парке “Валдайский” (Новгородская область, Валдайский район, 57°57.76′ с.ш., 33°20.34′ в.д., 218 м над ур. моря). По своему ботанико-географическому положению объект находится на границе южной тайги. Район исследований относится к восточно-европейским таежным ландшафтам (Исаченко, Шляпников, 1989) и располагается на холмисто-моренной возвышенности в области валдайского оледенения. Согласно рядам наблюдений на метеостанции “Валдай” (Научно-прикладной …, 1988), среднемноголетняя температура воздуха составляет 3.4°C, годовое количество осадков – 709 мм. Наиболее холодным месяцем в регионе является январь, средняя многолетняя температура которого –9.9°C, наиболее теплым – июль – с 16.7°C. Формирование устойчивого снегового покрова приходится на конец ноября, сход снега – конец второй декады апреля. Продолжительность безморозного периода по разным оценкам составляет 128–130 дней (Научно-прикладной …, 1988; Коротков, 1991).
Исследования СО2-эмиссионной активности проводили на крупном валеже ели, представленном упавшими на землю стволами разной степени разложения. Валежные фрагменты были расположены в типичных лесных биотопах южной тайги – старовозрастном (110-летнем) ельнике мелкотравно-зеленомошном Piceeta parviherboso-hylocomiosa (9Е1С, средний диаметр 37 см, полнота 0.4) и окне распада елового древостоя, представляющем собой сукцессионно-возрастную трансформацию елового древостоя. Согласно эколого-флористической классификации, старовозрастный ельник относится к ассоциации Maianthemo bifoliae – Piceetum abietis. Окно распада занято рябинником крупнопапоротниково-малиновым на месте выпавшего в 2012–2013 гг. елового древостоя, имевшего средний диаметр 28 см. В отсутствие древесного яруса подрост и подлесок имеют среднюю высоту 4.5 м и сомкнутость крон 0.4: доминирует рябина обыкновенная, участие других видов (ель, береза, бузина и крушина) незначительное. В исследуемых биотопах были проведены учеты комплексов дереворазрушающих грибов и измерены потоки эмиссии СО2 в атмосферу с поверхности валежа ели.
Оценку видового состава дереворазрушающих грибов производили в 2015–2016 гг. методом маршрутных учетов (Толмачев, 1974). Встречаемость определяли подсчетом числа валежных и сухостойных елей с найденными на них визуально диагностируемыми базидиокарпами ксилотрофных грибов одного вида, а их и относительное обилие – по отношению встречаемости грибов одного вида к общему числу обнаруженных дереворазрушающих грибов. По данным учетов был установлен видовой состав комплекса ксилотрофных грибов елового дебриса. Номенклатура грибов приведена по Mycobank Database (http://www.mycobank.org).
Исследования эмиссионного потока СО2 выполняли методом закрытых камер на случайно выбранных крупномерных фрагментах елового валежа разных стадий разложения. Фрагменты имели диаметр, близкий к среднему диаметру древостоя исследуемых биотопов. В верхнюю поверхность валежной древесины врезали непрозрачные открытые ПВХ-камеры цилиндрической формы. Камеры имели диаметр 10 см и высоту 9–15 см. Их внутренние объемы были определены перед началом измерений потоков СО2. С двух диаметрально противоположных сторон в камеры были врезаны штуцеры для герметичного крепления воздуховодов. Для обеспечения репрезентативности измерений на крупных древесных остатках ели было установлено в общей сложности 29 камер: 16 – в старовозрастном ельнике и 13 – в окне распада древостоя (рис. 1).
Рис. 1.
Еловый валеж в старовозрастном ельнике (а) и окне распада древостоя (б) с камерами для оценки эмиссионного потока СО2.

Измерения потоков диоксида углерода проводили ежемесячно, с апреля по октябрь 2015–2017 гг. Поток эмиссии СО2 определяли по изменению его концентрации в камерах портативным полевым газоанализатором, разработанным на биологическом факультете МГУ им. М.В. Ломоносова на базе инфракрасного СО2 – сенсора AZ 7752 (AZ Instrument Corp., Тайвань) и воздушной помпы Е134–11–120 (Hargraves Thechnologies Corp., США). Сенсор AZ 7752 имеет разрешающую способность 1 ppm (млн–1) и диапазон измерений CO2 0–2000 ppm. Мощность воздушной помпы Е134–11–120 – 2 л мин–1 (рис. 2).
Рис. 2.
Измерение эмиссии СО2 от крупных древесных остатков в старовозрастном ельнике анализатором на основе сенсора AZ 7752.

Во время измерений камеры, постоянно установленные на поверхности древесного дебриса, герметично закрывали крышкой и соединяли с газоанализатором полиуретановыми воздуховодами при помощи штуцеров. Объем замкнутой измерительной системы V складывался из суммы объемов камеры, воздуховодов и измерительной камеры сенсора. Продолжительность измерения составляла 3 мин. При каждом измерении, после околоминутного ожидания, фиксировали начальное и, через 3 мин, конечное значения концентрации СО2 в замкнутой системе. Результаты поверяли одновременными измерениями СО2, выполненными при помощи высокоточного инфракрасного газоанализатора LI-8100A (LiCor, Небраска, США) с разрешением 0.1 ppm. Поверки показали, что результаты оценок потоков, выполненных этими приборами, на одних и тех же камерах различаются не более чем на 5%. Величины потоков С-СО2 с поверхности древесного валежа рассчитывали на основе уравнения Менделеева—Клапейрона:
Отнесением DC к времени измерения и площади основания камеры получали величину интенсивности удельной эмиссии углерода от крупных древесных остатков ели (г C-СО2 м–2 мин–1). На основе отдельных измерений определяли средние величины месячных потоков СО2. Для средних месячных величин находили квадратичную функцию зависимости от порядкового номера месяца. Интегральный поток за весь сезон рассчитывали как площадь под параболой квадратичной функции в пределах ее пересечений с осью абсцисс.
При расчете годового потока величинами зимней эмиссии от валежа (период с устойчивыми отрицательными температурами) пренебрегали. За трехлетний период на крупных древесных остатках разных стадий разложения было выполнено в общей сложности 555 измерений в старовозрастном ельнике и 365 – в окне распада.
Данные по температуре воздуха, осадкам и атмосферному давлению брали по метеостанции “Валдай”, ближайшей к объекту исследований (в 4 км). Достоверность различий средних величин определяли с помощью непараметрического теста Манна—Уитни в статистическом пакете SPSS-20 (IBM Company, США). Связи между среднесуточной интенсивностью эмиссии СО2 и значимыми факторами окружающей среды оценивали с помощью корреляции Пирсона. Если в течение суток в одних и тех же камерах было выполнено несколько измерений эмиссии СО2, в анализе использовали среднесуточную величину.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В таблице 1 приведены результаты учета встречаемости визуально диагностируемых видов комплекса ксилотрофных грибов на сухостое ели. Как видно из приведенных в ней результатов выполненных микологических учетов, в комплексах дереворазрушающих грибов лишь три вида отличаются относительно высоким обилием – Fomitopsis pinicola (Sw.) P. Karst. (38 найденных базидиокарпов, 52.0%), Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden (15 находок, 20.5%) и Fomitopsis rosea (Alb. & Schwein.) P. Karst. (6 находок, 8.2%). Они принимают участие в разложении 81% елового валежа и сухостоя в пределах территории объекта исследования.
Таблица 1.
Видовой состав комплекса ксилотрофных грибов–деструкторов елового валежа и сухостоя в старовозрастном ельнике и в окне распада
| № | Вид | Встречаемость, шт. | Относительное обилие, % |
|---|---|---|---|
| 1 | Amylocystis lapponica (Romell) Bondartsev & Singer ex Singer | 1 | 1.4 |
| 2 | Dacrymyces sp. | 5 | 6.8 |
| 3 | Entoloma byssisedum (Pers.) Donk | 1 | 1.4 |
| 4 | Exidia saccharina (Alb. & Schwein.) Fr. | 1 | 1.4 |
| 5 | Fomitopsis pinicola (Sw.) P. Karst. | 38 | 52.0 |
| 6 | Fomitopsis rosea (Alb. & Schwein.) P. Karst. | 6 | 8.2 |
| 7 | Gloeophyllum odoratum (Wulfen) Imazeki | 1 | 1.4 |
| 8 | Neoantrodia serialis (Fr.) Audet | 1 | 1.4 |
| 9 | Pycnoporellus fulgens (Fr.) Donk | 4 | 5.5 |
| 10 | Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden | 15 | 20.5 |
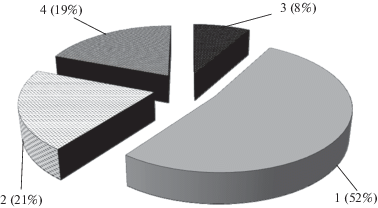
Аналогичный комплекс доминирующих ксилотрофных грибов – деструкторов елового дебриса описан для среднетаежных, южнотаежных и подтаежных ельников Западной Сибири. Доминантом в них является Fomitopsis pinicola, субдоминанты – Trichaptum abietinum и Fomitopsis rosea – наиболее распространенные деструкторы древесных остатков ели сибирской (Мухин, 1993). Значительно реже на еловом дебрисе в пределах территории национального парка “Валдайский” встречаются Amylocystis lapponica (Romell) Bondartsev & Singer ex Singer, Dacrymyces sp., Entoloma byssisedum (Pers.) Donk, Exidia saccharina (Alb. & Schwein.) Fr., Gloeophyllum odoratum (Wulfen) Imazeki, Neoantrodia serialis (Fr.) Audet, Pycnoporellus fulgens (Fr.) Donk. Суммарно они были обнаружены 14 раз, и их относительная доля составляет около 19.0% (рис. 3).
Рис. 3.
Дереворазрушающие грибы–деструкторы елового дебриса на территории национального парка “Валдайский” (в скобках приведено относительное обилие отдельных видов грибов): 1 – Fomitopsis pinicola (Sw.) P. Karst., 2 – Trichaptum abietinum (Dicks.) Ryvarden, 3 – Fomitopsis rosea (Alb. & Schwein.) P. Karst., 4 – другие виды.
На рис. 4 приведена динамика среднемесячного удельного потока С-СО2 от крупных древесных остатков ели разной степени разложения в старовозрастном ельнике (а) и в окне распада елового древостоя (б). По существу, на графике показан сезонный цикл активности комплекса дереворазрушающих грибов ели, полученный по данным трехлетних наблюдений. Если предположить, что найденная квадратичная функция корректно отражает динамику годовой эмиссии, можно заключить, что активность ксилотрофных грибов начинает интенсивно увеличиваться в марте и прекращается в ноябре.
Рис. 4.
Сезонная динамика потока С-СО2 с поверхности крупных древесных остатков ели (1) и температур воздуха (2) в старовозрастном ельнике (а) и окне распада елового древостоя (б). Приведены среднемесячные значения и их стандартные ошибки, а также аппроксимация потоков СО2 квадратичной функцией.
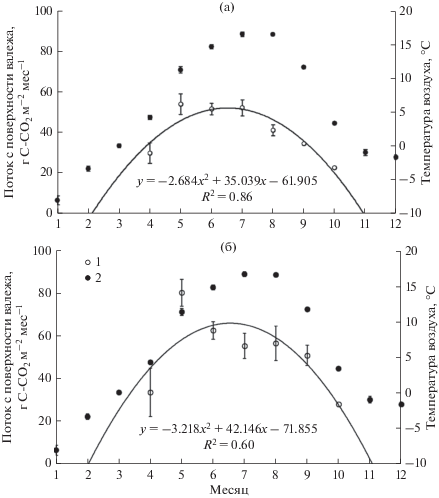
При рассмотрении ряда независимых переменных (количество осадков, температура древесины, температура воздуха) сильная положительная связь была выявлена только между эмиссионной активностью валежа и температурой воздуха. С допущением близких к нулю значений эмиссии в марте и ноябре коэффициент корреляции Пирсона между среднемесячными величинами потока СО2 и среднемесячной температурой составил rp = +0.94 (Р < 0.0001, N = 9) в ельнике и +0.73 (Р = = 0.025, N = 9) в окне распада древостоя.
Будучи основными деструкторами древесины, дереворазрушающие грибы являются уникальной биосферно-значимой группой организмов (Заварзин, 2006; Мухин, Воронин, 2007). Поскольку идентифицированные на крупных древесных остатках ксилотрофные грибы Fomitopsis pinicola, F. rosea и Trichaptum abietinum (рис. 3) доминируют в сходных типах леса и аналогичном климате в других регионах страны, можно сделать заключение, что этот комплекс дереворазрушающих грибов является типичным для южнотаежных еловых лесов России (Мухин, 1993). Вероятно, их можно считать ответственными за основную часть эмиссионного потока С-СО2 при разложении древесных остатков в темнохвойных лесах умеренной зоны.
СО2-эмиссионая активность древесного дебриса при его микогенном разложении в условиях умеренных широт имеет температурно-обусловленный годичный цикл. Как правило, он включает 4 периода, соответствующие четырем временам года. В частности, в зонально близких южнотаежным лесам подтаежных лесах Среднего Урала зимний период характеризуется устойчиво отрицательными среднесуточными температурами воздуха с ноября по февраль, и в это время СО2–эмиссионная активность древесных остатков пренебрежимо мала. Весенний период начинается в марте и характеризуется наличием температурно-обусловленных пауз в суточной эмиссии СО2. Переход же к круглосуточной эмиссии диоксида углерода означает начало летнего периода: май–сентябрь. Появление температурно-обусловленных пауз в суточной эмиссии СО2 в октябре – это начало осеннего периода (Мухин, Диярова, 2012).
Погодные условия в период наших исследований отличались от среднемноголетних данных (Научно-прикладной …, 1988; Коротков, 1991). Период устойчивых среднемесячных отрицательных температур, ранее составлявший 5 месяцев (Научно-прикладной …, 1988), в 2015–2017 гг. сократился до 4 месяцев (с ноября по февраль), что, по-видимому, связано с общей тенденцией к потеплению, наблюдающейся в последние десятилетия на территории Российской Федерации (Доклад …, 2019). Согласно полученной квадратичной аппроксимации данных эмиссии СО2 от древесного дебриса, его эмиссионная активность в обоих биотопах прекращалась к ноябрю и возобновлялась в марте (рис. 4). Таким образом, длительность периода низких температур, когда эмиссионная активность древесного валежа отсутствует, в целом согласуется с данными наблюдений в лесах Среднего Урала (Мухин, Диярова 2012), также как и предполагаемая продолжительность внутригодовой активности дереворазрушающих грибов. В бореальных лесах умеренной климатической зоны с длительностью зимнего периода от 4 мес. и более общая интенсивность разложения мертвой древесины группой грибов–ксилотрофов зависит, прежде всего, от их активности в безморозный период. Поэтому установленный период эмиссионной активности с марта по ноябрь, по нашему мнению, можно приближенно принять за продолжительность годовой эмиссии диоксида углерода в результате деятельности комплекса дереворазрушающих грибов южной тайги.
При попарных сравнениях среднемесячных значений эмиссии от валежа, в обоих изучаемых биотопах потоки с мая по июль значимо не отличаются (критерий Манна–Уитни, Р = 0.3–0.5). В всех остальных случаях отличия потоков в последовательные пары месяцев (апрель–май, июль–август, август–сентябрь, сентябрь–октябрь) значимы (критерий Манна–Уитни, Р < 0.05). Средние за весенние, летние и осенние сезоны значения потоков между сравниваемыми биотопами значимо отличаются (1.79 ± 0.08 (N = 281) > 1.48 ± 0.06 (N = = 486) г С-СО2 м–2 сут–1; Манн–Уитни, Р = 0.001). Однако в месячном масштабе эти различия оказались значимы только в мае, августе и сентябре (Манн–Уитни, Р < 0.05).
При статистической значимости внутрисезонной динамики потоков, которая определяется, в основном, температурой как ведущим фактором, нельзя не отметить заметную вариабельность среднемесячных оценок (рис. 4). Это обусловлено тем, что в массивах внутримесячных измерений усреднены значения эмиссионных потоков, полученных от крупных древесных остатков разных стадий разложения. Степень разложения древесины оказывает существенное влияние на наблюдаемую интенсивность эмиссионного потока СО2 с поверхности валежа. Различия в интенсивности эмиссии диоксида углерода от древесного дебриса разной степени разложения подробно рассмотрены в наших предыдущих работах (Сафонов и др., 2012; Гитарский и др., 2017). В настоящем исследовании данные помесячных измерений от отдельных стадий разложений объединены для выявления общих закономерностей сезонной динамики эмиссионного потока.
За трехлетний период наблюдений наибольшие среднесуточные значения температуры наблюдались в июле–августе. Тем не менее наибольших значений усредненный по стадиям разложения поток от дыхания поверхности валежа достигает в мае: 54.3 г C-СО2 м–2 мес.–1 в ельнике и 80.4 г C-СО2 м–2 мес.–1 в окне распада (рис. 4а, 4б). Различие средних значимо (критерий Манна–Уитни, Р = 0.019). Возможно, это связано с особенностями жизненной активности ксилотрофных грибов, имевших место в период наших исследований.
Более высокие по абсолютной величине и более сглаженные показатели сезонного потока диоксида углерода в окне распада елового древостоя обусловлены лучшей, чем под пологом леса, освещенностью и более высокой температурой открытой поверхности валежа. По-видимому, в летний период в окне распада создаются близкие к оптимуму условия жизнедеятельности ксилотрофных грибов. Аналогичное влияние микроклиматических факторов на скорость разложения валежа отмечено, в частности, в работе (Progar et al., 2000), авторы которой установили более высокую интенсивность дыхания древесных остатков на участках леса южных экспозиций.
Во время наших наблюдений при диапазоне среднемесячных температур от 3.5 до 16.7°C, абсолютные величины эмиссионного потока от древесного валежа изменялись от минимальных 22.9 г C-СО2 м–2 мес.–1 в октябре в ельнике до максимальных 80.4 г C-СО2 м–2 мес.–1 в мае в окне распада. После достижения максимальных значений в мае наблюдалось постепенное снижение эмиссии СО2 в обоих биотопах. При этом среднемесячная за трехлетний период температура воздуха в мае 11.4°C была ниже, чем аналогичные температуры июля (16.7°C) и августа (16.6°C). Таким образом, несмотря на установленную по нашим наблюдениям высокую корреляцию между дыханием валежа и температурой воздуха, в условиях южной тайги не подтверждается многократное увеличение интенсивности эмиссии СО2 с ростом температуры окружающей среды на каждые 10°C, часто отмечаемое в литературе (Chen et al., 2000; Li et al., 2007; Herrmann, Bauhus, 2013; Kahl et al., 2015). По всей вероятности, температура является не единственным главным фактором, определяющим интенсивность эмиссионного потока от крупных древесных остатков. Аналогичный вывод был сделан в работе (Bradford et al., 2014). Среди других предполагаемых факторов эмиссии СО2 из валежа могла быть влажность древесины, которая зависит от влажности воздуха. Возможно, майский максимум активности ксилотрофного грибного сообщества связан с оптимальным сочетанием температуры и влажности воздуха.
Полученные среднесуточные величины потока С-СО2 от древесных остатков хорошо согласуются с оценками А.Г. Молчанова с соавт. (2011) в близких лесорастительных условиях южной тайги, а также с оценками в старовозрастных хвойных лесах северо-запада США (Marra, Edmonds, 1994), что позволяет говорить о сходных условиях и закономерностях в разложении валежа в бореальных лесах. Однако мы считаем, что использование средних величин может приводить к смещению оценки годового потока С-СО2 от крупных древесных остатков в связи с высокой вариабельностью, связанной с составом валежа. Более корректной представляется оценка эмиссионного потока СО2 от отдельных стадий разложения крупных древесных остатков, как это было сделано в наших предыдущих работах (Сафонов и др., 2012; Гитарский и др., 2017). Такой подход позволяет точнее оценить площадной вклад дыхания валежа в общий углеродный баланс леса, если известен долевой запас различных стадий разложения валежа.
В нашем предыдущем исследовании оценка годовой эмиссии С-СО2 была выполнена исходя из средней длительности безморозного периода 128 дней, полученной по литературным данным (Коротков, 1991; Гитарский и др., 2017). Согласно проведенной аппроксимации, эмиссионная активность ксилотрофных грибов оказалась значительно продолжительней – с марта по ноябрь, что согласуется с данными исследований, выполненных в других регионах умеренной климатической зоны (Мухин, Диярова, 2012). Недоучет длительности наиболее активного периода жизнедеятельности дереворазрушающих грибов может привести к существенной недооценке их годовой эмиссионной активности. Поэтому для получения более точных площадных оценок потока С-СО2 от древесного дебриса, наряду с учетом его запасов по стадиям разложения, необходимо точно фиксировать фактические границы сезона с положительными температурами воздуха.
ЗАКЛЮЧЕНИЕ
В настоящее время старовозрастные леса, выбранные в качестве объектов исследования, преобладают в составе лесного фонда Российской Федерации. Ожидается, что в дальнейшем их доля в лесном фонде будет увеличиваться (Замолодчиков и др., 2013). Поэтому уточнение оценок вклада в углеродный бюджет лесов России эмиссионной составляющей крупных древесных остатков является важной задачей, значимость которой будет возрастать. Результаты наших исследований позволяют не только уточнить методологию оценок потока диоксида углерода от древесного дебриса, но и существенно повысить их точность. Оценки и закономерности, полученные для лесов южной тайги, могут быть применены в других лесах бореальной зоны, что позволит улучшить качество оценок их углеродного бюджета. Кроме того, происходящие изменения климата и связанные с ними процессы трансформации лесных экосистем и деградации древесного яруса диктуют необходимость более точного учета скорости деструкции древесного дебриса.
Список литературы
Бурова Л.Г. Экология грибов макромицетов. М.: Наука, 1986. 222 с.
Гитарский М.Л., Замолодчиков Д.Г., Мухин В.А., Грабар В.А., Диярова Д.К., Иващенко А.И. Поток углерода от валежа южнотаежных лесов Валдайской возвышенности // Экология. 2017. № 6. С. 447–453.
Доклад об особенностях климата на территории Российской Федерации за 2018 год. М.: Росгидромет, 2019. 79 с.
Заварзин Г.А. Углеродный баланс России // Возможности предотвращения изменения климата и его негативных последствий: проблема Киотского протокола: материалы Совета–семинара при президенте РАН / Под ред. Израэля Ю.А. М.: Наука, 2006. С. 134−151.
Замолодчиков Д.Г., Грабовский В.И., Коровин Г.Н., Гитарский М.Л., Блинов В.Г., Дмитриев В.В., Курц В.А. Бюджет углерода управляемых лесов Российской Федерации в 1990–2050 гг.: ретроспективная оценка и прогноз // Метеорология и гидрология. 2013. № 10. С. 73–92.
Иванов А.В., Браун М., Лошаков С.Ю., Потоцкий О.В., Замолодчиков Д.Г. Эмиссия углерода с поверхности валежа в кедровых лесах Южного Приморья // Экология. 2018. № 4. С. 275–281.
Исаченко А.Г., Шляпников А.А. Природа мира: ландшафты. М.: Мысль, 1989. 504 с.
Коротков К.О. Леса Валдая. М.: Наука, 1991. 157 с.
Кудеяров В.Н., Заварзин Г.А., Благодатский С.А., Борисов А.В., Воронин П.Ю., Демкин В.А., Демкина Т.С., Евдокимов И.В., Замолодчиков Д.Г., Карелин Д.В., Комаров А.С., Курганова И.Н., Ларионова А.А., Лопес де Гереню В.О., Уткин А.И., Чертов О.Г. Пулы и потоки углерода в наземных экосистемах России. М.: Наука, 2007. 315 с.
Молчанов А.Г., Татаринов Ф.А., Курбатова Ю.А. Эмиссия СО2 стволами живых деревьев и валежом в еловых лесах юго-запада Валдайской возвышенности. // Лесоведение. 2011. № 3. С. 14–25.
Мухин В.А. Биота ксилотрофных базидиомицетов Западно-Сибирской равнины. Екатеринбург: Наука, 1993. 230 с.
Мухин В.А., Воронин П.Ю. Микогенное разложение древесины и эмиссия углерода в лесных экосистемах // Экология. 2007. № 1. С. 24–29.
Мухин В.А., Диярова Д.К. Сезонная динамика конверсионной активности трутовых грибов // Грибные сообщества лесных экосистем. М.; Петрозаводск: Изд-во КарНЦ РАН, 2012. Т.3. С. 41–48.
Мухин В.А., Диярова Д.К., Веселкин Д.В. Влажность как фактор СО2-эмиссионной активности древесного дебриса // Лесоведение. 2015. № 3. С. 208–213.
Мухин В.А., Воронин П.Ю., Сухарева A.В., Кузнецов В.В. Грибное разложение древесины при потеплении климата в бореально-гумидной лесорастительной зоне // Доклады Академии наук. 2010. Т. 431. № 3. С. 423–425.
Мухортова Л.В., Ведрова Э.Ф. Вклад крупных древесных остатков в динамику запасов органического вещества послерубочных лесных экосистем // Лесоведение. 2012. № 6. С. 55–62.
Научно-прикладной справочник по климату СССР. Серия 3. Многолетние данные. Вып. 3. Ч. 1–6. Л.: Гидрометеоиздат, 1988. 693 с.
Сафонов С.С., Карелин Д.В., Грабар В.А., Латышев Б.А., Грабовский В.И., Уварова Н.Е., Замолодчиков Д.Г., Коротков В.Н., Гитарский М.Л. Эмиссия углерода от разложения валежа в южнотаежном ельнике // Лесоведение. 2012. № 5. С. 75–80.
Стороженко В.Г. Характеристика древесного отпада в коренных ельниках восточноевропейской тайги // Там же. 2012. № 3. С. 43–90.
Толмачев А.И. Введение в географию растений. Л.: Изд-во Ленинградского университета. 1974. 244 с.
Bradford M.A., Warren II R.J., Baldrian P., Crowther T.W, Maynard D.S., Oldfield E.E., Wieder W.R., Wood S.A., King J.R. Climate Fails to Predict Wood Decomposition at Regional Scales // Nature Climate Change. 2014. V. 4. № 7. P. 625–630.
Brown S. Measuring Carbon in Forests: Current Status and Future Challenges // Environmental Pollution. 2002. V. 116. № 3. P. 363–372.
Chen H., Harmon M.E., Griffiths R.P., Hicks W. Effects of Temperature and Moisture on Carbon Respired from Decomposing Woody Roots // Forest Ecology & Management. 2000. V. 138. P. 51–64.
Harmon, M.E., Franklin J.F., Swanson F.J., Sollins P., Gregory S.V., Lattin J.D., Anderson N.H., Cline S.P., Aumen N.G., Sedell J.R., Lienkaemper G.W., Cromack K., Cummins K.W. Ecology of Coarse Woody Debris in Temperate Ecosystems // Advances in Ecological Research. 1986. V. 15. P. 133–302.
Herrmann S., Bauhus J. Effects of Moisture, Temperature and Decomposition Stage on Respirational Carbon Loss from Coarse Woody Debris of Important European Tree Species // Scandinavian J. Forest Research. 2013. V. 28. № 4. P. 346–357.
Kahl T., Baber K., Otto P., Wirth C., Bauhus J. Drivers of CO2 Emission Rates from Dead Wood Logs of 13 Tree Species in the Initial Decomposition Phase // Forests. 2015. V. 6. P. 2484–2504.
Li Zh., Li-min D., Hui-yan G., Lei Zh. Review of the Decomposition and Influence Factors of Coarse Woody Debris in Forest Ecosystem // J. Forestry Research. 2007. V. 18. № 1. P. 48–54.
Marra J.L., Edmonds R.L. Coarse Woody Debris and Forest Floor Respiration in an Old-Growth Coniferous Forest on the Olympic Peninsula, Washington, USA // Canadian J. Forest Research. 1994. V. 24. № 9. P. 1811–1817.
Moore J.C., Berlow E.L., Coleman D.C., Ruiter de P.C., Dong Q., Hastings A., Johnson N.C., McCann K.S., Melville K., Morin P.J., Nadelhoffer K., Rosemond A.D., Post D.M., Sabo J.L., Scow K.M., Vanni M.J., Wall D.H. Detritus, Trophic Dynamics and Biodiversity // Ecology Letters. 2004. V. 7. № 7. P. 584–600.
Mycobank Database. Fungal Databases, Nomenclature & Species Banks [Электронный ресурс]. Режим доступа: http://www.mycobank.org, свободный. – Загл. с экрана. – Яз. англ. Дата обращения: 10.01.2020.
Pan Y., Birdsey R.A., Fang J., Houghton R., Kauppi P.E., Kurz W.A., Phillips O.L., Shvidenko A.Z., Lewis S.L., Canadell J.G., Ciais P., Jackson R.B., Pacala S.W, McGuire A.D., Piao S., Rautiainen A., Sitch S., Hayes D. A Large and Persistent Carbon Sink in the World’s Forests // Science. 2011. V. 333. Is. 6045. P. 988–993.
Progar R.A., Schowalter T.D., Freitag C.M., Morrell J.J. Respiration from Coarse Woody Debris as Affected by Moisture and Saprotroph Functional Diversity in Western Oregon // Oecologia. 2000. V. 124. № 3. P. 426–431.
Turner D.P., Koerper G.J., Harmon M.E., Lee J.J. A Carbon Budget for Forests of the Conterminous United States // Ecological Applications. 1995. V. 5. № 2. P. 421–436.
Wang C., Bond-Lamberty B., Gower S.T. Environmental controls on Carbon Dioxide Flux from Black Spruce Coarse Woody Debris // Oecologia. 2002. V. 132. № 3. P. 374–381.
Zhou L., Dai L., Gu H., Zhong L. Review on the Decomposition and Influence Factors of Coarse Woody Debris in Forest Ecosystem // J. Forestry Research. 2007. V. 18. № 1. P. 48–54.
Дополнительные материалы отсутствуют.