

Лесоведение, 2020, № 5, стр. 399-411
Моделирование лесного покрова бассейна реки Венгери на острове Сахалин с применением данных дистанционного зондирования
К. А. Корзников a, *, Н. Г. Беляева b, Р. Б. Сандлерский c
a Ботанический сад-институт ДВО РАН
690024 Владивосток, ул. Маковского, 142, Россия
b Институт географии РАН
119017 Москва, Старомонетный пер., 29, стр. 4, Россия
c Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский проспект, 33, Россия
* E-mail: korzkir@mail.ru
Поступила в редакцию 02.04.2018
После доработки 16.11.2018
Принята к публикации 06.06.2020
Аннотация
Выполнено моделирование актуального растительного покрова бассейна р. Венгери (восточное побережье центральной части Сахалина) площадью свыше 33 тыс. га. Картографическая модель создана при помощи алгоритма пошагового дискриминантного анализа на основе оригинальных геоботанических данных, цифровой модели рельефа и спектральных характеристик снимков Landsat 5. В растительном покрове выделено 9 физиономических типов растительных сообществ. Из них 3 – нелесные (скальная растительность, горные тундры, луга), 6 – различные типы лесов: пихтово-еловые (Abies sachalinensis, Picea ajanensis), лиственничники, лиственничные редины (Larix cajanderi), стланиковые сообщества (Pinus pumila, Dushekia fruticosa), каменноберезовые леса (Betula ermanii), мелколиственные и пойменные лиственные леса (Alnus hirsuta, Betula platyphylla, Chosenia arbutifolia, Populus suaveolens, Salix udensis и др.). Указана принадлежность физиономических типов лесов к синтаксонам эколого-фитоценотической и эколого-флористической классификационных систем. Лесные сообщества развиты на 89.1% территории водосбора. Наибольшую площадь занимают лиственничные леса и редколесья – 33.32% речного бассейна. Ландшафтно-климатические условия района способны поддерживать существование зональных пихтово-еловых лесов, но на их долю приходится только 11.23% территории. Широкое распространение лиственничников обусловлено естественной пирогенной трансформацией темнохвойных лесов. В условиях сильно расчлененного горного рельефа участки растительного покрова затененных склонов северных экспозиций не поддаются корректному автоматическому дешифрированию и моделированию. Восстанавливать растительный покров предлагается по данным визуального дешифрирования снимков высокого разрешения и материалам полевых работ.
Особенности применения методов дистанционного зондирования для установления пространственной структуры лесного покрова и решения спектра смежных задач изложены в большом числе российских и зарубежных публикаций (Reese et al., 2002; Bartalev et al., 2003; Hansen et al., 2005; Исаев и др., 2009; McRoberts et al., 2010; Chernen’kova et al., 2013, 2015; Пузаченко и др., 2014; Комарова и др., 2016; Пузаченко, Черненькова, 2016; Gomes et al., 2016; Zhu et al., 2016; Belyaeva, Chernen`kova, 2018; и др.). Использование данных дистанционного зондирования расширяет возможности для отображения пространственной неоднородности растительности (Черненькова, Морозова, 2017). Наличие в свободном доступе мультиспектральных космических снимков и цифровых моделей рельефа упрощает создание моделей растительного покрова, что особо актуально для инвентаризации растительности и растительных ресурсов труднодоступных малоизученных районов.
Хотя в мире и европейской части России методы дистанционного зондирования давно пополнили арсенал подходов к изучению растительного покрова, на территории российского Дальнего Востока широкого распространения они до сих пор не нашли. Из проведенных в регионе работ следует упомянуть моделирование растительного покрова с использованием морфометрических параметров рельефа для Ланжинских гор в Северной Охотии (Омелько и др., 2010; Omelko et al., 2012), а также для верхнего течения р. Соколовки, Южный Сихотэ-Алинь (Yakovleva, 2010). В горных регионах рельеф определяют локальные экологические условия, от него зависят микроклимат, почвы, режим локальных нарушений (например, интенсивность схода селей или лавин) и, в конечном счете, растительный покров. Использование морфометрических характеристик рельефа позволяет провести моделирование потенциальной, но не актуальной растительности. Поскольку размещение растительных сообществ определяется не только ландшафтными условиями, но и историей нарушений, распространение производных сообществ невозможно смоделировать, оперируя только геоморфологическими характеристиками местности.
Цель нашей работы заключалась в создании модели актуального растительного покрова, с фокусом на лесную растительность малоизученного района Восточно-Сахалинских гор на о. Сахалин – бассейна р. Венгери. Для достижения цели в ходе полевых исследований мы определили пространственное размещение различных типов растительных сообществ (“обучающая выборка”), а затем связали их со спектральными характеристиками космических снимков и морфометрическими переменными рельефа. Проведенное нами моделирование является первой работой подобного рода для территории Сахалина и позволяет оценить закономерности размещения растительных комплексов в труднодоступной горной части острова.
ОБЪЕКТЫ И МЕТОДИКА
Характеристика района исследования
Бассейн р. Венгери приурочен к центральной части Сахалина, восточному побережью острова (рис. 1). Река берет начало у вершин Набильского хребта Восточно-Сахалинских гор и впадает в Охотское море в 30 км к востоку от места истока. Площадь бассейна составляет свыше 33 тыс. га. Абсолютная высота над уровнем моря меняется от 0 до 1431 м, средняя высота составляет 442 м. Набильскому хребту соответствует структурно-денудационный, крутосклонный сильно расчлененный рельеф (глубина расчленения 700–1150 м). Восточная часть бассейна располагается в пределах Центрального хребта – сильно расчлененного (глубина расчленения 320–720 м над уровнем моря) денудационного низкогорья с крутой и средней крутизны склонами. Водораздельным участкам обоих хребтов местами соответствуют пологие поверхности выравнивания. Почвы горные лесные, кислые подзолистые, иллювиально-гумусные. В центральной части бассейна, между двумя хребтами, сформировалось денудационное грядовое предгорье с отметками высот 100–260 м, где выражены эрозионно-аккумулятивные формы рельефа – слабонаклонные горные шлейфы. Почвы бурые лесные и кислые подзолистые. Вдоль берега Охотского моря развиты морские а-бразионно-аккумулятивные слаборасчлененные террасы с активными клифами. Долина р. Венгери в верхнем течении узкая, v-образная, аллювиальные террасы отсутствуют. Полугорный характер русла р. Венгери сохраняется до устья. Участки развитой поймы выражены в среднем и нижнем течении.
По данным ближайшей к территории исследования метеостанции Пограничное (30 км к югу, побережье Охотского моря), среднегодовая температура воздуха составляет минус 1.6°C, среднегодовое количество осадков 772.4 мм. Климат муссонный. Вследствие длительного таяния морских льдов начало лета холодное. Охлаждающий эффект также создает холодное Восточно-Сахалинское течение. В конце лета и осенью часты тропические циклоны с обильными осадками и сильными порывами ветра. В горах термический режим более суровый, осадков выпадает больше, чем на побережье (Казаков, Генсиоровский, 2007). В то же время, защищенным от холодного морского воздуха речным долинам соответствует более мягкий по сравнению с побережьем микроклимат. Особенность погодного режима территории заключается в относительно частых грозах, возникающих вследствие столкновения поступающего с запада, прогретого над Тымь-Поронайской равниной, теплого воздуха с холодными воздушными массами Охотского моря.
В зональном отношении территория исследования соответствует северному пределу подзоны южной тайги. Зональные сообщества представлены темнохвойными лесами из Abies sachalinensis и Picea ajanensis, с преобладанием в древостое последнего вида. Участие растений умеренной флоры выражено слабее, чем в более южных районах острова. В растительном покрове очень редки широколиственные виды, такие как Fraxinus mandshurica, Quercus mongolica, Ulmus japonica. Полностью отсутствуют представители таких семейств как Araliaceae, Hydrangaceae, Rutaceae, нет деревянистых лиан Actinidia kolomikta и Schisandra chinensis. В вертикальной зональности растительности выражены три пояса: горнотундровый (с доминированием видов из семейства Ericaceae), редколесный (с доминированием Betula ermanii) и таежный (A. sachalinensis и P. ajanensis) (Зоны …, 1999).
Лесная растительность Сахалина сильно нарушена вследствие экстенсивного лесопользования и масштабных антропогенных пожаров (Харук и др., 2004). Бассейн р. Венгери был выбран в качестве территории исследования из-за отсутствия населенных пунктов, объектов транспортной инфраструктуры и хозяйственного освоения лесов. Весь бассейн реки входит в состав государственного природного заказника регионального значения “Восточный”. Все это позволяет говорить о минимальном антропогенном преобразовании состава и структуры растительных сообществ.
Сбор и анализ полевых данных
Полевые исследования проведены в 2015 и 2017 г. Полные геоботанические описания выполнены на пробных площадях различного размера, в зависимости от типа сообществ – от 25 до 400 м2. Географические координаты мест выполнения описаний фиксировали при помощи GPS-ГЛОНАСС приемника Garmin eTrex 30 с заявленной точностью позиционирования ±7 м. В ходе радиальных маршрутов, помимо полных геоботанических описаний, выполняли также краткие. В общей сложности сделано 60 полных и 20 кратких геоботанических описаний.
Описанные сообщества были отнесены к различным синтаксонам растительности в рамках эколого-фитоценотического (“доминантного”) и эколого-флористического (“метод Браун-Бланке”) подходов. Для создания картографической модели выделено 9 физиономических типов растительности, из которых 6 относятся к лесам, включая заросли стлаников Pinus pumila и Duschekia fruticosa (Тихомиров, 1946). Названия синтаксонов эколого-фитоценотической классификации приведены по работе В.Ю. Нешатаевой (2009), эколого-флористической классификации – по ряду источников (Vegetation …, 1988; Krestov, Nakamura, 2002; Krestov et al., 2009). Латинские названия сосудистых растений указаны согласно сводке С.К. Черепанова (1995), мхов – в соответствии с перечнем О.Ю. Писаренко с соавт. (2012), печеночных мхов – согласно перечню В.А. Бакалина (2012).
Моделирование
Создание картографической модели растительного покрова основано на интеграции результатов полевых исследований, данных дистанционного зондирования и цифровой модели рельефа. Использованы разносезонные безоблачные снимки Landsat 5 TM (24.05.2009 и 28.06.2010) и Landsat 8 OLI TIRS (30.09.2015), а также цифровая модель рельефа (SRTM, абсолютные отметки высот). На основе исходной модели высот в программе ArcGI-S 10 были вычислены значения крутизны и экспозиции склонов с пространственным разрешением данных 30 × 30 м. Обучающая выборка составлена из пикселей, соответствующих местам выполнения геоботанических описаний. Процедура классификации проведена методом линейного дискриминантного анализа. Информативный комплекс переменных определялся автоматически, методом пошагового отбора, алгоритмом с последовательным включением переменных (“forward stepwise”), при значениях F-включения, равного 3.84 и F-исключения, равного 2.71. Априорные вероятности отнесения к классу были заданы соответственно размерам выборки обучающего класса. Предсказанные принадлежности пикселей к различным физиономическим типам растительности визуализированы и переведены в векторный формат. На заключительном этапе была проведена генерализация картографической модели до минимальной площади выдела, равной 0.5 га.
В условиях сильно расчлененного рельефа склоны северных экспозиций затенены. Низкая отражающая способность растительных сообществ затененных склонов приводит к некорректным результатам их программного дешифрирования и моделирования. Пиксели затененных склонов северных экспозиций в 100% случаев распознавались как принадлежащие физиономическому классу темнохвойных лесов. В научной литературе проблема подобного рода подробно описана (Giles, 2001; Riano et al., 2003; Richter et al., 2009; Shahtahmassebi et al., 2013). Предложено несколько вариантов ее решения, например, путем внесения поправок в спектральные значения пикселей в зависимости от крутизны и экспозиции склонов. В нашем случае предложенные приемы устранения эффекта глубоких теней оказались недействующими. Поэтому восстановление растительного покрова затененных участков выполнено экспертным путем, с привлечением данных полевых наблюдений и визуального дешифрирования космических снимков высокого разрешения (использованы данные Digital Globe).
Работа с векторными и растровыми данными велась в программных продуктах ENVI 4.7, ERDAS Imagine 2014, а также ArcGIS 10. Дискриминантный анализ проведен в программе IBM SPSS Statistics 20. Описательная статистика и визуализация выполнены в программной среде R.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дешифрирование и моделирование
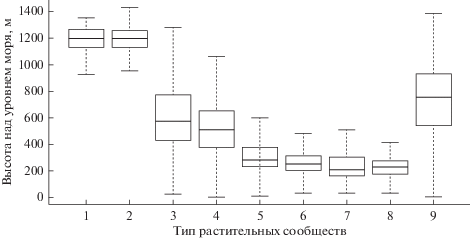
В результате дискриминантного анализа получено 5 осей, в наилучшей степени разделяющих физиономические типы растительности в пространстве признаков, отражаемых каналами съемки и морфометрическими характеристиками рельефа. В итоговой модели при составлении линейных дискриминантных уравнений с использованием алгоритма последовательного отбора признаков выявлено 5 значимых переменных: яркости каналов снимка Landsat 5 TM от 28 июня 2010 г. (каналы 3, 4, 5 и 7 по нумерации сенсора TM) и фактор высоты над уровнем моря (табл. 1). Включение в модель в качестве предикторов значений яркостей спектральных каналов других снимков (весеннего и осеннего), а также показателей крутизны и экспозиции склонов не улучшают ее прогнозных характеристик. Красный канал видимого спектра (3-й) и ближний инфракрасный канал (4-й) связаны с интенсивностью фотосинтеза. Средние инфракрасные каналы SWIR 1 (5-й) и SWIR 2 (7-й) интерпретируются как связанные с влажностью растительного покрова (Tucker, 1979, 1980). Таким образом, ведущими переменными при разделении выделенных физиономических типов растительности оказались интенсивность фотосинтеза растений и содержание влаги в экосистеме, а также высота над уровнем моря (рис. 2).
Таблица 1.
Переменные канонических функций, отобранные в ходе пошагового дискриминантного анализа
| Отобранная переменная | Толерантность | F искл. | λ | Структурные коэффициенты переменных по трем первым осям | ||
|---|---|---|---|---|---|---|
| Ось 1 (53.2%) | Ось 2 (23.4%) | Ось 3 (18.3%) | ||||
| Высота над ур. моря, м | 0.954 | 24.313 | 0.021 | 0.642 | –0.375 | –0.647 |
| Ближний инфракрасный (NIR) | 0.218 | 8.925 | 0.011 | 0.359 | –0.555 | 0.568 |
| Красный (Red) | 0.299 | 4.421 | 0.008 | 0.315 | 0.214 | 0.174 |
| Средний инфракрасный (SWIR 2) | 0.059 | 10.961 | 0.012 | 0.459 | 0.171 | 0.086 |
| Средний инфракрасный (SWIR 1) | 0.051 | 7.592 | 0.010 | 0.400 | -0.226 | 0.213 |
Рис. 2.
Распределение сообществ физиономических типов растительности на градиенте высот. Горизонтальные линии соответствуют медианам, прямоугольники – нижнему (25%) и верхнему (75%) квартилям, “усы” – минимальному и максимальному значениям. Тип растительности: 1 – скальная растительность, 2 – горные тундры, 3 – заросли кедрового и ольхового стлаников, 4 – каменноберезовые леса и криволесья, 5 – пихтово-еловые леса, 6 –мелколиственные и пойменные лиственные леса, 7 – лиственничные леса; 8 – лиственничные редколесья, 9 – луга.
Относительное качество классификации обучающей выборки по спектральным и морфометрическим данным составляет 90% (табл. 2). Для 6 из 9 физиономических типов растительности обучающая выборка классифицирована с максимальной точностью (100%). Пиксели, соответствующие стланиковым сообществам, частично были отнесены к горным тундрам и лиственничным редколесьям. С наименьшей точностью были классифицированы мелколиственные и пойменные леса – треть пикселей, принадлежащая этим сообществам, оказалась отнесена к темнохвойным лесам. Это связано с пространственной сопряженностью сообществ обоих типов в пределах шлейфов склонов Набильского хребта и частичным вкраплением массивов вторичных мелколиственных древостоев в контуры темнохвойных лесов. По той же причине небольшая часть пикселей лугов распозналась как каменноберезовые леса – сообщества соседствуют и постепенно переходят друг в друга по лощинам и тальвегам горных ручьев.
Таблица 2.
Качество классификации обучающей выборки
| Физиономический тип | № | Предсказанная принадлежность, % | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| Растительность скал и осыпей | 1 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Горные тундры | 2 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Стланиковые сообщества | 3 | 0 | 20 | 70 | 0 | 0 | 0 | 0 | 10 | 0 |
| Каменноберезовые леса | 4 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 |
| Пихтово-еловые леса | 5 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 |
| Мелколиственные и пойменные лиственные леса | 6 | 0 | 0 | 0 | 0 | 33.3 | 66.7 | 0 | 0 | 0 |
| Лиственничные леса | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 |
| Лиственничные редколесья | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 |
| Луга | 9 | 0 | 0 | 0 | 18.2 | 0 | 0 | 0 | 0 | 81.8 |
Модель растительного покрова
Согласно созданной картографической модели (рис. 3), площадь лесопокрытой территории в бассейне р. Венгери составляет 89.1% (табл. 3). Реальная площадь поверхности, с учетом кривизны рельефа, больше ее проекции на плоскость почти на 7150 га. Соответственно, растительные сообщества, преимущественно приуроченные к горным склонам, занимают несколько большую площадь.
Рис. 3.
Размещение физиономических типов растительности в бассейне р. Венгери по результатам моделирования: а – скальная растительность, б – горные тундры, в – стланики, г – каменноберезовые леса и криволесья, д – пихтово-еловые леса, е – мелколиственные и пойменные лиственные леса, ж – лиственничные леса, з – лиственничные редины, и – луга, к – высота над ур. моря.
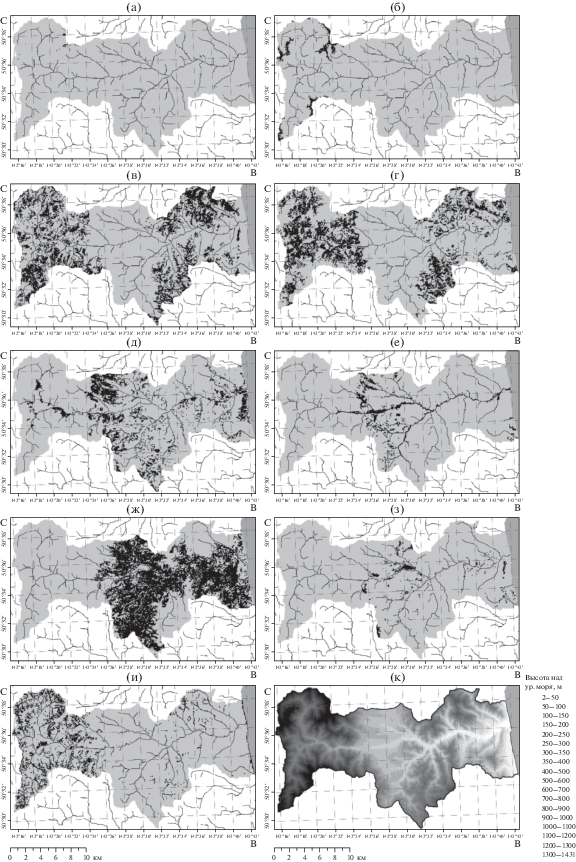
Таблица 3.
Площади сообществ различных физиономических типов растительности
| № | Физиономический тип | S1, га | S2, га | Доля, % | Save ±SD, га | Smin, га | Smax, га |
|---|---|---|---|---|---|---|---|
| 1 | Растительность скал и осыпей | 46.38 | 40.56 | 0.12 | 1.45 ± 0.97 | 0.54 | 4.32 |
| 2 | Горные тундры | 705.20 | 612.98 | 1.85 | 4.64 ± 12.51 | 0.53 | 80.64 |
| 3 | Стланиковые сообщества | 7675.36 | 6824.54 | 20.64 | 3.2 ± 10.090 | 0.50 | 206.64 |
| 4 | Каменноберезовые леса | 7562.85 | 6558.77 | 19.84 | 3.37 ± 8.60 | 0.54 | 238.23 |
| 5 | Пихтово-еловые леса | 4309.54 | 3714.95 | 11.23 | 2.49 ± 7.27 | 0.54 | 193.14 |
| 6 | Мелколиственные и пойменные лиственные леса | 1368.34 | 1345.06 | 4.07 | 1.95 ± 3.53 | 0.54 | 48.78 |
| 7 | Лиственничные леса | 10889.28 | 10363.55 | 31.34 | 5.52 ± 39.08 | 0.52 | 1219.38 |
| 8 | Лиственничные редколесья | 672.94 | 653.09 | 1.98 | 2.11 ± 5.70 | 0.54 | 85.14 |
| 9 | Луга | 6988.37 | 2952.55 | 8.93 | 2.66 ± 5.80 | 0.52 | 87.57 |
Нелесная растительность
В составе нелесной растительности выделено 3 физиономических типа: сообщества скал и осыпей, горных тундр, лугов. Сообщества скальной и осыпной растительности (тип 1) приурочены к выходам коренных пород и денудационным останцам, осыпям, курумам. Сомкнутый покров из сосудистых растений в таких местообитаниях обычно отсутствует. Горные тундры (тип 2) представлены растительными сообществами эрикоидных кустарничков и других арктоальпийских растений (Dryas ajanensis, Empetrum sibiricum, Ledum decumbens, Loiseleuria procumbens, Phyllodoce caerulea, Salix berberifolia, S. reticulata, Sieversia pentapetala и др.). Физиономический тип лугов (тип 9) объединяет травяные сообщества нивальных ложбин, горных склонов и морских террас. Заросли высокотравья (Aconogonon weyrichii, Filipendula camtschatica, Senecio cannabifolius) формируются по местам схода селевых потоков и лавин, в долинах ручьев и горных потоков.
Лесная растительность
Лесная растительность исследованного района образована сообществами как минимум 14 формаций эколого-фитоценотической или 7 союзов эколого-флористической классификации (табл. 4).
Таблица 4.
Принадлежность лесных сообществ к синтаксонам растительности
| Растительное сообщество | Формация согласно эколого-фитоценотической классификации | Синтаксоны согласно эколого-флористической классификации | ||
|---|---|---|---|---|
| Класс | Порядок | Союз | ||
| Сообщества кедрового стланика (3) | Pinetosum pumilae | Vaccinio-Piceetea | Vaccinio-Pinetalia | Vaccinio-Pinion pumilae |
| Сообщества ольхового стланика (3) | Alnetosum fruticosae | Betulo-Ranunculetea | Streptopo-Alnetalia | Smilacino-Betulion ermanii |
| Каменноберезовые леса и криволесья горных склонов (4) | Betuleta ermanii | Betulo-Ranunculetea | Streptopo-Alnetalia | Athyrio-Weigelion middendorffianae |
| Каменноберезовые леса на месте коренных темнохвойных (4) | Betuleta ermanii | неизвестно | неизвестно | неизвестно |
| Пихтово-еловые леса (5) | Piceetosum ajanensis | Vaccinio-Piceetea | Abieti-Piceetalia | Picenion jesoensis |
| Pino-Piceion jezoensis | ||||
| Белоберезовые леса и редколесья на месте коренных темнохвойных (6) | Betuleta platyphyllae | неизвестно | неизвестно | неизвестно |
| Осиновые леса и редколесья на месте коренных темнохвойных (6) | Populeta tremulae | неизвестно | неизвестно | неизвестно |
| Пойменные леса (6) | Saliceta udensis | Salicetea sachalinensis | неизвестно | Filipendulo-Salicionudensis |
| Чозениевые леса (6) | Chosenieta arbutifoliae | |||
| Тополевые леса (6) | Populetosum suaveolentis | |||
| Ольховые леса (6) | Alneta hirsutae | |||
| Черемуховые леса и заросли (6) | Padeta avi | |||
| Лиственничные леса (7) | Lariceta cajanderi | Vaccinio-Piceetea | Ledo-Laricetalia | Ledo-Laricion cajanderi |
| Лиственничные редколесья (8) | Sublariceta cajanderi | |||
Примечание: цифры после названий сообществ указывают на их принадлежность к физиономическим типам растительности (табл. 2, 3; рис. 2).
Стланиковые сообщества (тип 3) представлены как чистыми, так и смешанными зарослями Pinus pumila и Duschekia fruticosa. Они занимают верхние части горных склонов и привершинные участки, водоразделы, седловины. Сообщества P. pumila также развиваются на вершинах и склонах невысоких сопок, если те были пройдены пожарами. В благоприятных условиях высота стлаников может достигать 4–5 м, а в наиболее суровых, например, на подветренных крутых каменистых склонах, она не превышает 0.3–0.4 м. Пространственная структура стланиковых сообществ простая, а флористической состав небогатый. Из кустарников наиболее часто встречается Sorbus sambucifolia. Под плотно сомкнутыми кронами можно обнаружить Chamaepericlymenum canadense, Ledum decumbens, Maianthemum dilatatum, Rubus pedatus, Vaccinium vitis-idaea. Моховой покров развит слабо, представлен широко распространенными видами, такими как Pleurozium schreberi, Ptilium crista-castrensis, Dicranun spp. На старых ветвях произрастает Ptilidium pulcherrimum.
Сообщества каменноберезовых лесов (Betula ermanii, тип 4) занимают средние и верхние части горных склонов, формируют парковые разреженные древостои или криволесья. В долинах и нижних частях склонов представлены вторичными древостоями на месте коренных темнохвойных лесов. Высота древесного яруса зависит от конкретных экологических условий, в долинах может достигать 20 м и более, а в верхних частях горных склонов не превышает 4–5 м. В зависимости от условий местообитаний формируются кустарниковые, крупнотравные и разнотравные типы этих сообществ (Кабанов, 1940). К числу постоянных спутников B. ermanii относятся кустарники Sorbus sambucifolia и Weigela middendorffiana. Нередко кустарниковый ярус образуют стланики Pinus pumila и Duschekia fruticosa. Для крупнотравных вариантов характерно присутствие Aconogonon weyrichii, Cirsium kamtschaticum, Filipendula camtschatica, Senecio cannabifolius. К наиболее увлажненным вариантам горных каменноберезовых лесов приурочены охраняемые реликтовые виды травянистых растений Diphylleia grayi и Macropodium pterospermum. Каменноберезовые криволесья и заросли стлаников могут произрастать на одних и тех же высотах и соседствовать друг с другом, но при этом занимают различные элементы рельефа. Сообщества B. ermanii тяготеют к лощинам и бортам долин горных ручьев, где складываются условия накопления снега и защиты от иссушения, повреждения зимними ветрами.
Темнохвойные пихтово-еловые леса (Abies sachalinensis, Picea ajanensis, тип 5) являются зональными климаксовыми сообществами, но занимают лишь немногим более 11% территории бассейна р. Венгери. Основные массивы приурочены к предгорьям Набильского хребта в центральной части бассейна р. Венгери. Сообщества распространены в нижних частях склонов, на делювиальных шлейфах и коренных террасах речных долинах. В древостоях, помимо двух содоминантов A. sachalinensis и P. ajanensis, присутствуют Betula ermanii и Larix cajanderi. В зависимости от состава подчиненных ярусов выявлены следующие типы пихтово-еловых лесов, ранее подробно описанные Н.Е. Кабановым (1940): моховые (Hylocomium splendens, Pleurozium schreberi, Rhytidiadelphus triquetrus, Dicranum spp.), папоротниковые (Gymnocarpium dryopteris, Leptorumohra amurensis, Phegopteris connectilis), кустарниковые (Sorbus sambucifolia, Vaccinium ovalifolium,) и мелкотравные (Chamaepericlymenum canadense, Maianthemum bifolium, M. dilatatum, Oxalis acetosella). На гниющей древесине развиваются Bazzania trilobata, Macrodiplophyllum plicatum. В рамках эколого-флористической классификации разнообразие сообществ темнохвойных лесов укладывается в объем ассоциаций Weigelo middendorffianae–Piceetum jezoensis Krestov et Nakamura 2002 и Vaccinio vitis-idaeae–Piceetum jezoensis Krestov et Nakamura 2002.
Мелколиственные и пойменные лиственные (согласно А.Г. Крылову (1984)) леса (тип 6) – наиболее разнообразная по составу доминантов древесного яруса группа. Пойменные леса представлены комплексом сообществ с доминированием деревьев из семейства Salicaceae (Chosenia arbutifolia, Populus suaveolens, Salix cardiophylla, S. udensis). На участках речных долин с застойным увлажнением и вдоль тальвегов ручьев формируются сообщества с доминированием Alnus hirsuta, иногда редкостойные, переходящие во влажные заросли крупнотравья. Древостои с доминированием Betula platyphylla и Populus tremula существуют на месте коренных темнохвойных лесов в пределах межгорной депрессии между Набильским и Центральным хребтами, а также на ограниченных участках в долине р. Венгери.
Сообщества лиственничных лесов (Larix cajanderi, тип 7) занимают наибольшие площади в бассейне р. Венгери. Широкое развитие лиственничных лесов на месте коренных темнохвойных объясняется естественными пожарами различной давности. Почти все лиственничные леса исследованного района принадлежат к различным стадиям пирогенных сукцессий, поэтому состав и структура сообществ зависят от времени, прошедшего с момента пожара. Для наиболее молодых лесов отмечена простая вертикальная структура и небогатый видовой состав растений подчиненных ярусов (Pinus pumila, Ledum hypoleucum, Vaccinium vitis-idaeae, Polytrichum spp.). На местах давних пожаров древостой с 2–3 ярусами, второй ярус образован Picea ajanensis, реже Abies sachalinensis. Pinus pumila из состава таких лесов почти полностью выпадает, лишь иногда встречаются отдельные сильно угнетенные особи. В кустарниковом ярусе доминирует Vaccinium ovalifolium, в травяном – виды таежного мелкотравья и папоротники. Бывает выражен почти сплошной покров из зеленых мхов (Hylocomium splendes, Pleurozium schreberii, Dicranum spp.). Непирогенные лиственничники, формирование и существование которых определяется эдафическими условиями, приурочены к местообитаниям с недостаточным дренажем. На отдельных участках по долинам ручьев могут формироваться высокоствольные древостои с доминированием в травяном ярусе Osmun-dastrum asiaticum.
Лиственничные редины (тип 8) – это низкополнотные лишайниковые или кедровостланниковые пирогенные сообщества. Они почти никогда не занимают более или менее крутых склонов, соответствуют выположенным поверхностям, плоским вершинам невысоких сопок, и, по-видимому, формируются в условиях очень хорошего дренажа. Помимо Larix cajanderii в древесном ярусе могут встречаться отдельные деревья Betula platyphylla и Salix caprea. В кустарниковом ярусе присутствуют Pinus pumila и Sorbus sambucifolia, реже – Juniperus sibirica. В лишайниковых лиственничниках кустарниковый ярус обычно не сомкнут, а в кедровостланниковых Pinus pumila на отдельных площадях образует труднопроходимые заросли. В этих сообществах растения нижних ярусов имеют незначительное проективное покрытие. Как правило, в их составе присутствуют: Angelica maximowiczii, Antennaria dioica, Diphasiastrum complanatum, Spiraea betulifolia, Vaccinium vitis-idaea. Мхи встречаются в виде компактных куртин, обычно это Polytrichum spp., иногда Pleurozium schreberii. На отдельных площадях сплошной покров образуют виды рода Cladonia.
Пирогенная трансформация лесной растительности
В пределах бассейна р. Венгери территория, занятая производными лесами с доминированием Larix cajanderi, почти 3 раза превышает площадь зональных темнохвойных лесов из Abies sachalinensis и Picea ajanensis. Общепризнана точка зрения, согласно которой широкое распространение лиственничников на местообитаниях ели – это результат многовекового влияния огня на растительный покров. Как указывает Ю.И. Манько (1987), на Дальнем Востоке России лиственничные леса послепожарного происхождения носят как длительно-устойчивый характер, так и объединены с темнохвойными лесами различными стадиями восстановительных смен. Лиственничные леса обладают высокой горимостью, особенно сообщества сухих лишайниковых вариантов. Неоднократные пожары или беглые палы обычно уничтожают “связующие звенья” лиственничников с климаксовыми темнохвойными сообществами. Широкое развитие лиственничников в районе исследования, таким образом, по всей видимости, связано с частым возникновением естественных лесных пожаров, причиной которых являются обычные для района бассейна р. Венгери сухие грозы.
Особенности моделирования
Реально существующий лесной покров в пределах любого более или менее обширного участка земной поверхности не представляет собой однородного контура. Воздействие естественных факторов нарушений, таких как пожары, ветровалы, очаги развития насекомых-фитофагов, сели, наводнения и многие другие, приводит к формированию комплексной пространственной структуры растительного покрова. Поэтому для создания моделей актуального растительного покрова необходимо использовать не только внешние по отношению к растительным сообществам данные (такие как морфометрические показатели рельефа), но и данные, которые бы соответствовали признакам различных растительных комплексов, находящие отражение в яркостях спектральных каналов или их комбинаций.
Связанное с глубоким расчленением рельефа затенение поверхности горных склонов вносит существенные искажения в результаты автоматического дешифрирования и космических снимков и основанного на нем моделирования. Как показывает опыт нашей работы, в случае моделирования растительного покрова на относительно небольших по площади территориях, проблема наличия сильно затененных участков может быть решена путем экспертной оценки и визуального дешифрирования снимков высокого разрешения. Поэтому именно склонам северной экспозиции необходимо уделять особое внимание в ходе проведения этапа сбора полевых данных.
ЗАКЛЮЧЕНИЕ
Использование цифровой модели рельефа и спектральных характеристик растительных сообществ позволило провести моделирование актуального растительного покрова бассейна р. Венгери. Горный рельеф территории определяет существование высотных поясов растительного покрова. Наиболее возвышенные и крутосклонные формы рельефа заняты стланиковыми сообществами (Dushekia fruticosa, Pinus pumila) и древостоями из каменной березы (Betula ermanii). Нижний высотный пояс занят зональными темнохвойными лесами из Abies sachalinensis и Picea ajanensis, но на их долю приходится только около 11% территории речного бассейна. Более 33% территории речного бассейна занимают леса и редколесья из Larix cajanderi, в основном послепожарного происхождения. Поскольку пространственная структура растительного покрова зависит не только от ландшафтно-климатических факторов, но также и от истории нарушений (в т.ч. от лесных пожаров), то создание моделей актуального растительного покрова не может быть выполнено только на основании внешних по отношению к растительному покрову признаков. Безошибочное автоматическое дешифрирование и моделирование растительного покрова в регионах с горным рельефом невозможно из-за сильного затенения и отсутствия отражения от находящихся в глубокой тени склонов. Восстановление актуальной растительности для таких площадей возможно провести экспертным путем, с использованием снимков высокого разрешения и привлечением данных полевых исследований, на что необходимо обращать внимание при планировании экспедиционных работ.
Список литературы
Бакалин В.А. Конспект печеночников // Бриофлора Сахалина. Владивосток: Морской гос. университет, 2012. С. 44–112.
Зоны и типы поясности растительности России и сопредельных стран. Пояснительный текст и легенда карты / Отв. ред. Огуреева Г.Н. М.: ЭКОР, 1999. 64 с.
Исаев А.С., Князева С.В., Пузаченко М.Ю., Черненькова Т.В. Использование спутниковых данных для мониторинга биоразнообразия лесов // Исследование Земли из космоса. 2009. № 2. С. 55–66.
Кабанов Н.Е. Лесная растительность Советского Сахалина. Владивосток: Горнотаежная станция АН СССР, 1940. 210 с.
Казаков Н.А., Генсиоровский Ю.В. Влияние вертикального градиента осадков на характеристики гидрологических, лавинных и селевых процессов в низкогорье // Геоэкология, инженерная геология, гидрогеология, геокриология. 2007. № 4. С. 342–347.
Комарова А.Ф., Куксина Н.В., Зудкин А.Г. Картографирование темнохвойных лесов Северо-Западного Кавказа методом нейронных сетей // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 13. № 5. С. 157–166.
Крылов А.Г. Жизненные формы лесных фитоценозов. Л.: Наука, 1984. 184 с.
Манько Ю.И. Ель аянская. Л.: Наука, 1987. 280 с.
Нешатаева В.Ю. Растительность полуострова Камчатка. М.: Товарищество научных изданий КМК, 2009. 537 с.
Омелько А.М., Якубов В.В., Бакалин В.А., Великанов А.В., Черданцева В.Я., Скирина И.Ф., Яковлевка А.Н., Крестов П.В. Растительный покров Ланжинских гор (Охотия) // Комаровские чтения. 2010. Т. 57. С. 103–163.
Писаренко О.Ю., Игнатов М.С., Черданцева В.Я., Игнатова Е.А. Конспект мхов // Бриофлора Сахалина. Владивосток: Морской гос. ун-т, 2012. С. 123–268.
Пузаченко М.Ю., Черненькова Т.В. Определение факторов пространственного варьирования растительного покрова с использованием ДДЗ, ЦМР и полевых данных на примере центральной части Мурманской области // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 13. № 5. С. 167–191.
Пузаченко Ю.Г., Сандлерский Р.Б., Кренке А.Н., Пузаченко М.Ю. Мультиспектральная дистанционная информация в исследовании лесов // Лесоведение. 2014. № 5. С. 13–29.
Тихомиров Б.А. К происхождению ассоциаций кедрового стланика (Pinus pumila Rgl.) // Материалы по истории флоры и растительности СССР. Вып. 2. М.; Л.: Изд-во АН СССР, 1946. С. 491–537.
Харук В.И., Щербинина И.С., Якубайлик О.Э., Касишке Э. Анализ пространственного распределения гарей на острове Сахалин // Лесоведение. 2004. № 5. С. 66–80.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
Черненькова Т.В., Морозова О.В. Классификация и картографирование ценотического разнообразия лесов // Лесоведение. 2017. № 4. С. 243–255.
Bartalev S.A., Belward A.S., Erchov D.V., Isaev A.S. A new SPOT4-VEGETATION derived land cover map of Northern Eurasia // International J. Remote Sensing. 2003. V. 24. № 9. P. 1977–1982.
Belyaeva N.G., Chernen`kova T.V. Relationship between habitat properties and composition of communities in conifer-broadleaf Forest // Russian J. Ecology. 2018 V. 49. № 2. P. 111–118.
Chernen`kova T.V., Basova E.V., Puzachenko M.Y., Koroleva N.E. Assessment of forest spatial differentiation in Murmansk Province using field surveys and remote sensing data // Contemporary Problems of Ecology. 2013. V. 6. № 7. P. 746–754.
Chernen`kova T.V., Puzachenko M.Yu., Morozova O.V., Ogureeva G.N., Kuperman R.G. // An approach for mapping Northern Fennoscandian forests at different scales // Botanica Pacifica. 2015. V. 4. № 1. P. 37–46.
Giles P. Remote sensing and cast shadows in mountainous terrain // Photogrammetric Engineering & Remote Sensing. 2001. V. 67. № 7. P. 833–839.
Gomes C., White J.C., Wulder M.A. Optical remotely sensed time series data for land cover classification: A review // ISP-RS J. Photogrammetry and Remote Sensing. 2016. V. 116. P. 55–72.
Hansen M.C., Townshend J.R.G., DeFries R.S., Carroll M. Estimation of tree cover using MODIS data at global, continental and regional/local scales // International J. Remote Sensing. 2005. V. 26. № 19. P. 4359–4380.
Krestov P.V., Nakamura Yu. Phytosociological study of the Picea jezoensis forests of the Far East // Folia Geobotanica. 2002. V. 37. № 4. P. 441–473.
Krestov P.V., Ermakov N.B., Osipov S.V., Nakamura Yu. Classification and phytogeography of larch forests of Northeast Asia // Folia Geobotanica. 2009. V. 44. № 4. P. 323–363.
McRoberts R.E., Cohen W.B., Naesset E., Stehman S.V., Tomppo E.O. Using remotely sensed data to construct and assess forest attribute maps and related spatial products // Scandinavian J. Forest Research. 2010. V. 25. № 4. P. 340–367.
Omelko A.M., Krestov P.V., Yakovleva A.N. A topography-based model of the vegetation cover of the Lanzhinskie mountains // Botanica Pacifica. 2012. V. 1. № 1. P. 109–119.
Reese H., Nilsso M., Sandströ P., Olsso H. Applications using estimates of forest parameters derived from satellite and forest inventory data // Computers & Electronics in Agriculture. 2002. V. 37. № 1–3. P. 37–56.
Riano D., Chuvieco E., Salas J., Aduado I. Assessment of different topographic corrections on Landsat-TM data for mapping vegetation types // IEEE Transactions on Geoscience & Remote Sensing. 2003. V. 41. № 5. P. 1056–1061.
Richter R., Kellenberger T., Kaufmann H. Comparison of topographic correction methods // Remote Sensing. 2009. V. 7. № 1. P. 184–196.
Shahtahmassebi A., Yang N., Wang K., Moore N., Zhangquan Sh. Review of shadow detection and de-shadowing methods in remote sensing // Chinese Geographical Science. 2013. V. 23. № 4. P. 403–420.
Tucker C.J. Red and photographic infrared linear combinations for monitoring vegetation // Remote Sensing of Environment. 1979. V. 8. № 2. P. 127–150.
Tucker C.J. Remote sensing of leaf water content in the near-infared // Remote Sensing of Environments. 1980. V. 10. № 1. P. 23–32.
Vegetation of Japan. Vol. 9 Hokkaido Ed. A. Miyawaki. Tokyo, Shibundo, 1988. 563 p.
Zhu Z., Fu Y., Woodcock C.E., Olofsson P., Vogelmann J.E., Holden C., Wang M., Dai S., Yu Y. Including land cover change in analysis of greenness trends using all available Landsat 5, 7, and 8 images: A case study from Guangzhou, China (2000–2014) // Remote Sensing of Environment. 2016. V. 185. P. 243–257.
Yakovleva A.N. A model of vegetation pattern at the Verkhneussuriysky Biogeocenotic Station // Russian J. Ecology. 2010. V. 41. № 4. P. 307–315.
Дополнительные материалы отсутствуют.