

Лесоведение, 2021, № 1, стр. 28-41
Влияние пространственной структуры древостоев сосны на отпад и диаметр деревьев
М. В. Рогозин *
Пермский государственный национальный исследовательский университет
614990 Пермь, ул. Генкеля, 4, Россия
* E-mail: rog-mikhail@yandex.ru
Поступила в редакцию 05.03.2019
После доработки 12.08.2019
Принята к публикации 07.10.2020
Аннотация
С целью выяснить силу влияния густоты микро- и макроценозов на отпад и размер деревьев в количественном выражении изучено 184-летнее насаждение сосны обыкновенной I класса бонитета с полнотой 0.91–0.95. На площади 2.16 га живые и естественным образом отпавшие примерно за 60 лет деревья нанесли на план с точностью ±10–35 см, далее план оцифровали в программе ArcMap и механически разбили на 216 площадок (микроценозов) размером 0.01 га. Следы пней позволили установить густоту на них в возрасте около 120 лет, а также в возрасте 170 лет – до ветровала 14 лет назад, когда пострадало 7% деревьев. Силу влияния фактора густоты площадок на диаметр дерева рассчитали как среднее значение из трех показателей: детерминации по квадрату корреляции, аппроксимации тренда по линейной связи и среднего из аппроксимаций трендов по пяти типам полиномов. Установлено, что на уровне миди- и макроценозов на пробных площадях размером по 0.12 и 0.24 га естественные флуктуации их густоты в 120–170 лет повлияли на средний диаметр сосны в 184 года с силой в среднем 47%. В то же время густота микроценозов на площадках 10 × 10 м в тех же возрастах повлияла на средний диаметр сосны с силой от 8.8 до 11.8% (в среднем 10.1%), что в 4.7 раза слабее, чем действие густоты на уровне макроценоза. Этот факт позволяет предположить, что в микроценозах внутривидовая конкуренция между деревьями резко слабеет и меняется на взаимное партнерство. Кроме того, густота на площадках 0.01 га в 120 лет повлияла на отпад деревьев на них к 184 годам с силой всего лишь 4.7%. В остальных 95.3% случаев на отпад повлияли неустановленные факторы.
Густота признана одним из мощных биоценотических факторов, влияющих на рост древостоев, и публикаций на эту тему достаточно много (Рогозин, Разин, 2015). Однако сила ее влияния на развитие деревьев в микроценозах до сих пор точно не известна. Между тем в лесоводстве регуляцию густоты насаждения рубками осуществляют именно через микроценозы, вырубая в них часть живых деревьев. В горизонтальной структуре древостоев они играют главную роль, и для их изучения применяют целый спектр методов на основе изучения площадей питания деревьев (Мартынов, 1976; Борисов и др., 2014; Вайс, 2014) и достаточно сложные пространственные модели (Нагимов, 1999; Грабарник, 2010; Усольцев, 2013), в том числе с выяснением динамики прироста в течение сезона (Горячев, 1999) и анализом структуры биогрупп (Рогозин, 2018б).
Согласно анализу данных методов (Рогозин, 2018а), если в них принималось, что чем больше диаметр деревьев, тем больше приходящиеся на них площади питания, тогда использовали три зависимых переменных величины: текущий прирост дерева по диаметру (величина 1); диаметр дерева (величина 2) и площадь питания, определяемая делением расстояний от соседей пропорционально их диаметрам (величина 3). Так как первая и третья величины зависят от второй (являются ее частью при расчетах), то первая величина будет зависима от третьей. Но тогда площадь питания оказывается непригодна как фактор для выяснения силы ее влияния на размеры дерева. Поэтому независимой переменной величиной в площади питания дерева будет, по нашему мнению, ее значение, определяемое по полигонам питания с простым делением расстояний между соседями. Иными словами, следует выяснить, как дерево распорядилось доставшейся ему ранее площадью питания, причем безотносительно к размерам соседей, которые признаются все потенциально равными в ее освоении в прошлом. В этом отношении более адекватным является изучение отпада и структур фитоценоза при детальном картировании всех деревьев, а не только их моделей (Kenkel, 1988; Grenfell et al., 2011; Omelko et al., 2018).
По-видимому, следует выяснить влияние густоты фитоценоза на онтогенез деревьев на двух уровнях: всего древостоя (в макроценозах) и индивидуальных взаимодействий между деревьями (в микроценозах). Определение количественных значений влияния густоты на этих уровнях позволит понять, на какой из них следует воздействовать для улучшения роста – на общую густоту или на густоту локальную. Такая постановка вопроса вытекает из закона и моделей развития древостоев в зависимости от их начальной густоты, где ее регуляция эффективна только в фазе прогресса, т.е. в период повышения прироста. Этот период достаточно короткий и, например, в продуктивных ельниках продолжается до 45–50 лет, причем в изначально густых древостоях он заканчивается уже в 15–20 лет, после чего наступает фаза регресса, когда регуляция густоты рубками уже слабо влияет на развитие древостоя (Рогозин, Разин, 2015).
Между тем, действующие правила ухода за лесом (Правила …, 2017) предписывают регулировать текущую густоту насаждений вплоть до приспевающего возраста. Процедура ее регуляции решается через разреживание густых мест, т.е. через воздействие на микроценозы. Влияние их густоты проявится в конечном итоге как результат всего предшествующего развития деревьев в них только в спелом возрасте. Поэтому изучение структуры старовозрастных лесов оказывается весьма актуально для понимания того, как нам сохранить леса в устойчивом состоянии на возможно более длительное время. В этом случае особенности их структуры позволят взять ее как образец, к которому следует стремиться. Структуру малонарушенных бореальных лесов изучают с целью определения закономерности ее формирования (Linder et al., 1978; Kuuluvainen et al., 2014) и выяснения динамики послепожарных сукцессий в течение нескольких сотен лет (Горшков, и др., 2005; Ставрова и др., 2012, 2015), а также иных сильных воздействий на сложные по составу насаждения, включающие до пяти пород-доминантов. Сукцессии в таких лесах обусловлены мелкими и средними возмущениями их структуры с образованием прогалин, их заполнением подростом и выходом пород в верхний ярус или их подчинением другим породам, в зависимости от сочетания многих факторов (Omelko et al., 2018).
Для использования моделей даже в простых по составу насаждениях нужно знать силу (или долю) влияния факторов, образующих тот или иной тип их структуры. Точных сведений об этом крайне мало. Все прежние теоретические положения лесоводства разрабатывались на логических моделях, а также на основе опытных данных – при выращивании насаждений с использованием разреживаний (Сеннов, 1999) или на основе изучения культур разной густоты (Плантационное …, 2007).
В последнее время появились модели с включением до 20 параметров (Грабарник, 2010; Грабарник, Секретенко, 2015). Возник интерес к динамике лесных прогалин (концепция “gap-динамики”), где изучают появление подроста, образование и распад куртин в многоярусных насаждениях (Карев, 1999; Кислов и др., 2015). Для их изучения подбирают насаждения по возрастам и далее конструируют их состояние в возрастной динамике (Кузьмичев, 2013; Колобов, 2014), с использованием моделей стохастических процессов (Секретенко, 1984). Однако увлечение такими моделями заводит объяснения по их действию в некий логический тупик. Типы эти известны давно, но до понимания, почему они образуются в древостое, дело не доходит. Неясно, какие факторы формируют случайный, а какие кластерный типы поселения деревьев, а в более широком плане – поселение растений вообще. Происходит лишь фиксация этих структур, где фактором их образования принимается чаще всего конкуренция за свет (Бузыкин и др., 1985; Усольцев, 2013).
Обычно выделяют три типа размещения особей в фитоценозе: случайное, групповое и равномерное. Считается, что групповое размещение отражает повышенную напряженность конкуренции и преобладает над случайным в большинстве ценопопуляций. Мозаичность структуры свойственна всем лесным фитоценозам, в том числе лесным культурам с их первоначальным регулярным размещением (Усольцев, 2013). Древостой состоит из биогрупп – территориально сближенных деревьев, где проявляется так называемый “эффект группы” (эффект кооперации), который был сформулирован как принцип агрегации особей Олли (Alleе, 1931; Одум, 1975). Согласно этому принципу у особей, объединенных в группу, повышается устойчивость к неблагоприятным факторам, поскольку в группе поверхность их соприкосновения со средой меньше и группа способна изменять микросреду в благоприятном для себя направлении.
Так, исследования В.А. Усольцева (2013) показали, что в естественных сосняках и культурах в возрасте 20 лет преобладает групповое распределение деревьев, а к 40-летнему возрасту естественные сосняки приобретают уже случайное распределение деревьев. В результате индексы конкуренции, полученные 10 способами, объясняли изменчивость фитомассы и прироста деревьев соответственно, на 5 и 11%, а в 20-летних культурах эти значения были равны всего лишь 0.2 и 3%. Радиальный прирост модельных деревьев индексы конкуренции объясняли в естественных молодняках на 36%, а в культурах – на 11%.
В следующей работе В. А. Усольцева с соавт. (2018) было констатировано, что буквально единичные работы посвящаются теме партнерства между деревьями. Например, Ю.П. Демаков (2007) отмечает наличие в древостоях сосны сухих деревьев с живым комлем, регулярным приростом по диаметру и срастанием корневых систем, которые говорят о том, что насаждение – это единая система, где распространена взаимопомощь. А.И. Колтунова (2013) выяснила, что в сосняках в 19–60 лет на 1 га насчитывается до 1 тыс. деревьев, сросшихся корнями, и с возрастом их доля возрастает. Эти феномены расширяют адаптацию вида и показывают, что “…организация сообщества древесных растений построена на единстве процессов конкуренции и кооперации” (Усольцев и др., 2018, с. 238). Факты эти были известны уже давно (Крамер, Козловский, 1963; Рахтеенко, 1963; Калинин, 1983), однако они не поколебали идею верховенства конкуренции между деревьями и воспринимались скорее как редкие феномены.
Рассматривая далее внутри- и межвидовую конкуренцию в историческом аспекте В.А. Усольцев с соавт. (2018) отмечает, что еще в конце ХIХ в. в биологии начала доминировать теория отбора Ч. Дарвина и грубая модель эволюции была навязана всему миру воинствующими атеистами в политических целях (Баландин, 2010). Дарвинизм и поныне является отправной точкой любого биологического исследования по умолчанию (Костерин, 2007). Поэтому с учетом новейших представлений об эволюции исследования структуры древостоев должны включать изучение и конкуренции, и партнерства между деревьями, не отдавая предпочтения ни одному из этих столь различающихся концептуальных подходов.
Для выявления распределения деревьев в древостоях по различным биологическим параметрам – возрасту, размерам, текущей густоте, плотности заселения и другим, а также связей между ними в настоящее время изучают их структуру на некотором множестве небольших по размеру пробных площадей, например, 5 × 5 м в молодых и 10 × 10 м в спелых древостоях, число которых должно обеспечивать необходимую точность их статистического анализа (Linder at al., 1997; Grenfell et al., 2011; Ставрова и др., 2012; Omelko et al., 2018).
Таким образом, микроценозы и биогруппы как первичные структуры лесного сообщества, остаются изученными недостаточно. Возможно, конкуренция между деревьями в них ослабевает и уступает место партнерству. Поэтому мы выдвинули гипотезу ослабления в них конкуренции между деревьями в сравнении с ее действием на уровне древостоя в целом, которая проявляет себя через влияние локальной и общей густоты стояния деревьев на их размерные показатели.
В связи с этим была поставлена цель выяснить силу влияния густоты микро- и макроценозов на размер деревьев в них с расчетами показателей связи “густота × диаметр дерева”. Различия в уровне связей должны подтвердить или опровергнуть выдвинутую гипотезу. Для достижения этой цели необходимо было, последовательно объединяя условные микроценозы (площадки 10 × 10 м с деревьями на них) вначале перейти к уровню мидиценоза (среднему между микро- и макроценозом), а затем к уровню древостоя, т.е. макроценоза, с пробными площадями размером до 0.20–0.30 га, что возможно на территории насаждения площадью не менее 2 га.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены в старовозрастном насаждении сосны обыкновенной (Рinus sylvestris L.) в районе г. Пермь в кв. 61 Нижне-Курьинского участкового лесничества, выделе 3 площадью 10 га с полнотой 0.8. Данное насаждение выбрали после обследования всех лесов зеленой зоны вокруг г. Пермь в 2015 г. и признали наилучшим. Его структура была не нарушена, в отличие от других насаждений, так как оно не подвергалось выборочным рубкам с прорубкой волоков и вывоз древесины от регулярных санитарных рубок здесь проводили по просветам между деревьями. По нашим наблюдениям, в 2003 г. древостой был пройден ветровалом с вывалом 7% деревьев, которые были вывезены. Возраст определили по кернам у 40 деревьев, и в среднем он составил 184 года, с колебаниями от 173 до 195 лет. Насаждение расположено на надпойменной террасе р. Кама с мощностью песчаных отложений местами до 7–20 м. Выбранную для исследования территорию площадью 2.16 га разделили на две равные части, названные участок “Северный” и участок “Южный”. Проведено картирование участков с нанесением на план на двух листах формата А0 в масштабе 1 : 100. Территория представляет собой пологую песчаную дюну с ее центром вблизи середины территории и понижением рельефа к северо-восточному краю примерно на 3.0 м, и к юго-востоку на 1.5–2.0 м.
В насаждении у деревьев делали отметку на высоте 1.3 м, наносили его номер, измеряли окружность ствола и расчетным путем определяли диаметр с округлением до ±0.1 см. Деревья, живые и сухостойные, а также пни, валеж, подрост ели, подлесок рябины и березы наносили на план в масштабе 1 : 100 с точностью ±10–35 см. Высоты измерялись высотомером “Haglof” c двух-четырех позиций, так как деревья имели наклон вследствие ветровой нагрузки. План был оцифрован в программе “ArcMap-ArcViev” и территория разбита на 216 площадок-микроценозов размером 10 × 10 м (рис. 1).

Такие микроценозы лишены внешней физиономичности; они выхвачены из структуры связей с соседями, но при подобном их выделении исключен субъективный фактор, что немаловажно. Полученные данные были обработаны с использованием стандартных статистических процедур. Влияние густоты на отпад и средний диаметр деревьев определяли вначале в микроценозах 10 × 10 м, затем их объединяли в более крупные площади размером 20 × 20, 30 × 30, 30 × 40, 40 × 60 и 60 × × 60 м, и далее изучали влияние их густоты на средний диаметр деревьев. Определяли показатели корреляции и аппроксимации R2 для линейных и полиномиальных связей, обычно используемых в лесоводственных исследованиях (Маслаков, 1984; Усольцев, 2013; Демаков, 2018). Корреляционный анализ проведен для уровня значимости t0.05, и корреляции оказались достоверны. Так как форма связи не была известна, силу влияния фактора густоты площадок на диаметр дерева рассчитали как среднее значение из трех показателей: детерминации по квадрату корреляции, аппроксимации тренда по линейной связи и среднего значения из показателей аппроксимаций трендов по пяти типам полиномов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Таксационные показатели. Оба участка по наличию по всей их территории кислицы обыкновенной были отнесены к типу леса сосняк кисличниковый, но в кустарничковом ярусе были некоторые отличия. Таксационная характеристика участков следующая.
Участок “Южный” – состав 10С едЕ, возраст 184 г., средняя высота сосны 32.5 ± 0.24 м, средний диаметр 39.1 ± 0.4 см, кустарничковый ярус представлен черникой (70%) и брусникой (30%), класс бонитета 1, абсолютная полнота 42.8 м2 на 1 га, относительная полнота 0.91, запас 614 м3 на 1 га. (Если учесть ветровальные деревья, то полнота и запас могли бы увеличиться на 7.4% при полноте 0.98). Текущая густота 339 шт. на 1 га.
Участок “Северный” имеет тот же состав и возраст, но кустарничковый ярус представлен черникой уже на 95% и брусникой на 5%, что свидетельствует о несколько более влажных условиях. Средняя высота сосны 33.4 ± 0.27 м, средний диаметр 41.2 ± 0.43 см. Густота древостоя здесь меньше на 6%, но полнота выше и достигает 0.95, запас составляет 655 м3 на 1 га. (Если учесть ветровальные деревья, то густота, полнота и запас могли бы увеличиться здесь на 6.5% и составить ныне 700 м3 на 1 га при полноте 1.01). Текущая густота 324 шт. на 1 га.
Таким образом, исследуемая территория площадью 2.16 га характеризовалась следующими средними таксационными показателями: высота 32.8 м, диаметр 41.0 см, полнота 0.93, густота 334 шт. на 1 га и запас 630 м3 на 1 га. На обоих участках единично встречается ель в возрасте 120 лет с диаметрами 20–45 см, образующая невыраженный второй ярус. Подрост ели представлен поколением в возрасте 35 лет с высотой 3–6 м, возникшим после низового пожара и обильного урожая семян ели в 1982 г.
Сравнение показало достоверное увеличение на участке “Северный” среднего диаметра сосны на 5.5% при t = 3.64 > t0.01 = 2.7 и средней высоты на 2.9% при t = 2.60 > t0.05 = 2.0. Данные перечислительной таксации подтвердили предположения, основанные на большей встречаемости здесь черники, что на этом участке условия несколько более благоприятные. Они обусловлены большим увлажнением вследствие северной экспозиции склона примерно в 3° и меньшей на 6% густотой, при которой сформировалось больше крупных деревьев. В свою очередь, оба участка в своих частях также имели некоторые отличия по густоте и увлажнению. В целом же территория представляет собой единый массив с естественными флуктуациями почвенных условий и густоты ценозов.
Показатели густоты микро- миди- и макроценозов. Всего на 216 площадках размером по 0.01 га расположено 720 живых и 54 вываленных 14 лет назад деревьев. Прибавляя вываленные деревья к густоте в настоящее время, была рассчитана густота 14 лет назад, т.е. в возрасте 170 лет. Густоту насаждения нам удалось реконструировать и далее, до возраста примерно 120 лет, по следам полуразложившихся пней, которых насчитывалось 331 шт. на обоих участках. Возраст 120 лет был принят в предположении, что в условиях южной тайги места, где росли ранее отпавшие деревья сосны, можно опознать спустя примерно 60 лет по заметным отличиям в характере напочвенного покрова, а спустя 50 лет они хорошо угадываются по микроповышениям (Стороженко, 2007). Таким образом, реконструкция густоты в прошлое позволила оперировать густотой в возрасте 184, 170 и 120 лет.
В результате обработки данных картирования число деревьев на одной площадке в возрасте 184 и 170 лет колебалось в пределах 0–7 шт., а в возрасте 120 лет – 1–10 шт. Среднее число деревьев на них в возрасте 170 и 120 лет составляло, соответственно, 3.63 и 5.12 шт. Получается, что их густота за 50 лет снизилась на 29%.
Отпад в лесных насаждениях представляют в абсолютных или относительных значениях по классам возраста при составлении таблиц хода роста (Рогозин, Разин, 2015). Для нашего же случая важно было получить его средние величины по классам густоты микроценозов, которых в возрасте 120 лет было 10, от 0–1 шт. деревьев на площадке до 10 шт. По этим классам густоты мы построили регрессионные уравнения, характеризующиеся высокими значениями (0.650 и 0.954) показателей аппроксимации трендов. Следовательно, их можно использовать для моделирования отпада сосны от 120 до 170 лет, в зависимости от густоты микроценозов на площадках 10 × 10 м (рис. 2).
Рис. 2.
Средний отпад сосны на площадках 10 × 10 м в возрасте от 120 до 170 лет: а – в абсолютном выражении (шт.); б – в относительном выражении (%).
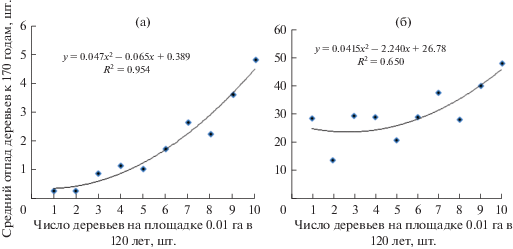
Однако точность прогноза по таким моделям отпада остается не ясной, так как показатели аппроксимации R2 на таких диаграммах, с усреднением данных по вертикальной оси, не показывают их изменчивость. То есть нужно знать весь спектр отпада деревьев, который колеблется здесь в пределах классов густоты от 0 до 100%. В этом случае связь оказывается уже совершенно другой (рис. 3).
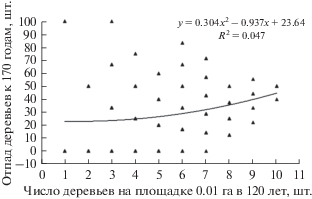
Разброс данных получается настолько большим, что при густоте в 6–7 шт. встречаются площадки как с нулевым отпадом, так и с отпадом 71–83%. При этом даже малая густота, по. 1–3 шт. деревьев на площадке, не гарантировала им сохранность и имел место даже их полный отпад. На это можно возразить, что они могли соседствовать с густыми площадками, и те влияли на процесс отпада. Это вполне возможно, и в этом недостаток микроценозов, выделенных механическим способом; однако недостаток этот преодолевается увеличением объема выборки, и здесь он достаточно большой (216 шт.). Но на диаграмме число точек получилось намного меньше, так как их значения часто совпадали и накладывались одно на другое.
Если взять значения с линии тренда на рис. 3, то при числе деревьев на площадке 1–3 шт. отпад к 170 годам составит в среднем 23%, а при 7–9 шт. – 37%. Таким образом, с повышением густоты микроценоза от 1–3 до 7–9 шт. (в среднем в 4 раза) процент отпада за 50 лет увеличился в среднем лишь в 1.6 раза. Слабое влияние густоты площадок на отпад подтверждает и показатель аппроксимации, равный R2 = 0.047. Это низкий уровень, и сила влияния густоты деревьев на площадке на будущий отпад составляет всего лишь 4.7%. А это значит, что в подавляющем большинстве случаев (95.3%) деревья отмирают в микроценозах в период от 120 до 170 лет по причинам, не связанным с их густотой; вероятно, здесь влияют факторы другого плана, определяемые, например, литологией конкретных мест и флуктуациями почвенной среды геофитоценоза.
На средний диаметр деревьев густота микроценозов в 120 лет также повлияла довольно слабо (рис. 4).
Рис. 4.
Влияние густоты площадок 10 × 10 м в 120 лет на средний диаметр деревьев в 184 г. Прерывистая линия – средний диаметр древостоя.
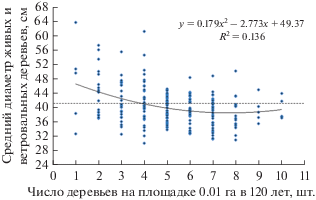
Здесь фактор густоты повлиял на диаметр сосны с силой 13.6% (R2 = 0.136); это показатель аппроксимации для тренда полинома, а для линейного тренда он равен R2 = 0.108 при коэффициенте корреляции r = –0.329. Между тем, показанный на рисунке 4 тренд – только один из множества построенных нами линий связи, которые включали в себя 6 типов; их описание приведено ниже (табл. 1).
Таблица 1.
Влияние густоты на пробных площадях разного размера (в микро-, миди и макроценозах) в разном возрасте на средний диаметр сосны в возрасте 184 г.
| №№ | Размер пробной площади, га | Число пробных площадей, шт. | Корреляция “густота × × диаметр дерева” (r) | Показатель аппроксимации линий тренда R2 (сила влияния фактора густоты) | Среднее влияние густоты: (r2 + + R2 + $R_{{{\text{cp}}}}^{2}$) : 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| линейная cвязь R2 | полиномы, степень | среднее из полиномов ($R_{{{\text{cp}}}}^{2}$) | |||||||||
| 2 | 3 | 4 | 5 | 6 | |||||||
| Микроценозы: | |||||||||||
| в 184 года | |||||||||||
| 1 | 0.01 | 216 | –0.304 | 0.078 | 0.089 | 0.094 | 0.096 | 0.096 | 0.102 | 0.095 | 0.088 |
| в 170 лет | |||||||||||
| 2 | 0.01 | 216 | –0.304 | 0.092 | 0.101 | 0.106 | 0.109 | 0.113 | 0.117 | 0.109 | 0.098 |
| в 120 лет | |||||||||||
| 3 | 0.01 | 216 | –0.329 | 0.108 | 0.136 | 0.136 | 0.137 | 0.138 | 0.138 | 0.137 | 0.118 |
| участок “Южный” (более сухой) | |||||||||||
| 4 | 0.01 | 108 | –0.263 | 0.069 | 0.089 | 0.096 | 0.112 | 0.112 | 0.114 | 0.105 | 0.081 |
| в т. ч. более густое место | |||||||||||
| 5 | 0.01 | 54 | (–0.220) | 0.048 | 0.070 | 0.077 | 0.086 | 0.086 | 0.087 | 0.081 | 0.059 |
| участок “Северный” (более влажный) | |||||||||||
| 6 | 0.01 | 108 | –0.346 | 0.133 | 0.177 | 0.191 | 0.219 | 0.239 | 0.240 | 0.213 | 0.155 |
| в т.ч. более редкое место | |||||||||||
| 7 | 0.01 | 54 | –0.346 | 0.119 | 0.180 | 0.223 | 0.249 | 0.300 | 0.303 | 0.251 | 0.163 |
| Мидиценозы в 120 лет | |||||||||||
| 8 | 0.04 | 54 | –0.459 | 0.21 | 0.27 | 0.293 | 0.313 | 0.329 | 0.333 | 0.308 | 0.243 |
| 9 | 0.09 | 24 | 0.59 | 0.348 | 0.454 | 0.509 | 0.568 | 0.632 | 0.64 | 0.561 | 0.419 |
| 10 | 0.12 | 18 | –0.605 | 0.366 | 0.394 | 0.516 | 0.54 | 0.567 | 0.609 | 0.525 | 0.419 |
| Мидиценозы в 170 лет | |||||||||||
| 11 | 0.04 | 54 | –0.542 | 0.293 | 0.294 | 0.369 | 0.394 | 0.408 | 0.408 | 0.375 | 0.321 |
| 12 | 0.09 | 24 | –0.576 | 0.332 | 0.399 | 0.399 | 0.4 | 0.431 | 0.461 | 0.418 | 0.361 |
| 13 | 0.12 | 18 | –0.705 | 0.497 | 0.538 | 0.573 | 0.573 | 0.631 | 0.642 | 0.591 | 0.528 |
| Макроценозы в 170 лет | |||||||||||
| 14 | 0.24 | 9 | –0.650 | 0.422 | 0.445 | 0.495 | 0.565 | 0.585 | 0.677 | 0.553 | 0.466 |
| 15 | 0.36 | 6 | –0.747 | 0.557 | 0.693 | 0.990 | 0.999 | 1.000 | 1.000 | 0.936 | 0.684 |
В этой сложной таблице сведены вместе данные из множества точечных диаграмм, подобных показанной на рис. 4. Для отслеживания изменений в №№ 1–7 в таблице использованы пробные площади размером 10 × 10 м, в №№ 8–13 – пробные площади размером 20 × 20, 30 × 30 и 40 × 30 м, а в №№ 14, 15 – пробные площади по 0.24 и 0.36 га. Поясним, что для анализа данных мы провели еще одно разделение массива сосны, на этот раз на четыре части (участки “Южный” и “Северный” и их половины, которые показаны курсивом). Это сделано с целью проследить, как естественные флуктуации густоты и увлажнения биотопа влияют на показатели связи “густота × диаметр” по четырем частям этого массива. В нижних №№ 8–15 таблицы показано, как меняется эта связь от уровня миди- (0.04–0.12 га) к уровню макроценоза (0.24–0.36 га). Анализ данных этой таблицы показал, что в возрасте 184 года показатели аппроксимации R2 наименьшие: 0.078 для линейной связи и в среднем по пяти полиномам $R_{{{\text{ср}}}}^{{\text{2}}}$ = 0.095. Заметим, что корреляция r = –0.304 при возведении в квадрат дает детерминацию связи “густота×диаметр” r2 = 0.092, что близко к значениям $R_{{{\text{ср}}}}^{{\text{2}}}$. Для общей оценки влияния фактора густоты можно принять среднее из этих трех значений: (0.078 + 0.095 + 0.092) : 3 = = 0.088 или 8.8%, которое приведено в последнем столбце табл. 1.
Так как в возрасте 170 лет в массиве сосны прошел ветровал, мы пролонгировали у вываленных деревьев их диаметры на настоящее время и далее провели их обработку вместе с живыми деревьями. Здесь для линейной связи “густота × диаметр” показатель R2 = 0.092, а в среднем по пяти типам полиномов $R_{{{\text{ср}}}}^{{\text{2}}}$ = 0.109. Корреляция r = –0.304 дает детерминацию r2 = 0.092. Из двух значений R2 и одного r2 получаем для 170-летней густоты микроценоза силу ее влияния на диаметр сосны в 184 г., в среднем равной 0.098, или 9.8%.
Такие же расчеты для 120-летней густоты микроценоза показывают силу ее среднего влияния на диаметр сосны в 184 г. 0.118 или 11.8% (последний столбец в таблице).
На участке “Южный” (более сухом) в 120 лет на 108 площадках по 0.01 га среднее влияние густоты оказалось равно 0.081, в том числе в более густом месте массива на 54 площадках оно снизилось до 0.059. На участке “Северный” (более влажном) на 108 площадках среднее влияние густоты равно 0.155, в том числе в более редком месте 0.163. Однако увеличение влияния не достоверно из-за слабых корреляций. При этом оценка по средним значениям из полиномов ($R_{{{\text{ср}}}}^{{\text{2}}}$) показала увеличение силы влияния густоты от 0.081 до 0.251, т.е. в 3.1 раза. Поэтому вполне возможно, что увеличение густоты древостоя в целом (густоты макроценоза) ослабляет ее влияние в микроценозах, т.е. на уровне индивидуальных взаимодействий соседствующих деревьев.
В целом полученные данные характеризуют площадки 0.01 га как сообщества со слабым влиянием их густоты на диаметр деревьев. При усреднении показателей детерминации и аппроксимации линий трендов в возрасте 184, 170 и 120 лет их густота действует на диаметр деревьев с силой, соответственно, 8.8, 9.8 и 11.8%. Усреднение этих данных дает нам силу влияния густоты микроценозов на диаметр сосны равную 10.1%.
Далее рассмотрим нижнюю часть таблицы, с данными пробных площадей размером 0.04–0.12 га. Эти размеры – уже уровень мидиценоза (среднего между микро- и макроценозом), с приближением к макроценозу, т.е. полноценному древостою.
На пробных площадях размером 0.04 га оценки силы влияния густоты в последнем столбце таблицы в возрасте 120 и 170 лет составили, соответственно, 0.243 и 0.321; далее, для пробных площадей размером 0.09 га, они были равны 0.419 и 0.361, и для 0.12 га они возросли до 0.419 и 0.528. В сравнении с микроценозами (площадками по 0.01 га) эти значения выше в среднем в 4.0 раза. Здесь повышение корреляции “густота × диаметр” достоверно, и при переходе от площадей размером 0.01 га к размеру 0.04 га для возраста 170 лет они были равны r = –0.304 ± 0.062 и r = –0.542 ± ± 0.096, при достоверности различия между ними t = 2.07 > t0.05 = 1.98.
Дальнейшее увеличение размера пробных площадей до 0.24 и 0.36 га приводит к увеличению корреляций “густота × диаметр” до –0.650 и –0.747 и показателям R2 для линейных связей до 0.422 и 0.557 соответственно. Они выше, чем их значения на меньших пробных площадях размером 0.09–0.12 га, где они были равны для корреляций –0.576 и –0.705, и для показателей R2 линейных связей 0.332 и 0.497. Однако малые выборки из 9 и 6 пар значений для крупных пробных площадей размером 0.24–0.36 га это повышение доказать не позволяют. Кроме того, построенные по 6 точкам полиномы 3–6-й степени имеют значения R2 = = 0.990–1.000, которые вообще нельзя использовать, так как тренд просто соединяет отдельные точки. И мы приводим их лишь как пример значений, на которых заканчивается корректный регрессионный анализ.
Поэтому, подводя итог расчетам силы влияния густоты на уровне древостоя на диаметр деревьев в 184 г., можно взять за основу результаты определения ее влияния по 18 пробным площадям размером по 0.12 га, т.е. на уровне мидиценоза. Здесь и корреляции, и показатели аппроксимации трендов связей “густота × диаметр” достоверны, и при усреднении оценок, полученных для линейных и криволинейных связей, для густоты в 120 и 170 лет они были в среднем равны, соответственно, 0.419 и 0.528. То есть в целом для насаждения сосны флуктуации густоты на уровне мидиценозов в возрасте 120–170 лет повлияли на диаметр деревьев в 184 г. в среднем с силой (0.419 + 0.528) : 2 = 0.474, или 47.4%.
Полученные здесь результаты интересны в сравнении с результатами, полученными другими методами и другими авторами. Например, при выделении в структуре насаждения биогрупп с предельно малым расстоянием между деревьями. Они представляют немалый интерес, так как деревья, оставшиеся в одиночестве после их разреживания, существенно не увеличивали прирост (Сеннов, 1999). Кроме того, несмотря на стремление к равномерному размещению деревьев после рубок ухода за лесом в таких ухоженных лесах со временем все равно образовывались прогалины, а биогруппы сохранялись (Нестеров, 1961).
Наличие биогрупп, где формируется 28–57% деревьев, является своеобразным атрибутом древостоев (Рогозин, Разин, 2015). В изучаемом здесь насаждении в них продолжает расти 40.4% деревьев, и диаметры деревьев в них и в древостое отличаются недостоверно. Кроме того, деревья в них взаимно коррелировали диаметрами при r = = 0.47–0.58 (Рогозин, 2018а). Это свидетельствует о партнерстве деревьев-соседей, так как конкуренция приводила бы к противоположному результату – рассогласованию их роста и отрицательной корреляции.
Между тем, действующая парадигма предполагает, что более плотное размещение растений усиливает конкуренцию, и она должна снижать размер особей. И если в лесу будут густые места, то диаметры деревьев в них должны быть меньше. В этой логике общую закономерность, действующую на уровне всего ценоза, априорно перемещают на уровень индивидуальных взаимодействий. На первый взгляд, это справедливо, потому что густота – независимая переменная и главный фактор в основной закономерности морфогенеза одноярусных древостоев (Разин, 1979), которой далее был присвоен ранг закона (Рогозин, Разин, 2015). Наряду с ним, в древостоях действуют еще три закона: закон естественного изреживания (Морозов, 1949), ранговый закон роста деревьев (Маслаков, 1984), а также гомеостаз частот деревьев правых и левых форм (Голиков, 2011). В последнем находит свое воплощение всеобщий закон “Золотой пропорции”, и по нему частоты этих форм стремятся к соотношению 50:50 либо 62:38. При этом левые формы хорошо адаптированы к более сухим почвам и росту на свободе, а правые – к влажным почвам и конкуренции.
Проявление Золотой пропорции обнаружено нами и в исследуемом здесь насаждении (Рогозин, 2019). Установлено, что на расстояниях между соседями до 4.5 м их диаметры коррелировали с r = 0.52 ± 0.04. Кроме того, у них было рассчитано отношение диаметров (максимального к минимальному). Оно оказалось равно 1.273 и совпадало с “постоянной Хатчинсона”, применяемой для оценки экологических ниш (Розенберг, 2007). В нашем случае это отношение характеризует древостой как сообщество с минимальной конкуренцией между деревьями за ресурсы питания. Наиболее же ярко подтверждают такую минимизацию конкуренции данные В.М. Горячева (1999) о наступлении у деревьев-соседей пиков прироста с различиями в 1–2 недели, что свидетельствует о партнерских отношениях и резком снижении конкуренции и, тем самым, разделении экологической ниши между соседствующими деревьями во времени. Впервые же о толерантности деревьев упоминалось при исследовании модельных культур ели (Кайрюкштис, Юодвалькис, 1976). Если же вернуться к полученному нами отношению диаметров (1.273) и возвести его в квадрат, то получим значение 1.62, которое почти совпадает с Золотой пропорцией, равной ≈1.618.
В связи с этим для понимания характера отношений деревьев в микроценозах перечисленных четырех законов уже недостаточно. Не исключено, что в микроценозах действует еще один закон – закон минимизации конкуренции между деревьями и смены ее на взаимное партнерство, где как раз и найдется место для объяснения роли правых и левых форм, взаимной корреляции их диаметров и гармоничных отношений диаметров деревьев в соответствии с законом Золотой пропорции. Если же рассуждать методом от противного и полагать, что в микроценозах конкуренция имеет такую же силу, как и в древостое, то биогруппы должны обязательно изреживаться и в конце концов деревья будут располагаться по-одиночке. Но в динамике биогрупп таких тенденций нет – они сохраняются до спелого возраста и с прежней долей деревьев в них (Марченко, Брайко, 1985; Маслаков, 1999; Рогозин, 2018а). Среди травянистых растений явление партнерства также имеет место, где оно получило название “эффекта Олли” (Allee, 1931), а также “эффекта группы” (Титов, 1978). Поэтому есть все основания полагать, что явление ослабления конкуренции между деревьями в микроценозах имеет общий характер. И его также следует отнести к закону, действующему в лесных насаждениях.
Перечисленные работы, а также наши исследования дают экспериментальные факты, выводы по которым выходят далеко за рамки традиционных представлений о внутривидовых взаимоотношениях деревьев. Они показывают, что на индивидуальном уровне конкуренция между деревьями за свет и элементы питания, если ее оценивать по густоте микроценозов, отнюдь не главный фактор. Следует учитывать и влияние генотипа, например, правых и левых форм деревьев. Эти формы имеют доказанные генетические отличия по наблюдаемой гетерозиготности (Голиков, 2011). Но важно иметь хотя бы приблизительные оценки по влиянию генотипа на размер деревьев, и сравнить их с действием густоты с тем, чтобы воздействовать лесохозяйственными мерами на более сильный из них. В исследованиях лесных генетиков и селекционеров влияние генотипа на рост деревьев оценивают в 20–40% (Исаков, 1999; Тараканов, 2003; Царев, 2013). Получается, что генотип – в 3 раза более сильный фактор, чем густота микроценоза, для которой мы определили ее влияние в 10.1%. Поэтому безусловным приоритетом при уходе за лесом должно быть создание комфортных условий для деревьев-лидеров, размеры которых определяет их генотип. Вместе с тем, исходя из совместной доли влияния густоты и генотипа примерно на 40%, следует признать, что в гораздо большей мере на рост дерева действуют неустановленные и действующие локально факторы. Возможно, это флуктуации литосферы, увлажнения и состава почвы в местах микоценозов, т.е. особенности насаждения как геофитоценоза.
В данной работе получены также оценки влияния густоты на уровне миди- и макроценозов. Их густота в 120–170 лет повлияла на средний диаметр сосны в 184 г. с силой, соответсвтвенно, 47.4 и 46.6% (в среднем 47%). Это сильное влияние, но оно оказалось в два раза ниже, чем у других авторов, где ее влияние достигало 88–93% (Нагимов, 2000). Это вполне объяснимо, так как там З.Я. Нагимовым использовались данные по насаждениям из разных лесничеств, отличающихся по истории их формирования гораздо больше, чем в нашем случае. Нами же был рассмотрен единый массив сосны, где изменчивость показателей была намного ниже.
ЗАКЛЮЧЕНИЕ
Изучение микро- миди- и макроценозов в массиве 184-летнего насаждения сосны первого класса бонитета с высокой полнотой показало, что естественные флуктуации густоты микроценозов в 120 лет на площадках 0.01 га повлияли на отпад деревьев к 184 годам с силой 4.7%, а в остальных 95.3% случаев на отпад повлияли пока неустановленные факторы, действующие как особенности среды геофитоценоза. Густота мидиценозов на площадях размером 0.12 га в 120–170 лет повлияла на средний диаметр сосны в 184 г. с силой в среднем 47.4%. На уровне макроценозов на площадях по 0.24 га также получено близкое (46.6%) ее влияние, тогда как в указанных возрастах густота микроценозов повлияла на средний диаметр сосны в среднем на 10.1%, что в 4.7 раза слабее. Эти факты позволяют выдвинуть положение о том, что в микроценозах внутривидовая конкуренция к спелому возрасту ослабевает и меняется на партнерство. В связи с этим и сведениями из литературы можно также полагать, что если генотип влияет на размеры дерева примерно на 30%, а густота микроценоза, по нашим данным, – на 10%, то на долю пока неизвестных факторов, действующих в насаждении как геофитоценозе, приходится до 60% их влияния на рост деревьев в микроценозах в период от 120 до 184 лет.
Список литературы
Баландин Р.К. Анти-Дарвин. Миражи эволюции. М.: Яуза Эксмо, 2010. 350 с.
Борисов А.Н., Иванов В.В., Екимов Е.В. Метод оценки пространственного распределения ресурса в экологической нише // Сибирский лесной журн. 2014. № 5. С. 113–121.
Бузыкин А.И., Гавриков В.Л., Секретенко О.П., Хлебопрос Р.Г. Анализ структуры древесных ценозов. Новосибирск: Наука, 1985. 94 с.
Вайс А.А. Научные основы оценки горизонтальной структуры древостоев для повышения их устойчивости и продуктивности (на примере насаждений Западной и Восточной Сибири): Автореф. дис. …. д-ра с.-х. наук (спец. 06.03.02 – лесоведение, лесоводство, лесоустройство и лесная таксация). Красноярск, 2014. 33 с.
Голиков А.М. Эколого–диссимметрийный и изоферментный анализ структуры модельных популяций сосны обыкновенной // Лесоведение. 2011. № 5. С. 46–51.
Горшков В.В., Ставрова Н.И., Баккал И.Ю. Динамика восстановления лесной подстилки в бореальных сосновых лесах после пожаров//Лесоведение. 2005. № 3. С. 37–45.
Горячев В.М. Влияние пространственного размещения деревьев в сообществе на формирование годичного слоя древесины хвойных в южнотаежных лесах Урала // Экология. 1999. № 1. С. 9–19.
Грабарник П.Я. Анализ горизонтальной структуры древостоя: модельный подход // Лесоведение. 2010. № 2. С. 77–85.
Грабарник П.Я., Секретенко О.П. Анализ горизонтальной структуры древостоев методами случайных точечных полей // Сибирский лесной журн. 2015. № 3. С. 32–44.
Демаков Ю.П. Сухостой с живым комлем? // Научные труды государственного природного заповедника “Большая Кокшага”. Йошкар-Ола: Поволжский государственный технологический университет, 2007. Вып. 2. С. 331–333.
Демаков Ю.П. Структура и закономерности развития лесов республики Марий Эл. Йошкар-Ола: Поволжский государственный технологический университет, 2018. 432 с.
Исаков Ю.Н. Эколого-генетическая изменчивость и селекция сосны обыкновенной: Автореф. … д-ра. биол. наук (спец. 03.00.15 – генетика 06.01.05 – селекция и семеноводство). СПб: Санкт-Петербургская лесотехническая академия, 1999. 36 с.
Кайрюкштис Л.В., Юодвалькис А.И. Явление смены характера взаимоотношений между индивидами внутри вида // Лесоведение и лесное хозяйство. Минск, 1976. Вып. II. С. 16–24.
Калинин М.И. Формирование корневой системы деревьев. М.: Лесн. пром-сть, 1983. 152 с.
Карев Г.П. Эргодические свойства стационарных состояний лесных экосистем // Проблемы экологического мониторинга и моделирования экосистем. СПб.: Гидрометеоиздат, 1999. Вып. 17. С. 122–141.
Кислов Д.Е., Прилуцкий А.Н., Брижатая А.А. Статистический анализ неоднородности горизонтальной структуры древостоев // Поволжский экологический журн. 2015. № 3. С. 268–276.
Колобов А.Н. Моделирование пространственно-временной динамики древесных сообществ: индивидуально-ориентированный подход // Лесоведение. 2014. № 5. С. 72–82.
Колтунова А.И. О формировании горизонтальной структуры и срастании корневых систем в древостоях сосны // Эко-потенциал. 2013. № 3–4. С. 136–142.
Крамер П.Д., Козловский Т.Т. Физиология древесных растений. М.: Гослесбумиздат, 1963. 627 с.
Костерин О.Э. Дарвинизм как частный случай “бритвы Оккама” // Вестник ВОГиС. 2007. Т. 11. № 2. С. 416–431.
Кузьмичев В.В. Закономерности динамики древостоев. Новосибирск: Наука, 2013. 208 с.
Мартынов А.Н. Зависимость биометрических показателей сосны от площади питания // Лесоведение. 1976. № 5. С. 85–88.
Маслаков Е.Л. Формирование сосновых молодняков. М.: Лесн. пром-сть, 1984. 168 с.
Маслаков Е.Л. Генезис и динамика социальных структур сосны в фазе индивидуального роста // Таежные леса на пороге XXI в. СПб.: Санкт-Петербургский научно-исследовательский институт лесного хозяйства, 1999. С. 42–51.
Марченко И.С., Брайко В.Б. Деревья главного пользования// Молодые ученые 40-летию победы и 1000-летию г. Брянска. Брянск: Брянский государственный технологический институт,1985. С. 38–39.
Морозов Г.Ф. Учение о лесе. Изд. 7-е / Под ред. Нестерова В.Г. М.: Гослесбумиздат, 1949. 453 с.
Нагимов З.Я. Оценка методов определения площадей роста деревьев // Леса Урала и хозяйство в них. Екатеринбург: Уральская государственная лесотехническая академия, 1999. Вып. 19. С. 82–98.
Нагимов З.Я. Закономерности роста и формирования надземной фитомассы сосновых древостоев: Дис. … д‑ра с.-х. наук (спец. 06.03.03 – лесоведение и лесоводство; лесные пожары и борьба с ними). Екатеринбург: Уральская государственная лесотехническая академия, 2000. 409 с.
Нестеров В. Г. Вопросы современного лесоводства. М.: ГСХИ, 1961. 384 с.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
Плантационное лесоводство / Под ред. Шутова И.В. СПб.: Изд-во Политехнического университета, 2007. 366 с.
Правила ухода за лесами. Утверждены приказом Минприроды России от 22.11.2017 № 626.
Разин Г.С. Динамика сомкнутости одноярусных древостоев // Лесоведение. 1979. № 1. С. 23–25.
Рахтеенко И.Н. Рост и взаимодействие корневых систем древесных растений. Минск: Изд-во АН БССР, 1963. 254 с.
Рогозин М.В. Биогруппы в старых насаждениях сосны // Вестник Пермского университета. Серия Биология. 2018а. Вып. 2. С. 150–158.
Рогозин М. В. Площадь питания дерева: анализ методов // Бюллетень науки и практики. 2018б. Т. 4. № 7. С. 34–37.
Рогозин М.В. Постоянная Хатчинсона и конкуренция в сосновом древостое // Научное обозрение. Биологические науки 2019. № 1. С. 51–55.
Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы Пермь: Пермский государственный национальный исследовательский университет, 2015. 277 с.
Розенберг Г.С. Экология в картинках (Учебное пособ.). Тольятти: Институт экологии Волжского бассейна РАН. 2007. 218 с.
Секретенко О.П. Метод анализа пространственной структуры древостоев // Исследование структуры лесонасаждений. Красноярск: Институт леса и древесины, 1984. С. 88–101.
Сеннов С.Н. Итоги 60-летних наблюдений за естественной динамикой леса // С-Пб.: Санкт-Петербургский научно-исследовательский институт лесного хозяйства, 1999. 98 с.
Ставрова Н.И., Горшков В.В., Катютин П.Н. Возрастная и пространственная структура ценопопуляций Pinus sylvestris (Pinaceae) в условиях северной тайги (Кольский п-ов) // Растительные ресурсы. Т. 48. 2012. № 1. С. 16–34.
Ставрова Н.И, Горшков В.В., Катютин П.Н. Формирование структуры ценопопуляций лесообразующих видов в процессе послепожарного восстановления северотаежных лесов // Труды Карельского научного центра РАН. № 12. 2015. С. 1–9.
Стороженко В.Г. Устойчивые лесные сообщества: (теория и эксперимент). М.: Институт лесоведения РАН, 2007. 190 с.
Титов Ю.В. Эффект группы у растений. Л.: Наука, 1978. 151 с.
Тараканов В.В. Структура изменчивости, селекция и семеноводство сосны обыкновенной в Сибири. Дис. … д-ра с.-х. наук (спец. 06.03.01 – лесные культуры, селекция и семеноводство). Новосибирск: Институт леса СО РАН, 2003. 454 с.
Усольцев В.А. Продукционные показатели и конкурентные отношения деревьев. Исследование зависимостей. Екатеринбург: УГЛТУ, 2013. 556 с.
Усольцев В.А., Часовских В.П., Акчурина Г.А., Осмирко А.А., Кох Е.В. Фитомасса деревьев в конкурентных условиях: исследование системных связей средствами информационных технологий. Екатеринбург: Уральский государственный лесотехнический университет, 2018. 526 с.
Царев А.П. Программы лесной селекции в России и за рубежом. М.: Московский государственный университет леса, 2013. 164 с.
Allee W.C. Animal aggregations: A study in general sociology. Chicago, Illinois: University of Chicago Press, 1931. 431 p.
Grenfell R., Aakala T., Kuuluvainen T. Microsite occupancy and the spatial structure of understorey regeneration in three late-successional Norway spruce forests in northern Europe // Silva Fennica. 2011. V. 45(5). P. 1093–1110.
Kenkel N.C. Patterns of self-thinning in jack-pine: testing of the random mortality hypothesis // Ecology. 1988. V. 69. P. 1017–1024.
Kuuluvainen T., Wallenius T.H., Kauhanen H., Aakala T., Mikkola K., Demidova N., Ogibin B. Episodic, patchy disturbances characterize an old-growth Piceaabies dominated forest landscape in northeastern Europe // Forest Ecology and Management. 2014. V. 320. P. 96–103.
Linder P., Elfving B., Zackrisson O. Stand structureand successional trends in virgin boreal forest reserves in Sweden // Forest Ecology and Management. 1997. V. 98. P. 17–33.
Omelko A., Ukhvatkina O., Zhmerenetsky A., Sibirina L., Petrenko T., Bobrovsky M. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pinebroadleaved forest // Forest Ecology and Management. 2018. V. 411. P. 46–66.
Дополнительные материалы отсутствуют.