

Лесоведение, 2020, № 4, стр. 301-313
Вегетационный индекс NDVI как индикатор фотосинтетического потенциала бореальных лесов Волжского бассейна
Э. Г. Коломыц a, *, Л. С. Шарая a
a Институт экологии Волжского бассейна РАН
445003 Тольятти, ул. Комзина, 10, Россия
* E-mail: egk2000@mail.ru
Поступила в редакцию 03.02.2019
После доработки 15.06.2019
Принята к публикации 06.04.2020
Аннотация
По цифровым материалам спутникового зондирования лесного покрова России составлена региональная карта нормализованного разностного вегетационного индекса NDVI бореальных лесов Волжского бассейна. Проведен эколого-географический анализ индекса, характеризующего физиологические основы их продуктивности. Выполнен статистический анализ связей фотосинтетического потенциала трех групп бореальных лесов с климатом и равнинным рельефом, а также с продуктивностью лесных формаций. Определены соответствующие орографические и климатические условия максимальных и минимальных значений NDVI. Выявлена структура экологических пространств растительности, продуктивности и вегетационного индекса, установлены закономерности физиологической адаптации лесных сообществ к изменениям оро-климатических факторов.
В решении современных задач рационального управления лесными ресурсами в глобальном масштабе на основе прогнозирования их климатогенных изменений все большее значение приобретают методы дистанционного зондирования лесов из космоса, открывающие новые возможности построения автоматизированных технологий их непрерывного долговременного мониторинга (Барталев и др., 2010; Исаев, 2012). Картографирование лесов по данным дистанционного зондирования Земли основано на автоматизированной системе фиксации сканирующим радиометром пространственных изменений весьма специфичных, не традиционных для геоботанической картографии отражательных характеристик лесного покрова, – главным образом жизненных форм растений, типов вегетативных органов (листьев и хвои), а также и их фенологической динамики. Это обеспечивает унифицированное картографирование растительного покрова больших территорий по указанным фитоценологическим признакам, что и было реализовано в рамках международного проекта Global Land Cover 2000 (Bartholome, Belward, 2005). На основе данных спутниковой системы SPOT-Vegetation были созданы карты наземных экосистем Северной Евразии (Bartalev et al., 2003), а по данным Terra-MODIS – карта растительного покрова России (Барталев и др., 2011).
В соответствии с возможностями дешифрирования космических снимков легенда к карте России включает несколько классов и 18 типологических единиц фитоценологических ареалов, которые авторами названы “типами растительности”. Для лесной зоны выделены шесть таких типов: 1) хвойные вечнозеленые; 2) лиственные; 3) смешанные с преобладанием хвойных, 4) смешанные; 5) смешанные с преобладанием лиственных: 6) хвойные листопадные.
Следует отметить, что эти дефиниции совершенно не соответствуют принятому в традиционной геоботанике содержательному смыслу понятий “классы растительных формаций” и “типы растительности” (Сукачев, 1975; Сочава, 1979; Вальтер. 1982), на основе которых создана известная классификация растительного покрова Европейской России и построены соответствующие мелкомасштабные карты (Исаченко, Лавренко, 1974; Грибова и др. 1980; Котова, 1987). В связи с этим для лесов Волжского бассейна дефиниции новой геоботанической систематики будем называть условно группами бореальных и неморальных лесов.
Данные группы характеризуют в неявном виде физиологические аспекты функциональной организации растительного (в том числе лесного) покрова, лежащие в основе его продуктивности, и в этом их несомненные преимущества по сравнению с выделением традиционных геоботанических единиц. Одним из таких преимуществ являются уже широко используемый в аэро-космической лесной экологии численный показатель – нормализованный разностный вегетационный индекс NDVI (Normalized Difference Vegetation Index).
В международной литературе чаще всего рассматриваются изменения NDVI растительных сообществ, связанные с климатическими колебаниями (Nath, 2014; Song et al., 2014; Aburas et al., 2015; Park et al., 2015; Zhang et al., 2015; и др.), однако причинные механизмы этих связей остаются слабо изученными. В настоящем сообщении изложен опыт изучения закономерностей дифференциации NDVI бореальных лесов крупного экорегиона в пространствах современного климата и равнинного рельефа. Установленная оро-климатическая ординация фотосинтетической активности лесного покрова может в определенной степени имитировать ее климатогенную динамику во времени (свойство эргодичности экологической системы).
ВЕГЕТАЦИОННЫЙ ИНДЕКС, ФОТОСИНТЕЗ И ПРОДУКТИВНОСТЬ
Вегетационный индекс вычисляется по следующей формуле:
где Nir и Red суть коэффициенты спектральной яркости в ближней инфракрасной (NIR) и красной (RED) областях спектра электромагнитного излучения. Типичная кривая отражения имеет максимум в области NIR из-за сильного отражения листьями и минимум в RED из-за поглощения хлорофиллом, поэтому NDVI отражает количество хлорофилла в зеленых листьях растений (Tucker, Sellers, 1986). Будем полагать, что этим параметром оценивается фотосинтетический потенциал растительного сообщества. Значения NDVI коррелируют со значениями хлорофиллового индекса, выражаемого в весовых единицах содержания хлорофилловых зерен.Основополагающую, глобальную роль хлорофилла не только в первичном биопродукционном процессе, но и в формировании всего живого вещества биосферы весьма ярко охарактеризовал еще в 1871 г. К.А. Тимирязев: " … зерно хлорофилла – тот орган, в котором неорганическое вещество, углекислота и вода превращаются в органическое, … тот фокус, та точка в мировом пространстве, в которой живая сила солнечного луча, превращаясь в химическое напряжение, слагается, накопляется для того, чтобы впоследствии исподволь освобождаться в тех разнообразных проявлениях движения, которые нам представляют организмы, как растительные, так и животные. Таким образом. зерно хлорофилла – исходная точка всякого органического движения, всего того, что мы разумеем под словом жизнь" (1947, с. 17).
Начальным звеном процесса автотрофного биогенеза является восстановление хлорофилловыми зернами зеленого листа/хвои поступающего из атмосферы СО2 при активном воздействии энергии солнечного света. Эта энергия расходуется на данный процесс, на разложение (фотолиз) поступающей из почвы воды, с выделение кислорода, а также на образование эмбрионального органического вещества общей формулы СН2О. При этом фотолиз воды – ключ ко всему процессу фотосинтеза (Гэлстон и др., 1983). Газо- и водообмен зеленого листа/хвои с атмосферой осуществляются через устьица в хлоропластах.
Вегетационный индекс рассматривается в самом общем виде как показатель обилия зеленой массы. Он конформирует с листовым индексом и концентрацией хлорофилла (Gobron et al., 2002; Myneni et al., 2002; Walker et al., 2003; Raynolds et al., 2012; Медведева и др., 2016). Для засушливых земель получена достаточно высокая корреляция NDVI с зеленой массой и ее продукцией (Золотокрылин и др., 2000). Другие эксперименты показали также, что и продуктивность лесов связана с NDVI (Елсаков, 2012). Однако анализ наших эмпирических данных по Большому Кавказу (Залиханов и др., 2010) не подтверждает такого вывода. Связь оказалась весьма слабой: коэффициент корреляции Спирмана rs = 0.29 и вероятность ошибки P < 0.01.
Дело в том, что параметр NDVI индицирует лишь начальное звено весьма сложного биопродукционного механизма: по содержанию зерен хлорофилла в зеленом листе оценивается скорость поглощения хлоропластами СО2 из атмосферы и интенсивность его превращения в углеводы, которые поставляют исходные вещества для дальнейших ассимиляционных процессов (Лархер, 1978). Соответственно хлорофилловый индекс может служить показателем стока углерода в растительные ценозы (Цельникер, 1994). Однако первые же экспериментальные исследования эколого-физиологических адаптаций растений к свету (Любименко, 1935) показали, что интенсивность фотосинтеза листа не связана прямой зависимостью с количеством хлорофилла в нем. По этой причине известное положение о том, что количество хлорофилла является показателем “производственной мощности” фотосинтетического аппарата (Горышина. 1979), остается достаточно проблематичным, и его мы принимает только как рабочую гипотезу.
Общая схема создания первичной продукции (после осуществления фотосинтеза), по (Риклефс, 1979), имеет вид (рис. 1). Внешние факторы влияют на эти процессы хотя и в том же направлении, но часто в иной степени, чем на СО2-газообмен, поэтому не всегда следует ожидать полного соответствия между газообменом и продуктивностью растений (Лархер, 1978). Экспериментально и эмпирически доказано, что растения в ценозах обладают примерно одинаковой способностью поглощать энергию фотосинтетической радиации (Ничипорович, 1973), однако величина утилизации этой энергии определяется обеспеченностью растений материально-энергетическими ресурсами среды, прежде всего ресурсами тепла и влаги, а также минерального питания (Абатуров, 1973), и эти ресурсы определяются внешними географическими факторами.
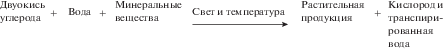
ЭКОЛОГИЧЕСКИЕ ПРОСТРАНСТВА РАСТИТЕЛЬНОСТИ, ПРОДУКТИВНОСТИ И ВЕГЕТАЦИОННОГО ИНДЕКСА
Под экологическим пространством объекта понимается совокупность экологических факторов, определяющих его структурную и/или функциональную организацию. Это пространство может быть как параметрическим, так и территориальным.
Полный спектр зональных типов растительных формаций основного водосбора Волжского бассейна (от среднетаежных до южностепных и северопустынных) соразмерен с наиболее крупными, субпланетарными единицами биосферы – географическими поясами и физико-географическими странами. На данном иерархическом уровне биосферы растительный покров и его продуктивность формируются в достаточно сложной системе вещественно-энергетических потоков в абиотической среде (рис. 2). Трансформация поступающих радиационного тепла и атмосферной влаги осуществляется последовательно четырьмя эшелонами промежуточных преобразователей (ретрансляторов). Наиболее высокие импульсы воздействия проходят по каналам связей от осадков теплого периода и теплоэнергетических показателей (средней июльской температуры, суммы биологически активных температур и испаряемости) к годовому и летнему коэффициентам увлажнения. Затем идет некоторое ослабление передачи сигналов к гидрологическим факторам (эвапотранспирации, поверхностному и подземному стоку), от которых воздействие передается к заключительному эшелону абиотических связей – летнему содержанию продуктивной влаги в почве. Этот сигнал совместно с годовым коэффициентом увлажнения формирует основные зональные типы растительных формаций и их региональные варианты.
Рис. 2.
Общая информационно-статистическая модель межкомпонентных связей в Волжском бассейне. Нормированные коэффициенты сопряженности межкомпонентных признаков, по (Пузаченко, Скулкин, 1981): 1 – 0.081–0.120; 2 – 0.121–0.160; 3 – 0.161–0.200; 4 – 0.201–0.240; 5 – 0.241–0.300; 6 – >0.300.
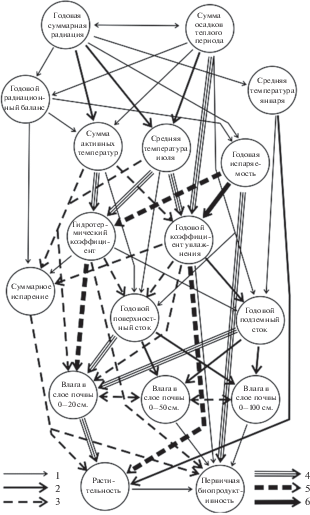
Первичная же биопродуктивность напрямую определяется главным образом тремя атмосферными гидротермическим параметрами – испаряемостью, гидротермическим коэффициентом Селянинова и суммарным годовым испарением. Гидрологический режим почвы в продукционном процессе играет второстепенную роль. Сравнительно слаба также зависимость продуктивности от типа растительных формаций. Таким образом, подтверждается известный тезис (Пузаченко, Скулкин, 1981) о том, что продуктивность леса является достаточно независимым (автономным) функциональным параметром по отношению к его фитоценологической структуре. Соответственно мало обнадеживающими становятся поиски прямых корреляций вегетационного индекса NDVI с зонально-региональной структурой лесного покрова.
В субконтинентальном масштабе (всего Волжского бассейна) связь нетто-продукции NPP c NDVI имеет вид определенного, хотя и сильно “размытого” нелинейного положительного тренда (рис. 3), который описывается кубической регрессией:
(2)
$\begin{gathered} {\text{NPP}} = - 44.00NDV{{I}^{3}} + 78.17NDV{{I}^{2}} - \\ - \,\,35.70NDVI + 11.89;\,\,\,\,{{r}_{s}} = 0.458;\,\,\,P < {{10}^{{--6}}}.~ \\ \end{gathered} $Рис. 3.
Распределение нетто-продуктивности (NPP) лесов Волжского бассейна естественного растительного покрова в пространстве значений вегетационного индекса на основном водосборе Волжского бассейна. В правом верхнем углу пунктиром очерчена область распространения лесных сообществ в зонах средней и южной тайги, а также северной и средней лесостепи. Как видно, здесь какая-либо значимая пространственная корреляция NDVI и NPP отсутствует.
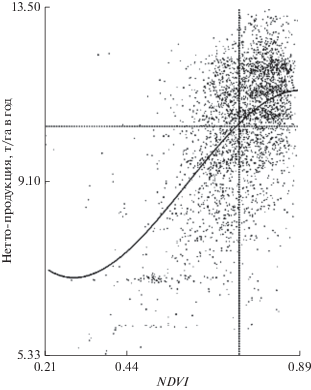
Если же мы ограничим пространственный анализ группой лесных природных зон, т.е. перейдем на более низкий (зонально-региональный) иерархический уровень, то получим овальный рой точек (рис. 3). Этот рой указывает на то, что фотосинтетический потенциал лесных формаций однозначно не реализуется в их конечной нетто-продукции, значения которой существенно определяются целым рядом гидролого-климатических и почвенно-геоморфологических факторов на постфотосинтезном этапе ассимиляционных процессов (рис. 1). Таким образом, региональный уровень природно-териториальной организации, отличающийся, как известно (Сочава, 1978), наибольшей сложностью географических пространств, демонстрирует эти свойства и в основном процессе функционирования экосистем – в создании первичного органического вещества.
ОБЩИЙ ЭКОЛОГО-ГЕОГРАФИЧЕСКИЙ АНАЛИЗ ФОТОСИНТЕТИЧЕСКОГО ПОТЕНЦИАЛА БОРЕАЛЬНЫХ ЛЕСОВ
Влияние экологических факторов на фотосинтез достаточно хорошо изучено в лабораторных условиях для отдельных (преимущественно травяных) растительных индивидов и для их популяций в искусственных фитотронах (Иванов, 1946; Лархер, 1978; Культиасов, 1982; Гэлстон и др., 1983; и др.). Проведены также многочисленные натурные исследования (в частности в горах Северной и Центральной Европы) гидротермического режима и фотосинтеза у хвойных пород деревьев в различные сезоны года (Вальтер, 1974).
Вместе с тем сам эколого-физиологический анализ (как лабораторный, так и натурный) остается в рамках аутэкологии, а экстраполяция его результатов на биогеосистемы разномасштабного географического пространства носит весьма гипотетический характер и по существу остается еще нерешенной задачей. В настоящем исследовании проведен опыт интерпретации первичной биопродуктивности лесных экосистем зонально-регионального уровня по вегетационному индексу на основе анализа его экологических связей с климатом и рельефом.
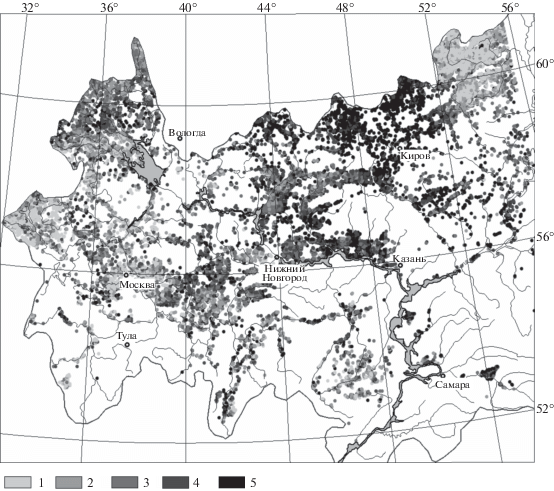
На карте распределения вегетационного индекса бореальных лесов Волжского бассейна (рис. 4) точечные данные представляют 86 000 участков площадью 4 км2, которые получены путем совмещения данных из цифровых матриц NDVI и растительного покрова, сформированных в Институте космических исследрований РАН (Барталев и др., 2004) с помощью разработанной П.А. Шарым программы “Аналитическая ГИС Эко”, версия 1.08r. В итоге для анализа мы имели наборы точек со значениями NDVI для трех групп бореальных лесов: 1 – темнохвойных (не менее 80% площади крон представлено елью), 2 – светлохвойных (не менее 80% занято сосной), 3 – смешанных, с преобладанием хвойных (60–80%). Температура и осадки, осредненные за 50 лет, взяты (в виде матриц с разрешением в 1 км) из базы данных WorldClim (Hijmans et al., 2005).
Наиболее высоким фотосинтетическим потенциалом (NDVI > 0.83) выделяются: 1) подтаежные леса обширной области Низменного Заволжья, охватывающей бассейны Унжи, Ветлуги и верховьев Вятки; 2) южнотаежные сообщества обрамляющей их гряды Северных Увалов; 3) смешанные леса заболоченной низменности к северо-западу от Рыбинского водохранилища; 4) хвойные леса заболоченной Мещерской низины по левобережью Оки; 5) более мелкие ареалы лесов по долине Цны и в верховьях Волги; 6) отдельный изолированный ареал Бузулукского бора в подзоне северных степей Самарского Заволжья. Во всем лесном поясе Русской равнины это почти исключительно хвойные сообщества низменных зандровых полесий (Коломыц, 2005), где преобладают малоплодородные песчаные и супесчаные, часто переувлажненные почвы.
Лесные же формации с минимальным вегетационным индексом (≤0.75) распространены довольно мелкими ареалами от южной тайги до северной лесостепи на хорошо дренируемых местоположениях Валдайской, Смоленско-Московской, Среднерусской и Приволжской возвышенностей, где господствуют местообитания с суглинистыми наиболее высокотрофными почвами. Обширный ареал крайне низких значений NDVI (до 0.20) расположен в среднетаежном Камско-Вишерском бассейне.
Таким образом, возвышенно-низменный макрорельеф Русской равнины предопределяет общую мозаичную картину распределения вегетационного индекса бореальных лесных формаций через посредство создаваемых им местообитаний, контрастных по мехсоставу и влажности почвообразующего субстрата. В наименее благоприятных местообитаниях переувлажненных зандровых низменностей и крупных речных долин хвойные и смешанные леса имеют наиболее высокую плотность хлорофилловых зерен и, следовательно, обладают максимально возможным фотосинтетическим потенциалом. При переходе же к хорошо дренируемым местообитаниями суглинистых возвышенностей, где водный и минеральный режимы почв наиболее благоприятны для растений, NDVI хвойных лесов заметно снижается. В этих геоморфологических и гидроэдафических условиях лесные сообщества в большей мере обеспечены вещественно-энергетическими ресурсами и могут достичь достаточно высокого уровня продуктивности при гораздо более низком фотосинтетическом потенциале. Как видим, в первом случае лесное сообщество стремится компенсировать дефицит вещественно-энергетических ресурсов увеличением содержания хлорофилла в хвое/листьях в целях поддержания по возможности более высокого продукционного процесса. Здесь отчетливо проявляется известный докучаевский закон компенсирующих экологических факторов, о котором речь пойдет ниже.
ОБЛАСТИ ДОМИНИРОВАНИЯ МАКСИМАЛЬНЫХ ЗНАЧЕНИЙ ВЕГЕТАЦИОННОГО ИНДЕКСА
Данным областям отвечают территории с определенными значениями сезонных температур и годового количества осадков. На основе данных по температуре и осадкам построены карты областей наибольшей фотосинтетической активности (NDVImax) для трех групп бореальных лесов Волжского бассейна (рис. 5). Рассмотрим диспозиции этих областей по холодно-слабогумидному климатическому тренду (юго-запад → северо-восток), с которым обнаружена наибольшая конформность пространственных изменений вегетационного индекса. Отметим, какие конкретные гидротермические изменения происходят на этом общеклиматическом тренде, используя материалы Атласа-монографии (Коломыц, 2005).
Обратимся сначала к ареалу NDVImax для темнохвойных лесов как наиболее ярких представителей бореального пояса (рис. 5а). Данный ареал занимает относительно небольшую область на самом северо-востоке бассейна – в Предуралье на Верхнекамской возвышенности, в верховьях Камы и на южных склонах Северных Увалов (рис. 4). Это по сути конечный пункт регионального холодно-слабогумидного вектора. В направлении “юго-запад → северо-восток”, т.е. от южных границ подтаежной зоны (примерно от восточной части Московской возвышенности и левобережья Окского бассейна) до Предуральских районов южной и средней тайги (в верховьях бассейнов Ветлуги и Камы) прослеживаются следующие изменения климатических параметров, которые назовем параметрическими трендами.
Рис. 4.
Распределение значений вегетационного индекса по ареалам всех трех групп бореальных лесов Волжского бассейна. Ареалы лесов даны по Геоботанической карте Европейской России (Грибова и др., 1980). Значения NDVI: 1 – 0.14–0.69; 2 – 0.69–0.75; 3 – 0.75–0.79; 4 – 0.79–0.83; 5 – 0.83–0.93.
Тренд 1. Ухудшается в целом световой режим местообитаний в связи со снижением поступающей радиационной энергии; годовая суммарная радиация уменьшается с 4200–4000 до 3200–3000 МДж/м2, а в солнечном спектре возрастает доля рассеянной радиации в связи с увеличением количества облачных погод. В этих условиях ослабление интенсивности света частично компенсируется повышением (до 50–60%) в фотосинтетически активной радиации доли физиологически более качественной рассеянной радиации (Культиасов, 1982).
Рис. 5.
Области максимальных значений (1) фотосинтетической активности групп бореальных лесов: темнохвойных (а), светлохвойных (б) и смешанных с преобладанием хвойных (в), – на территории Волжского бассейна. Темносерой отмывкой показан возвышенный рельеф бассейна.

Тренд 2. Годовой радиационный баланс как энергетическая база всех биогеоценотических процессов уменьшается с 1800–1600 до 1200–1000 МДж/м2; в соответствии с этим происходит снижение средних январских и июльских температур соответственно с –(11–12)°C до –(16.0–16.5)° и от 18.5–19.0° до 17.0–16.5°. При этом континентальность климата меняется незначительно и неоднозначно по сезонам года: коэффициент январской широтной континентальности Полозовой возрастает с 74 до 79%, а коэффициент июльской континентальности – наоборот, уменьшается с 42 до 37%. В этих условиях происходит смена ели европейской (Picea excels) на ель сибирскую (Picea abovata), к которой примешиваются кедр и лиственница (Толмачев, 1974).
Тренд 3. Сумма биологически активных температур падает с 2200–2000 до 1800–1600°, т.е. до нижнего теплоэнергетического уровня для ели и пихты (Пузаченко, Скулкин, 1981); при этом сокращается продолжительность вегетационного периода – с 180–175 до 150–145 дней, а коэффициент снежности зимы Осокина, включающий длительность залегания снежного покрова, увеличивается в 2–2.5 раза.
Тренд 4. Годовое количество осадков увеличивается с 650–700 до 800–850 мм, что и позволяет считать данный тренд слабогумидным.
Тренд 5. Годовой коэффициент увлажнения Высоцкого-Иванова возрастает с 1.1–1.2 до 1.7–1.8, а гидротермический коэффициент Селянинова – с 1.8–2.0 до 3.0–3.2 единиц и более, что свидетельствует о значительном росте атмосферного увлажнения территории до уровня его существенного переувлажнения, свойственного уже северотаежным ландшафтам.
Тренд 6. В направлении от подтаежных пихто-ельников Верхневолжья к кедрово-лиственнично-еловым лесам средней тайги Прикамья таксономическая норма годового суммарного испарения, которое при сомкнутом лесном покрове представлено почти исключительно транспирацией (Раунер, 1972), сокращается на 15% – с 578 до 502 мм. Одновременно снижается норма летних запасов продуктивной влаги в метровой толще почвы – с 167 до 134 мм. Очевидно, сказывается рост по данному направлению континентальности климата. При этом в районе Поветлужья происходит переход летних влагозапасов в почве через критический уровень в 150 мм, когда суммарное испарение практически равно испаряемости (Зубенок, 1976). Таким образом, если в начале рассматриваемого климатического тренда транспирация древостоев в вегетационный период регулируется главным образом температурой, что подтверждается специальными опытами (Иванов, 1946), то в его конце (в районе NDVImax) она определяется также влагобеспеченностью, по правилу лимитирующих экологических факторов Либиха (Одум, 1975). Это особенно должно сказаться на произрастании ели, которая весьма требовательна к влаге.
Итак, второй и третий, а также в меньшей степени шестой параметрические тренды являются решающими в ухудшении общих климатических условий произрастания темнохвойных пород, что должно негативно отразиться и на их продуктивности. Прежде всего, понижение температуры уменьшает скорость и величину открывания устьиц в хлоропластах, а также сокращает поглощение и восстановление углерода, что изначально снижает верхнюю границу нетто-фотосинтеза (Лархер, 1978). Далее, с понижением температуры в вегетационный период уменьшается проницаемость цитоплазмы; тем самым сокращается приток транспирируемой влаги и соответственно поступление минеральных питательных веществ из почвы к ассимилирующим органам, что ослабляет рост растений (по экспоненциальному закону Вант-Гоффа). Задерживаются также поглощение азота корнями, передвижение его по растению и переведение в органические соединения (Культиасов, 1982). В холодное время года удлиняется период нахождения хвои в замерзшем состоянии, что задерживает наступление весеннего срока восстановления нормального фотосинтеза и в целом снижает его летний максимум (Вальтер, 1974).
Четвертый и пятый тренды (совместно со вторым) обусловливают смену (прежде всего на плакорных местообитаниях) зональных типов темнохвойных лесов. В данном случае происходит смена подтаежных и южнотаежных неморальнотравяных и кислично-разнотравных пихто-ельников на северотаежные ельники (с примесью кедра и лиственницы) чернично-сфагново-долгомошные и кустарничково-зеленомошные (Грибова и др., 1980). Согласно нашим эмпирическим данным (Коломыц, 2008), такая фитоценологическая перестройка происходит в условиях повышения в конце весны – начале лета уровня грунтовых вод в среднем с 90–120 до 30–70 см. В последнем случае неизбежен регулярный подъем зеркала грунтовых вод до поверхности почвы (Вальтер, 1974). Избыточное переувлажнение ризосферы вызывает, во-первых, развитие в анаэробных условиях токсичных глеевых почвенных процессов, а во-вторых, – снижение осмотического давления в тканях растений (их сосущей силы) ниже допустимого предела, что приводит к сокращению минерального питания. В результате деревья угнетаются и соответственно снижается их фотосинтетический потенциал. Так, в черничном ельнике средней тайги содержание хлорофиллла в хвое оказывается выше, чем в заболоченных ельниках северной тайги (Тужилкина, Бобкова, 2010). Наконец, как отмечал еще К.А. Тимирязев (1947), высокая влажность воздуха замедляет сам процесс фотосинтеза, поскольку водяные пары поглощают тепловые лучи.
На фоне ухудшения всех этих лесорастительных условий по направлению “юго-запад → северо-восток” средняя первичная биопродуктивность региональных ландшафтов в доминирующих значениях своих экологических ниш уменьшается сравнительно мало – с 12.0–11.5 до 10.5–10.0 т/га год (табл. 1). При этом объем экологической ниши остается сравнительно узким (охватывает три–четыре продукционной градации), а экологический доминант продуктивности почти сохраняет свою повышенную вероятность (65% → 61%): продукционный процесс по всему вектору остается достаточно чувствительным к изменения климата.
Таблица 1.
Распределение групп бореальных лесных формаций Волжского бассейна в пространстве первичной биологической продуктивности. Нормированные частные коэффициенты связей, по (Пузаченко, Скулкин, 1981)
| Группы лесных формаций | Первичная биопродуктивность, т/г год | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 7.0–7.6 | 7.6–8.3 | 8.3–9.0 | 9.0–9.5 | 9.5–10.0 | 10.0–10.5 | 10.5–11.0 | 11.0–11.5 | 11.5–12.0 | 12.0–12.5 | |
 1 1
|
0.02 | 0.01 | 0.06 | 0.13 | 0.06 | 0.06 | 0.66 | |||
 2 2
|
0.08 | 0.11 | 0.29 | 0.36 | 0.16 | |||||
 3 3
|
0.02 | 0.11 | 0.23 | 0.30 | 0.31 | 0.03 | ||||
 4 4
|
0.02 | 0.16 | 0.17 | 0.28 | 0.37 | |||||
 5 5
|
0.16 | 0.04 | 0.16 | 0.38 | 0.14 | 0.10 | 0.02 | |||
 6 6
|
0.36 | 0.26 | 0.30 | 0.06 | 0.01 | 0.01 | ||||
 7 7
|
0.06 | 0.08 | 0.15 | 0.16 | 0.21 | 0.21 | 0.03 | |||
 8 8
|
0.07 | 0.09 | 0.18 | 0.11 | 0.10 | 0.16 | 0.15 | 0.14 | ||
 9 9
|
0.15 | 0.32 | 0.33 | 0.14 | 0.06 | |||||
Примечание. Зонально-провинциальные группы растительных формаций, по (Грибова и др., 1980): 1–6 – темнохвойные и широколиственно-темнохвойные леса: 1, 2, 3 – восточноевропейские (среднетаежные, южнотаежные и подтаежные), 4, 5, 6 – камско-печерские (средне- и южнотаежные, подтаежные пихтово-еловые сложные и смешанные); 7–9 – сосновые и широколиственно-сосновые леса: 7 – средне- и южнотаежные, 8, 9 – подтаежные сосняки с дубом в подлеске и смешанные. Жирным шрифтом выделены доминирующие области экологических ниш групп лесных формаций.
Одновременно происходит общий значительный рост вегетационного индекса темнохвойных лесов, который достигает максимума в указанном Верхнекамско-Предуральском регионе (см. рис. 5а) с экстремально негативными для европейской средней тайги климатическими параметрами. В более западном бассейне верховьев Северной Двины концентрация хлорофилла в хвое ельников продолжает возрастать и по мере перехода от средней тайги к северной (Тужилкина, Бобкова, 2010).
Правомерно полагать, что фотосинтетический аппарат темнохвойных лесов стремится поддержать “запрограммированность”, по терминологии В.Б. Сочавы (1978), стратегии растительного покрова к максимальной продуктивности. Эта гипотеза подтверждается описанной выше региональной картиной распределения интегрального индекса NDVI всех трех групп бореальных лесов по возвышенно-низменному макрорельефу Волжского бассейна. Основываясь на результаты исследований, представленных в работах (Любименко, 1935; Харитонович, 1968; Лархер, 1978; Горышина, 1979; Культиасов, 1982; Гэлстон и др., 1983), можно обрисовать следующие механизмы такого распределения.
Теневые листья (хвоя) деревьев, находящиеся в условиях более низких температур и повышенной влажности воздуха, имеют обычно на единицу поверхности минимальное число относительно крупных устьиц, через которые осуществляется газообмен растений с атмосферой. В то же время они содержат больше хлорофилла, чем листья световые, где более высокая температура и пониженная влажность воздуха обеспечивают относительно высокую скорость фотосинтеза, чему способствует также повышенная удельная плотность более мелких устьиц. Повышенная концентрация хлорофилла у тенелюбов способствует более энергичному фотосинтезу при уменьшении освещенности – ослаблении потока солнечной радиации. Таким образом, происходит определенное выравнивание суммарной фотосинтетической активности листьев (хвои), находящихся в неодинаковых гидротермических условиях – в тени и на свету. Соответственно происходит выравнивание и конечной нетто-продукции, чему способствует также более выгодный баланс между фотосинтезом и дыханием у теневыносливых растений.
Реализация стратегии максимальной продуктивности по региональному холодно-слабогумидному климатическому вектору в данном случае осуществляется за счет увеличения удельной плотности хлорофилла в хвое ельников, т.е. роста их фотосинтетической активности, которое компенсирует в той или иной мере отмеченное выше ухудшение климатических условий для роста растений. Налицо одно из проявлений известного докучаевского закона компенсирующих экологических факторов (Докучаев, 1953). Согласно известной классификации экологических факторов (Реймерс, 1990), плотность хлорофилловых зерен в листве/хвое, индицируемая параметром NDVI, можно отнести по происхождению к биотическим факторам, а по среде возникновения – к фактору физиологическому. Докучаевский закон как нельзя лучше применим именно к темнохвойным лесам, которые относятся, как известно (Мазинг, 1970), к наиболее высокоорганизованным и наиболее развитым в эволюционном отношении лесным сообществам, что и проявляется в их способности сохранить при негативном воздействии внешних экологических факторов максимально возможный высокий уровень продукционного процесса путем увеличения своего фотосинтетического потенциала.
Перейдем к рассмотрению вегетационного индекса у групп светлохвойных и смешанных лесов. Ареал максимальных значений параметра NDVI светлохвойных (главным образом сосновых) лесов охватывает значительную площадь центральной части Волжского бассейна, подходя к его границам на северо-западе (рис. 5б). Сосна обыкновенная (Pinus silvestris) является светолюбивой породой и обладает, как известно, широкой экологической амплитудой. Она произрастает в интервале температур от –50 до +40° и при годовой сумме осадков от 300 до 700 мм. С одной стороны, она засухоустойчива и индифферентна к испаряемости, а с другой – способна занимать заболоченные местоположения (Харитонович, 1968). В результате ее NDVImax охватывает весьма широкий ареал: от южных районов неморальнолесной подзоны (в верховьях Суры на Приволжской возвышенности) на юге до северных границ южнотаежной зоны (верховьев Сухоны, Ветлуги и Камы) на севере и от верховьев Волги и северо-западных истоков Рыбинского водохранилища на западе до средней части левобережного бассейна Камы в Предуралье на востоке.
Смешанные (с преобладанием хвойных пород) леса имеют относительно протяженную по вектору “северо-запад → юго-восток” область значения NDVImax в северо-восточной части бассейна (рис. 5в). Эта область сближается с ареалом максимума для темнохвойных пород, поэтому на нее можно распространить те же закономерности описанных региональных экологических связей. Вместе с тем, примесь к медленно растущим ели и пихты быстрорастущих сосны, лиственницы и мелколиственных пород означает, что фотосинтетический потенциал этой группы хвойных лесов в целом существенно возрастает. Соответственно их чувствительность к изменениям климата снижается и наступление NDVImax по северо-восточному климатическому вектору здесь наступает несколько раньше – при меньшем ухудшении лесорастительных условий, чем это свойственно группе чистых пихто-ельников.
ЗАКЛЮЧЕНИЕ
По материалам спутникового зондирования проведены региональное картографирование и эколого-географический анализ вегетационного индекса NDVI бореальных лесов Волжского бассейна, что позволило обрисовать причинные механизмы формирования фотосинтетического потенциала лесов в различных условиях равнинного рельефа и климата. Описаны пространственные климатические тренды в данном регионе по направлению “юго-запад → северо-восток”, с которым обнаружена наибольшая конформность пространственных изменений вегетационного индекса. Выполнен сопряженный анализ векторных гидротермических полей, а также полей продуктивности и параметра NDVI группы темнохвойных лесов как основных зональных представителей бореальной биоклиматической системы. Установлено достаточно четкое проявление докучаевского закона взаимозаменяемости экологических факторов в формировании фотосинтетического потенциала бореальных лесов.
Список литературы
Абатуров Б.Д. Биопродукционный процесс в наземных экосистемах. М.: Наука, 1973. 128 с.
Барталев С.А., Ершов Д.В., Исаев А.С., Лупян Е.А., Уваров И.А. Карта растительного покрова России. М.: Институт космических исследований РАН и Центр экологии и продуктивности лесов РАН, 2010. Масштаб 1 : 5 000 000. 2 л.
Барталев С.А., Егоров В.А., Ильин В.О., Лупян Е.А. Синтез улучшенных сезонных изображений Северной Евразии для картографирования и мониторинга динамики растительности по данным SPOT-VEGETATION // Дистанционное зондирование поверхности Земли и атмосферы. 2004. Вып. 5. С. 12–14.
Барталев С.А., Егоров В.А., Ершов Д.В., Исаев А.С., Лупян е.А., Плотников Д.Е., Уваров И.А. Спутниковое картографирование растительного покрова России по данным спектрорадиометра MODIS // Современные проблемы дистанционного зондирования Земли из космоса. 2011. Т. 8. № 4. С. 285–302.
Вальтер Г. Растительность Земного Шара. Эколого-физиологическая характеристика. М.: Прогресс, 1974. Т. II. 423 с.
Вальтер Г. Общая геоботаника. Пер. с немецкого. М.: Мир, 1982. 261 с.
Горышина Т.К. Экология растений. М.: Высшая школа, 1979. 368 с.
Грибова С.А., Исаченко Т.И., Лавренко Е.М., ред. Растительность Европейской части СССР. Л.: Наука, 1980. 429 с.
Гэлстон А., Девис П., Сэттер Р. Жизнь зеленого растения / Пер. с англ. М.: Мир, 1983. 549 с.
Докучаев В.В. Наши степи прежде и теперь. М.: Сельхозгиз, 1953. 152 с.
Елсаков В.В. Спутниковая съемка в оценке продуктивности экосистем Европейского Севера / Современные проблемы дистанционного зондирования Земли из космоса. 2012. Т.9. №1. С. 71–79.
Залиханов М.Ч., Коломыц Э.Г., Шарая Л.С., Цепкова Н.Л., Сурова Н.А. Высокогорная геоэкология в моделях. М.: Наука, 2010. 487 с.
Золотокрылин А.Н., Коняев К.В., Титкова Т.Б. Зависимость между аномалиями индекса вегетации и аномалиями месячных сумм осадков в зоне умеренного и недостаточного увлажнения // Исследования Земли из космоса. 2000. № 6. С. 74–78.
Зубенок Л.И. Испарение на континентах. Л.: Гидрометеоиздат, 1976. 264 с.
Иванов Л.А. Свет и влага в жизни древесных пород. М.; Л.: Изд-во АН СССР, 1946. 60 с.
Исаев А.С., ред. Разнообразие и динамика лесных экосистем России. Книги 1. М.: Товарищество научных изданий КМК, 2012. 461 с.
Исаченко Т.И., Лавренко Е.М., ред. Карта растительности Европейской части СССР. М-б 1 : 2 500 000. М.: ГУГК, 1974. 6 л.
Коломыц Э.Г. Бореальный экотон и географическая зональность: Атлас-монография. М.: Наука, 2005. 390 с.
Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
Котова Т.В., ред. Растительность Европейской части СССР и Кавказа. М-б 1 : 2 000 000. М.: ГУГК, 1987. 4 л.
Культиасов И. М. Экология растений. М.: Изд-во МГУ, 1982. 380 с.
Лархер В. Экология растений. Пер. с немецк. М.: Прогресс, 1978. 185 с.
Любименко В.Н. Фотосинтез и хемосинтез в растительном мире. М.–Л.: Сельхозгиз, 1935. 320 с.
Мазинг В.В. К вопросу эволюции биоценотических систем // Теоретические проблемы фитоценологии и биогеоценологии. М.: Наука, 1970. С. 95–107.
Медведева М.А., Елсаков В.В., Савин И.Ю., Барталёв С.А. О связи фенологического развития растительности таежной зоны с величиной NDVI, определенной по спутниковым данным // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 7. № 1. С. 319–329.
Ничипорович А.А. Основы фотосинтетической продуктивности растений // Современные проблемы фотосинтеза. М.: Изд-во МГУ, 1973. С. 17–43.
Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. 740 с.
Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: Системный анализ. М.: Наука, 1981. 275 с.
Раунер Ю.Л. Тепловой баланс растительного покрова. М.: Наука, 1972. 210 с.
Реймерс Н.Ф. Природопользование. Словарь–справочник. М.: Мысль, 1990. 639 с.
Риклефс Р.Э. Основы общей экологии. Пер. с англ. М.: Мир, 1979. 424 с.
Сочава В.Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 189 с.
Сукачев В.Н. Избранные труды. Том III. Проблемы фитоценологии. Л.: Наука, 1975. 543 с.
Толмачев А.И. Введение в географию растений. Л.: Изд-во ЛГУ, 1974. 244 с.
Тимирязев К.А. Избранные работы по хлорофиллу и усвоению света растениями. М.: Изд-во АН СССР, 1947. 351 с.
Тужилкина В.В., Бобкова К.С. Хлорофильный индекс в фитоценозах коренных ельников Европейского Северо-Востока // Лесной журнал. 2010. № 2. С. 17–23.
Харитонович Ф.И. Биология и экология древесных пород. М.: Лесная промышленность, 1968. 304 с.
Цельникер Ю.Л. Хлорофилловый индекс как показатель годичной аккумуляции углерода древостоями леса // Физиология растений. 1994. Т. 41. С. 325–330.
Aburas M.M., Abdullah S.H., Ramlia M.F., Ashaaria Z.H. Measuring land cover change in Seremban, Malaysia using NDVI Index // Procedia Environmental Sciences. 2015. V. 30. P. 238–243.
Bartalev S.A., Belward A.S., Ershov D.V., Isaev A.S. A new SPOT4-VEGETATION derived Land Cover Map of Nortehrn Eurasia // International. Journal of Remote Sensing. 2003. V. 24. № 9. P. 1977–1982.
Bartholome E., Belward A.S. GLC2000: a new approach to global land cover mapping from Earth observation data // Intetnational Journal Remote Sensing. 2005. V. 26. P. 1959–1977.
Gobron, N., Pinty, B., Verstraete, M. M., Widlowski J.-L., D.J. Diner D.J. Uniqueness of multiangular measurements. II. Joint retrieval of vegetation structure and photosynthetic activity from MISR // IEEE Transactions on Geoscience and Remote Sensing. 2002. V. 40. № 7. P. 1574–1592.
Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., Jarvis A. Very high resolution interpolated climate surfaces for global land areas // International J. Climatology. 2005. V. 25. № 15. P. 1965–1978.
Myneni R.B., Hoffman S., Knyazikhin Y., Privette J.L., Glassy J., Tian Y., Wang Y., Song X., Zhang Y., Smith G.R., Lotsch A., Friedl M., Morisette J.T., Votava P., Nemani R.R., Running S.W. Global products of vegetation leaf area and fraction absorbed PAR from year one of MODIS data // Remote Sensing of Environment. 2002. V. 83. P. 214–231.
Nath B. Quantitative Assessment of Forest Cover Change of a Part of Bandarban Hill Tracts Using NDVI Techniques // Journal of Geosciences & Geomatics. 2014. V. 2. № 1. P. 21–27.
Park H., Jeong S., Ho C., Kim J.,Brown M.E, Schaepman M.E. Nonlinear response of vegetation green-up to local temperature variations in temperate and boreal forests in the Northern Hemisphere // Remote Sensing of Environment. 2015. V. 165. P. 100–108.
Raynolds M.K., Walker D.A., Epstein H.E., Pinzon J.E., Tucker C.J. A new estimate of tundra-biome phytomass from trans-Arctic field data and AVHRR NDVI // Remote Sensing Letters. 2012. V. 3. Is. 5. P. 403–411.
Song X.P., Huang C., Sexton J.O., Channan S., Townshend J.R. Annual Detection of Forest Cover Loss Using Time Series. Satellite Measurements of Percent Tree Cover // Remote Sensing. 2014. № 6. P. 8878–8903.
Tucker C.J., Sellers P.J. Satellite remote sensing of primary production // International J. Remote Sensing. 1986. V. 7. № 11. P. 1395–1416.
Walker D.A., Epstein H.E., Jia G.J., Balser A., Copass C., Edwards E. J., Gould W. A., Hollingsworth J., Knudson J., Maier H. A., Moody A., Raynolds M. K. Phytomass, LAI, and NDVI in northern Alaska: relationships to summer warmth, soil pH, plant functional types, and extrapolation to the circumpolar Arctic // J. Geophysical. 2003. V. 108. P. 1–7.
Zhang, Z., Xue Y., MacDonald G., Cox P.M., Collatz G.J. Collatz Investigation of North American vegetation variability under recent climate: A study using the SSiB4/TRIFFID biophysical/dynamic vegetation model // J. Geophysical Research.: Atmospheres. 2015. V. 120. P. 1300–1321.
Дополнительные материалы отсутствуют.