

Лесоведение, 2020, № 6, стр. 483-492
Радиальный прирост и анатомическая структура древесины стволов здоровых и суховершинных деревьев лиственницы на многолетней мерзлоте
Д. А. Машуков a, *, А. В. Бенькова a, В. Е. Бенькова a, А. В. Шашкин a, А. С. Прокушкин a
a Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: mashukov1988@gmail.com
Поступила в редакцию 15.03.2018
После доработки 22.05.2018
Принята к публикации 06.06.2020
Аннотация
Приведены результаты исследования динамики радиального прироста и ксилотомической структуры вдоль ствола суховершинных и здоровых деревьев лиственницы Гмелина (Larix gmelinii (Rupr.) Rupr.), произрастающих в одновозрастном древостое (64°19′23″ с.ш., 100°13′28″ в.д.) на многолетнемерзлой почве склона северной экспозиции. Измерены ширина годичных колец, радиальные размеры люменов и стенок трахеид на 1/4, 1/2, 3/4 высоты стволов и у верхушки на двадцати пяти не поврежденных и суховершинных деревьях. Выявлен отрицательный градиент средних значений этих показателей от комля к верхушке, особенно четко выраженный у суховершинных деревьев. В годичных кольцах на верхних уровнях высоты стволов за последние 20 лет у суховершинных деревьев произошли модификации анатомической структуры: нарушение рядности трахеид, утоньшение стенок ранних и поздних трахеид. По скользящим климатическим функциям отклика радиального прироста показано, что деревья на склоне северной экспозиции с конца мая до двадцатых чисел июня испытывают водный дефицит. Наличие суховершинных и здоровых деревьев на склоне северной экспозиции может быть обусловлено высокой вариабельностью гидротермических свойств почвы в связи с высокой пространственной мозаичностью мохово-лишайникового покрова. Со временем обостренная реакция некоторых деревьев на водный дефицит может привести к усыханию верхушек.
В северо-таежных древостоях, развивающихся на многолетней мерзлоте, распространено явление суховершинности. Одна из возможных причин появления суховершинности – острый водный дефицит (Поздняков, 1983; Николаев и др., 2011), который могут испытывать деревья в специфических гидротермических условиях многолетнемерзлых почв. Несмотря на обилие влаги в сезонно-талом слое, деревья могут испытывать “физиологическую засуху” (Kramer, Boyer, 1995) из-за низкой температуры почвенной воды, которая обусловливает пониженную вязкость последней и пониженную функциональную активность корневой системы. При этом верхушки деревьев, имеющие низкий водный потенциал по сравнению с расположенными ниже частями дерева (Kramer, Boyer, 1995), оказываются в наиболее неблагоприятных условиях. Недостаточное водообеспечение верхушек может привести к усыханию.
Как было показано в работах по исследованию северо-таежных лесов, наиболее неблагоприятные для роста деревьев гидротермические почвенные условия сложились на склоне северной экспозиции, где, по сравнению со склоном южной экспозиции, сравнительно низкая поступающая солнечная энергия и мощный теплоизолирующий мохово-лишайниковый покров обусловливают небольшую глубину сезонно-талого слоя, который имеет низкую температуру в течение всего вегетационного сезона. Деревья, произрастающие на склонах северной экспозиции, находятся в угнетенном состоянии, часто имеют усохшие верхушки, характеризуются низкими таксационно-биометрическими показателями и скоростями роста в высоту и по диаметру (Yanagihara и др., 2000), отличаются низкой интенсивностью фотосинтеза (Koike et al., 2010).
Данная статья посвящена выявлению различий между суховершинными и здоровыми деревьями, произрастающими на склоне северной экспозиции, по динамике их радиального роста и анатомическому строению древесины вдоль ствола от комля к верхушке и является продолжением ранее проведенных исследований (Бенькова и др., 2015). Действительно, движение воды снизу вверх по стволу происходит за счет уменьшения водного потенциала в направлении от комля к вершине (Kramer, Boyer, 1995). Скорость уменьшения этого показателя определяет разность между водным потенциалом листа, зависящем от транспирации, и водным потенциалом почвы. Разный водный потенциал вдоль ствола дерева обусловливает различия в функционировании камбия и дифференциации его производных. На камбиальную активность косвенно, через количество доступной влаги в почве (водный потенциал почвы), влияют и погодные условия. Таким образом, в характеристиках радиальных приростов на разных уровнях высоты ствола “зашиты” эти оба вида воздействия на камбий, что не может не отразиться на анатомической структуре годичных колец. В задачи работы входило выявить различия в динамике радиального роста (интенсивности и чувствительности к климатическим факторам) и анатомической структуре годичных колец на разной высоте стволов у суховершинных и здоровых деревьев лиственницы Гмелина, произрастающих на многолетней мерзлоте в экстремальных гидротермических условиях склона северной экспозиции.
ОБЪЕКТЫ И МЕТОДИКА
Исследование проводилось в бассейне р. Нижняя Тунгуска, вблизи пос. Тура, на базе Эвенкийского опорного экспедиционного пункта Института леса СО РАН (64°19′23″ с.ш., 100°13′28″ в.д.), в лиственничнике, который сформировался после сильного низового пожара 1899 г. (датировка пожара проведена А.А. Кнорре с соавт. (2009)) на мерзлотных почвах склона северной экспозиции с уклоном 7°–8°, расположенного в нижнем течении ручья Кулингдакан. Лиственничник характеризуется как багульниково-бруснично-голубичный лишайниково-зеленомошный. Подлесок представлен ивами боганидской и енисейской (Salix boganidensis Trautv., S. jenisseensis (Fr. Schmidt) B. Floder), распространен по площади равномерно. Напочвенный покров относительно однороден, общее проективное покрытие 35–40%. Мохово-лишайниковый покров представлен в основном Pleurozium schreberi (Brid.) Mitt. и Aulocomnium turgidum (Hedw.) Schwaegr, в меньшей степени Hylocomium splendens (Hedw.) B.S.G. Густота древостоя 4075 дер. га–1, полнота 0.43, запас 41.7 м3 га–1, бонитет Vб. Более 60% деревьев лиственницы было с сухими верхушками. Общая климатическая характеристика района исследования, более подробное описание пробной площади и древостоя даны в работе А.В. Беньковой с соавт. (2015).
Почвы имеют среднесуглинистый гранулометрический состав (доля физической глины 41%), характеризуются повышенной влажностью и наличием надмерзлотного стока воды. Сезонное оттаивание зависит от мозаичности напочвенного покрова и микрорельефа (Ловелиус, 1978) и в среднем составляет 42 ± 10 см в буграх мерзлотного пучения и 6 ± 6 см в западинах. Мощность органического горизонта варьирует в пределах 11–25 см. Глубина корнеобитаемого слоя не превышает 25 см (особенности расположения корневой системы деревьев лиственницы на поверхности и внутри сезонно-талого слоя исследовались С.Г. Прокушкиным с соавт. (2008) и T. Kajimoto (2010)). Сравнительно небольшая глубина сезонного оттаивания почвы обусловлена наличием развитого мохово-лишайникового покрова (7–15 см), препятствующего прогреву. В течение всего вегетационного периода почва имеет довольно низкую температуру. Так, на конец июля (к этому времени уже завершаются основные процессы формирования годичного прироста) почва на глубине 10 см прогрелась всего до 6.5°С, а объемная влажность составила 40%.
На пробной площади отобрали тринадцать не поврежденных суховершинностью и двенадцать соседних с ними суховершинных деревьев. На момент взятия образцов (07.2009) деревья с сухими верхушками отстали в росте от соседних здоровых деревьев: средние значения диаметра на высоте груди у первых были на 27% меньше (47.4 и 65.2 мм соответственно), высоты – на 26% меньше (7.36 и 8.29 м), протяженности кроны на 14.6% меньше (3.79 и 4.44 м).
Со стволов выбранных деревьев лиственницы взяли по пять спилов: у комля, на 1/4, 1/2, 3/4 высоты ствола дерева и у верхушки, на 20–30 см ниже апекса. Спилы, взятые у комля, использовали для определения возраста деревьев, но из дендроэкологического анализа они были исключены, так как на особенности формирования годичных приростов древесины в комлевой части ствола существенно влияет близко расположенная корневая система. Место, с которого брался самый верхний спил у суховершинного дерева, внешне не имело визуально диагностируемых признаков усыхания, тогда как на 1–3 см выше него эти признаки были выраженными. Всего было исследовано 65 спилов с неповрежденных и 60 – с суховершинных деревьев.
На спилах по двум радиусам измерили ширину годичных колец. Бо́льшую часть измерений провели на измерительном комплексе LINTAB v3.0 (точность 0.01 мм). Ширину очень узких и плохо различимых годичных колец, которые сформировались у верхушек, измерили в проходящем свете на установке Axio Imager MAT, Version 04.14.2005 (Karl Zeis Light AGMicroscopy (точность 0.003 мм) по микропрепаратам, содержащим поперечные микросрезы древесины толщиной 15–25 мкм. Для резки использовался санный микротом, для окрашивания – сафранин.
Построение индексированных древесно-кольцевых хронологий осуществляли стандартными методами (Fritts, 1976; Cook, 1990; Шиятов и др., 2000) с использованием общепринятых в дендрохронологии программ Arstan и Cofecha. Возрастной тренд был аппроксимирован отрицательной экспоненциальной функцией. Индексированные хронологии анализировали во временно́м интервале 1934–2009 г., общем для хронологий на всех уровнях высоты стволов, за исключением самого верхнего, “верхушки” (при выборе интервала учитывалось, что суточные климатические данные по метеостанции “Тура” в доступной базе данных представлены с 1934 г.). Общий период для хронологий “верхушек” – 1974–2009 г.
С целью выделения периодов времени в пределах сезона вегетации со значимым влиянием климатических факторов (температуры воздуха и количества осадков) на радиальный прирост рассчитывали скользящие корреляционные климатические функции отклика. Расчеты проводили c “окном” в 20 дней и “шагом” в 5 дней с 10 апреля по 28 августа, т.е., за период, когда климатические факторы могут заметно повлиять на рост годичного кольца. Ранее было показано, что для мест с коротким сезоном вегетации эти значения “окна” являются оптимальными (Симанько и др., 2013). Суточные значения температуры воздуха и количества осадков по метеостанции “Тура” взяты с сайта “National Weather Service. Internet Weather Source”” ftp:/ftp.ncdc.noaa.gov/pub/data/ghcn/daily.
Для проведения ксилотомических исследований использовали те же микропрепараты, что и для измерения ширины очень узких годичных колец. Измерения радиального размера люменов трахеид и толщины их стенок проводили в проходящем свете на упомянутом выше оборудовании Axio Imager MAT с точностью 0.003 мм.
Изменение температуры активных горизонтов почвы проводилось в непрерывном режиме термодатчиками TR–51–A (Япония), которые были установлены непосредственно под мохово-лишайниковым покровом и на глубине 5 и 10 см.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Индексированные хронологии радиального прироста на исследованных уровнях высоты стволов суховершинных и здоровых деревьев характеризуются стандартным набором показателей. Коэффициенты чувствительности всех хронологий превышают значение 0.20 (табл. 1), что свидетельствует о выраженной реакции деревьев на влияние климатических факторов (Шиятов, 1986). Судя по значениям коэффициента чувствительности (табл. 1), “чувствительность” радиального прироста к влиянию изменяющихся погодных условий у суховершинных деревьев выше, чем у здоровых (при Р ≥ ≥ 0.95) на всех уровнях высоты ствола (за исключением самого верхнего, “верхушки”). В то же время у здоровых деревьев радиальный прирост у верхушки отличается большей “чувствительностью” (при Р ≥ 0.95) по сравнению с приростом на нижних уровнях высоты, а у суховершинных деревьев такая тенденция не прослеживается.
Таблица 1.
Характеристики индексированных хронологий радиального прироста на 1/4, 1/2, 3/4 уровнях высоты ствола (1934–2009 г.) и у верхушки (1974–2009 г.) деревьев лиственницы Гмелина на склоне северной экспозиции
| Уровень высоты ствола | Коэффициент | Коэффициент | ||
|---|---|---|---|---|
| вариации | чувствительности | вариации | чувствительности | |
| здоровые деревья | суховершинные деревья | |||
| 1/4 | 0.48 | 0.29 | 0.64 | 0.33 |
| 1/2 | 0.60 | 0.24 | 0.75 | 0.29 |
| 3/4 | 0.60 | 0.24 | 0.87 | 0.29 |
| “Верхушка” | 0.20 | 0.34 | 0.28 | 0.29 |
Вариабельность радиального прироста (судя по значениям коэффициента вариации) у суховершинных деревьев выше, чем у здоровых, на всех уровнях высоты ствола (табл. 1).
У суховершинных деревьев средняя ширина годичных колец на всех уровнях высоты ствола заметно меньше (при P ≥ 0.95), чем у здоровых (табл. 2): годичные кольца у́же в 1.5 раз на уровне 1/4 высоты ствола и в 2.5 раз – у верхушки. Очень узкий прирост вблизи сухой верхушки указывает на сравнительно низкую камбиальную активность.
Таблица 2.
Средняя за 1974–2009 г. ширина годичных колец на разных уровнях высоты ствола суховершинных и здоровых деревьев на склоне северной экспозиции
| Уровень высоты ствола | Ширина годичных колец, мм | |
|---|---|---|
| здоровые деревья | суховершинные деревья | |
| “Комель” | 0.20 (0.05) | 0.140 (0.039) |
| 1/4 | 0.14 (0.04) | 0.095 (0.023) |
| 1/2 | 0.11 (0.02) | 0.081 (0.020) |
| 3/4 | 0.12 (0.02) | 0.066 (0.013) |
| “Верхушка” | 0.12 (0.03) | 0.048 (0.014) |
Примечание. Здесь и в табл. 2: В скобках приведено стандартное отклонение.
Корреляционная связь между индексированными хронологиями, полученными на одинаковых уровнях высоты ствола у здоровых и суховершинных деревьев, довольно тесная: при Р ≥ 0.95 коэффициенты парной корреляции составили от 0.67 до 0.89. Очевидно, что заметное влияние на радиальный рост здоровых и суховершинных деревьев на соответствующих уровнях высоты ствола оказывают одинаковые внешние (главным образом, климатические) факторы. Корреляция индексированной хронологии радиального прироста у верхушек с таковыми на нижних уровнях высоты ствола у суховершинных и здоровых деревьев также довольно высока, но у суховершинных деревьев она гораздо слабее (табл. 3). По нашему мнению, этот эффект может быть обусловлен, например, водным дефицитом, отрицательное влияние которого, как известно (например, Kramer, Boyer, 1995), в большей степени сказывается на верхушках деревьев, и суховершинные деревья оказались к нему более чувствительными. Это не могло не сказаться на анатомическом строении стволовой древесины.
Таблица 3.
Коэффициенты парной корреляции между индексированной хронологией радиального прироста у верхушки и хронологиями на 1/4, 1/2, 3/4 уровнях высоты ствола здоровых и суховершинных деревьев (1974–2009 г.)
| Уровень высоты ствола | Здоровые деревья | Суховершинные деревья |
|---|---|---|
| 1/4 | 0.60 | 0.45 |
| 1/2 | 0.55 | 0.34 |
| 3/4 | 0.85 | 0.62 |
Примечание. Здесь и в табл. 3: Р ≥ 0.95.
Действительно, в строении годичных колец на верхних уровнях высоты стволов (на 3/4 высоты и у верхушки) суховершинных деревьев мы обнаружили признаки “нарушенности” (рис. 1а, 1б), которые отсутствовали на нижних уровнях (на 1/2 и 1/4 высоты ствола). Оказалось, что у разных деревьев “нарушенности” в строении древесины появились не одновременно, а с разбросом в несколько лет, с 1985 по 1997 г. Основные признаки “нарушенности”: нарушается рядность трахеид (стрелки на рис. 1а, 1б); становятся более тонкими стенки ранних и поздних трахеид и со временем позднюю зону в годичных кольцах, так же как и сами годичные кольца, трудно, а иногда невозможно различить.
Рис. 1.
Ксилотомическое строение годичных колец под верхушкой (а), на 3/4 высоты (б) и на 1/2 высоты ствола (в) суховершинных деревьев лиственницы. Стрелками указан год появления нарушений строения: а – 1994 г.; б – 1989 г. На 1/2 высоты (в) нарушенности отсутствуют (последнее годичное кольцо на а, б и в образовано в один и тот же год).

Сравнительный анализ анатомических характеристик древесины ствола (радиального размера люмена и толщины стенки трахеид), средних за период 1985–2009 гг. (рис. 2), характеризующихся наличием “нарушенностей” в строении годичных колец, выявил следующее. У обеих групп деревьев средний размер люмена трахеид и толщина их стенок уменьшается по высоте ствола. Убывающий тренд люмена снизу вверх по стволу является типичным для всех деревьев (Schweingruber, 1996; Антонова, 1999). В нашем случае он более четко выражен у суховершинных деревьев по сравнению со здоровыми (рис. 2). Средние значения размера люмена и толщины стенок трахеид под усохшей верхушкой в 1.5 и 2 раза (соответственно) меньше, чем на 1/4 высоты ствола, а под неповрежденной верхушкой – в 1.2 и 1.4 раза меньше. Обе группы деревьев слабо различаются по среднему радиальному размеру люмена и средней толщине стенок трахеид в нижней части ствола (на 1/4 высоты). Различие между обеими группами деревьев четко проявилось вблизи верхушек: у суховершинных деревьев средний радиальный размер люмена достоверно (при P ≥ 0/95) меньше (в 1.3 раза), а стенки трахеид примерно в 2 раза тоньше, чем у здоровых.
Рис. 2.
Средние за период 1985–2009 гг. радиальный размер люмена и толщина клеточной стенки трахеид на высоте 1/4 ◼ и верхушки ◻ у суховершинных (а, в) и несуховершинных (б, г) деревьев.
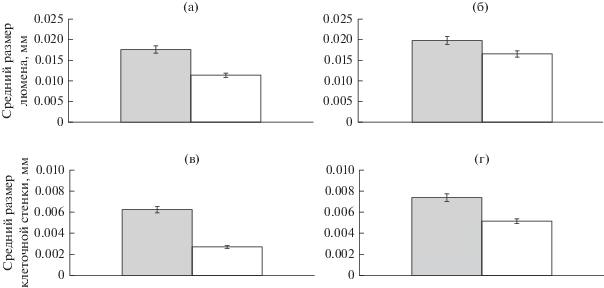
Отмеченные особенности могут быть вызваны недостатком воды, поступающей к верхним частям ствола деревьев, и пониженным их водным потенциалом во время процесса дифференциации клеток камбия (Larson, 1963). Низкий водный потенциал обусловливает низкий тургор, при котором клетки растут сравнительно медленно и делятся с меньшей скоростью (Nonami, Boyer, 1990; Lockhart, 1965). В результате формируются сравнительно узкие годичные кольца, состоящие из малочисленных мелких узкополостных трахеид. Эффективность водопроведения таких трахеид низкая, и это усугубляет дефицит поставляемой к верхушкам влаги. Последний приводит к уменьшению фотосинтеза, и, соответственно, углеводов, расходуемых на синтез стенок трахеид.
Для выявления различия в реакции суховершинных и здоровых деревьев на климатические факторы использовались скользящие корреляционные климатические функции отклика индексов радиального прироста, поскольку они детально отражают внутрисезонное влияние погодных условий (рис. 3). В связи с тем, что появление признаков нарушенности в строении годичных колец датировалось второй половиной 1980-х годов, расчет функций отклика проводили за 1985–2009 г. На рис. 3 в качестве примера представлены три уровня высоты ствола: 1/4, 3/4 и “верхушка”.
Рис. 3.
Скользящие 20-дневные корреляционные климатические функции отклика индексов радиального прироста суховершинных (а, в) и здоровых деревьев (б, г) на уровнях 1/4 (1), 3/4 (2) высоты ствола и у верхушки (3) на температуру воздуха и количество осадков. 4 и 5 – среднемноголетние значения среднесуточной температуры и суточных осадков, “сглаженные” в окне в 20 дней (1985–2009 гг.). Коэффициенты корреляции R ≥ 0.41 значимы при P ≥ 0.95.
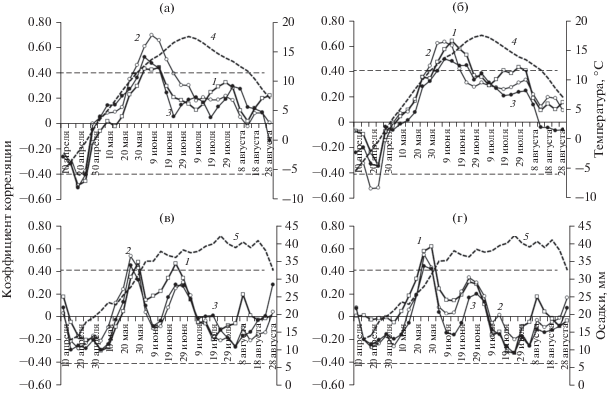
Выделяются два временны́х отрезка, когда радиальный прирост суховершинных и здоровых деревьев достоверно коррелирует с климатическими факторами (рис. 3). Первый период длится примерно две недели (третья декада апреля – начало мая) и характеризуется отрицательной корреляцией с температурой воздуха. У суховершинных деревьев радиальный прирост на всех уровнях высоты ствола. а у здоровых деревьев – на уровне 3/4 отрицательно связан с температурой воздуха. Отмеченный период предваряет начало радиального роста лиственницы Гмелина в данной местности (Yasue et al., 2010). Относительно высокие (выше среднемноголетних) температуры воздуха в конце апреля могут “спровоцировать” начало процесса предсезонной реактивации камбия. Однако за оттепелью неизбежно следуют морозные периоды; процесс реактивации камбия может быть приостановлен, что негативно отразится на ширине и анатомическом строении годичного кольца. Часто встречающиеся морозобойные кольца у деревьев лиственницы на полярной границе леса (Гурская, Бенькова, 2013) могут рассматриваться как подтверждение негативного влияния на камбий повышенной температуры воздуха ранней весной.
Второй период характеризуется положительной корреляцией радиального прироста на всех уровнях высоты ствола с обоими климатическими факторами. Периоды заметного влияния температуры воздуха на прирост на разной высоте ствола у здоровых и суховершинных деревьев могут незначительно различаться по срокам и длительности (рис. 3). Так, у суховершинных деревьев температурное влияние на прирост на 3/4 высоты ствола сказывается с конца мая до двадцатых чисел июня, на прирост у верхушки и в нижней части ствола начинает проявляться примерно на 5 дней позже, в начале июня, и заканчивает примерно на пять дней раньше. У здоровых деревьев влияние температуры на прирост на 3/4 высоты ствола продолжается до середины июня, на прирост у верхушки и в нижней части ствола – до двадцатых чисел июня.
Период положительного влияния количества осадков на прирост гораздо короче, чем температуры, и проявляется раньше: это последняя декада мая (рис. 3). Для приростов на всех исследуемых уровнях высоты у суховершинных и здоровых деревьев этот период один и тот же. У суховершинных и здоровых деревьев выделяется еще один период с положительной корреляцией прироста с количеством осадков: это двадцатые числа июня – самое теплое и достаточно сухое время сезона (кривые 3 и 4). Однако влияние осадков заметно проявляется только на уровне 1/4 высоты ствола.
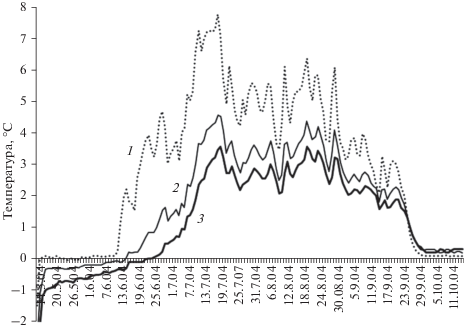
В конце мая–начале июня возобновляются физиологические процессы в деревьях после зимнего покоя. Это время старта и интенсивного радиального роста лиственницы Гмелина в исследуемом районе (Yasue et al., 2010). Между тем на склоне северной экспозиции деятельный слой почвы имеет температуру, близкую к нулю (рис. 4). Основным источником воды для растений в этот период оказываются осадки. Они имеют более высокую температуру, чем температура почвы, и ускоряют ее оттаивание. Обильные осадки и повышенная температура воздуха в этот период сезона оказывают заметное положительное влияние на темпы радиального роста суховершинных и здоровых деревьев.
Рис. 4.
Среднесуточная температура под моховым покровом на поверхности почвы (1), на глубине 10 см (2) и 20 см (3) в сезон 2004 г.
В более поздний период сезона, со второй декады июня, повышенные температуры воздуха обусловливают поступление к корням деревьев сравнительно теплой воды, что облегчает ее всасывание; высокие температуры воздуха к тому же определяют высокую интенсивность фотосинтеза и образование значительного количества метаболитов – все это положительно сказывается на радиальном приросте. Отсутствие заметной реакции у деревьев на температуру позже, в июле, может означать, что либо к этому времени годичные кольца сформировались, либо что температура уже не лимитирует рост и на нeм в большей мере сказываются осадки. Суховершинные деревья оказались к ним более чувствительными (по радиальному росту), но только в нижней части ствола, где сравнительно высокий водный потенциал и значительный радиальный прирост (Nonami, Boyer, 1990).
ЗАКЛЮЧЕНИЕ
Результаты анализа анатомического строения стволовой древесины и скользящих климатических функций отклика радиального прироста находятся в согласии с гипотезой о том, что все деревья на склоне северной экспозиции при пониженных (ниже среднемноголетних) значениях температуры воздуха и количества осадков с конца мая до двадцатых чисел июня испытывают водный дефицит. Отрицательное влияние последнего сказалось на деревьях в древостое по-разному. Разная реакция деревьев проявилась не столько в зависимости радиального прироста от климатических факторов, сколько в анатомическом строении древесины на верхних уровнях высоты ствола (на 3/4 высоты и под усохшей верхушкой). Появление нарушенностей в анатомическом строении годичных приростов древесины свидетельствует о нарушении самого процесса дифференциации меристематических клеток у определенных деревьев, верхние части стволов которых оказались в более жестких по водообеспечению условиях. В конечном итоге со временем такая реакция деревьев может привести к усыханию верхушек. Неодинаковые для всех деревьев в древостое условия произрастания могут быть связаны высокой пространственной неоднородностью гидротермических почвенных характеристик, связанной с высокой пространственной мозаичностью мохово-лишайникового покрова. Вариабельность его термоизоляционных свойств определяется пространственным распределением и соотношением микроассоциаций видов-доминантов Pleurozium schreberi, Aulocomnium turgidum, Hylocomium splendens (Зырянова и др., 2006).
Список литературы
Антонова Г.Ф. Рост клеток хвойных. Новосибирск: Наука, 1999. 232 с.
Бенькова А.В., Машуков Д.А., Бенькова В.Е., Прокушкин А.С., Шашкин А.В. Значение экспозиции склонов для роста лиственницы Гмелина в мерзлотных условиях Средней Сибири I. Различия в динамике радиального прироста деревьев на склонах северной и южной экспозиции // Сибирский лесной журн. 2015. № 4. С. 18–29.
Гурская М.А., Бенькова В.Е. Типы светлых колец у Larix sibirica и L. gmelinii на верхней границе леса в урало-сибирской субарктике // Ботанический журнал. 2013. Т. 98. № 8. С. 1037–1054.
Зырянова О.А., Абаимов А.П., Бугаенко Т.Н. Оценка видового разнообразия и структуры лиственничных ассоциаций криолитозоны Сибири // Биоразнообразие и динамика экосистем: информационные технологии и моделирование. Новосибирск: Наука, 2006. С. 495–504.
Кнорре А.А., Кирдянов А.В., Прокушкин А.С. Оценка скорости подъема мерзлоты в лиственничниках Центральной Эвенкии методами дендрохронологии // Лесоведение. 2009. № 2. С. 77–80.
Ловелиус Н.В. Снежный покров и мерзлота // Ары-Мас: Природные условия, флора и растительность самого северного в мире лесного массива. Л.: Наука, 1978. С. 21–30.
Николаев А.Н., Федоров П.П., Десяткин А.Р. Влияние гидродинамического режима мерзлотных почв на радиальный прирост лиственницы и сосны в центральной Якутии // Сибирский экологический журн. 2011. № 2. С. 189–201.
Поздняков Л.К. Лес на вечной мерзлоте. Новосибирск: Наука, 1983. 96 с.
Прокушкин С.Г. Структурно-функциональные особенности лиственницы Гмелина в криолитозоне Центральной Эвенкии. Красноярск: Изд-во Института леса СО РАН “ДарМа-печать”, 2008. 161 с.
Симанько В.В, Бенькова А.В., Шашкин А.В. Применение метода “скользящих функций отклика” для выявления влияния климатических факторов на радиальный рост деревьев // Вестник КрасГАУ. 2013. № 7. С. 188–194.
Шиятов С.Г. Дендрохронология верхней границы леса на Урале. М.: Наука, 1986. 136 с.
Шиятов С.Г., Ваганов Е.А., Кирдянов А.В., Круглов В.Б., Мазепа В.С., Наурзбаев М.М., Хантемиров Р.М. Методы дендрохронологии. Часть I. Основы дендрохронологии. Сбор и получение древесно-кольцевой информации. Красноярск: КрасГУ, 2000. 80 с.
Cook E. Methods of Dendrochronology: Applications in the Environmental Sciences (Eds. Cook E.R., Kairiukstis L.A.). IIASA. Dordrecht; Boston, London: Kluwer Academic Press., 1990. 394 p.
Fritts H.C. Tree-rings and climate. London; N.-Y., San-Francisco: Academic Press, 1976. 576 p.
Kajimoto T. Root System Development of Larch Trees Growing on Siberian Permafrost / Permafrost ecosystem: Siberian Larch Forest, Ecological studies (eds. Osawa A., Zyryanova O.A., Matsuura Y., Kajimoto T., Wein R.W). Dordrecht: Springer, 2010. Vol. 209. P. 303–330.
Koike T., Mori S., Zyryanova O.A., Kajimoto T., Matsuura Y. Abaimov, A.P. Photosynthetic characteristics of trees and shrubs growing on the north and south facing slopes in Central Siberia / Permafrost ecosystem: Siberian Larch Forest, Ecological studies (eds. Osawa A., Zyryanova O.A., Matsuura Y., Kajimoto T., Wein R. W). Dordrecht: Springer, 2010. V. 209. P. 273–287.
Kramer P.J., Boyer J.S. Water Relations of Plants and Soils. San Diego: Academic Press, 1995. 495 p.
Larson P.R. The indirect effect of drought on tracheid diameter in red pine // Forest Science. 1963. № 9. P. 52–62.
Lockhart J.A. An analysis of irreversible plant cell elongation // J. Theoretical Biology.1965. № 8. P. 264–275.
“National Weather Service. Internet Weather Source”” URL: ftp:/ftp.ncdc.noaa.gov/pub/data/ghcn/daily (дата обращения 15.06.2017).
Nonami H., Boyer J.S. Primary events regulating stem growth at low water potentials. // Plant Physiology. 1990. № 93. P. 1601–1609.
Schweingruber F.H. Tree Rings and Environment. Dendroecology. Bern; Stuttgart; Vienna: Haupt Publ. 1996. 609 p.
Yanagihara Y., Koike T., Matsuura Y., Mori S., Shibata H., Satoh F., Masuyagina O.V. Zyryanova O.A., Prokushkin A.S., Prokushkin S.G., Abaimov A.P. Soil respiration on the contrasting north and south facing slopes of a Larch forest in Central Siberia // Eurasian J. Forest Research 2000. № 1. P. 19–29.
Yasue K., Kujansuu J., Kajimoto T., Nakai Y., Koike T., Abaimov A.P., Matsuura Y. Seasonal Changes in Stem Radial Growth of Larix gmelinii in Central Siberia in Relation to its Climatic Responses // Permafrost ecosystem: Siberian Larch Forest, Ecological studies (eds. Osawa A., Zyryanova O.A., Matsuura Y., Kajimoto T., Wein R.W). Dordrecht: Springer, 2010. V. 209. P. 332–344.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение