

Лесоведение, 2021, № 1, стр. 3-10
Диагностика состояния лесов на основе ретроспективного анализа спутниковых данных MODIS
А. В. Ковалев a, *, В. И. Воронин b, **, В. А. Осколков b, В. Г. Суховольский c, ***
a ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50, Россия
b Сибирский институт физиологи и биохимии растений СО РАН
664033 Иркутск, Лермонтова, 132, Россия
c Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: sunhi.prime@gmail.com
** E-mail: bioin@sifibr.irk.ru
*** E-mail: soukhovolsky@yandex.ru
Поступила в редакцию 13.02.2017
После доработки 03.12.2019
Принята к публикации 07.10.2020
Аннотация
Рассмотрена возможность оценки состояния древесных растений по данным дистанционного зондирования. Для оценки использованы интегральные показатели кривой сезонной динамики вегетационного индекса (NDVI), которые в течение 2003–2017 гг. сопоставлялись для контрольных и поврежденных пробных площадей в зоне Хамар-Дабана вблизи побережья о. Байкал (Иркутская область). Показано, что использование предложенных интегральных показателей сезонной динамики NDVI позволяет классифицировать изученные пробные площади по состоянию деревьев.
ВВЕДЕНИЕ
Контроль и мониторинг состояния древесных насаждений в лесах Сибири крайне затруднителен в связи с их гигантскими площадями. К тому же оценки по визуальным признакам (состоянию кроны, повреждениям стволов) появляются на тех этапах патологических процессов, когда управление состоянием этих насаждений становится практически невозможным. Для оценки состояния насаждений на обширных таежных территориях единственно возможным методом мониторинга является дистанционное зондирование. Однако неясно, какие дистанционные характеристики можно использовать для оценки состояния насаждений и насколько заблаговременно эти показатели могут сигнализировать об изменении состояния древесных растений. В настоящей работе рассмотрены возможности использования вегатационного индекса (Tucker, 1979) для оценки состояния древесных насаждений на территории Иркутской области, пораженных бактериальной водянкой (Воронин и др., 2013; Воронин, 2018).
Всплеск бактериальных болезней хвойных и лиственных пород произошéл в последние десятилетия. При этом неблагополучное состояние темнохвойных лесов Прибайкалья отмечается уже по крайней мере в течение последних сорока лет (Воронин, Соков, 2005; Плешанов, Морозова, 2009). Ухудшение состояния деревьев связывали с воздействием поллютантов, массовых грибных эпифитотий и размножением насекомых-дендрофагов (Плешанов, Морозова, 2009). В последние же годы картина еще более усложнилась в связи с обнаружением в районах Прибайкалья больших очагов бактериозов хвойных (Воронин и др., 2013; Воронин, 2018). Бактериальные болезни лесов, получившие широкое распространение в последние десятилетия, с большой долей вероятности вызваны климатическими изменениями (Краснобаев, Воронин, 2011).
Бактериальная водянка хвойных пород, поражающая, в том числе и кедр, представляет собой ранее несвойственное для Сибири заболевание. Впервые гибель лесов предположительно от этой болезни обнаружена в 2012 г. на стыке трех муниципальных районов, в Усольском, Слюдянском и Шелеховском лесничествах Иркутской области, относящихся к Байкальской природной территории. Симптомами бактериальной водянки хвойных являются ослабление и усыхание деревьев, поперечные и продольные трещины в коре и активное смолотечение из них, наличие на поперечном срезе древесины ствола “мокрого ядра”, а у сильно ослабленных и недавно усохших деревьев – характерного “темного водослоя” (Щербин–Парфененко, 1963). Очень часто первым внешне заметным признаком болезни дерева является изменение окраски хвои: вначале она желтеет, обычно начиная с концов, а затем довольно быстро приобретает оранжево-красную или розовато-красную окраску, засыхает, но сразу обычно не осыпается. Это может произойти в любой части кроны – в вершине, сбоку, снизу, в середине кроны, нередко на отдельных мутовках и даже на отдельной ветви, а в некоторых случаях только на концах ветвей, иногда по всей кроне сразу. Для контроля заболеваемости лесов крайне важно своевременно выделить лесные территории, на которых происходят изменения состояния деревьев. В настоящей работе рассмотрена возможность использования для мониторинга состояния лесных насаждений методов дистанционного зондирования.
Вегетационные индексы широко используются для изучения состояния и комплексных изменений растительного покрова на различных пространственных масштабах.
В настоящей работе для классификации растительных сообществ с помощью дистанционных методов использовался нормализованный разностный вегетационный индекс растительности (NDVI), основанный на разнице отражения красного и ближнего инфракрасного излучения, полученного спутниковыми датчиками (Tucker, 1979).
Вегетационный индекс связан с продуктивностью и биофизическими свойствами экосистемы (Liu et al., 2017; Barr et al., 2009; Rechid et al., 2009; Richardson et al., 2010) и представляет собой спектральный индикатор фотосинтеза и интенсивности метаболизма растительности (Bayarjargal et al., 2006; Cunha, Richter, 2014; Ma et al., 2013). При изучении динамики вегетационных индексов растительного покрова рассматриваются как внутригодовые, так и межгодовые изменения, связанные с изменчивостью климата (Jacquin et al., 2010; Tottrup, Rasmussen, 2004) или с естественными или антропогенными изменениями растительного покрова (de Beurs, Henebry, 2005; Verbesselt et al., 2009).
Показатели NDVI успешно используются для различных оценок состояния и изменений в растительном покрове, поскольку значения NDVI связаны с фотосинтетически активным излучением. Дистанционное зондирование широко используется для картирования пространственной динамики вспышек насекомых (Verbesselt et al., 2012; Liang et al., 2014; Senf et al., 2017). Однако основной целью таких исследований является оценка ущерба, нанесенного вспышкой, и расчет площади потери. Попытки использовать данные дистанционного зондирования для оценки состояния (и, самое главное, устойчивости) лесных насаждений к внешним воздействиям не приводят к успеху (Spruce et al., 2011; Thayn, 2013; Olsson et al., 2016). Разработка таких методов необходима, так как важно определить районы будущих вспышек насекомых или воздействия болезней, по крайней мере, за один или два сезона до начала повреждения. Такая информация может улучшить прогнозы будущих воздействий и позволит заблаговременно разработать эффективные профилактические меры по снижению потерь в насаждениях.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводились в зоне взаимопроникновения растительных сообществ Хамар-Дабанской гольцово-горно-таежной провинции Южносибирской горно-таежной области и Байкальской озерно-котловинной провинции Байкало-Джугджурской гольцово-горно-таежной области (Белов и др., 2002). Эдификаторами лесов в районе выступают кедр сибирский (Pinus sibirica), пихта сибирская (Abies sibirica) и ель обыкновенная (Picea obovata), при участии сосны обыкновенной (Pinus sylvestris) и лиственницы сибирской (Larix sibirica) (Растительность …, 1988).
Исследовались деревья в возрасте 100–120 лет на контрольных пробных площадях Выдриной (51°28′16.1″ с. ш., 104°51′21.6″ в. д.), и Танхой (51°32′35.47″ с. ш., 105°7′9.68″ в. д.) размером 50 × 50 м, представлявших собой кедрач чернично-зеленомошный с породным составом 10К + П Б, средней высотой около 20 м и средним диаметром кедра в насаждении – 27 см. Кроме того, изучалось состояние деревьев с повреждениями на двух площадях тех же размеров. Насаждение на пробной площади Бабушкин (51°29′53.1″ с. ш., 105°59′09.5″ в. д.) характеризуется как кедрач чернично-зеленомошный в возрасте от 100 до 120 лет с породным составом 9К1П, средней высотой 18 м и средним диаметром кедра в насаждении 20.5 см. Насаждение на пробной площади Култук (51°46′06.6″ с. ш., 103°37′16.6″ в. д.) размером 50 × × 50 м представляет собой пихтарник лиственнично-кедровый багульниково-зеленомошный с породным составом 9Л1К + Б, средней высотой около 17 м и средним диаметром кедра в насаждении 16.5 см.
Показатель NDVI вычисляется по стандартной формуле:
где NIR и RED – нормализованные значения интенсивности отражения в ближнем инфракрасном и красном диапазонах спектра для данной точки земной поверхности.Территория вокруг пробных площадей была обследована с помощью квадрокоптера. Установлено, что характеристики насаждений вблизи пробных площадей такие же, как и на пробных площадях, после чего показатели NDVI рассчитывались для пикселей 250 × 250 м с центром на пробной площади.
Показатели были рассчитаны с помощью продукта MYD09Q1 измерительного комплекса MODIS (Moderate-resolution Imaging Spectroradiometer), установленного на международном научно-исследовательском спутникe AQUA (EOS PM-1). Продукт MYD09Q1 включает в себя два спектральных значения для пикселя размером 250 × 250 метров. Для каждого пикселя формируется композит за 8-дневный период по критерию максимального качества с учетом атмосферных условий. Данные получены с ресурса The Earth Observing System Data and Information System EOSDIS (earthdata.nasa.gov). Значения NDVI представлены за период с 2003 по 2017 г. Типичный сезонный временной ряд NDVI приведен на рис. 1.
Рис. 1.
Типичный временной ряд NDVI(n) на пробной площади Выдринной в 2003 г. (n – номер дня в течение года): 1 – сезонный ряд NDVI, 2 – фильтрованный ряд 1, 3 – ряд 2, обрезанный по критическому значению NDVIFкрит = NDVIFmin + 0.2(NDVImax – ‒ NDVImin), 4 – параболическая аппроксимация ряд-а 3; 5 – критическое значение NDVI(n).
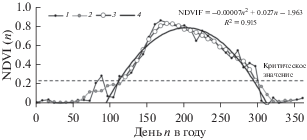
Как видно из рис. 1, кривая сезонной динамики NDVI(n) имеет сложную форму, и для упрощения ее описания предложена следующая процедура “свертки” сезонного временного ряда значений NDVI(i) в год i (Ковалев и др., 2019):
1. Определялись максимальное NDVI(i)max и минимальное NDVI(i)min значения NDVI для сезона i.
2. Временной ряд NDVI(n), в интервале значений от n(i)0 до n(i)f подвергался фильтрации высокочастотным скользящим фильтром по пяти точкам:
3. Определялось критическое значение ND VIF(i)крит за сезон: NDVIF(i)крит = NDVIFmin + + 0.20 (NDVIF(i)max – NDVIF(i)min).
4. Определялись значение n(i)0 – начальная временная точка, до которой все значения NDVIF текущего года ниже значения NDVIF(i)крит, и значение n(i)f– конечная временная точка, после которой все значения NDVIF текущего года ниже значения NDVIF(i)крит.
5. Полученный фильтрованный ряд {NDVIF(n, i)} аппроксимировался уравнением параболы ${\text{NDVIF}}(n,i)$ = $a(i){{n}^{2}} + b(i)n + c(i)$ с помощью нелинейного метода наименьших квадратов, в результате сезонная динамика NDVIF(n) в диапазоне значений (n0, nf) характеризовалась тремя параметрами a(i), b(i), c(i);
6. Вместо параметров a(i), b(i), c(i) использовались две новых переменных, определяемых по уравнению NDVIF(n, i) сезонной динамики NDVIF:
– максимальное сезонное значение NDVIF(i)max, находившееся стандартным способом из уравнения $\frac{{d{\text{NDVIF}}(n,i)}}{{dn}}$ = $2a(i)n + b(i) = 0;$ ${{n}_{{\max }}} = \frac{{ - b(i)}}{{2a(i)}};$ ${\text{NDVIF}}{{(i)}_{{\max }}}$ = $a(i)n_{{\max }}^{2} + b(i){{n}_{{\max }}} + c(i);$
– скорость $\frac{{d{\text{NDVIF}}(i)}}{{dn}}$ = $2a(i){{n}_{{{1 \mathord{\left/ {\vphantom {1 2}} \right. \kern-0em} 2}}}} + b(i)$ нарастания NDVIF в точке $n{{(i)}_{{{1 \mathord{\left/ {\vphantom {1 2}} \right. \kern-0em} 2}}}}$ = $\frac{{(n{{{(i)}}_{{\max }}} - n{{{(i)}}_{0}})}}{2} + n{{(i)}_{0}},$ расположенной в середине интервала между точками n(i)0 и n(i)max. Эта характеристика примерно соответствует дате начала распускания листьев (Суховольский и др., 2017).
Таким образом, для классификации растительных сообществ предложено использовать два “интегральных” показателя фенологической динамики растительных сообществ NDVIF(i)max и dNDVIF/dn. Использование этих фенологических маркеров вегетационной динамики растительных сообществ позволяет уменьшить число переменных, используемых для анализа и классификации.
Для сопоставления данных дистанционного зондирования с наземными данными на исследованных пробных площадях проводился перечет деревьев с использованием визуальной шкалы категорий состояния деревьев (табл. 1)
Таблица 1.
Визуальная шкала категорий состояния деревьев
| Характеристики деревьев | Категории состояния | |||||
|---|---|---|---|---|---|---|
| 1 – без признаков ослабления | 2 – ослабленное | 3 – сильно ослабленное | 4 – усыхающее | 5 – сухостой текущего года (свежий) | 6 – сухостой прошлых лет (старый) | |
| Дефолиация (опадение листьев/хвои) | 0–5% | 10–25% | 25–60% | 60–100% | 100% | 100% |
| Состояние кроны | Крона густая, симметричная, прирост нормальный | Крона слабо ажурная, прирост немного уменьшен | Крона ажурная, прирост заметно уменьшен | Крона изрежена, часто ненормального цвета. | Хвоя/листья усохли, ветви засыхают | Хвоя/листья и мелкие ветви опали |
| Состояние ствола | Нормальное | Нормальное | Повреждения коры ветвей и стволов стволовыми вредителями | Заметные повреждения ствола вредителями, наличие трутовых грибов | Кора начинает отслаиваться, грибные мицелии под корой | Кора отслаивается на большой площади, вылетные отверстия насекомых |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ.
В таблице 2 приведены результаты наземных учетов состояния деревьев на исследованных пробных площадях, проведенных в августе 2018 г.
Таблица 2.
Распределение деревьев по категориям состояния на обследованных пробных площадях
| Пробная площадь | Категория состояния* | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Бабушкин | 0 | 0 | 0.276 | 0.414 | 0.310 |
| Култук | 0 | 0.176 | 0.353 | 0.235 | 0.235 |
| Выдринная | 1 | 0 | 0 | 0 | 0 |
| Танхой | 1 | 0 | 0 | 0 | 0 |
* Подробнее категории см. в табл. 1.
Как видно из табл. 2, все деревья на пробных площадях Выдринная и Танхой относятся в категории 1, и на них отсутствуют видимые признаки повреждений. На пробной площади Култук примерно 20% деревьев принадлежит к категории 2, остальные деревья – к категориям 3, 4 и 5. Большннство (>70%) деревьев на пробных площадях Бабушкин и Култук можно отнести к погибшим (категории 4 и 5).
На рисунке 2 приведены сезонные показатели дистанционного зондирования для изученных пробных площадей в течение 2003–2017 гг. Как видно из рис. 2, дистанционные характеристики в течение 2003–2017 гг. на пробных площадях с поврежденными и неповрежденными деревьями существенно различаются. В таблице 3 приведена классификационная матрица дискриминантного анализа для неповрежденных (пробные площади 1 (Выдринная) и 2 (Танхой)) и поврежденных (площади 3 (Бабушкин) и 4 (Култук)) насаждений.
Рис. 2.
Значения дистанционных показателей N-DVImax и d(NDVI)/dt на пробных площадях в 2003–2017 г. (1 – Выдринная, 2 – Танхой, 3 – Бабушкин, 4 – Култук).
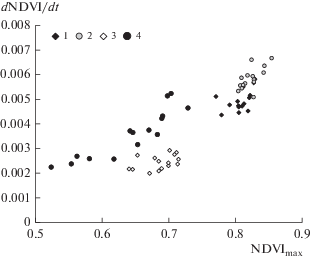
Таблица 3.
Классификационная матрица для классификации четырех пробных площадей в течение 15 лет (2003–2017 гг.) по дистанционным данным
| Пробные площади по наземным оценкам | Процент точно классифированных данных | Пробные площади по дистанционным оценкам | Всего лет | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||
| 1 | 73.3 | 11 | 4 | 0 | 0 | 15 |
| 2 | 93.3 | 1 | 14 | 0 | 0 | 15 |
| 3 | 100.0 | 0 | 0 | 15 | 0 | 15 |
| 4 | 100.0 | 0 | 0 | 0 | 15 | 15 |
| Всего | 91.7 | 12 | 18 | 15 | 15 | 60 |
Из классификационной матрицы для 2003–2017 гг. следует, что все пробные площади практически безошибочно классифицируются по данным дистанционного зондирования (с некоторыми ошибками классифицируются неповрежденные пробные площади 1 и 2) (табл. 4). Качество дискриминации оценивалось по значению λ-критерия Вилкса. Значение λ = 0.036 в табл.4 указывает на хорошую дискриминацию данных. Таким образом, возникает возможность оценки состояния насаждений по дистанционным показателям.
Таблица 4.
Статистическая достоверность (по критерию λ-Вилкса) классификации пробных площадей в течение 2003–2009 гг. по дистанционным данным
| переменные | Значение λ-Вилкса | F-критерий | Уровень значимости F-критерия |
|---|---|---|---|
| Всего | 0.036 | 77.65 | 0 |
| NDVImax | 0.167 | 65.405 | 0.000 |
| dNDVI/dt | 0.163 | 63.643 | 0.000 |
Как можно будет использовать полученные результаты для мониторинга состояния насаждений только по показателям NDVI? Для оценки состояния насаждений, для которых не проводились наземные, а лишь аэровизуальные обследования, был выбрана трансекта, начинающаяся вблизи пробной площади 3 (Бабушкин) (точка М01) направлением к побережью Байкала. Вдоль выбранной трансекты через каждые 1000 м рассчитывались показатели NDVI за период 2003–2017 гг.
Для точек М01–М18 на трансекте для периода с 2003 по 2017 гг. были вычислены средние многолетние значения сезонных параметров NDVImax и d(NDVI)/dt. На рисунке 3 приведены эти значения, а также средние многолетние показатели NDVImax и d(NDVI/dt для пробных площадей 3 (Бабушкин) и 1 (Выдринная).
Рис. 3.
Средние многолетние значения сезонных показателей NDVImax и dNDVI/dt для точек M01–M07 (кластер 1) и M08–M18 (кластер 2) на трансекте, а также для пробной площади 1 (Выдринная, точка 3) и пробной площади 4 (Бабушкин, точка 4).
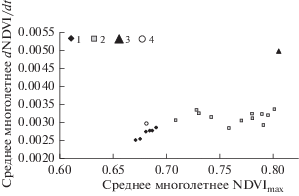
Как видно из рис. 3, все точки на трансекте можно разделить на две группы: точки М01–М07, близкие по своим средним многолетним показателям к средним многолетним показателям поврежденного насаждения на пробной площади 3 (Бабушкин), и точки М08-М018 со средними многолетними показателями, близкими к средним многолетним показателям для неповрежденного насаждения на пробной площади 1 (Выдринная).
ЗАКЛЮЧЕНИЕ
Анализ возможностей использования показателей сезонной динамики вегетационного индекса для оценки состояния лесных насаждений показал, что дистанционные характеристики насаждений, классифицируемых в 2018 г. по наземным оценкам, значимо различаются в предыдущие 15 лет, с 2003 по 2017 гг. (более ранние дистанционные данные отсутствуют). Так как индикаторные параметры NDVImax и d(NDVI)/dt оцениваются с использованием кривой сезонной динамики NDVI, оценить состояние насаждения по дистанционным данным возможно только в конце сезона. При этом данные по сезонной динамике NDVI для всей территории лесов Сибири находятся в открытом доступе, что открывает возможности дистанционного мониторинга обширных лесных территорий. Для дальнейшего анализа возможностей использования дистанционных показателей для построения шкал состояния насаждений предполагается продолжить сопряженный анализ наземных и дистанционных показателей состояния.
Список литературы
Белов А.В., Лямкин В.Ф., Соколова Л.П. Картографическое изучение биоты. Иркутск: Облмашинформ, 2002. 160 с.
Воронин В.И. Бактериальная водянка хвойных в байкальских лесах: причины возникновения и риск эпифитотии // Механизмы устойчивости растений и микроорганизмов к неблагоприятным условиям среды. Сб. матер. Годичного собрания Общества физиологов растений России, Всеросс. науч. конф. с междунар. участием и школы молодых ученых. Иркутск: Изд-во Института географии им. В.Б. Сочавы СО РАН. 2018. С. 9–12.
Воронин В.И., Соков М.К. Влияние сероорганических компонентов атмосферных выбросов на пихту сибирскую // Лесоведение. 2005. № 2. С. 62–64.
Воронин В.И., Морозова Т.И., Ставников Д.Ю., Нечесов И.А., Осколков В.А., Буянтуев В.А., Михайлов Ю.З., Говорин Я.В., Середкин А.Д., Шуварков М.А. Бактериальное повреждение кедровых лесов Прибайкалья // Лесн. хоз-во. 2013. № 3. С. 39–41.
Ковалев А.В., Воронин В.И., Суховольский В.Г. Оценка повреждений кедровых насаждений южного берега оз. Байкал на основе многолетних спутниковых наблюдений MODIS/AQUA // Аэрокосмические методы и геоинформационные технологии в лесоведении, лесном хозяйстве и экологии: Доклады VII Всерос. конф. (Москва, 22–24 апреля 2019 г.). М.: ЦЭПЛ РАН, 2019. С. 63–64.
Краснобаев В.А., Воронин В.И. Аномальные оттепели как одна из причин повреждений кроны молодых хвойных деревьев в южном Прибайкалье // География и природные ресурсы. 2011. № 2. С. 75–78.
Плешанов А.С., Морозова Т.И. Микромицеты пихты сибирской и атмосферное загрязнение лесов. Новосибирск: Гео, 2009. 115 с.
Растительность хребта Хамар-Дабан. Новосибирск: Наука. 1988. 113 с.
Суховольский В.Г., Иванова Ю.Д., Овчинникова Т.М., Ботвич И.Ю. Моделирование фенодинамики листопадных древесных пород // Лесоведение. 2017. № 4. С. 293–302.
Щербин-Парфененко А.Л. Бактериальные заболевания лесных пород. М.: Гослесбумиздат, 1963. 148 с.
Barr A., Black T.A., McCaughey H. Climatic and phenological controls of the carbon and energy balances of three contrasting boreal forest ecosystems in western Canada // Phenology of Ecosystem Processes: Applications in Global Change Research. 2009. P. 3–34.
Bayarjargal Y., Karnieli A., Bayasgalan M., Khudulmur S., Gandush C., Tucker C.J. A comparative study of NOAA-AVHRR derived drought indices using change vector analysis // International Journal of Remote Sensing. 2006. V. 105. № 1. P. 9–22.
Cunha M., Richter C. A time-frequency analysis on the impact of climate variability with focus on semi-natural montane grassland meadows // IEEE Transection Geoscience and Remote Sensing. 2014. V. 52. № 10. P. 6156–6164.
de Beurs K.M., Henebry G.M. Land surface phenology and temperature variation in the International Geosphere-Biosphere Program high-latitude transects // Global Change Biology. 2005. V. 11. № 5. P. 779–790.
earthdata.nasa.gov
Jacquin A., Sheeren D., Lacombe J.P. Vegetation cover degradation assessment in Madagascar savanna based on trend analysis of MODIS NDVI time series // International J. Applied Earth Observation and Geoinformation. 2010. V. 12. P. S3–S10.
Liang L., Chen Y., Hawbaker T., Zhu Z., Gong P. Mapping mountain pine beetle mortality through growth trend analysis of time-series landsat data // Remote Sensing. 2014. V. 6. P. 5696–5716.
Liu Yan, Hill M.J., Zhang Xiaoyang, Wang Zhuosen, Richardson A.D., Hufkens K., Filippa G., Baldocchi D.D., Ma Siyan, Verfaillie J., Schaaf C.B. Using data from Landsat, MODIS, VIIRS and Pheno Cams to monitor the phenology of California oak/grass savanna and open grassland across spatial scales // Agricultural and Forest Meteorology. 2017. V. 237–238. P. 311–325.
Ma X., Huete A., Yu Q., Coupe N.R., Davies K., Broich M., Ratana P., Beringer J., Hutley L.B., Cleverly J., Boulain N., Eamus D. Spatial patterns and temporal dynamics in savanna vegetation phenology across the North Australian Tropical Transect // Remote Sensing of Environment. 2013. V. 139. P. 97–115.
Olsson P.O., Lindstrom J., Eldundh L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI // Remote Sensing of Environment. 2016. V. 181. P. 42–53.
Rechid D., Raddatz T.J., Jacob D. Parameterization of snow-free land surface albedo as a function of vegetation phenology based on MODIS data and applied in climate modelling // Theoretical and Applied Climatology. 2009. V. 95. P. 245–255.
Richardson A.D., Black A.T., Ciais P., Delbart N., Friedl M.A., Gobron N., Hollinger D.Y., Kutsch W.L., Longdoz B., Luyssaert S., Migliavacca M., Montagnani L., Munger W.J., Moors E., Piao S., Rebmann C., Reichstein M., Saigusa N., Tomelleri E., Vargas R., Varlagin A. Influence of springand autumn phenological transitions on forest ecosystem productivity. // Philosophical Transactions of the Royal Society B: Biological Sciences. 2010. V. 365. P. 3227–3246.
Senf C., Seidl R., Hostert P. Remote sensing of forest insect disturbances: Current state and future directions // International J. Applied Earth Observation and Geoinformation. 2017. V. 60. P. 49–60.
Spruce J.P., Sader S., Ryan R.E., Smoot J., Kuper P., Ross K., Prados D., Russell J., Gasser G., McKellip R. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks // Remote Sensing of Environment. 2011. V. 115. P. 427–437.
Thayn J.B. Using a remotely sensed optimized Disturbance Index to detect insect defoliation in the Apostle Islands, Wisconsin, USA // Remote Sensing of Environment. 2013. V. 136. P. 210–217.
Tottrup C., Rasmussen M.S. Mapping long-term changes in savannah crop productivity in Senegal through trend analysis of time series of remote sensing data // Agriculture, Ecosystems and Environment. 2004. V. 103. № 3. P. 545–560.
Tucker C.J. Red and photographic infrared linear combinations for monitoring vegetation // Remote Sensing of Environment. 1979. V. 8. P. 127–150.
Verbesselt J., Zeileis A., Herold M. Near real-time disturbance detection using satellite image time series // Remote Sensing of Environment. 2012. V. 123. P. 98–108.
Verbesselt J., Robinson A., Stone C., Culvenor D. Forecasting tree mortality using change metrics derived from MODIS satellite data // Forest Ecology and Management. 2009. V. 258. №.7. P. 1166–1173.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение