

Лесоведение, 2021, № 1, стр. 78-92
Биологическая активность почв в поясе широколиственных лесов Центрального Кавказа
О. Н. Горобцова a, *, Т. С. Улигова a, Ф. В. Гедгафова a, Р. Х. Темботов a, Е. М. Хакунова a
a Институт экологии горных территорий им. А.К. Темботова РАН
360000 Кабардино-Балкарская Республика, Нальчик, ул. И. Арманд, 37-а, Россия
* E-mail: ecology_lab@mail.ru
Поступила в редакцию 07.06.2018
После доработки 06.12.2018
Принята к публикации 07.10.2020
Аннотация
Биологические показатели характеризуют состояние почвенной системы, ее способность к выполнению экологических функций. Их определение особенно актуально в условиях горных ландшафтов, где возрастающее антропогенное воздействие (вырубка леса, повышение рекреационной нагрузки) приводит к быстрому, а иногда необратимому изменению почвенных свойств и нарушению стабильности лесного биогеоценоза. Основная цель работы состояла в определении параметров и общего уровня биологической активности основных типов горных почв – бурые лесные глеевые, серые лесные, дерново-карбонатные, формирующие почвенный покров пояса широколиственных лесов терского варианта поясности Центрального Кавказа (в границах Кабардино-Балкарии). Проведен анализ профильного распределения биотических показателей, а также определены их средние значения в верхних горизонтах (0–20 см) изученных горных почв. Общий уровень биологической активности охарактеризован на основе интегрального показателя эколого-биологического состояния почв (ИПЭБСП, %), суммирующего изученные биологические характеристики. Показано, что максимальный уровень профильной биологической активности наблюдается в горных дерново-карбонатных почвах, который на 59–41% превышает аналогичный показатель остальных типов. Биологическая активность верхних горизонтов, определенная по ИПЭБСП (отн. %) убывает в следующем ряду: горные дерново-карбонатные (100%) > горные бурые лесные глеевые (64%) > горные серые лесные (55%). Проведенные исследования показывают, что формирование общего уровня биологической активности определяется характером напочвенного растительного покрова и спецификой почвообразовательных процессов, образующих тот или иной почвенный тип.
Высотная поясность является мощным механизмом формирования биологического разнообразия горных территорий, оказывающим влияние на специфику биотического и почвенного покрова как равнин, так и гор Кавказа (Соколов, Темботов, 1989). Восточная часть Центрального Кавказа (бассейн р. Черек) относится к терскому варианту поясности, в котором пояс широколиственных лесов занимает высотные пределы 500–1700 м над ур. моря. Терский вариант граничит с эльбрусским по линии Дыхтау–Каракая – нижнее течение р. Баксан. Более сухой и континентальный климат эльбрусского варианта обусловливает ксерофитизацию практически всех ландшафтов и является причиной отсутствия пояса широколиственных лесов в его поясном спектре. В условиях сравнительно более мягкого и влажного климата терского варианта этот пояс хорошо выражен и имеет четкие границы. Широколиственные леса распространены в терском варианте поясности на северных склонах Скалистого и Лесистого хребтов, в бассейнах рек Черек и Нальчик. С учетом трехмерности рельефа они занимают площадь 283 990 га. Покрытая лесом территория, на которой произрастает 15 типов буковых, грабовых и дубовых лесов, охватывает 158 300 га (Темботова и др., 2012). Определение современного состояния ненарушенных естественных почв пояса широколиственных лесов необходимо для дальнейшего контроля степени их изменения, так как стабильность почвенных свойств обуславливает и устойчивость всего лесного биогеоценоза. Возрастание антропогенной нагрузки в условиях горных ландшафтов приводит к быстрой, а иногда необратимой деградации почвенного покрова, что в свою очередь вызывает потерю ценнейших лесных угодий. Контроль биологических параметров естественных лесных почв позволяет определить стабильность функционирования почвенной системы, фиксировать развитие негативных процессов на самых ранних стадиях (Стольникова и др., 2011).
Данные, характеризующие различные показатели и общий уровень биологической активности (БА) естественных лесных почв, являются основой дальнейших исследований по определению степени трансформации лесных почв при различных видах антропогенного воздействия (сельскохозяйственного использования, рекреационных нагрузок, вырубки лесов и т.д.). Эффективность определения показателей БА отражена в исследованиях, посвященных изучению лесных почв Западного Кавказа (Казеев и др., 2002, 2013; Даденко и др., 2016). Биологические свойства почв пояса широколиственных лесов Центрального Кавказа в настоящее время изучены недостаточно.
Цель данного исследования: оценить параметры и общий уровень БА основных типов естественных почв, формирующих почвенный покров пояса широколиственных лесов терского варианта поясности северного макросклона Центрального Кавказа (в границах Кабардино-Балкарии).
Определение общего уровня БА основных типов почв пояса широколиственных лесов основано на методологических подходах, использующих сравнительно-географический и профильно-генетический методы исследования (Вальков и др., 1999). Так как БА есть производное от целого ряда биотических почвенных показателей, определяющим является выбор небольшого количества характеристик, которые охватывают основные аспекты, формирующие тот или иной ее уровень. К такому набору предлагается отнести: содержание углерода органического вещества; респираторные показатели – скорость базального и субстрат-индуцированного дыхания (БД и СИД), а также содержание углерода микробной биомассы (Смик), определенное методом СИД; активность ряда почвенных ферментов, отражающую биохимические процессы функционирования почвенной системы.
Для того чтобы установить общий уровень БА почв, применяется интегральный показатель эколого-биологического состояния почв (ИПЭБСП). Выраженный в относительных процентах, ИПЭБСП является суммирующим производным комплекса контролируемых параметров БА (Вальков и др., 1999; Казеев и др., 2004).
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили на бурых лесных глеевых почвах по (Классификация …, 1977), бурозем темный по (Классификация …, 2004), Endogleic Cambisols по (World Reference …, 2014); серых лесных почвах по (Классификация …, 1977; Классификация …, 2004), Greyic Phaeozems Albic по (World Reference …, 2014); дерново-карбонатных почвах по (Классификация …, 1977), Карболитозем темногумусовый (рендзина) по (Классификация …, 2004), Rendzic Leptosols Eutric по (World Reference …, 2014).
Бурые лесные глеевые и серые лесные почвы сформировались под буковыми и буково-грабовыми лесами. Буковые леса, занимающие в районе исследования площадь 72300 га, наилучшего развития достигают на высоте 800–1300 м над ур. моря. В их составе преобладает бук восточный (Fagus orientalis Lipsky), спутником которого обычно являются граб кавказский (Сarpinus caucasica Grossh.), а также клен (Aser sp.), липа (Tilia caucasica), вяз горный (Ulmus glabra). Основными типами букового леса являются мертвопокровный, ясменниково-папоротниковый и др. (Темботова и др., 2012).
Наиболее распространенными типами буково-грабовых лесов являются азалиевые, ясменниковые, бузинные, разнотравные, мертвопокровные. Среди грабовых лесов чаще всего встречаются мертвопокровные, папоротниковые, орешниково-щитовниковые. В составе подлеска распространены лещина обыкновенная (Corylus avellana L.), бересклет (Euonymus europaeus L.), бузина (Sambucus nigra L.). В местах с хорошо развитым подростом бука и граба и подлеском из кустарников травяной покров отсутствует. В широколиственных лесах общее проективное покрытие (ОПП) напочвенного травяного покрова может составлять 50–60%. В его составе обычны сныть (Aegopodium podаgrarai L.), подмаренник (Galium aparine L.), медуница (Pulmonaria officinalis L.), ясменник (Asperula graveolens L.), а также злаки: овсяница (Festuca pratensis L.), мятлик (Poa annua L.) и др.
Горные дерново-карбонатные почвы исследуемых территорий функционируют на безлесных склонах, покрытых богатой разнотравно-злаковой растительностью, образующей мощную дернину. Высота травостоя достигает 50–60 см, ОПП – 100%. Наиболее распространены следующие виды злаковых: ежа сборная (Dactylis glomerata), овсец опушечный (Helictotricho pubescens), тонконог Люерсена (Koeleria luerssenii), пырей средний (Elytrigia intermedia), мятлик луговой (Poa pratensis), костер береговой (Bromopsis riparia), тимофеевка луговая (Phleum pratense), трясунка южная (Briza australis), овсяница красная (Festuca rubra) и др. Разнотравье представлено: клевером луговым (Trifolium pratense), лядвенцем кавказским (Lotus caucasicus), подмаренником обыкновенным (Galium verum),душицей обыкновенной (Origanum vulgare), шалфеем мутовчатым (Salvia verticillata), чабрецом пастбищным (Thymus pastoralis), подорожником средним (Plantago media), манжеткой жесткой (Alchemilla rigida), колокольчиком Гогенакера (Campanula hohenackeri), вероникой колосистой (Veronica spicata), льном жилковатым (Linum nervosum), эспарцетом невооруженным (Onobrychis inermis) и др.
Изучение почвенного покрова пояса широколиственных лесов осуществляли в процессе экспедиционных выездов в первой декаде июля 2017 г. Территория исследования (160 тыс. га) расположена на высоте 507–1048 м над ур. моря в границах координат 43°14′–43°30′ с. ш., 43°55′–43°89′ в. д. (рис. 1).
Рис. 1.
Район исследований – пояс широколиственных лесов терского варианта поясности Центрального Кавказа (в границах Кабардино-Балкарии).
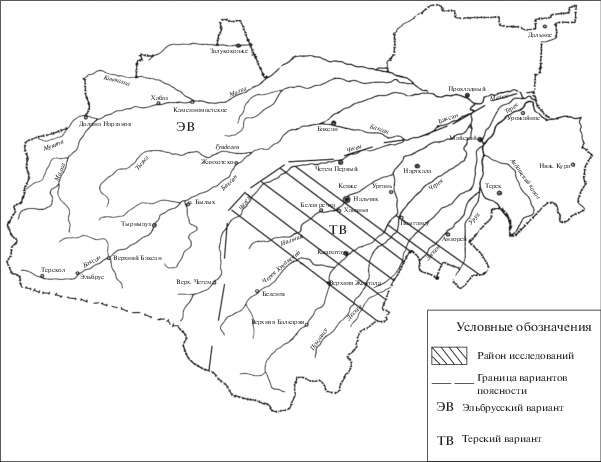
При определении мест отбора для закладки разрезов использовали почвенную карту исследуемых территорий масштаб 1 : 100 000 (Молчанов и др., 1990). Закладку почвенных разрезов и отбор образцов из генетических горизонтов горных почв проводили на наиболее типичных участках естественных биогеоценозов. Диагностику исследуемых почв осуществляли в соответствии с генетической классификацией почв (Классификация …, 1977). Ниже представлено морфогенетическое описание четырех полнопрофильных разрезов исследуемых типов почв, обладающих наиболее характерными свойствами. Места заложения разрезов указаны в табл. 1.
Таблица 1.
Местоположение разрезов, характеризующих основные типы почв широколиственных лесов терского варианта поясности Центрального Кавказа (в границах Кабардино-Балкарии)
| № | Географическое положение разреза | Название почвы (Классификация …, 1977) | Растительное сообщество |
|---|---|---|---|
| 1 | Окрестности села Белая Речка 43°42′405″ с.ш., 43°46′690″ в.д., 778 м над ур. моря |
Горная бурая лесная глеевая, среднегумусная, слабокаменистая, тяжелосуглинистая, на желто-бурых глинах | Буково-грабово-ясменниковый лес, средняя высота стволов 17–19 м, сомкнутость крон 0.7, ОПП 30%, в травостое ясменник душистый, земляника лесная, герань Роберта, кислица обыкновенная, гравилат приречный, любка двулистная, фиалка душистая |
| 2 | За селом Ерокко 43°31′955″ с.ш., 43°88′658″ в.д. 543 м над ур. моря |
Горная серая лесная, среднемощная, среднесуглинистая, малокаменистая, на бурых глинах с включениями щебня, гальки и крупных валунов | Грабово-буково-разнотравный лес, опушка, помимо основных пород дикая груша, дикая алыча, шиповник, боярышник, злаковое разнотравье ОПП 80%, высота травостоя 30 см, в травостое злаковые, бобовые, тысячелистник, душица обыкновенная, шалфей мутовчатый, скабиоза белая |
| 3 | Окрестности села Псыгансу 43°38′892″ с.ш., 43°80′432″ в.д., 577 м над ур. моря |
Горная серая лесная, среднемощная, среднесуглинистая на коричнево-буром суглинке | Грабово-буково-мертвопокровный лес, высокосомкнутый (0.9), высота стволов 16–17 м, подрост и подлесок отсутствует, мощность лесной подстилки (опад) 5 см |
| 4 | Между селом Герпегеж и поселком Кашхатау 43°33′549″ с.ш., 43°65′433″ в.д., 642 м над ур. моря |
Горная дерново-карбонатная, типичная, многогумусная среднемощная, среднекаменистая, среднесуглинистая на карбонатном элювио-делювии известняковых пород |
Горный луг, злаково-разнотравное сообщество, ОПП 100%, высота травостоя 40–60 см, мощная дернина (10 см), в травостое злаковые, бобовые, душица обыкновенная, подорожник средний, погремок малый, мята кавказская, лапчатка прямая, незабудка мелкая, шалфей сухостойный и др. |
Дополнительно проводили отбор проб из верхних (наиболее биогенных) горизонтов горных почв (0–20 см), чтобы определить усредненные значения контролируемых показателей. Для каждого типа почв было заложено 5 пробных площадок (100 м2), на которых (в соответствии со стандартными методиками) провели геоботанические описания и отобрали смешанные почвенные образцы (ГОСТ 17.4.4.02-84; Миркин и др., 1989).
Лабораторные анализы выполняли в 3–9-кратной повторности. Содержание углерода органического вещества определяли по методу Тюрина в модификации Никитина (Казеев и др., 2003), рН водной суспензии (1 : 2.5) – потенциометрически. Активность ферментов (инвертазы, уреазы, фосфатазы, дегидрогеназы) определяли колориметрически, каталазы – газометрически по методикам Галстяна в модификации Ф.Х. Хазиева (1982). Показатели активности ферментов оценивали по шкале Э.И. Гапонюк, С.В. Малахова (1985). Суммарную профильную ферментативную активность для каждого фермента и каждого генетического горизонта рассчитали в соответствии с методикой Д.Г. Звягинцева (Звягинцев, 1978), в соответствии с формулой:
М = аbd,
где а – активность фермента в расчете на 1 г почвы; b – мощность горизонта, см; d – плотность почвы (г см–3).
Далее суммировали активность ферментов во всем профиле почвы.
Физиологическую активность микробной биомассы установили по БД и СИД (Anderson, 2003). Определение проводили в соответствии с методическими разработками Н.Д. Ананьевой (2003; Ананьева и др., 2011). Для оценки скорости выделения СО2 использовали классическую методику, предложенную А.Ш. Галстяном (1961) и рекомендуемую другими авторами (Казеев и др., 2003). Метод основан на том, что продуцированный при дыхании микроорганизмов во влажной почве (60% полной влагоемкости) СО2 поглощается раствором NaOH (0.1-нормальным) с последующим титрованием непрореагировавшей щелочи (0.05-нормальным НСl) после 24 ч инкубации почвенного образца массой 10 г при 22°С. По разнице результатов титрования контрольного (без почвы) и опытного образцов определяли количество выделившегося СО2 (мг 100 г–1 24 ч–1). Используемый метод, хотя он и уступает приборным в скорости определения, имеет высокую воспроизводимость, не требует специального оборудования и позволяет решить поставленные в данном исследовании задачи. Прединкубацию образцов осуществляли при оптимальной влажности почв (60%) в течение 7 сут при температуре 22°С в полиэтиленовых пакетах с воздухообменом. Скорость СИД оценивали по скорости дыхания микроорганизмов после обогащения почвы глюкозой (0.2 мл г–1 сухой почвы; титр 0.05 г глюкозы) и инкубирования в течение 4 ч при температуре 22°С. Для расчетов содержания углерода микробной биомассы (Смик) скорость СИД выражали в мкл СО2 г–1 почвы ч–1.
Смик (мкг С г–1 почвы) =
= СИД (мкл СО2 г–1 почвы ч–1) × 40.04 +
+ 0.37 (Anderson, Domsch, 1978).
Запас углерода микробной биомассы в слое мощностью 20 см рассчитывали с учетом плотности исследуемых почв:
запас Смик (г м–2) = Смик (г г–1 почвы) × d (г см–3) × V,
где V – объем почвы в слое 20 см и площади 1 м2 = = 10 000 см2.
Плотность почв (d (г см–3)) определяли в соответствии с ГОСТ 5180-2015.
Удельное дыхание микробной биомассы определили как отношение скорости БД к содержанию углерода микробной биомассы:
qСО2 (мкг СО2-С мг–1 Смик ч–1) = БД/Смик.
Коэффициент микробного дыхания (не имеющий размерности) рассчитали как отношение скорости БД к скорости СИД:
QR = БД : СИД.
Долю углерода микробной биомассы в общем органическом углероде почвы рассчитали как Смик : Сорг, %) (Ананьева, 2003).
Для определения и сравнения общего уровня биологической активности различных типов горных почв использовали методику расчета интегрального показателя эколого-биологического состояния почв (ИПЭБСП) (Колесников и др., 2002; Казеев и др. 2004). Для расчета ИПЭБСП (отн. %) максимальное значение каждого из показателей в выборке принимали за 100% и по отношению к нему в процентах выражали значение этого же показателя в относительных процентах в остальных образцах:
БI = (Бх/Бmax) × 100%,
где Б1 – относительный балл показателя; Бх – фактическое значение показателя; Бmax – максимальное значение показателя.
Затем суммировали относительные значения контролируемых показателей, абсолютные величины которых суммированы быть не могут, так как имеют разные единицы измерения, и рассчитывали средний оценочный балл изученных показателей:
Бср = (Б1 + Б2 + Б3 … + Бn)/N,
где Бср – средний оценочный балл показателей; N – число показателей. Интегральный показатель эколого-биологического состояния почвы рассчитывали по формуле:
ИПЭБСП = (Бср/Бср. mах) × 100%,
где Бср – средний оценочный балл всех показателей; Бср. mах – максимальный оценочный балл всех показателей.
При вычислении ИПЭБСП (отн. %) использовали данные, характеризующие активность пяти ферментов, скорость БД, запасы углерода органического вещества и углерода микробной биомассы, определенные с учетом плотности сложения верхних горизонтов и более точно отражающие свойства исследуемых почв.
Статистическую обработку полученных данных осуществляли в программе “Statistica-10.0”. Рассчитали средние, ошибки средних, коэффициенты корреляции и детерминации. Достоверности различия изученных почвенных характеристик оценивали с помощью t-критерия Стьюдента. В исследовании принят уровень значимости, равный 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Горная бурая лесная глеевая почва (разрез № 1) обладает слабо дифференцированным профилем. Почва влажная, мажущаяся, тяжелосуглинистая, каменистые включения наблюдаются с поверхности и по всему профилю (галька 0.5–5 см). Под лесной подстилкой, состоящей из опада в разной степени разложения, расположен наиболее гумусированный горизонт А, темно-серый, с бурым оттенком. Ниже бурые тона усиливаются, содержание углерода микробной биомассы резко снижается (на 59%), увеличивается плотность сложения (табл. 2). Комковатая структура верхнего горизонта меняется на крупнокомковато-глыбистую, ниже горизонта В1 структура не выражена. Горизонты В2 и ВС неоднородно окрашены (желто-бурые с сизыми пятнами глея), плотные, глинистые, мокрые.
Таблица 2.
Изменение физико-химических и биологических показателей в профиле горной бурой лесной глеевой почвы (разрез № 1)
| Горизонт/глубина отбора образцов, см | Плотность, г cм–3 | рН (Н2О) |
Содержание C, % |
Скорость БД, мкг СО2 1 г–1 ч–1 |
Скорость СИД, мкг СО2 1 г–1 ч– 1 |
Смик, мкг С г–1 |
|---|---|---|---|---|---|---|
| А 0–12 | 1.1 | 5.20 | 4.7 | 11 | 56.2 | 1217 |
| АВ 12–20 | 1.3 | 5.20 | 1.9 | 3.9 | 27.9 | 609 |
| В1 20–45 | 1.5 | 5.52 | 0.7 | 3.2 | 23.9 | 518 |
| В2g 45–65 | 1.3 | 5.60 | 0.4 | 3.1 | 17.6 | 386 |
| ВС 65–100 | 1.3 | 6.43 | 0.5 | 1.9 | 18.4 | 396 |
| Активность ферментов | ||||||
| Горизонт/глубина отбора образцов, см | Инвертаза, мг глюкозы 1 г–1 24 ч–1 |
Фосфатаза, мг Р2О5 100 г–1 ч–1 |
Уреаза, мг NH3 10 г–1 24 ч–1 |
Каталаза, мг О2 г–1 мин–1 |
Дегидрогеназа, мг ТФФ 10 г–1 24 ч–1 |
|
| А 0–12 | 10.8 | 75.1 | 34.4 | 2.1 | 1.1 | |
| АВ 12–20 | 8.3 | 70.2 | 5.5 | 0.3 | 0.9 | |
| В1 20–45 | 6.6 | 22.8 | 0.4 | 0 | 0.5 | |
| В2g 45–65 | 6.6 | 3.7 | 0 | 0.2 | 0.3 | |
| ВС 65–100 | 8.0 | 0 | 0 | 0.6 | 0.2 | |
Переувлажнение, обусловливающее процессы оглеения нижних горизонтов, вызвано боковыми подтоками поверхностных и грунтовых вод. Содержание углерода органического вещества в нижней части профиля очень низкое, каменистых включений становится больше, их размер увеличивается (крупная галька диаметром около 10 см). Реакция почвенного раствора кислая по всему профилю, но с глубиной значения рН постепенно повышаются.
Скорость БД и СИД резко снижается уже в горизонте АВ. Следует отметить, что описываемая почва обладает самыми низкими показателями интенсивности БД в ряду изученных естественных почв. Соответственно показателям СИД, ниже горизонта А содержание Смик резко снижается (на 50%) с высокого (более 1001 мкг С г–1) до среднего уровня (501–1000 мкг С г–1). В горизонте В1 содержание Смик низкое (201–500 мкг С г–1). Оценка содержания Смик дана в соответствии с используемой градацией (Ананьева, 2003). Скорость СИД в глеевом горизонте увеличивается и, соответственно, содержание Смик несколько возрастает на фоне продолжающей уменьшаться скорости фонового БД. Можно предположить, что причина заключается в проявлении сохранившейся потенциальной физиологической активности почвенной микробной биомассы. Вероятно, аэробные микроорганизмы сохраняют свою жизнеспособность и в глеевых горизонтах, на что указывают значения скорости СИД и содержания Смик, полученные в лабораторных условиях оптимального увлажнения и достаточного количества питательного субстрата. При функционировании в условиях повышенного увлажнения метаболизм аэробных микроорганизмов подавляется (Алехина, 2001; Головченко, 2001), поэтому скорость фонового микробного дыхания (БД) в глеевых горизонтах снижается.
При анализе данных, характеризующих каталитическую деятельность изученных ферментов в профиле бурой лесной глеевой почвы (табл. 2), установлена очень слабая активность оксидоредуктаз (каталазы и дегидрогеназы), в том числе и в горизонте А. Ниже по профилю указанные ферменты практически не проявляют активности. Возможно, в условиях высокого увлажнения и недостатка кислорода в сочетании с кислой реакцией почвенного раствора, происходит ингибирование оксидоредуктаз. Проведенные нами ранее исследования показали, что активность каталазы выше в карбонатных почвах, обладающих слабощелочной реакцией (темно-каштановых, черноземах южных) по сравнению с кислыми лесными почвами (Гедгафова и др., 2015; Горобцова и др., 2015).
Активность гидролаз (инвертазы, фосфатазы, уреазы) в верхних горизонтах профиля заметно выше, чем оксидоредуктаз. Среди перечисленных ферментов лидирует фосфатаза, высокие показатели которой установлены в горизонтах А и АВ, а средние – в горизонте В1. В нижних горизонтах активность данного фермента резко снижается (на 84%) и в горизонте ВС не обнаружена. Уреаза высокоактивна только в верхнем горизонте. Ниже (в горизонта В) показатель ее деятельности падает на 84% и соответствует очень слабому уровню, ниже действие этого фермента практически не проявляется. Инвертаза наименее активна в ряду гидролаз – ее активность слабая во всем профиле, однако даже при переходе к материнской породе этот фермент сохраняет свою функцию, причем, показатели горизонтов А и ВС вполне сопоставимы (табл. 2).
Полученные данные свидетельствуют о достаточно резком снижении большинства показателей БА вниз по профилю. При этом верхний горизонт, обладающий высоким содержанием углерода органического вещества и Смик, демонстрирует низкие значения некоторых рассмотренных параметров БА (активность оксидоредуктаз, инвертазы и скорость БД).
Естественные горные серые лесные почвы представлены двумя разрезами, один из которых заложен на опушке, под злаково-разнотравным сообществом (разрез № 2), а другой – на мертвопокровном участке грабово-букового леса (разрез № 3). Строение профиля и биологические показатели отражают особенности условий, при которых функционируют описываемые почвы, максимальные различия наблюдаются в верхних горизонтах.
Верхний горизонт почвы разреза № 2 – дерновый (Ад), около 30% объема которого занимает корневая система травянистых растений. Темно-серый, комковато-ореховатый, насыщен корнями, среднесуглинистый, высокогумусированный (табл. 3), слабощелочной, множественные включения мелкого щебня и гальки. Горизонт Ад характеризуется максимальными в ряду изученных почв показателями респираторной функции микробной биомассы, содержания Смик, активности всех ферментов, за исключением фосфатазы. Слабощелочная реакция водной суспензии, нехарактерная для серых лесных почв, выделяет этот горизонт (ниже по профилю реакция нейтральная).
Таблица 3.
Изменение физико-химических и биологических показателей в профиле горных серых лесных почв, под различными типами леса
| Горизонт/глубина отбора образцов, см | Плотность, г cм–3 | рН (Н2О) |
Содержание C, % |
Скорость БД, мкг СО2 1 г–1 ч–1 |
Скорость СИД, мкг СО2 1 г–1 ч–1 |
Смик, мкг С г–1 |
|---|---|---|---|---|---|---|
| Горная серая лесная под грабово-буково-разнотравным лесом (разрез № 2) | ||||||
| Ад 0–5 | 1.1 | 7.9 | 6.6 | 31.7 | 204.3 | 4321 |
| А1 5–17 | 1.3 | 6.5 | 3.8 | 12.8 | 76.7 | 1643 |
| А1А2 17–32 | 1.3 | 6.6 | 2.3 | 11.4 | 42.5 | 913 |
| В 32–63 | 1.5 | 7.0 | 0.7 | 7.2 | 33.0 | 700 |
| ВС 63–110 | 1.4 | 7.0 | 0.5 | 6.8 | 23.9 | 518 |
| Горная серая лесная под грабово-буково-мертвопокровным лесом (разрез № 3) | ||||||
| А0 0–5 | 0.9 | 6.2 | 4.1 | 10.4 | 62.5 | 1339 |
| А1 5–16 | 0.9 | 6.5 | 3.4 | 8.9 | 56.4 | 1217 |
| А1А2 16–25 | 1.1 | 6.6 | 1.8 | 6.8 | 42.0 | 913 |
| А2В 25–43 | 1.3 | 6.9 | 0.8 | 6.5 | 45.9 | 974 |
| В 43–70 | 1.4 | 7.2 | 0.7 | 4.3 | 36.8 | 791 |
| ВС 70–100 | 1.4 | 7.4 | 0.5 | 1.4 | 38.7 | 822 |
| Активность ферментов | ||||||
| Горизонт/глубина отбора образцов, см | Инвертаза, мг глюкозы 1 г–1 24 ч–1 |
Фосфатаза, мг Р2О5 100 г–1 ч–1 |
Уреаза, мг NH3 10 г–1 24 ч–1 |
Каталаза, мг О2 г–1 мин–1 |
Дегидрогеназа, мг ТФФ 10 г–1 24 ч–1 |
|
| Горная серая лесная под грабово-буково-разнотравным лесом (разрез № 2) | ||||||
| Ад 0–5 | 27.1 | 26.1 | 44.8 | 9 | 8.1 | |
| А1 5–17 | 10.5 | 13.3 | 42.9 | 2.7 | 1.1 | |
| А1А2 17–32 | 7.8 | 12.7 | 9.5 | 0.8 | 0.4 | |
| В 32–63 | 5.5 | 2.0 | 0 | 1 | 0.5 | |
| ВС 63–110 | 5.3 | 0 | 0 | 1.4 | 0.2 | |
| Горная серая лесная под грабово-буково-мертвопокровным лесом (разрез № 3) | ||||||
| А0 0–5 | 7.7 | 18.5 | 44.1 | 4.4 | 3.6 | |
| А1 5–16 | 8.3 | 12.8 | 28.8 | 3.9 | 1.5 | |
| А1А2 16–25 | 6.8 | 6.7 | 10.3 | 2.1 | 1.0 | |
| А2В 25–43 | 5.4 | 0.8 | 0.8 | 2.2 | 0.5 | |
| В 43–70 | 5.8 | 0.6 | 0 | 1.7 | 0.1 | |
| ВС 70–100 | 5.6 | 0 | 0 | 0.4 | 0 | |
Органогенный горизонт (А0) горной серой лесной почвы разреза № 3 представляет собой хорошо разложившуюся часть лесной подстилки, пронизанную грибным мицелием. Верхняя часть подстилки состоит из неразложившегося опада бука восточного и граба кавказского. Горизонт А0 рыхлый, обилие трухи (сильно разложившегося лиственного опада), свежий, темно-серый, мелкокомковато-порошистый, среднесуглинистый, каменистые включения отсутствуют, многочисленные ходы дождевых червей, белые нити мицелия. Отличием строения профиля почвы под мертвопокровным лесом является наличие переходного горизонта А2В (табл. 3) серо-бурого, неоднородно окрашенного, сырого, комковато-глыбистого, тяжелосуглинистого, присутствуют корни различного диаметра и червороины.
Определенные различия наблюдались и в биологических характеристиках верхних горизонтов сравниваемых серых лесных почв. В горизонте А0 установлено достаточно высокое содержание углерода органического вещества, однако оно значительно (на 39%) ниже, чем в дерновом горизонте почвы горной серой лесной почвы под грабово-буковым разнотравным лесом (разрез № 2). Вниз по профилю разреза № 3 содержание углерода органического вещества заметно снижается (на 19%), но резкое падение (на 43%) при переходе к горизонту А1, зафиксированное в горной серой лесной почве под грабово-буковым разнотравным лесом, отсутствует. Органогенный горизонт А0 разреза № 3 обладает меньшими показателями дыхательной активности почвенной микробной биомассы (БД слабее на 67%, СИД и содержание Смик – на 69%). Активность всех ферментов в горизонте А0 ниже, чем в Ад. Показатели уреазы, инвертазы и дегидрогеназы переходят в более низкую категорию (шкала оценки (Гапонюк, Малахов, 1985)) фосфатаза и каталаза сохранили среднюю активность, но абсолютные значения, характеризующие их деятельность, ниже, чем в соответствующих горизонтах разреза № 2 на 30 и 56% соответственно.
Горизонты А1 и А1А2 сравниваемых почв характеризуются усилением бурого цвета в окраске, увеличением влажности и плотности, более грубой крупнокомковатой и глыбистой структурой, среднесуглинистым гранулометрическим составом. Отличие профиля разреза № 2 – обилие каменистых включений, которые отсутствуют в профиле разреза № 3, и проникновение корневой системы травянистых растений на глубину до 50 см. К общим чертам можно отнести множество червороин, присутствие дождевых червей и других представителей почвенной мезофауны. Эти горизонты обладают более близкими значениями контролируемых параметров БА, однако большинство абсолютных значений, характеризующих содержание углерода органического вещества и Смик, скорость БД и СИД, активность гидролитических ферментов, несколько выше в профиле почвы № 2. Активность каталазы и дегидрогеназы незначительно выше в профиле почвы № 3. Рассматриваемые показатели различаются в меньшей степени, чем в поверхностных горизонтах (на 1–33%). В горизонтах В и ВС наблюдаются сопоставимые значения, характеризующие деятельность оксидоредуктаз в описываемых почвах.
Представленные данные свидетельствуют, что, несмотря на близость расположения описанных разрезов, строение профиля и биологические свойства почв под различными типами леса имеют определенные особенности. В наибольшей степени различаются верхние горизонты, морфологические признаки и биологические показатели которых, вероятно, зависят от характера напочвенного покрова (травянистый или мертвопокровный).
Горная дерново-карбонатная почва (разрез № 4), относящаяся к фациальному подтипу теплых периодически промерзающих, функционирует на карбонатном элювио-делювии известняковых пород, под богатой горно-луговой растительностью. В биоэкологических условиях пояса широколиственных лесов – промывного водного режима и обилия органических остатков формируется почвенный профиль с хорошо выраженным гумусовым горизонтом, слабощелочной реакцией почвенного раствора (табл. 4).
Таблица 4.
Изменение физико-химических и биологических показателей в профиле горной дерново-карбонатной почвы (разрез № 4)
| Горизонт/глубина отбора образцов, см | Плотность, г cм–3 |
рН (Н2О) |
Содержание C, % |
Скорость БД, мкг СО2 1 г–1 ч–1 |
Скорость СИД, мкг СО2 1 г–1 ч–1 |
Смик, мкг С г–1 |
|---|---|---|---|---|---|---|
| Ад 0–10 | 1.1 | 8.0 | 4.3 | 20.0 | 73.4 | 1562 |
| А 10–36 | 1.2 | 7.8 | 4.2 | 19.1 | 67.7 | 1440 |
| АВ 36–70 | 1.5 | 8.2 | 2.0 | 18.7 | 35.9 | 770 |
| Вса 70–100 | 1.5 | 8.4 | 1.6 | 14.5 | 23.7 | 507 |
| ВС 100–120 | 1.5 | 8.4 | 1.4 | 12.3 | 22.7 | 487 |
| Активность ферментов | ||||||
| Горизонт/глубина отбора образцов, см | Инвертаза, мг глюкозы 1 г–1 24 ч–1 |
Фосфатаза, мг Р2О5 100 г–1 ч–1 |
Уреаза, мг NH3 10 г–1 24 ч–1 |
Каталаза, мг О2 г–1 мин–1 |
Дегидрогеназа, мг ТФФ 10 г–1 24 ч–1 |
|
| Ад 0–10 | 11.1 | 5.7 | 103.3 | 7.8 | 7.8 | |
| А 10–36 | 9.9 | 0.6 | 90.5 | 7.5 | 7.5 | |
| АВ 36–70 | 7.6 | 0.2 | 30.5 | 6.2 | 3.9 | |
| Вса 70–100 | 7.6 | 0 | 2.5 | 5.1 | 2.2 | |
| ВС 100–120 | 7.3 | 0 | 0 | 3.0 | 1.3 | |
Рассматриваемую почву отличает высокое содержание каменистых включений (30–35% от массы почвы). Частицы щебня разного диаметра от очень мелких до значительных (5 см и более) бурно реагируют с кислотой. С глубиной содержание щебнистых включений возрастает и в нижней части профиля составляет более 50% от массы почвы, в горизонте ВС щебень перемешан с желто-бурой карбонатной глиной.
Поверхностный горизонт Ад темно-серый, почти черный, с белыми и серыми вкраплениями щебня, слегка влажный, зернистый (почвенные “бусы”), среднесуглинистый, около 50% объема занимает переплетенная корневая система травянистых растений, дождевые черви. Бурно вскипает, переход постепенный. Нижележащий горизонт А обладает таким же высоким содержанием углерода органического вещества, как и дерновый, что проявляется в очень темном цвете и комковато-ореховатой хорошо выраженной структуре. Гранулометрический состав среднесуглинистый, присутствуют многочисленные корни и червороины, возрастает плотность сложения, размер и количество щебнистых включений, переход постепенный, заметен по окраске.
В горизонте АВ проявляются бурые тона, неоднородность окраски усиливается обилием беловато-серых щебнистых включений, возрастает влажность почвы, структура грубая, крупнокомковато-глыбистая. В горизонте Вса отмечаются выцветы карбонатов, большая влажность и каменистость, цвет буровато-серый, неоднородно окрашенный, гранулометрический состав среднесуглинистый, структура не выражена. Нижняя часть профиля (горизонт ВС) каменистая, частицы почвы между щебнем и глиной придают горизонту пеструю окраску (возможно механическое перемещение почвенной массы между обломками щебня), структура не выражена, гранулометрический состав – тяжелосуглинистый. Почвенная масса и каменистые включения бурно вскипают с поверхности и по всему профилю, что позволяет отнести почву к подтипу дерново-карбонатных типичных.
Показатели биологической активности дерново-карбонатной почвы отличаются от аналогичных характеристик рассмотренных ранее лесных почв. Наблюдается высокое содержание углерода органического вещества, вплоть до горизонта АВ. Гумусированные средняя и нижняя части профиля, вероятно, являются следствием заполнения частицами почвы пространства между щебнистыми включениями. Скорости БД самые высокие в ряду изученных почв, значения скорости СИД и содержания Смик уступают только показателям верхних горизонтов горной серой лесной почвы под грабово-буковым разнотравным лесом (табл. 4).
Деятельность ферментов в дерново-карбонатной почве имеет определенные особенности. В условиях слабощелочной реакции почвенного раствора активность оксидоредуктаз достигает в верхних горизонтах среднего уровня (каталаза среднеактивна даже в горизонте ВС). Среди гидролаз наиболее высокими значениями (вплоть до Вса) характеризуется уреаза, а показатели фосфатазы самые низкие в ряду изученных почв. Активность фосфатазы слабая даже в дерновом горизонте, а ниже по профилю – близка к нулевым значениям. Инвертаза слабоактивна, но сохраняет близкие значения абсолютных показателей вплоть до материнской породы. Параметры биологической активности горной дерново-карбонатной почвы близки к аналогичным показателям горных черноземов, функционирующих на высоте 500–1000 м над ур. моря в условиях эльбрусского варианта поясности (Хакунова и др., 2018), что, вероятно, является следствием влияния богатой злаково-разнотравной растительности, характерной и для горных черноземов, и для дерново-карбонатных почв.
Как следует из проведенного анализа материалов полевых и лабораторных исследований, каждая рассмотренная почва имеет свои особые черты проявления биологических свойств, поэтому представленное выше подробное описание профильного распределения параметров БА различных почв является необходимым компонентом почвенно-экологических исследований. Однако только совокупность полученных данных является основой, благодаря которой может быть установлен общий уровень профильной БА каждой почвы. Для того, чтобы сопоставить полученные данные и сравнить БА основных типов горных почв пояса широколиственных лесов, необходим единый оценочный показатель. Критерием оценки уровня БА изученных почв является ИПЭБСП (отн. %), эффективность использования которого была доказана при изучении естественных и агрогенных почв равнинной части Кабардино-Балкарии (Гедгафова и др., 2015; Горобцова и др., 2015, 2017).
Как показывает представленная диаграмма (рис. 2б), максимальный уровень БА (принятый за 100%) установлен в профиле горной дерново-карбонатной почвы. Значительно более высокая БА дерново-карбонатных почв, по сравнению с лесными, обусловлена факторами почвообразования, формирующими эти почвы – мощная травяная растительность, обилие органических остатков, достаточное количество тепла и влаги, высокая карбонатность профиля.
Рис. 2.
ИПЭБСП (%), характеризующие биологическую активность изученных почв пояса широколиственных лесов: а – значения верхних горизонтов (0–20 см): 1 – Горные дерново-карбонатные; 2 – Горные бурые лесные глеевые; 3 – Горные бурые лесные; 4 – Горные серые лесные. б – суммарные профильные значения: 1 – Горная дерново-карбонатная; 2 – Горная бурая лесная глеевая; 3 – Горная серая лесная (разнотравный напочвенный покров); 4 – Горная серая лесная (мертвопокровный).
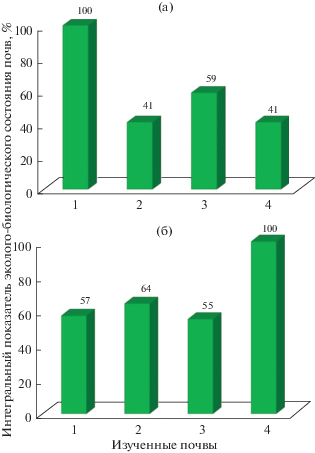
Различия профильных значений ИПЭБСП (отн. %) рассмотренных горных серых лесных почв являются следствием влияния напочвенного растительного покрова. Более высокая БА установлена в почве под грабово-буковым разнотравным лесом по сравнению с почвой под мертвопокровным лесом. Особенности растительного сообщества влияют на почвенные характеристики, а также интенсивность и направленность почвенных процессов. В свою очередь, свойства почвы определяют состав растительного сообщества, а также направление и скорость аутогенных сукцессий (Rode, 1999; Лукина и др., 2008).
Следует отметить, что общий уровень профильной БА горной бурой лесной глеевой почвы совпадает с аналогичной характеристикой горной серой лесной под мертвопокровным лесом. Вероятно, в данном случае проявляется влияние не только напочвенного растительного покрова, достаточно развитого под бурой лесной глеевой почвой (ОПП 30%), но и процессов переувлажнения и оглеения, снижающих ряд контролируемых параметров БА (содержание углерода органического вещества, микробные показатели, активность уреазы, каталазы, дегидрогеназы) в средних и нижних горизонтах профиля этой почвы (табл. 2, 3).
Проведенные исследования показали, что в профилях рассмотренных типов естественных горных почв пояса широколиственных лесов динамика изученных характеристик и общий уровень БА обладают различиями, которые определяются характером напочвенного растительного покрова и спецификой почвообразовательных процессов.
Представленные выше данные описывают каждый тип почвы в конкретной точке ее формирования, а высокая вариабельность составляющих БА требует формирования репрезентативной выборки, проведения статистического анализа достаточно большого объема данных и определения средних значений контролируемых показателей. Поэтому, помимо профильных исследований, было проведено подробное изучение верхних, наиболее биогенных горизонтов (0–20 см) горных почв пояса широколиственных лесов, сведения о которых приведены в табл. 5.
Таблица 5.
Средние значения физико-химических и биологических показателей верхних горизонтов (0–20 см) естественных горных почв пояса широколиственных лесов терского варианта поясности Центрального Кавказа (в пределах Кабардино-Балкарии)
| Почвы | ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ | Плотность, г см–3 |
Углерод органического вещества | |
|---|---|---|---|---|
| содержание С, % | запасы С, т га–1 | |||
| Горные бурые лесные глеевые | 5.02 ± 0.21 | 1.0 ± 0.09 | 4.6 ± 1.7 | 97.7 ± 45.6 |
| Горные серые лесные | 6.32 ± 0.29 | 0.9 ± 0.04 | 4.3 ± 0.7 | 86.0 ± 19.3 |
| Горные дерново-карбонатные | 7.68 ± 0.12 | 0.7 ± 0.09 | 7.6 ± 3.0 | 102.9 ± 41.4 |
| Показатели микробиологической активности | ||||
|---|---|---|---|---|
| Почвы | Скорость БД мкг СО2 1 г–1 ч–1 |
Скорость СИД мкг СО2 1 г–1 ч–1 |
Смик, мкг С г–1 | Запас Смик, г м–2 |
| Горные бурые лесные глеевые | 9.3 ± 3.1 | 54.2 ± 14.7 | 1200 ± 326 | 193 ± 68 |
| Горные серые лесные | 11.9 ± 1.5 | 76.0 ± 9.8 | 1681 ± 216 | 336 ± 42 |
| Горные дерново-карбонатные | 24.3 ± 1.1 | 162.6 ± 18.0 | 3078 ± 652 | 492 ± 59 |
| Активность ферментов | |||||
|---|---|---|---|---|---|
| Почвы | Дегидрогеназа, мг ТФФ 10 г–1 24 ч–1 |
Каталаза,
мг О2 г–1 мин–1 |
Инвертаза, мг глюкозы 1 г–1 24 ч–1 |
Уреаза,
мг NH3 10 г–1 24 ч–1 |
Фосфатаза, мг Р2О5 100 г–1 ч–1 |
| Горные бурые лесные глеевые | 0.74 ± 0.74 | 2.08 ± 0.52 | 17.37 ± 3.27 | 18.42 ± 4.59 | 107.59 ± 37.45 |
| Горные серые лесные | 1.40 ± 0.68 | 3.20 ± 0.55 | 16.51 ± 1.50 | 19.33 ± 3.86 | 22.79 ± 5.70 |
| Горные дерново-карбонатные | 8.28 ± 1.74 | 6.43 ± 0.62 | 26.71 ± 3.46 | 81.48 ± 20.97 | 26.96 ± 3.16 |
Кислотно-щелочные условия существенно (t > 2.99; Р < 0.02) различаются в верхних горизонтах дерново-карбонатных и лесных почв, что определяет особенности протекания биохимических и микробиологических процессов. Наиболее гумусированы горные дерново-карбонатные почвы. Содержание углерода органического вещества в верхних органогенных горизонтах лесных почв высокое, а полученные средние показатели вполне сопоставимы. Высокие запасы углерода органического вещества в горизонте 0–20 см всех изученных типов почв говорят об активном протекании процессов гумификации растительных остатков, активности микробиологических и биохимических процессов. Это подтверждают данные респираторной активности почвенной микробной биомассы, высокие для всех изученных почв. Следует отметить, что микробные показатели горных дерново-карбонатных почв выделяются на фоне данных, характеризующих лесные почвы (табл. 5).
Проведенные исследования свидетельствуют, что скорость БД и СИД в верхних горизонтах дерново-карбонатных почв статистически значимо выше (более чем на 50%), чем в лесных почвах (t > 4.5; Р < 0.03). Условия, в которых сформировались дерново-карбонатные почвы, способствуют активному развитию и функционированию почвенных микроорганизмов. Это подтверждается данными, характеризующими содержание и запас микробной биомассы (Смик) в изученных почвах. Указанные показатели в дерново-карбонатных почвах существенно выше, чем в лесных. Обнаруженные различия также статистически значимы (t > 2.3; Р < 0.05).
Расчет микробных метаболических коэффициентов (qСО2; QR; Смик/Сорг) показал, что верхние горизонты исследуемых почв обладают близкими значениями удельного дыхания микробной биомассы (qСО2 находится в пределах 0.15 ± 0.01–0.16 ± 0.02 мкг СО2-С/мг Смик/ч) и коэффициентов микробного дыхания (QR составляет 6.69 ± ± 0.82–7.35 ± 0.87). Статистически значимых различий данных показателей в различных типах почв не установлено. Представленные значения свидетельствуют об устойчивом функционировании микробных процессов, связанных с трансформацией соединений углерода, и отсутствии стресса в микробных сообществах естественных горных почв.
Доля углерода микробной биомассы в общем органическом углероде почв (Смик : Сорг ) достаточно высока в верхних горизонтах горных дерново-карбонатных и горных серых лесных почв (4.22 ± 0.48 и 4.10 ± 0.34% соответственно). В горных бурых лесных глеевых почвах этот показатель значимо (t > 2.41; Р < 0.04) ниже и составляет 2.64 ± 0.44%. Возможно, полученные данные отражают генетические особенности глеевых почв, функционирующих в условиях периодического переувлажнения и анаэробиозиса. Резкие колебания водного и воздушного режимов могут приводить к уменьшению части “живого” углерода в бурых лесных глеевых почвах.
Деятельность изученных ферментов в верхних горизонтах рассматриваемых типов почв заметно различается. Оксидоредуктазы наиболее активны в горных дерново-карбонатных почвах, обладающих слабощелочной реакцией почвенного раствора. Наименьший уровень активности этих ферментов зафиксирован в горных бурых лесных глеевых почвах, характеризующихся кислыми условиями. Ингибирование оксидоредуктаз в кислой среде и активизация их деятельности в щелочной отмечались нами в более ранних исследованиях, а также и другими авторами (Галстян, 1974; Хазиев, 1982; Гедгафова и др., 2015; Горобцова и др., 2015, 2017).
Гидролитические ферменты характеризуются, в основном, средним уровнем активности. Во всех изученных почвах наблюдаются сопоставимые значения показателей инвертазной активности. Средний уровень активности уреазы практически совпадает в верхних горизонтах лесных почв, а наиболее высокими значениями обладают горные дерново-карбонатные почвы. Высокая активность фосфатазы зафиксирована только в горных бурых лесных глеевых почвах, верхние горизонты остальных почв незначительно различаются по этому показателю и соответствуют среднему уровню.
Для сравнения общего уровня деятельности всех изученных ферментов в различных подтипах горных почв рассчитали показатели суммарной относительной ферментативной активности, интегрирующие данные, полученные для всех изученных ферментов (Звягинцев, 1978). Максимальный уровень ферментативной активности (принятый за 100%) установлен для дерново-карбонатных почв, далее следуют бурые лесные глеевые (54%) и серые лесные почвы (41%). Представленные значения наглядно демонстрируют значительно более высокий уровень ферментативной активности дерново-карбонатных почв, вероятно, связанный с высоким содержанием органического вещества (Галстян, 1974; Хазиев, 1982). В изученных подтипах горных почв установлены высокие коэффициенты детерминации между содержанием углерода органического вещества и активностью ферментов (R2 = 0.7, при P = 0.0009), а также респираторными показателями микробной биомассы (R2 = 0.9, при P = 0.009).
Представленные на диаграмме (рис. 2а) значения ИПЭБСП (отн. %) верхних горизонтов (0–20 см) естественных горных почв, также как и профильные показатели, демонстрируют существенно более высокую БА верхних горизонтов горных дерново-карбонатных почв. Лесные горные почвы обладают сопоставимыми значениями интегрального показателя, а несколько большая величина ИПЭБСП (отн. %) горной бурой лесной глеевой почвы является, вероятно, следствием очень высокой активности фосфатазы, установленной в этих почвах. Выяснение причин высокой фосфатазной активности в верхних горизонтах горных бурых лесных глеевых почв требует дальнейших исследований.
ЗАКЛЮЧЕНИЕ
Проведен анализ профильного распределения морфогенетических, физико-химических и биологических показателей основных типов почв изученных территорий. На основе совокупности полученных данных рассчитаны значения ИПЭБСП (отн. %), которые являются критерием оценки общего уровня их профильной БА. Горные почвы образуют следующий ряд по убыванию профильной БА: горная дерново-карбонатная под горно-луговой растительностью (100%) > горная серая лесная под грабово-буково-разнотравным лесом (59%) > горная серая лесная под грабово-буково-мертвопокровным лесом (41%) = горная бурая лесная под буково-грабово-ясменниковым лесом (41%). БА профиля горной дерново-карбонатной почвы существенно превышает аналогичные характеристики лесных почв вследствие благоприятных для протекания активных биологических процессов условий, в которых сформировался и функционирует данный тип почв. Сравнение значений ИПЭБСП (отн. %) в группе горных лесных почв позволяет предположить, что на уровень БА могут оказывать влияние характер напочвенного растительного покрова, а также процессы, обусловленные генетическими особенностями изученных почв.
Значения ИПЭБСП (отн. %), полученные на основе репрезентативной выборки, позволяют оценить общий уровень БА верхних горизонтов (0–20 см) изученных естественных почв: горные дерново-карбонатные (100%) > горные бурые лесные глеевые (64%) > горные серые лесные (55%). Данные, установленные на основе средних показателей параметров БА, в целом согласуются с теми, которые получены при описании конкретных профилей горных почв. Несколько более высокий уровень БА в верхних горизонтах горных бурых лесных глеевых почв, по сравнению с профильными данными, вероятно, обусловлен тем, что процессы переувлажнения, оглеения и анаэробные условия, ингибирующие активность ряда биологических процессов, характерны для средних и нижних горизонтов данного типа почв. ИПЭБСП (отн. %) горных дерново-карбонатных почв почти вдвое превышает значения, установленные для группы лесных почв.
При возрастании антропогенного воздействия на почвенный покров горных территорий (сельскохозяйственное использование, рубка леса, возрастающая рекреационная нагрузка) получение и накопление данных, характеризующих свойства почв, которые функционируют в еще сохранившихся естественных условиях, необходимо для оценки состояния нарушенных лесных биогеоценозов.
Список литературы
Алехина Л.К. Оценка бактериального разнообразия в почвенных микрокосмах при разной влажности // Почвоведение. 2001. № 6. С. 847–854.
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 222 с.
Ананьева Н.Д., Сусьян Е.А., Гавриленко Е.Г. Особенности определения углерода микробной биомассы почвы методом субстрат-индуцированного дыхания // Почвоведение. 2011. № 11. С. 1327–1333.
Вальков В.Ф., Казеев К.Ш., Колесников С.И. Методология исследования биологической активности почв на примере Северного Кавказа // Научная мысль Кавказа. Ростов-на-Дону. Издательство Северо-Кавказского научного центра высшей школы. 1999. № 1. С. 32–37.
Галстян А.Ш. Дыхание почвы как один из показателей ее биологической активности // Сообщение лаборатории агрохимии АН АрмССР. Биологические науки. 1961. № 5. С. 69–74.
Галстян А.Ш. Ферментативная активность почв Армении. Ереван: Айастан, 1974. 275 с.
Гапонюк Э.И., Малахов С.В. Комплексная система показателей экологического мониторинга почв // Труды 4-го Всесоюзного совещания. Л.: Гидрометеоиздат, 1985. С. 3–10.
Гедгафова Ф.В., Улигова Т.С., Горобцова О.Н., Темботов Р.Х. Биологическая активность черноземных почв Центрального Кавказа (в пределах терского варианта поясности Кабардино-Балкарии) // Почвоведение. 2015. № 12. С. 1474–1482.
Головченко А.В., Добровольская Н.Г. Численность и запасы микроорганизмов в пойменных почвах реки Протва // Почвоведение. 2001. № 12. С. 1460–1464.
Горобцова О.Н., Улигова Т.С., Темботов Р.Х., Хакунова Е.М. Оценка уровня биологической активности агрогенных и естественных черноземов Кабардино-Балкарии // Почвоведение. 2017. № 3. С. 1–10.
Горобцова О.Н., Хежева Ф.В., Улигова Т.С., Темботов Р.Х. Эколого-географические закономерности изменения биологической активности автоморфных почв равнинных и предгорных территорий северного макросклона Центрального Кавказа (в пределах Кабардино-Балкарии) // Почвоведение. 2015. № 3. С. 347–359.
ГОСТ 17.4.4.02-84. Охрана природы. Почвы. Методы отбора и подготовки проб для химического, бактериологического, гельминтологического анализа.
ГОСТ 17.4.2.01 – Охрана природы. Почвы. Номенклатура показателей санитарного состояния.
ГОСТ 5180-2015. Грунты. Методы лабораторного определения физических характеристик.
Даденко Е.В., Казеев К.Ш., Колесников С.И. Влияние пастбищной нагрузки на ферментативную активность лесных почв Северо-Западного Кавказа // Известия Самарского НЦ РАН. 2016. Т. 18. № 2. С. 345–348.
Звягинцев Д.Г. Биологическая активность почв и шкалы для оценки некоторых ее показателей // Почвоведение. 1978. № 6. С. 48–55.
Казеев К.Ш., Козин В.К., Колесников С.И., Вальков В.Ф. Биологические особенности влажных почв субтропиков // Почвоведение. 2002. № 12. С. 1474–1478.
Казеев К.Ш., Колесников С.И., Вальков В.Ф. Биологическая диагностика и индикация почв: методология и методы исследований // Ростов-на-Дону: Изд-во ЦВВР. 2003. 204с.
Казеев К.Ш., Колесников С.И., Вальков В.Ф. Биология почв юга России // Ростов-на-Дону: Изд-во ЦВВР. 2004. 350 с.
Казеев К.Ш., Тер-Мисканянц Т.А., Колесников С.И., Козунь Ю.С. Биодиагностика экологического состояния почв Западного Кавказа после вырубки леса // Известия Самарского НЦ РАН. 2013. Т. 15. № 3. С. 1299–1301.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Классификация и диагностика почв СССР. М.: Колос, 1977. 225 с.
Колесников С.И., Казеев К.Ш., Вальков В.Ф. Экологические функции почв и влияние на них загрязнения тяжелыми металлами // Почвоведение. 2002. № 12. С. 1509–1514.
Лукина Н.В., Полянская Л.М., Орлова М.А. Питательный режим почв северотаежных лесов. М.: Наука, 2008. 342 с.
Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов. М.: Наука, 1989. 223 с.
Почвенная карта Кабардино-Балкарской АССР. М. 1 : 100 000. ГУГК СССР, М. 1990. Отв. редактор Молчанов Э.Н.
Соколов В.Е., Темботов А.К. Позвоночные Кавказа. Млекопитающие. Насекомоядные. М.: Наука. 1989. 547 с.
Стольникова Е.В., Ананьева Н.Д., Чернова О.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов европейской территории России // Почвоведение. 2011. № 4. С. 479–494.
Темботова Ф.А., Пшегусов Р.Х., Тлупова Ю.М. Леса Северного Макросклона Центрального Кавказа (эльбрусский и терский варианты поясности) // Разнообразие и динамика лесных экосистем России. М.: Товарищество научных изданий КМК, 2012. Т. 1. С. 227–251.
Хазиев Ф.Х. Системно-экологический анализ ферментативной активности почв. М.: Наука. 1982. 203 с.
Хакунова Е. М., Горобцова О.Н., Гедгафова Ф.В., Улигова Т.С., Темботов Р.Х. Изменение биологической активности горных черноземов Центрального Кавказа в результате сельскохозяйственного использования (в границах эльбрусского варианта поясности Кабардино-Балкарии) // Агрохимия. 2018. № 3. С. 12–18
Anderson J.P.E., Domsch K.H. A phisiological method for the quantitative measurement of microbial biomass in soils // Soil Biology and Biochemistry. 1978. V. 10. № 3. P. 215–221.
Anderson T.H. Microbial eco-physiological indicators to assess soil quality // Agriculture, Ecosystems & Environment. 2003. V. 98. P. 285–293.
Rode M.W. Influence of forest growth on former heathland on nutrient input and its consequences for nutrition and management of heath and forest // Forest Ecology and Management. 1999. V. 114. P. 31–43.
World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome. 2014. 181 p.
Дополнительные материалы отсутствуют.