

Лесоведение, 2020, № 6, стр. 560-574
Изменение биологической активности почв в хвойных насаждениях после пожара в средней тайге Карелии
М. В. Медведева a, *, О. Н. Бахмет b, В. А. Ананьев a, С. А. Мошников a, А. В. Мамай a, Е. В. Мошкина a, В. В. Тимофеева a
a Федеральный исследовательский центр “Карельский научный центр Российской академии наук” Институт леса КарНЦ РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, 11, Россия
b Федеральный исследовательский центр “Карельский научный центр Российской академии наук”
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: mariamed@mail.ru
Поступила в редакцию 24.04.2018
После доработки 14.06.2018
Принята к публикации 06.06.2020
Аннотация
Впервые в пирогенно трансформированных лесных экосистемах среднетаежной подзоны Карелии дана комплексная оценка состояния микробного сообщества почв. Установлено, что ель более чувствительна к огневому воздействию, чем сосна. На основании данных о проективном покрытии растений напочвенного покрова выделили импактную зону пирогенного воздействия (100% гибель растений) и буферную (до 50%). Контролем служил ненарушенный участок. Изменение фракционного состава лесных подстилок трансформированных участков подтверждает их гетерогенность, нарушение, а также зонирование территории. Выявлено, что под влиянием пожара происходят изменения морфологического строения почв, кислотно-щелочных свойств, содержания зольных элементов и гумуса. Показано, что на фоне пирогенеза происходит изменение структурно-функциональной организации микробоценоза почв. Выявлено изменение численности микроорганизмов, осуществляющих круговорот азота и углерода. Установлено увеличение численности актиномицетов в почвах импактного пирогенного воздействия. Снижение активности каталазы в верхнем органогенном горизонте почв, подверженных наибольшему пирогенному воздействию, свидетельствует об изменении окислительно-восстановительных условий превращения органического вещества. Показано, что содержание углерода микробной биомассы и эмиссия СО2 зависит от свойств почв, зоны пирогенного воздействия. Под влиянием пирогенеза происходит нарушение ресурса органического вещества и его состава, связанное с изменением фракционного состава лесных подстилок, что указывает на изменение условий функционирования микробиоты. Снижение деструкционных процессов на участках импактной зоны определяется невысокой потребностью микроорганизмов в субстрате, а также неблагоприятными эдафическими условиями. В настоящее время возможно формирование отрицательного углеродного баланса, когда потери углекислоты не будут компенсироваться поступлением растительных остатков в экосистему. Данные проведенных исследований могут быть использованы при проведении экологического мониторинга природной среды, а также могут быть основой при создании концептуально-балансовых моделей эмиссии СО2 на территории России.
Лесные пожары оказывают всеобъемлющее и длительное воздействие на различные компоненты экосистемы (Ollinger, 2002; Горшков, 2009; Chapin et all, 2010; Дымов и др., 2015). Ежегодно в России происходят десятки тысяч пожаров, а их площадь составляет миллионы гектаров. Только за 2015 г. их количество составило более 11 тысяч, а охваченная огнем площадь – более 2 млн. га. В республике Карелия в 2015 г. зафиксировано 73 лесных пожара, при этом общая площадь, охваченная огнем, составила более 83 тыс. га. В ходе прямого огневого контакта повреждается или уничтожается значительная часть основных компонентов биогеоценоза: фитоценоз – от мохово-лишайниковой растительности до древостоя, почва, биота. Изменение состава и количества трофического ресурса на фоне пирогенного воздействия может способствовать смене разнообразия биотипов, составляющих это сообщество. Так, гибель или ослабление древесных растений зачастую вызывает резкий рост численности насекомых-ксилофагов, например, короеда типографа в ельниках (Маслов, 2010). М.А. Шешукова с соавт. (1992) отмечают, что пожары вызывают коренные изменения в процессах естественного лесовозобновления, роста и развития древесных пород. Велика роль лесных экосистем в поддержании баланса углерода на планете (Bolin, 1977; Горшков и др., 1995). Баланс органического углерода антропогенно нарушенной лесной экосистемы отмечается как при краткосрочных воздействиях – в результате выброса СО2 в процессе горения, так и длительных – в выделении его при разложении древесины погибших деревьев. Именно пожары, по мнению Д.Г. Замолодчикова с соавт. (2013), являются основной причиной межгодовых вариаций углеродного баланса лесов России.
Работы по изучению влияния пожаров на разные уровни организации наземных экосистем проводятся достаточно давно. Основное внимание уделялось основному биотическому компоненту – древостою (дендроценозу). Как и древесный ярус, важным составным компонентом бореальных лесов является напочвенный покров, основная роль которого – регуляция гидротермического режима верхних горизонтов почвы и стабилизация условий для распада органического вещества и поглощения минеральных веществ корнями растений (Горшков, Ставрова, 2009; Ковалева, Иванова, 2012). В результате большинства пожаров травяно-кустарничковый и мохово-лишайниковый покровы уничтожаются практически полностью, а также выгорает значительная часть лесной подстилки (Горшков, Ставрова, 2009; Динамика лесных сообществ …, 2009; Тарасова и др., 2011).
Процессы восстановления напочвенного покрова в хвойных лесах бореальной зоны начинаются сразу поле пожара. На первом этапе (0–50 лет) происходит интенсивный рост травяно-кустарничкового яруса, в покрове доминируют травянистые растения, разрастаются ксерофитные виды мхов. Через 50–100 лет постепенно стабилизируются параметры мохово-лишайникового покрова и снижается доля травянистых видов растений. Полная стабилизация напочвенного покрова нижних ярусов еловых и сосновых лесов наблюдается через 120–200 лет после пожара (Баккал и др., 2009; Горшков, Ставрова, 2009; Ярмишко, 2009; Gleixner et al., 2001; Lindholm, Vasander, 1987).
Известно, что почва как часть лесных экосистем испытывает сильное влияние пирогенного воздействия (Богданов и др., 2009; Дымов и др., 2015). При этом отрицательное влияние этого воздействия на свойства лесных почв обусловлено природными предпосылками, в том числе самими эдафическими условиями, которые определяют пирологический режим всей таежной зоны. В Карелии доминируют почвы, в морфологическом облике которых выделяется верхний органогенный горизонт – лесная подстилка (Морозова, 1991). Пересыхая в летнее время года, она может создавать предпосылки для возникновения пожароопасной ситуации, последствия которой отмечаются даже спустя 200 лет после пожара. Резкая смена растительности, подверженной влиянию пирогенного стресса, нарушает морфологический профиль почв, изменяет направленность трансформации органического вещества, в целом ход почвообразовательного процесса. Минерализация поверхности почвы в результате пожаров существенно изменяет условия развития для микробиоты (Vazquez et al., 1993; Безкоровайная и др., 2005; Богородская, Кукавская, 2016). При этом резкие изменения светового, питательного и водного режимов почвы также могут способствовать смене видового состава микроорганизмов.
Вопросы прямого воздействия и последствий пожара на почвы и происходящие в ней процессы остаются малоизученными. Хотя именно эти компоненты во многом предопределяют пути, сроки и восстановления всей экосистемы, формирования устойчивости древостоя к неблагоприятным факторам среды (Мухортова и др., 2015). В этой связи необходимо проведение комплексных исследований абиотических и биотических факторов, которые определяют сукцессионную направленность постпирогенного развития всего биогеоценоза. Пирогенные трансформации растительности являются результатом интегрального воздействия совокупности факторов: особенностей рельефа, разнообразия и структуры растительного покрова, вида и силы пожара, условий его возникновения. В связи с тем, что влияние пирогенного фактора на свойства почв зависит от интенсивности и давности пожара, а также от типа древостоя, в котором идет формирование почв, целью настоящего исследования послужило изучение изменения свойств почв при пирогенной сукцессии хвойного древостоя в условиях средней тайги.
ОБЪЕКТЫ И МЕТОДИКА
Национальный парк (НП) “Водлозерский” (площадь 472 тыс. га) расположен на территории Пудожского района республики Карелия и Онежского района Архангельской области (рис. 1). Выбор данного места для исследования постпирогенных преобразований природной среды не был случайным. Он обусловлен тем, что важной особенностью территории НП является удаленность от крупных населенных пунктов (расстояние до г. Петрозаводск составляет более 140 км), промышленных предприятий (Кондопожский ЦБК – 135 км, Сегежский ЦБК – 212 км, Архангельский ЦБК – 312 км), иных потенциальных источников аэротехногенного загрязнения (космодром Плесецк – 184 км), расстояние до ближайшей автодороги федерального значения А-119 “Вологда–Медвежьегорск” более 50 км. Другой особенностью НП является сохранившийся массив коренных таежных лесов крупнейший в Европе. Таким образом, территория НП “Водлозерский” является одним из немногочисленных естественных природных “островков”, почти не затронутых антропогенной деятельностью, не только Северо-запада России, но и Европы в целом.
По ландшафтному районированию национальный парк находится в пределах равнинного, нормально и кратковременно избыточно увлажненного, холмистого и грядово-волнистого типа ландшафта. Абсолютные высоты территории парка в среднем составляют 100–200 м над уровнем моря.
Климатические условия района исследований характеризуются умеренным вегетационным периодом (безморозный период длится до 120 дней), средней температурой воздуха (число дней с температурой воздуха выше 10°С – 100–110 дней), вследствие умеренного выпадения осадков (650–725 мм в год) высокой влажность воздуха. По условиям теплообеспеченности почв район является умеренно теплым – температура почв в июле около 16.6–17.5°С.
Территория парка имеет достаточно однородное геологическое строение. Коренные породы (кристаллическое основание) представлены нерасчлененными, преимущественно гранитоидными породами и мигматитами. Четвертичные отложения мощные, озерно-ледниковые, ледниковые, флювиогляциальные песчаные, супесчаные, реже суглинистые. Территория расположена на Карельской границе главных стадий поздневалдайского оледенения.
Необходимо отметить, что сочетание рельефа, ландшафта, четвертичных отложений, а также наличие озер (о. Водлозеро, о. Онежское) и их водосборов определили специфику водно-климатического режима. Так, сток в период весеннего половодья здесь составляет более 220 мм, для сравнения – на севере Карелии он менее 100 мм.
Елово-сосновые сообщества, произрастающие на территории НП, неоднократно подвергались воздействию природных факторов. Среди них можно выделить ветровальное воздействие, транзитом прошедшее через всю территорию парка и изменившее его ландшафт. Также сильное влияние на коренные леса оказывают пожары разной интенсивности. Крупный пожар, который произошел более 3 лет назад, стал объектом пристального внимания ученых.
На территории НП была заложена постоянная пробная площадь (62°16.021 с.ш., 36°41.913 в.д.). Она представляет собой суходольный участок, расположенный среди большого болотного массива. Елово-сосновый древостой, подвергшийся природному пирогенному воздействию в 2013 г., в настоящий момент находится на начальной стадии восстановительной послепожарной сукцессии. Пожар, охвативший данную территорию, оценивается как низовой, средней интенсивности. На данном участке наблюдаются обгоревшие корни и стволы сосны, усыхающие ели. Официальной причиной возгорания данного древотстоя считается сухая гроза, в пользу этого свидетельствует удаленность участка от населенных пунктов, дорог и основных туристических маршрутов.
При осуществлении лесоводственной части исследования применяли стандартную методику (ОСТ 56-69-83). Проводили учет основных таксационных показателей и их изменения в результате пожара. Возрастную структуру древостоя определяли путем отбора кернов у 15–20 деревьев буравом Пресслера. Сплошной перечет деревьев с разделением на растущие и погибшие с указанием причины позволил установить структуру древостоя. Оценку естественного возобновления производили с учетом степени его жизнеспособности по категориям крупности подроста (<0.25, 0.25–0.5, 0.5–1.0, 1.0–1.5, 1.5–2.0, >2 м). В ходе камеральной обработки были рассчитаны основные таксационные показатели насаждения (состав, полнота, запас, класс бонитета и т.д., данные по естественному возобновлению), определены потери (по количеству деревьев, запасу и т.д.) в результате пожара. Таксационная характеристика древостоя до и спустя год после пожара представлена в табл. 1. Описание нижних ярусов лесных фитоценозов проводилось при помощи стандартных методик проведения геоботанических исследований (Полевая геоботаника 1964, 1976; Сукачев, Зонн, 1961).
Таблица 1.
Лесоводственно-таксационная характеристика естественного и пирогенно нарушенного древостоя
| Ярус | Состав древостоя | Возраст, лет | Число стволов, шт. га–1 |
Средние | Полнота | Запас растущий (наличный), м3 га–1 | ||
|---|---|---|---|---|---|---|---|---|
| диаметр, см | высота, м |
абсолютная, м2 га–1 | относи- тельная |
|||||
| До пожара | ||||||||
| 1 | 10С | 210 | 132 | 30.0 | 18.4 | 9.3 | 0.28 | 81 |
| 2 | 10Е | 90 | 1698 | 12.3 | 11.3 | 20.1 | 0.85 | 128 |
| Всего | 1830 | 209 | ||||||
| После пожара | ||||||||
| 1 | 10С | 210 | 132 | 30.0 | 18.4 | 9.3 | 0.28 | 81 |
| 2 | 10Е | 90 | 1032 | 12.7 | 11.6 | 13.0 | 0.52 | 82 |
| Всего | 1164 | 163 | ||||||
В связи с большой гетерогенностью экологических условий и контрастностью свойств педосреды, на данной пробной площади были выделены однородные по микрорельефу участки площадью 60 м2, на которых и проводили исследование свойств почв. На основании степени обгорания подстилки и напочвенного покрова выделяли зоны интенсивного пожара (импактную зону пирогенного воздействия) и среднюю (буферную зону пирогенного воздействия). Территория с ненарушенным растительным покровом служила контролем. Почвенные разрезы были заложены в разных зонах пирогенного воздействия и на контрольном участке.
Разрез 1 заложен в импактной зоне пирогенного воздействия. Наблюдения выявили 100%-е выгорание лесной подстилки, при этом черный налет “сплошным ковром” покрывал изучаемую область. Вечнозеленые кустарнички черники единично произрастали на участке, состояние их можно было оценить как удовлетворительное, что являлось хорошей предпосылкой к восстановлению напочвенного покрова. Почвенный покров был сильно нарушен, турбирование органогенного и верхнего минерального горизонтов почв выявлено повсеместно. Почва, сформировавшаяся в данной зоне, – антропогенно нарушенный подзол иллювиально-железисто-гумусовый (Podzols) имеет следующее морфологическое строение: Оpir(0–1 см)–Еpir(1–3 см)–ВFHpir(3–12 cм)–В2(12–23 см)–ВС(23–48 см)–С(ниже 48 см).
Разрез 2 заложен в буферной зоне пирогенного воздействия. Степень обгорания подстилки составляла менее 35%. Несмотря на наличие областей полного выгорания лесной подстилки, напочвенный растительный покров находится в удовлетворительном состоянии, поэтому может быстро адаптироваться к новым педоусловиям. Почва экотона – подзол иллювиально-гумусовый супесчаный на водно-ледниковых отложениях (Podzols). Почва имеет следующее морфологическое строение: Оpir(0–3 см)–Еpir(3–7 см)–В-Hpir(7–17 cм)–В2(17–29 см)–ВС(29–53 см)–С(ниже 53 см).
Разрез 3, заложенный на ненарушенном участке, рассматривался в качестве контроля. Однако надо подчеркнуть, что вследствие поступления пыли, сажи, семян растений, резкого изменения гидротермического режима близлежащей территории, а также изменение поступления опада (например, отпада коры хвойных деревьев) область охвата пирогенеза не ограничивается исключительно импактной зоной воздействия. Происходит постепенное изменение всех уровней организации биогеоценоза, прилегающего к зоне максимального природного воздействия. В этой связи участок “контроль” можно назвать относительным. Почва – подзол иллювиально-железисто-гумусовый супесчаный (Podzols), имеет следующее морфологическое строение: О(0–5 см)–Е(5–11 см)–ВFH(11–22 cм)–В2(22–36 см)–ВС(36–60 см)–С(ниже 60 см).
Исследовали древесные фитоценозы, которые подверглись влиянию низовых подстилочно-гумусовых пожаров разной давности и интенсивности. Свойства почв и их динамику на фоне пирогенного фактора изучали на постоянных пробных площадях (ПП).
В полевых условиях на каждой пробной площади закладывали почвенные разрезы, для более детального исследования почвенного покрова делались прикопки. Таксономическая принадлежность исследуемых почв устанавливалась в соответствии с региональной классификацией (Морозова, 1991), также приведена корреляция названий исследуемых почв в соответствии с международной реферативной базой почв WRB (World reference base for soil resources 2014, 2015).
Отбор образцов верхнего органогенного горизонта почв проводили при помощи рамки-шаблона, при этом измеряли его мощность. Отобранные образцы лесных подстилок в лаборатории высушивали до абсолютно сухого веса, взвешивали, после чего вычисляли их объемный вес и запас. За основу изучения компонентного состава подстилок были взяты общетеоретические разработки Л.О. Карпачевского (1981). В связи с различной скоростью разложения фракций, а также устойчивости к пирогенному воздействию проводили разделение образцов лесной подстилки ситовым методом на неактивные (>5 мм) и активные (<5 мм) фракции. После взвешивания каждой фракции находили их процентное содержание от массы всего образца.
Для определения химических свойств почв отбирали образцы из почвенных разрезов по генетическим горизонтам. Определяли гранулометрический состав почв методом Качинского, зольность для органогенных горизонтов и потерю при прокаливании для минеральных горизонтов, показатель рН – потенциометрическим методом, гидролитическую кислотность и сумму обменных оснований – по Каппену, содержание органического углерода – методом Тюрина, азота – методом Кьельдаля, по общепринятым в почвоведении методикам (Аринушкина, 1970).
При исследовании микробного сообщества анализировались смешанные образцы, отобранные на участках. Отбор почв для микробиально-биохимических анализов был произведен по генетическим горизонтам на стенке разреза из пяти точек на всю глубину профиля. Численность почвенных гетеротрофных микроорганизмов, которые принимают участие в трансформации азот- и углеродсодержащих соединений, определяли методом посева на плотные элективные питательные среды (Методы почвенной микробиологии, 1991). При этом количество бактерий, использующих органические формы азота, учитывали на мясопептонном агаре (МПА), ассимилирующие минеральный азот – на крахмало-аммиачном агаре (КАА), олигонитрофилы – на среде Эшби, олиготрофные микроорганизмы – на почвенном агаре (ПА). Численность актиномицетов определяли на КАА. Комплекс аэробных целлюлозоразрушающих микроорганизмов изучался на среде Гетчинсона. Микроскопические грибы подсчитывали на среде Чапека со стрептомицином, в качестве ингибитора роста бактерий. Активность каталазы устанавливали газометрически (Круглов, Пароменская, 1966).
В отобранных образцах почв определяли субстрат-индуцированное дыхание (СИД) по скорости начального максимального дыхания микроорганизмов после обогащения почвы глюкозой (Anderson, Domsch 1978, 2010). Углерод микробной биомассы рассчитывали по формуле: Смик (мкг С/г почвы) = (мкл СО2/г почвы/ч) × 40.04 + 0.37. Базальное дыхание (БД) определяли по скорости выделения CO2 почвой за 24 часа ее инкубации при 22°С и 60% полной полевой влагоемкости. Скорость БД выражали в мкг C-CO2/г почвы/ч. Микробный метаболический коэффициент (qCO2) рассчитывали, как отношение скоростей выделения CO2 из необогащенной почвы (БД) и почвы, в которую внесен избыток глюкозы (СИД) (Благодатская, Ананьева, 1996; Ананьева и др., 2002, 2009). Для расчета содержания азота в микробной биомассе (Nмик) использовали соотношение Nмик/Смик = 0.15 (Anderson, Domsch, 1980; Сусьян и др, 2009). Содержание микробного углерода и азота рассчитывали как процент от общего содержания C и N, соответственно.
Данные химических анализов были получены с использованием оборудования ЦКП “Аналитическая лаборатория” Института леса Карельского НЦ РАН. Статистическая обработка данных осуществлялась в среде Microsoft Excel.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Проведенные исследования показали, что различные компоненты биогеоценоза, находящиеся на начальной стадии постпирогенной сукцессии, имели различный адаптационный потенциал.
Древостой. В первом ярусе древостоя, представленном перестойной сосной в возрасте 210 лет, спустя год после пирогенного воздействия изменений отмечено не было. Это возможно обусловлено тем, что сосны отреагировали на пожар снижением прироста, при этом сохранив жизнеспособность. Основные изменения произошли во втором ярусе, сформированном елью. Спустя год погибшие деревья составили 39.2% их общего числа. Запас породы сократился на 35.9%, потери в целом по древостою составили 22%. В наибольшей степени повреждению подверглись деревья в ступенях толщины 4–10 см, где усохло 50%, в низших ступенях (4–6 см) погибли все растения. Основной причиной гибели растений на период исследования являлось непосредственное огневое воздействие. Полученные данные согласуются с результатами других исследователей, отмечавших большую чувствительность ели к высокотемпературному прессу (Баккал и др., 2009; Богданов и др., 2009; Горшков, Ставрова, 2009).
Степень повреждения растений на изучаемом участке была крайне неравномерна и определялась несколькими основными факторами: древесной породой, микрорельефом поверхности, мощностью и влажностью лесной подстилки. Высокая повреждаемость ели при низовых пожарах даже невысокой интенсивности обусловлена породными особенностями, в частности, низко опущенной кроной, тонкой корой ствола и концентрацией значительной части корней в верхней части минерального горизонта почвы и в подстилке. Усилению пирогенного воздействия способствовал слой хвойного опада вокруг стволов деревьев, иногда достигающий 10-ти см. По-видимому, это связано со специфическим строением кроны ели, способствующим накоплению опавшей хвои на относительно небольшом удалении от ствола, а также с замедленным разложением хвои из-за ее химического состава и неблагоприятного теплового режима. Сочетание всех этих факторов в исследуемом насаждении привело к повреждению не только коры и наружных проводящих тканей ствола и корней, но и глубокому (до 4–5 см) прогоранию древесины по всей окружности ствола отдельных деревьев.
Сосновая часть древостоя пострадала в значительно меньшей степени, вероятно не только из-за высоко расположенной кроны и мощной защиты от термического воздействия в виде толстой коры в нижней части ствола, но и меньшей толщины хвойного опада, сосредоточенной у корневой шейки деревьев этой породы.
Подрост. Естественное возобновление на участке на период проведения исследования полностью отсутствовало, весь подрост, сформированный до пожара елью с небольшой примесью сосны, погиб. В то же время пожарная минерализация поверхности почвы и обогащение ее элементами питания создали хорошие условия для укоренения семян и развития всходов древесных растений. Можно предположить, что в течение ближайших трех лет здесь появится подрост березы, несмотря на отсутствие ее в составе древостоя до пожара. Источники обсеменения хвойных, как сосны, так и ели, на участке и вокруг него имеются, однако сроки зависят от близости семенного года. Перспективу участия подроста сосны в составе формирующегося насаждения, с учетом высокого возраста деревьев материнского полога и светолюбивые породы, в дальнейшем можно оценить как невысокую. Ель, как теневыносливая порода, вероятно, примет определенное участие в составе, однако сейчас рассуждать о ее доле сложно. Это будет во многом зависеть от периодичности семенных лет, обильности плодоношения и степени развитости к тому времени травяно-мохового покрова.
Уже на следующий после пожара год на участке появились признаки активизации деятельности насекомых-вредителей, в частности короеда-типографа. Можно предположить, что из-за пирогенного увеличения кормовой базы в течение ближайших 1–3 лет произойдет резкое увеличение численности. В результате следует ожидать дальнейшего усиления изреживания, куда перейдет еще часть ослабленных пожаром деревьев, а впоследствии, возможно, и растения с окружающих, не поврежденных пожаром участков леса. Восстановление древостоя будет происходить замедленно вследствие неблагоприятных погодно-климатических условий, а также низкоплодородных почв.
Растительный покров. В результате пожара 2014 г. в смешанном сосново-еловом лесу до минерального горизонта почвы выгорело более 60% напочвенного покрова и лесной подстилки. По сравнению с незатронутым пожаром однотипным участком леса (контроль) на горельнике проективное покрытие травяно-кустарничкового яруса сократилось в 1.5 раза, мохово-лишайникового – почти в 4 раза (табл. 2). В первый год после пожара среди трав и кустарничков наибольшее покрытие сохранилось у доминантного вида – Vaccinium myrtillus (свыше 23%), при этом резко в виде отдельных “пятен” возросло участие в покрове V. vitis-idaea (максимальное покрытие – 15%). Другим отличием послепожарного участка леса от не горелого является разрастание в травяно-кустарничковом ярусе злака Avenella flexuosa и появление в покрове нескольких компактных популяций Chamaenerion angustifolium (максимальное покрытие на площадке – 15–20%). Лесные мхи сохранились в виде отдельных куртин только на не выгоревших участках выдела. На сильно нарушенных пожаром местах с выгоревшей до 100% лесной подстилкой встречаются только мхи рода Polytrichum и пионерный вид печеночных мхов – Marchantia polymorpha, покрывая от 3 до 60% поверхности почвы на разных участках выдела.
Таблица 2.
Геоботаническое описание естественных и пирогенно нарушенных участков
| Вид | Контроль | Буферная зона | Импактная зона |
|---|---|---|---|
| Кустарниковый ярус (проективное покрытие, %) | |||
| Sorbus aucuparia L. | + | Не обн. | Не обн. |
| Травяно-кустарничковый ярус (проективное покрытие, %) | |||
| Avenella flexuosa (L.) Drej. | <1 | Не обн. | Не обн.. |
| Empetrum hermaphroditum Hagerup | <1 | Не обн. | Не обн. |
| Vaccinium myrtillus L. | 45 | 20 | 10 |
| V. uliginosum L. | Не обн. | Не обн. | 3 |
| V. vitis-idaea L. | 10 | 3 | 1 |
| Общее проективное покрытие видов травяно-кустарничкового яруса, % | 50 | 20 | 10 |
| Мохово-лишайниковый ярус (проективное покрытие, %) | |||
| Pleurozium schreberi (Bird.) Mitt. | Не обн. | 2 | Не обн. |
| Polytrichum cf. junipeinum Hedw. | 8 | не обн. | <1 |
| Ptilium crista-castrensis (Hedw.) De Not. | 35 | Не обн. | Не обн. |
| Общее проективное покрытие мохово-лишайникового яруса, % | 90 | 2 | <1 |
| Общее проективное покрытие живого напочвенного покрова, % | 98 | 20 | 10 |
Морфологические свойства почв. Результаты показали, что изучаемые почвы имеют общие морфологические свойства, которые проявляются в строении минеральной толщи: хорошо развитом иллювиальном горизонте, относительно невысокой каменистости. Нижняя граница иллювиального горизонта легко диагностируется по влажности, цвету, плотности сложения, а также наличию средних и крупных корней. Насыщенность минеральной толщи корнями постепенно убывает по мере продвижения в глубь почвенной толщи. Изменение окраски происходит от ярко-ржавой в верхней части минеральной толщи до серо-коричневых тонов в переходном в минеральную породу горизонте.
Вместе с тем, несмотря на общие морфометрические свойства почв, можно выделить и особенные, обусловленные влиянием пирогенного воздействия. Прежде всего, в органогенном горизонте почв импактной зоны пирогенного воздействия отмечали изменение цвета (потемнение), состава, мощности (резкое снижение) и наличие большого количества углей. При этом подзолистый горизонт также претерпевал заметные изменения, которые выражались в изменении цвета, мощности, появлении многочисленных потеков гумуса, наличии не идентифицируемой границы перехода между органогенным и минеральным горизонтами. В почвах буферной зоны, где интенсивность пожара была менее выраженной, изменения морфометрических показателей были менее заметны и касались в основном только верхнего органогенного горизонта, поэтому восстановление данных участков будет происходить более интенсивно, чем в зоне максимального пирогенеза.
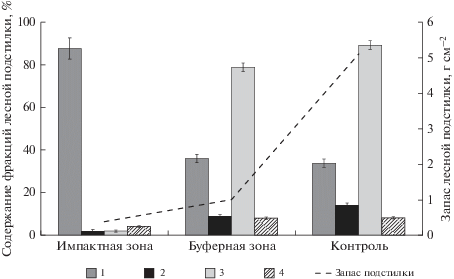
Запас подстилок и содержание компонентов в подстилках на исследуемых участках приводится на рис. 2. Результаты показали, что происходило резкое уменьшение запаса подстилок на участках наибольшее пирогенного воздействия. Также отмечали резкое нарушение в содержании как крупных, так и мелких фракций в составе лесных подстилок. Особенно резко увеличилось содержание крупных фракций, основную долю которых составляли артефакты (уголь), шишки, ветки, кора, хвоя, обгорание которых происходило не до конца. В меньшей степени были представлены мелкие компоненты (частицы размером 0.25–7 мм), выгорание которых происходило быстрее, а, следовательно, доступность их для микробиологической трансформации возрастала.
Рис. 2.
Изменение состава лесных подстилок на фоне пирогенного воздействия (размеры фракций: 1– более 7 мм; 2 – 7–5 мм; 3 – от 5 до 0.25; 4 – менее 0.25 мм).
Гранулометрический состав и физико-химические свойства почв. Почвы по гранулометрическому составу относятся к легким супесчаным, поэтому реабилитация их после снятия антропогенного влияния, в данном случае пирогенного, будет происходить более интенсивно по сравнению с почвами тяжелого механического состава. Это во многом обусловлено тем, что вследствие фронтального промачивания почв легкого механического состава восстановление их морфологического облика будет происходить быстро, а значит и интенсивнее будет восстановление структурно-функциональной организации микробного сообщества, осуществляющего круговорот элементов минерального питания растений. Последнее будет определяющим при формировании продукционной способности древостоя, а, значит, и устойчивости его к неблагоприятным факторам среды.
Как отмечено выше, изучаемые почвы относятся к кислым, что является следствием поступления хвойно-листового опада с низким значением рН, недостаточно высоким содержанием щелочноземельных металлов в почвообразующей породе, представленной озерно-ледниковыми отложениями, заторможенностью процессов выветривания первичных минералов (табл. 3). Изменение кислотности закономерно уменьшается по мере продвижения в глубь почвенной толщи. По мере приближения к центру пирогенного воздействия происходило возрастание зольности и одновременно снижение кислотности верхнего органогенного горизонта почв, что согласуется с результатами других исследователей (Безкоровайная и др., 2005; Динамика лесных сообществ, 2009; Дымов, 2015). Изменение суммы и степени насыщенности почв основаниями в почвах пирогенно нарушенных участков свидетельствует об изменении почвенно-поглощающего комплекса, его микростроения, что может оказывать влияние на микробиологические свойства почв. Возможно, что биопленки, формирующиеся на поверхности почвенных частиц и являющиеся местом жизни микроорганизмов, разрушаются под действием высоких температур, поэтому происходит деформация внутренней организации микробного сообщества. Нарушения структуры почвенно-поглощающего комплекса диагностируются снижением степени насыщенности и суммы поглощенных оснований. Это в целом свидетельствует об изменении микролокусности почв, нарушении среды обитания для микроорганизмов.
Таблица 3.
Физико-химические свойства почв, подверженных воздействию низового пожара
| Горизонт почв | Зольность/потеря при прокаливании, % | рНвод | S* , моль (экв.)/кг | V**,% | С, % | N, % | C/N | Содержание фракций, % | |
|---|---|---|---|---|---|---|---|---|---|
| <0.001 | <0.01 | ||||||||
| Импактная зона | |||||||||
| Opir | 24.85 | 5.32 | 4.42 | 3.5 | 26.9 | 1.04 | 25.9 | 5.2 | 13.7 |
| Еpir | 9.51 | 4.74 | 0.00 | 0.0 | 7.2 | 0.17 | 42.4 | 1.6 | 7.7 |
| BFHpir | 5.99 | 5.98 | 0.00 | 0.0 | 2.7 | 0.12 | 22.5 | 2.4 | 6.8 |
| Буферная зона | |||||||||
| O | 12.41 | 4.87 | 33.54 | 28.5 | 30.7 | 1.16 | 26.5 | 3.2 | 13.3 |
| E | 6.61 | 5.14 | 0.20 | 3.4 | 3.0 | 0.11 | 27.3 | 2.9 | 7.0 |
| BH | 10.58 | 5.09 | 1.45 | 11.4 | 3.6 | 0.16 | 22.5 | 1.6 | 9.7 |
| Контроль | |||||||||
| O | 6.48 | 4.52 | 22.12 | 17.1 | 39.5 | 0.98 | 40.3 | 3.3 | 11.8 |
| E | 5.68 | 4.74 | 27.12 | 0.0 | 2.1 | 0.10 | 21.0 | 2.0 | 8.4 |
| BH | 4.69 | 5.64 | 0.00 | 18.0 | 2.8 | 0.16 | 17.5 | 2.0 | 9.7 |
Заметные изменения исследуемых химических свойств почв происходили на фоне пирогенного воздействия. Прежде всего, выявлено увеличение зольности верхнего органогенного и подподстилочного горизонтов почв импактного участка. Это связано с поступлением большого количества золы в результате 100%-го выгорания подстилки и живого напочвенного покрова. Нейтрализующее действие золы проявлялось в увеличении рНводн верхнего и подподстилочного горизонтов почв. При этом нижние горизонты почв имели кислую реакцию и не отличались от таковых, сформировавшихся под пологом леса.
В почвах, испытывающих наибольшее пирогенное воздействие, выявлено снижение содержания Собщ, которое происходит в результате выгорания подстилки. Как известно, при беглом низовом пожаре средней силы в подстилке на стадии тления при температуре <3000°С происходит образование меньшего количества угля, которое и обусловливает более низкое его содержание на исследованных гарях по сравнению с контролем (Gleixner et al., 2001). Однако содержание валового углерода, азота увеличивалось в подподстилочном горизонте почв, что свидетельствует об изменении направленности почвообразовательного процесса: в почвах ненарушенных экосистем отмечается резкое снижение элементов – биофилов в подзолистом горизонте почв.
В исследуемых почвах пирогенно трансформированных участков отмечено невысокое содержание валового азота, что связано, возможно, с его изначально низким содержанием в исследуемых почвах и пирогенным превращением в летучие соединения (Богданов и др., 2009). Сгорание травяно-мохового покрова, подстилки, почвенного перегноя ведет к единовременному высвобождению из сгорающих органических веществ заключенных в них зольных элементов и улетучиванию в атмосферу углерода, азота. При этом отмечается изменение соотношения указанных элементов в пирогенно трансформированных горизонтах почв, изменение минерализационных процессов по сравнению с контролем.
Таким образом, проведенные исследования показали, что на фоне пирогенного фактора происходят заметные изменения морфологических и химических свойств почв. Степень трансформации почв зависит от интенсивности пожара: наибольшие изменения прослеживаются на участке импактной зоны пирогенного воздействия. Изменение морфометрических и химических свойств почв ухудшает выполнение ими концентрационно-трофических функций, изменяет условия функционирования для микробиоты (Безкоровайная и др., 2005; Сорокин, 2009). Микробиологические исследования показали, что на фоне пирогенного воздействия произошли изменения эколого-трофической структуры микробного сообщества (табл. 4). Прежде всего, установлено уменьшение численности микроорганизмов, утилизирующих органические соединения азота. Это связано во многом со снижением поступления свежего растительного опада, нарушением трофобеспеченности почв. Однако возрастание спорообразующих прокариот в составе аммонификаторов может свидетельствовать о более глубокой минерализации органического вещества в почве возможно вследствие поступления корневого опада. Подтверждением сдвига минерализационных процессов в почве является снижение содержания микроорганизмов, использующих минеральные формы. Однако возрастание численности актиномицетов в их составе, нуждающихся в полном наборе элементов питания, подтверждает более глубокую минерализацию органического вещества, обеспеченность почв элементами минерального питания. Интересно, что подобное увеличение доли актиномицетов в составе микроорганизмов, осуществляющих трансформацию трудногидролизуемых соединений азота, наблюдали в зоне аэротехногенного воздействия Костомукшского ГОКа (Медведева и др., 2008). Возможно, что это обусловлено благоприятным сочетанием увеличениия рН, поступлением элементов питания и изменением теплообеспеченности почв. Последнее связано с потемнением верхних горизонтов в результате выгорания подстилки. Возрастание численности микроорганизмов олиготрофной группы свидетельствует, что на данном этапе развития почв идет интенсивное использование микробиотой собственно органического вещества почв (Ананьева и др., 2002 Безкоровайная и др., 2005; Дымов и др., 2015; Sollins et al., 1996). Можно предположить, что микробиота использует для своих конструктивных и энергетических целей периферические, легко отщепляемые гетероциклические цепочки фульвокислот. Снижение численности микромицетов в почве связано с изменением не только экологических условий, но и изменением структуры и состава микробного сообщества. Последнее хорошо диагностировалось изменением комплекса целлюлозоразрушающих микроорганизмов: появление прокариот усиливало процессы разрушение целлюлозы, создавало предпосылки для увеличения скорости круговорота углерода в почве.
Таблица 4.
Численность микроорганизмов основных эколого-трофических групп в изучаемых почвах, тыс./г почвы
| Горизонт почв | Бактерии, утилизирующие | Олиго- нитро- филы |
Олиго-трофы | Микро-мицеты | КЦМ* | Коэффициент минерализации $\frac{{{\text{KAA}}}}{{{\text{MПА**}}}}$ | Индекс олиготрофности$\frac{{{\text{ПA}}}}{{{\text{MПА**}}}}$ |
||
|---|---|---|---|---|---|---|---|---|---|
| N-NH2 | N-NH4 | ||||||||
| общее число | споровые | общее число | |||||||
| Импактная зона | |||||||||
| Opir | 3250 | 1560 | 1350 | 12 410 | 3900 | 150 | 22 | 0.4 | 1.2 |
| Еpir | 1105 | 264 | 2360 | 1440 | 520 | 56 | 7 | 2.1 | 0.5 |
| BFHpir | 910 | 582 | 1191 | 4100 | 132 | 102 | 4 | 1.3 | 0.1 |
| Буферная зона | |||||||||
| O | 5920 | 2427 | 4144 | 10 100 | 5100 | 340 | 10 | 0.7 | 0.9 |
| E | 620 | 198 | 810 | 1420 | 110 | 112 | 5 | 1.3 | 0.2 |
| BH | 810 | 575 | 1510 | 3050 | 70 | 241 | 1.9 | 0.1 | |
| Контроль | |||||||||
| O | 6210 | 745 | 4968 | 5720 | 1960 | 410 | 9 | 0.8 | 0.3 |
| E | 510 | 71 | 702 | 810 | 57 | 101 | 6 | 1.4 | 0.1 |
| BH | 820 | 508 | 1100 | 1620 | 81 | 250 | 1.3 | 0.1 | |
* КЦМ – комплекс целлюлозоразрушающих микроорганизмов. ** Коэффициент минерализации – отношение численности микроорганизмов, выросших на крахмало-аммиачном агаре (КАА), к численности микроорганизмов, выросших на мясо-пептонном агаре (МПА). *** Индекс олиготрофности – отношение численности микроорганизмов, выросших на почвенном агаре (ПА), к численности микроорганизмов, выросших на мясо-пептонном агаре (МПА).
При анализе микробного сообщества ценную информацию дает пересчет количества микроорганизмов на почвенные горизонты (Методы почвенной микробиологии, 1991). Результаты показали, что, несмотря на более высокие показатели численности микроорганизмов в почвах импактного пирогенного воздействия, их численность в профиле почв снижалась. Это связано с нарушением морфологического облика почв, процессами выгорания подстилки, изменением хода почвообразовательных процессов.
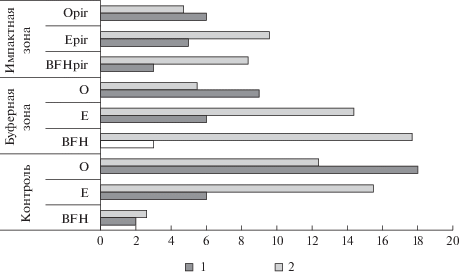
Структурные изменения микробного сообщества оказали влияние и на его функциональную активность. Выявлено снижение активности каталазы в почвах наибольшего пирогенного воздействия (рис. 2). Как известно, каталаза является ферментом, который участвует в расщеплении токсичной для микроорганизмов перекиси водорода, образуемой при расщеплении органических соединений. Снижение активности связано с увеличением содержания минеральной части почвы (золы) в составе лесных подстилок, изменением кислотно-основных свойств почв, а также, возможно, экранированием активных центров фермента продуктами пирогенеза лигнофицированной массой. Медленно высвобождаемые химически инертные молекулы гетерополимеров могут оказывать ингибирующее влияние на работу фермента, в конечном итоге тормозят образование фермент-субстратного комплекса (Sollins et al., 1996).
Рис. 3.
Изменение показателей микробиологической активности почв в результате пирогенеза (1 – С мик г м–2; мл О2 5 мин–1 г почвы–1).
Для объяснения изменения микробиологических основ функционирования лесных сообществ, подверженных влиянию пирогенного фактора, необходимо обратиться к данным таблицы, которая характеризует эко-физиологическое состояние микробиоты и ее изменение на фоне пирогенеза (табл. 5). При анализе процессов микробной трансформации органического вещества в исследованных почвах, следует отметить общую тенденцию их изменений, связанную с комплексным воздействием (Сорокин, 2009).
Таблица 5.
Важнейшие эко-физиологические показатели микробного сообщества изучаемых почв
| Горизонт почв | БД, мкг СО2-С г–1 час–1 | Смик, г С м–2 | qCO2, мкг СО2-С мг Смик–1 ч–1 | Смик/Сорг, % |
|---|---|---|---|---|
| Импактная зона | ||||
| Opir | 19.30 ± 4.47 | 4.79 ± 1.26 | 5.69 ± 0.20 | 1.27 ± 0.34 |
| Еpir | 4.59 ± 0.19 | 9.57 ± 0.43 | 9.70 ± 0.02 | 0.66 ± 0.03 |
| BFHpir | 0.21 ± 0.00 | 8.44 ± 0.02 | 3.91 ± 0.05 | 0.20 ± 0.00 |
| Буферная зона | ||||
| O | 14.05 ± 2.99 | 5.54 ± 0.54 | 4.51 ± 0.53 | 1.00 ± 0.10 |
| E | 3.02 ± 0.31 | 14.42 ± 1.43 | 5.30 ± 0.03 | 1.90 ± 0.19 |
| BH | 0.60 ± 0.05 | 17.65 ± 0.46 | 3.65 ± 0.38 | 0.45 ± 0.01 |
| Контроль | ||||
| O | 15.20 ± 0.70 | 12.35 ± 0.61 | 4.31 ± 0.02 | 0.89 ± 0.04 |
| E | 1.91 ± 0.02 | 15.50 ± 0.34 | 4.75 ± 0.05 | 1.91 ± 0.04 |
| BH | 0.10 ± 0.04 | 2.58 ± 0.21 | 2.89 ± 0.90 | 0.13 ± 0.01 |
Так, базальное дыхание увеличивается в органогенном и минеральном горизонтах почв пирогенно нарушенных экосистем. Такая же тенденция изменения функционирования микробоценоза отмечена для микробного метаболического коэффициента. Увеличение доли микробного углерода в органическом углероде почв свидетельствует о незрелости микробного сообщества, формировании эфимерных трофических связей между отдельными участниками микробиально-биохимических процессов, осуществляющих превращение органического вещества (Harris, 2003). Снижение запасов углерода микробной биомассы, подвергнутых пожару, объясняется уменьшением мощности органогенного и подподстилочного горизонтов, вследствие их выгорания, а также турбулентными нарушениями морфологического строения почв.
ЗАКЛЮЧЕНИЕ
На фоне пирогенного воздействия произошли нарушения на всех уровнях организации экосистемы. Выявлены изменения структуры и состава древостоя, растений напочвенного покрова. Негативное влияние пирогенного воздействия на свойства почв хорошо проявилось в первый послепожарный год. При этом наибольшие изменения установлены для верхнего органогенного горизонта почв, в нижележащих горизонтах почв изменения свойств носят менее выраженный характер. Выявлено изменение естественного морфологического строения профиля почв, реакции среды, содержания элементов-биофилов в верхних горизонтах почв. На фоне пирогенного воздействия произошло изменение фракционного состава подстилок и снижение активности каталазы, свидетельствующие об изменении окислительно-восстановительных условий трансформации органического вещества почв. Анализ микробного сообщества почв, подвергнутых влиянию пожара, привел к выводу о том, что эко-физиологические показатели микробиоты могут быть хорошим биологическим индикатором состояния природной среды. Они характеризуют условия существования микробиоты как со стороны современного их состояния, так и их прогнозного развития. Последнее имеет большое значение в связи с глобальным потеплением климата, а, следовательно, изменением эдафических условий произрастания древостоя в условиях заповедной территории.
Список литературы
Ананьева Н.Д., Благодатская Е.В., Демкина Т.С. Оценка устойчивости микробных комплексов почв к природным и антропогенным воздействиям // Почвоведение. 2002. № 5. С. 580–587.
Ананьева Н.Д., Сусьян Е.А., Рыжова И.М. Бочарникова Е.О., Стольникова Е.В. Углерод микробной биомассы и микробное продуцирование двуокиси углерода дерново-подзолистыми почвами постагрогенных биогеоценозов и коренных ельников южной тайги (Костромская область) // Там же. 2009. № 9. С. 1109–1116.
Аринушкина Е.В. Руководство по химическому анализу почвы. М.: Изд-во МГУ, 1970. 487 с.
Баккал И.Ю., Горшков В.В., Ставрова Н.И. Динамика восстановления основных компонентов бореальных сосновых лесов после пожаров // Проблемы экологии растительных сообществ. СПб.: ООО “ВВМ”, 2009. С. 273–283.
Безкоровайная И.Н., Иванова Г.А., Тарасов П.А., Сорокин Н.Д., Богородская А.В., Иванов В.А., Конард С.Г., Макрае Д.Дж. Пирогенная трансформация почв сосняков средней тайги Красноярского края // Cибирский экологический журн. 2005. № 1. С. 143–152.
Благодатская Е.В., Ананьева Н.Д. Оценка устойчивости микробных сообществ в процессе разложения поллютантов в почве // Почвоведение. 1996. № 11. С. 1341–1346.
Богданов В.В., Прокушкин А.С., Прокушкин С.Г. Влияние низовых пожаров на подвижность органического вещества почвы в лиственничниках криолитозоны Средней Сибири // Вестник КрасГАУ. 2009. Вып. 2. С. 88–93.
Богородская А.В., Кукавская Е.А. Состояние микробных сообществ в почвах лиственных и светлохвойных лесов средней Сибири после рубок и пожаров // Лесоведение. 2016. № 5. С. 383–396.
Горшков В.В., Баккал И.Ю., Ставрова Н.И. Восстановление нижних ярусов сосновых лесов Кольского полуострова после пожаров // Ботанический журн. 1995. Т. 80. № 5. С. 35–46.
Горшков В.В., Ставрова Н.И. Пожары как фактор нарушения бореальных лесов // Проблемы экологии растительных сообществ. СПб.: ООО “ВВМ”, 2009. С. 237–238.
Динамика лесных сообществ северо-запада России // Отв. ред. Ярмишко В.Т. СПб.: ООО ВВМ, 2009. 276 с.
Дымов А.А., Дубровский Ю.А., Габов Д.Н., Жангуров Е.В., Низовцев Н.А. Влияние пожара в северотаежном ельнике на органическое вещество почвы // Лесоведение. 2015. № 1. С. 52–62.
Замолодчиков Д.Г., Грабовский В.И., Шуляк П.П., Честных О.В. Влияние пожаров и заготовок древесины на углеродный баланс лесов России // Лесоведение. 2013. № 5. С. 36–49.
Карпачевский Л.О. Лес и лесные почвы. М.: Лесная промышленность, 1981. 264 с.
Ковалева Н.М., Иванова Г.А. Динамика живого напочвенного покрова после низовых пожаров в сосновых насаждениях (Нижнее Приангарье) // Известия Самарского научного центра Российской академии наук. 2012. № 1–5. Т. 14. С. 1264–1267.
Круглов Ю.В., Пароменская Л.Н. Модификация газометрического метода определения каталазной активности // Почвоведение. 1966. № 1. С. 93–95.
Маслов А.Д. Короед-типограф и усыхание еловых лесов. Пушкино: ВНИИЛМ, 2010. 38 с.
Медведева М.В., Бахмет О.Н., Яковлев А.С. Микробиологическая и биохимическая индикация состояния почв Карелии, подверженных воздействию аэротехногенного загрязнения // Почвоведение. 2006. № 1. С. 72–76.
Методы почвенной микробиологии и биохимии / Под ред. Звягинцева Д.Г. М.: МГУ, 1991. 304с.
Морозова Р.М. Лесные почвы Карелии. Л.: Наука, 1991. 184 с.
Мухортова Л.В., Кривобоков Л.В., Харпухаев Т.М., Найданов Б.Б. Влияние пожаров на запасы корней и подземного детрита в горнотаежных лиственничниках Прибайкалья // Лесоведение. 2015. № 4. С. 282–292.
ОСТ 56-69-83. Площади пробные лесоустроительные. Метод закладки. М.: ЦБНТИ гослесхоза СССР, 1983. 60 с.
Полевая геоботаника. М.;Л.: Наука, 1964. Т. 3. 530 с.; 1976. Т. 5. 320 с.
Сорокин Н.Д. Микробиологический мониторинг нарушенных наземных экосистем Сибири // Известия РАН. Сер. Биол. 2009. № 6. С. 728–733.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Сусьян Е.А., Ананьева Н.Д., Гавриленко Е.Г., Чернова, О.В., Бобровский М.В. Углерод микробной биомассы в профиле лесных почв южной тайги // Почвоведение. 2009. № 10. С. 1233–1240.
Тарасова В.Н., Горшков В.В., Жулай И.А. Динамика восстановления видового разнообразия лишайников в сосняке зеленомошном после низового пожара 1994 г. в заповеднике “Кивач” (южная Карелия) // Отечественная геоботаника: основные вехи и перспективы: Матер. Всерос. конф. СПб.: Изд-во ООО Бостон-спектр. 2011. Т. 2. С. 237–240.
Шешуков М.А., Савченко А.П., Пешков В.В. Лесные пожары и борьба с ними на севере Дальнего Востока. Хабаровск: ДальНИИ лесн. хоз-ва, 1992. 97 с.
Ярмишко В.Т., Ярмишко М.А. Воздействие лесных пожаров на растения нижних ярусов северотаежных сосновых лесов (Кольский полуостров) // Проблемы экологии растительных сообществ. СПб.: ООО ВВМ, 2009. С. 284–293.
Anderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biology & Biochemistry. 1978. V. 10. P. 215–221.
Anderson J.P.E., Domsch K.H. Quantities of plant nutrients in the microbial biomass of selected soils // Soil Science. 1980. V. 130. № 4. P. 211–216.
Anderson T.H., Domsch K.H. Soil microbial biomass: The eco-physiological approach // Soil Biology & Biochemistry. 2010. V. 42. P. 2039–2043.
Bolin B. Changes of land biota and their importance for the carbon cycle // Science. 1977. V. 196. P. 613–615.
Gleixner G., Czimczik C., Kramer C., Luhker B.M. and. Schmidt M.W. I Plant compounds and their turnover and stability as soil organic matter in Global Biogeochemical Cycles in the Climate System London, UK, Academic. 2001. P. 201–213.
Harris J.A. Measurements of the soil microbial community for estimating the success of restoration // European J. Soil Science. 2003. V. 54. P. 801–808.
Lindholm T., Vasander H. 1987. Vegetation and stand development of mesic forest after prescribed burning // Silva Fennica. V. 21(3). P. 259–278.
Ollinger S.V. Forest Ecosystems. Encyclopedia of life science. Basingstoke. Macmillan Publishers: Ltd. Nature Publishing Group. 2002. P. 1–10.
Sollins P., Homann P., Caldwell B.A. Stabilization and destabilization of soil organic matter: mechanisms and controls // Geoderma. 1996. V. 74. P. 65–105.
Vazquez F.J., Acea M.J., Carhallas T. Soil microbial populations after wildfire // Microbiology Ecology. 1993. V. 13. P. 93–104.
World reference base for soil resources 2014. International soil classification system for naming soils and creating legends for soil maps. World soil resources reports/ IUSS Working Group WRB. Rome: FAO. 2015. 203 p.
Дополнительные материалы отсутствуют.