

Лесоведение, 2021, № 1, стр. 42-51
Определение переходов между онтогенетическими состояниями лиственницы, сосны и кедра в северной тайге Западной Сибири
В. Р. Цибульский a, *, И. Г. Соловьев a, b, Д. В. Московченко a, c, Д. А. Говорков a, b
a ФИЦ “Тюменский научный центр СО РАН”, структурное подразделение “Институт проблем освоения Севера”
625026 Тюмень, ул. Малыгина, 86, Россия
b Тюменский индустриальный университет
625000 Тюмень, ул. Володарского, 38, Россия
c Тюменский государственный университет
625003 Тюмень, ул. Володарского, 6, Россия
* E-mail: v-tsib@yandex.ru
Поступила в редакцию 30.07.2018
После доработки 30.12.2018
Принята к публикации 07.10.2020
Аннотация
Для оценки изменения биоразнообразия на уровне видов необходима динамическая модель популяции эдификатора, определяющего соответствующий биом территории. В статье предложен аналитический метод определения возрастных границ между ювенильным и виргинильным, виргинильным и генеративным онтогенетическими состояниями деревьев хвойных пород, имеющих важное значение при идентификации популяционных моделей. В основе метода определения указанных границ принята функция роста Томазиуса, применимая как для объемных оценок, так и для высоты и диаметра деревьев. В нашем случае она описывает рост радиуса на уровне 1.3 м как наиболее доступный параметр при проведении натурных наблюдений. Эта функция представляет собой симметричную сигмоиду, что не соответствует реальным данным роста для хвойных пород. Для устранения этого недостатка предложено ввести нелинейность в степенной показатель модели. Идентификация модели осуществлялась путем последовательных приближений модели и кривых роста, полученных как усреднение реальных временных рядов на основе древесно-кольцевых хронологий деревьев – эдификаторов северной тайги Западной Сибири: лиственницы, сосны и кедра. Ранее авторами была опробована методика определения искомых границ онтогенетических состояний в моменты равенства нулю второй и третьей производных кривых роста, заданных в виде полиномов третьей и четвертой степени и полученных на основе статистической обработки реальных данных. В данном случае предлагается границы определять по функциям, имеющим биофизический смысл, т.е. дожитие и рождаемость или рост и торможение. Кроме того, границы определяются по второй производной кривой роста по достижении ею в первом случае максимума, во втором – минимума. Это значительно облегчает понимание и получение результатов. В статье приведены усредненные кривые роста лиственницы, сосны и кедра для районов вблизи поселков Сидоровск, Вынгапур, Красноселькуп и Толька (Ямало-Ненецкий автономный округ), а также временные отметки границ исследуемых онтогенетических состояний и коэффициенты моделей для них.
Проблема оценки устойчивости биоразнообразия на севере климатической зоны Западной Сибири в значительной степени определяется деревьями-эдификаторами, отражающими специфический для них биом и в конечном счете – фитоценоз территории (Свирежев, Логофет, 1978; Лукина и др., 2015). На границе лесотундры и тундры такими эдификаторами являются лиственница, сосна и кедр. Искомая оценка может быть реализована на основе динамических математических моделей популяции указанных деревьев (Смирнова и др., 1993).
Для проведения исследований влияния различных внешних факторов на продуктивность древостоя во времени разрабатывались динамические модели начиная от Г. Бэкмана, Х. Томазиуса (Лир и др., 1974) и заканчивая более поздними публикациями (Хильми, 1957; Петровский, 1968; Карев, 1983; Кулль, Оя, 1984; Носова и др., 2005; Желдак и др., 2013; Колобов, 2014). Большинство из указанных моделей являются нелинейными и содержат различные трудно определяемые показатели. Модели полученные на основе теории вероятностей и “теории встреч”, впервые предложенные Р. Глокером (Лир и др., 1974) успешно применяются и сейчас.
В настоящее время некоторые ученые рассматривают модели динамики древостоев с позиций уточнения влияния изреживания и возрастного умирания на воспроизводимость: например, это (Березовская, Карев, 2015; Soukhovolsky, Ivanova, 2018). Для мониторинга биоразнообразия лесов интересны работы по моделям биологических сообществ в виде проекционных матриц (Уланова и др., 2002; Кузнецов, 2006, Авдеева, Кузьмичев, 2007). Эти модели базируются на возрастной оценке фаз роста популяций и периодических натурных наблюдениях плотности в этих фазах. Имеются только отдельные попытки разработать и идентифицировать такие модели для лесных сообществ (Фардеева, Исламова, 2007; Евстигнеев, 2014).
Одной из проблем в таких работах является соответствие возрастного разделения фаз роста и данных по рождаемости, полученных при натурных мониторинговых наблюдениях. Если плотность по возрастам можно определить, применяя возрастную классификацию (Загреев и др., 1992), то для определения границ фаз необходимо использовать традиционные показатели: высота, высота основания кроны, порядок ветвления, диаметр ствола на уровне 1.3 м и др. (Евстигнеев, 2014). Это затрудняет работу при коротких экспедиционных исследованиях.
Авторами предпринята попытка определить эти границы по изгибу кривых роста на модельных деревьях самых больших диаметров и высоты, чаще всего субсенильного возраста, из предположения, что выделенный участок является частью нормальной ценопопуляции по Т.А. Работнову (Смирнова и др., 1993). По Г.П. Кареву (1999) это ближе всего к “ценнону”, т.е. участку (“ячейке”) в географическом и фазовом пространстве с однородной лесной субпопуляцией. Такой метод позволяет более определенно, в математическом смысле, привязывать границы фаз онтогенеза дерева к свойствам функции роста (прироста).
Конечной целью наших исследований является разработка модели популяции дерева-эдификатора на основе получения информации (кернов) роста по радиусу (диаметру) внутри каждого класса роста, в том числе и в субсенильной части возраста.
В данной статье предлагается формализация известной методики выделения “фаз развития” точками перегиба хода роста древостоя (см., например, (Березовская, Карев, 2015)), используя кривые роста (прироста) по диаметру модельного дерева (группы) в субсенильной фазе.
ОБЪЕКТЫ И МЕТОДИКА.
Для апробации предлагаемой методики были взяты по четыре временных ряда древесно-кольцевых хронологий (ДКХ) деревьев-эдификаторов: лиственницы и сосны в международной базе данных древесных колец (The International Tree-Ring Data Bank of National Centers for Environmental Information) вблизи поселков Сидоровск и Вынгапур (Ямало-Ненецкий автономный округ) и два ряда кедра около поселков Красноселькуп и Толька (Ямало-Ненецкий автономный округ). И первые и вторые полученны С.П. Арефьевым в результате экспедиционных исследований. Районы исследований с координатами отбора кернов и номерами временных рядов приведены в табл. 1, что позволяет любому исследователю найти соответствующие ряды данных. Для усреднения кривых роста временные ряды приведены к единице путем деления на rmax на дату взятия пробы (керна) на уровне 1.3 м. Временные ряды выбирались для деревьев без угнетения роста, т.е., для более освещенных участков леса.
Таблица 1.
Временные ряды древесно-кольцевых хронологий лиственницы, сосны и кедра
| Сидоровск (66°40′ с.ш., 82°20′ в.д.) (Лиственница) |
Вынгапур (62°93′ с.ш., 76°38′. в.д.) (Сосна) |
Толька (64°00′ с.ш., 82°02′ в.д.), Красноселькуп (65°07′ с.ш., 82°46′ в.д.) (Кедр) |
|||
|---|---|---|---|---|---|
| № | годы | № | годы | № | годы |
| 863 051 | 1786–1990 | 080 421 | 1796–1994 | С-10 | 1729–1994 |
| 863 071 | 1779–1990 | 080 431 | 1792–1994 | С- 0 | 1585–1994 |
| 863 072 | 1786–1990 | 080 441 | 1802–1994 | ||
| 863 102 | 1767–1990 | 080 442 | 1796–1994 | ||
При выборе выделенного участка для идентификации модели популяции рекомендуется для определения границ фаз использовать деревья субсениального возраста со здоровой кроной.
Для хвойных пород различают онтогенетические состояния как у большинства деревьев. При описании их будем отмечать только признаки, говорящие о скорости роста по высоте и диаметру и биофизическом смысле этих состояний.
1. Проростки, всходы (p). Начинаются после выхода зародыша из оболочки семени (прорастание) и длятся до появления первых хвоинок-семядолей (чаще 6–7 шт.) не более года. Завершается образованием верхушечной почки, из которой на второй год формируется центральный побег со своей хвоей. Корневая система представлена обычно осевым корнем. Чаще всего большинство авторов объединяют эти два состояния, называя предювенильной фазой (Николаева и др., 2008), проростками (Фардеева, Исламова, 2007). Для хвойных растений – лиственницы сибирской, сосны и кедра средняя высота 4–9 см, средняя глубина проникновения корневой системы от 4 до 12 см. Биофизический смысл этих состояний заключается вначале в гетеротрофном, а затем в автотрофном питании.
На второй год (автотрофный) происходит формирование центрального неветвящегося побега со своей хвоей и корневой системой с малочисленными боковыми корнями. Верхушечная почка раскрывается и образует прирост ствола в высоту (1–4) см – надсемядольное междоузлие (эпикотиль). Корень проникает в почву на глубину до 10–15 см. Продолжительность фазы составляет 2–5 лет до отмирания семядолей и появления укороченных побегов (Николаева и др., 2008; Евстигнеев, 2014; Evstigneev, Korotkov, 2016).
2. Ювенильная фаза (j) начинается с отмирания семядолей до первого ветвления и продолжается 2–3 г. В этой фазе годичный прирост в высоту невелик 5–10 см, корни углубляются на 5–10 см, образуются боковые корни с ответвлениями. Растения находятся в травяно-кустарничковом ярусе (Смирнова и др., 2001; Николаева и др., 2008; Евстигнеев, 2014). Недоразвитость системы питания (корневая и ветвление) не позволяют обеспечивать значительный рост в высоту и в диаметре. При этом у большинства хвойных в этом состоянии наблюдается хорошая теневыносливость.
3. Имматурная фаза (im) начинается с появления боковых ветвей. Происходит усиление роста ствола в высоту, при этом его диаметр в первые годы остается небольшим. Формируются боковые и придаточные корни. Фаза длится 2–9 лет, в течение которой растения выходят к кустарниковому ярусу (Смирнова, Бобровский, 2001). Годичный прирост составляет по высоте 3–20 см, по диаметру 0.05–1.5 мм (Фардеева, Исламова, 2007; Николаева и др., 2008; Евстигнеев, 2014).
Эта фаза считается промежуточной между ювенильной и виргинильной, в ней уже в достаточной степени организованны процессы фотосинтеза и ассимиляции (Крамер, Козловский, 1983; Смирнова, Бобровский, 2001), что создает предпосылки для перехода от небольшого прироста к максимально возможному.
4. Виргинильная фаза (v) характеризуется сформировавшейся удлиненной кроной с отсутствием у лиственницы и сосны сухих боковых ветвей на большей части высоты ствола (до 60%). Происходит фотосинтез, ассимиляция и функционирование меристем в полном объеме. При этом ускоряется рост центрального побега и боковых. Наблюдается формирование значительного количества мощных боковых корней. В конце виргинильной фазы растения переходят в древесный ярус (Смирнова, Бобровский, 2001), появляются мужские и женские стробилы и после опыления первые шишки. Годичный прирост по высоте в южных зонах может достигать 60 см, в северных – до 30 см, по радиусу 0.3–5 мм. (Фардеева, Исламова, 2007; Николаева и др., 2008; Евстигнеев, 2014).
Именно в этой фазе происходит перегиб кривой роста, т.е. достижение максимального значения годичного прироста. Принято считать, что появление первых шишек является пограничным признаком виргинильной и генеративной фаз.
5. Генеративная фаза (g). Характерными процессами являются цветение, образование шишек и семян и их распространение на фоне замедления процессов относительного роста. По публикациям многих авторов генеративная фаза начинается от 60 до 100 лет и длится около 200 лет (Каппер, 1954). Характерно для этой фазы уменьшение скорости роста и по высоте, и по диаметру ствола.
6. Фазы старости: субсенильная (SS) и сенильная (S) характеризуются слабым вегетативным ростом, уменьшением образования стробил и созревания шишек.
Таким образом, даже при кратком описании онтогенетических состояний очевидно, что максимальный рост во всех рассматриваемых деревьях находится в виргинильной фазе. Кроме того, между более медленной ювенильной и замедляющейся генеративной и быстрой виргинильной фазами должен быть перегиб (граница). На рисунке 1 приведена типичная кривая прироста, на которой отмечены фазы и места перегиба.
Рис. 1.
Типичная кривая прироста радиуса дерева t1 – первый момент перегиба кривой радиуса дерева, t2 – второй момент перегиба, p – момент окончания стадии всходов, j – момент окончания ювенильной стадии, im – момент окончания имматурной стадии.
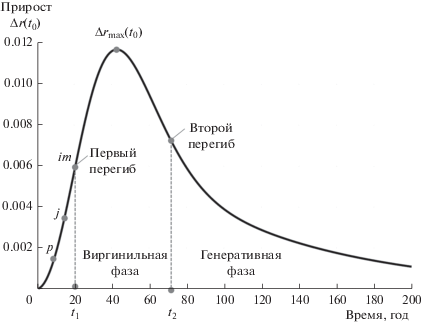
При разработке популяционной матрицы важно знать границы между фазами ювенильной и виргинильной, виргинильной и генеративной, причем не статистические или взятые из справочника, а полученные от модельного дерева исследуемого выделенного участка.
Авторы в своих исследованиях предлагали при обработке временных рядов на основе ДКХ определять границы ювенильной, виргинильной и генеративной фаз путем установления точек перегиба на кривых роста и прироста радиуса r(t) после сглаживания временных рядов авторегрессионным методом. Однако разброс при этом в зависимости от порядка полинома был достаточно велик (около 50%). На рисунках 2–4 приведены усредненные кривые роста лиственницы, сосны и кедра. Они показывают, что определить точку перегиба между усиленным ростом и началом замедления на них достаточно сложно.
Рис. 2.
Кривая роста лиственницы, ее аппроксимация и ее производные первой и второй степени. а – кривая роста лиственницы, ее аппроксимация и первая производная от аппроксимации; б – графики второй производной от аппроксимации. 1 – кривая роста лиственницы, 2 – аппроксимация кривой по функции (1), 3 – аппроксимация кривой по функции (2), 4 – первая производная от аппроксимированной функции (2), 5 – вторая производная от аппроксимированной функции (1), 6 – вторая производная от аппроксимированной функции (2).
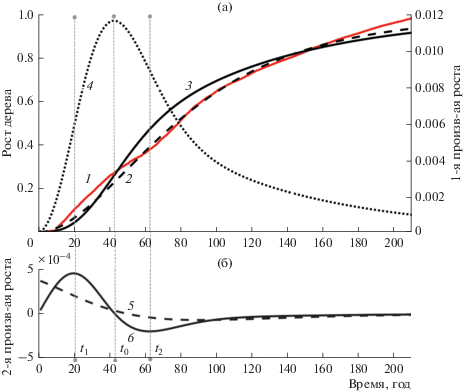
Рис. 3.
Кривая роста сосны, ее аппроксимация и ее производные первой и второй степени. а – кривая роста сосны, ее аппроксимация и первая производная от аппроксимации; б – графики второй производной от аппроксимации. 1 – кривая роста сосны, 2 – аппроксимация кривой по функции (1), 3 – аппроксимация кривой по функции (2), 4 – первая производная от аппроксимированной функции (2), 5 – вторая производная от аппроксимированной функции (1), 6 – вторая производная от аппроксимированной функции (2).
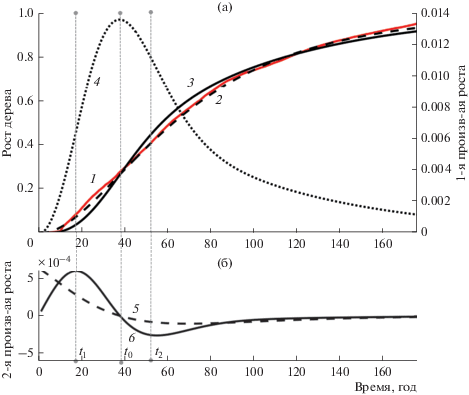
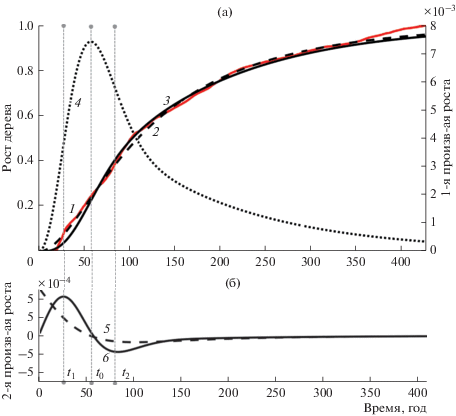
Рис. 4.
Кривая роста кедра, ее аппроксимация и ее производные первой и второй степени. а – кривая роста кедра, ее аппроксимация и первая производная от аппроксимации; б – графики второй производной от аппроксимации. 1 – кривая роста кедра, 2 – аппроксимация кривой по функции (1), 3 – аппроксимация кривой по функции (2), 4 – первая производная от аппроксимированной функции (2), 5 – вторая производная от аппроксимированной функции (1), 6 – вторая производная от аппроксимированной функции (2).
Нами предложено аппроксимировать усредненную кривую роста известной функцией роста Глокерса, распространенной на деревья, в том числе хвойные (Thomasius, 1963; Лир и др., 1974). Ее нормированный (приведенный к единице) вид выглядит для радиуса ствола дерева следующим образом:
где r(t) – радиус ствола на отметке 1.3 м, a и c – скоростные показатели роста и замедления соответственно.Исследования показали, что приведенная функция роста (1) отличается от реальной кривой роста в начале и конце (рис. 2–4) и от кривых, приведенных, например, для хвойного подроста (Норин, 1958). На рисунках видно, что вторая производная этой кривой (1) не имеет первого максимума. Это создает трудности получения уверенного определения границ.
Для уточнения кривой роста предлагается ввести нелинейность в показатели замедления роста. В этом случае формула (1) будет выглядеть следующим образом:
Раскроем скобки при показателе и запишем в виде удобном для интерпретации:
Из этого выражения понятно, что второе слагаемое представляет собой относительный (приведенный) рост с показателем, изменяющимся в зависимости от квадрата произведения скорости процесса замедления, отражая влияние не только генеративных функций, но и исчерпания физиологических возможностей прежнего роста растения.
Аналитическая форма кривой роста (2), полученная путем аппроксимации реальных временных рядов, позволяет более уверенно определить точки перегиба. На кривой роста первая точка перегиба t0 определяется по условию $\frac{{dr}}{{dt}} = \max .$ Кроме того видно, что это соответствует максимуму кривой прироста ∆r(t). Аналогично, если мы хотим определить перегибы на кривой прироста, что соответствует искомым границам t1, t2, необходимо взять вторую производную $\frac{{{{d}^{2}}r(t)}}{{d{{t}^{2}}}}$ от аналитической кривой роста. Первый перегиб будет соответствовать максимуму, второй – минимуму $\frac{{{{d}^{2}}r(t)}}{{d{{t}^{2}}}}.$
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Наиболее распространенными деревьями эдификаторами на севере Западной Сибири являются лиственница, сосна и кедр. В таблице 1 приведены координаты отбора их кернов на территории лесотундры и тайги Ямало-Ненецкого автономного округа.
Из предыдущих исследований авторов известно, что аппроксимация r(t) значительно отклоняется от реального ряда на начальном участке (от 0 до 45 лет) (Цибульский и др., 2016). Это связано с тем, что для получения ДКХ керн берется на уровне 1.3 м и часть возраста не учитывается: для лиственницы и сосны это 10–12 лет, для кедра – 10–20 лет (Норин, 1958; Загреев и др., 1992).
Приведенные на рис. 2–4 кривые дополнены временным интервалом: для лиственницы и сосны он составляет 10 лет, для кедра – 20 лет по линейному закону от 0 до радиуса на отметке 1.3 м.
Как указывалось выше, из рисунков понятно, что во всех трех случаях аппроксимация (1) не позволяет получить временную отметку t1 так как ее вторая производная не имеет максимума. Напротив, предлагаемая модель (2) и ее вторая производная имеют ярко выраженный максимум в точке t1.
Таким образом аппроксимация кривой роста на основе ДКХ по предлагаемой модели (2) позволяет получить численным графоаналитическим путем временные границы онтогенетических состояний: ювенильного, виргинильного и генеративного, характерные для данного ландшафта и почвенно-климатических условий. Для выбранных модельных эдификаторов значения коэффициентов a и c и границ фаз ювенильной и виргинильной t1 и виргинильной и генеративной t2 и максимального значения роста t0 сведены в табл. 2.
Таблица 2.
Модели роста лиственницы, сосны, кедра и границы онтогенетических состояний
| Показатель | Лиственница | Сосна | Кедр |
|---|---|---|---|
| a | 0.012 | 0.014 | 0.007 |
| c | 0.022 | 0.025 | 0.017 |
| t1 | 20 | 17 | 26 |
| t0 | 43 | 37 | 58 |
| t2 | 63 | 55 | 83 |
| Модель | $r(t) = 1 - {{e}^{{ - 0,012t\left( {1 - {{e}^{{ - {{{\left( {0,022t} \right)}}^{2}}}}}} \right)}}}$ | $r(t) = 1 - {{e}^{{ - 0,014t\left( {1 - {{e}^{{ - {{{\left( {0,025t} \right)}}^{2}}}}}} \right)}}}$ | $r(t) = 1 - {{e}^{{ - 0,007t\left( {1 - {{e}^{{ - {{{\left( {0,017t} \right)}}^{2}}}}}} \right)}}}$ |
После получения значений коэффициентов a и c возможно уточнение модели (2). Получение возрастных границ фаз за счет дополнения начальных значений кривой роста уже не по линейному закону, как это было сделано в начале, а по полученному (2). Такое уточнение влияет в основном на первую границу между ювенильной и виргинильной фазами на 1–2 года. В результате появляется реальная возможность разработки и идентификации модели популяции видов-эдификаторов и программного продукта на ее основе и проведения мониторинга устойчивости биоценоза на территории их преобладания. Для этого необходимо:
1) Определить выделенный участок леса, отвечающий требованиям для мониторинга состояния.
2) Разбить на классы роста и в каждом получить усредненные значения плотности. Построить кривую плотности выделенного участка по возрастам.
3) Определить урожайность шишек в определенных возрастных классах и построить кривую урожайности.
4) Для уточнения границ ювенильной, виргинильной и генеративной фаз получить керн в трех взаимно перпендикулярных направлениях здорового представителя суперсениальной фазы, усреднить по предложенной методике определить эти границы.
5) Составить проекционную матрицу, имея информацию пп. 2 и 4 и определить ее собственное число. Если оно меньше единицы, т.е. популяция деградирует, если больше, то развивается. Если получать такую информацию, например, каждые 10 или 20 лет, то можно построить траекторию поведения популяции, оценить и прогнозировать устойчивость биоценоза.
В настоящей статье приведено решение задачи по пункту 4.
Для определения разброса значений t0, t1 и t2 для одного вида (лиственницы) по широтному “ходу” авторы на основе ДКХ полученных С.П. Арефьевым в результате экспедиционных исследований сформировали усредненные модельные деревья для районов долины реки Полуй (г. Салехард, Ямало-Ненецкий автономный округ) и долины реки Пур (п. Самбург, Ямало-Ненецкий автономный округ). Временные ряды также дополнены в начале ряда недостающими значениями в виде линейной зависимости от 0 до первого значения ширины кольца. Номера кернов, район взятия проб и возраст деревьев приведены в таблице 3. Там же приведены значения полученных временных отметок t0, t1 и t2. Разброс значений для лиственницы включая таблицу 2 (п. Сидоровск, Ямало-Ненецкий автономный округ) достаточно велик и подтверждает предположение, что для каждой выбранной площадки необходимо определять свои значения.
Таблица 3.
Временные ряды ДКХ лиственницы
| Данные по керну | р. Полуй | г. Надым | п. Самбург | |||
|---|---|---|---|---|---|---|
| керн | возраст | керн | возраст | керн | возраст | |
| Лц 7 – 1.2 | 1533–2013 | Лц 9 – 1.2 | 1581– 991 | Лц 5 – 1.2.3 | 1503–2014 | |
| Лц 6 – 1.2 | 1542–2013 | Лц 25 – 1.2 Лц 27 – 1 |
1650–1991 1651–1991 |
Лц 1 – 3.2.1 | 1557–2014 | |
| Местность | Долина реки лиственнично-брусничный лес 66°20ʹ. 691 с.ш. 67°32ʹ. 12 в.д. |
Лиственичник, лиственнично-травяно-пустырничковое редколосье 65°53ʹ с.ш. 72°51ʹ в.д. |
Долина р. Пур, разнотравянное лиственничное редколесье 67°00ʹ с.ш. 78°13ʹ в.д. |
|||
| t1 t0 t2 |
6 15 20 |
24 54 78 |
8 19 27 |
|||
Список литературы
Авдеева Е.В., Кузьмичев В.В. Специфика онтогенеза и индикаторная роль лиственницы сибирской (Larix Sibirica Ledeb) в условиях городской среды // Хвойные бореальные зоны. 2007. № 4–5. С. 362–367.
Березовская Ф.С., Карев Г.П. Моделирование динамики древостоев // Сибирский лесной журн. 2015. № 3. С. 7–19.
Евстигнеев О.И. Поливариантность сосны обыкновенной в брянском полесье // Лесоведение. 2014. № 2. С. 69–77.
Желдак В.И., Чижов Б.Е., Гаркунов Г.А., Захаров А.И. О лесоводственных мероприятиях в притундровых лесах Западной Сибири // Лесоведение. 2013. № 3. С. 19–29.
Загреев В.В., Сухих В.И., Швиденко А.З., Гусев Н.Н., Мощнасеев А.Г. Общесоюзные нормативы для таксации лесов. М.: Колос, 1992. 495 с.
Каппер О.Г. Хвойные породы. Л.: Гослесбумиздат, 1954. 303 с.
Карев Г.П. Математическая модель роста в светолимитированных древостоях // Журн. общей биологии. 1983. Т. XLIV. № 4. С. 474–479.
Карев Г.П. Структурные модели лесных экосистем // Сибирский Экологический журн. 1995. № 4. С. 381–396.
Колобов А.Н. Моделирование пространственно-временной динамики древесных сообществ: индивидуально-ориентированный подход // Лесоведение. 2014. № 5. С. 72–82.
Крамер П.Д., Козловский Т.Т. Физиология древесных растений. М.: Лесн. пром-сть, 1983. 464 с.
Кузнецов В.И. Динамические системы в задачах вычислительной экологии леса. М.: Политбук Мультимедия, 2006. 111 с.
Кулль К., Оя Т. Структура физиологических моделей роста деревьев // Известия АН ЭССР. Биология, 1984. Т. 33. Выпуск 1. С. 33–40.
Лир Х., Польстер Г., Фидлер Г.Н. Физиология древесных растений / Пер. с немецкого Н. В. Лобанова. М.: Лесн. пром-сть, 1974. с. 423 с.
Лукина Н.В., Исаев А.С., Крышень А.М., Онучин А.А., Сирин А.А., Гагарин Ю.Н., Барталев С.А. Приоритетные направления развития лесной науки как основы устойчивого управления лесами // Лесоведение. 2015. № 4. С. 243–254.
Николаева С.А., Велисевич С.Н., Савчук Д.А. Онтогенез кедра сибирского в условиях Кеть-Чулымского междуречья // Вестник Томского государственного университета. Биология. 2008. № 3(4). С. 24–34.
Норин Б.Н. К познанию семенного и вегетативного возобновления древесных пород в лесотундре // Растительность Крайнего Севера СССР и ее освоение.М.; Л.: АН СССР, 1958 Вып. 3. С. 154–244.
Носова Л.М., Тихонова Е.В., Леонова Н.Б. Воздействие деревьев-эдификаторов на биологическое разнообразие лесных экосистем // Лесоведение. 2005. № 4. С. 40–48.
Петровский В.С. Математические модели лиственничных стволов, их точность и применение. // Лиственница. Красноярск: Изд-во СТИ, 1968. Т. 3. С. 76–85.
Свирежев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. М.: Наука, 1978. 352 с.
Смирнова О.В., Бобровский М.В. Оонтогенез дерева и его отражение в структуре и динамике растительного покрова // Экология. 2001. № 3. С. 177–181.
Смирнова О.В., Заугольнова Л.Б., Попадюк Р.В. Популяционная концепция в биогеоценологии // Журн. общей биологии. 1993. Т. 54. № 3. С. 438–448.
Уланова Н.Г., Демидова А.Н., Клочкова И.Н., Логофет Д.О. Структура и динамика популяций вейника седеющего calamagrostis canescens: модельный подход // Журн. общей биологии. Т. 63. № 6. С. 509–521.
Фардеева М.В., Исламова Г.Р. Особенности популяционной организации древесных видов хвойно-широколиственных лесов // Вестник Татарского государственного гуманитарно-педагогического университета. 2007. № 2–3(9–10). С. 112–121.
Хильми Г.Ф. Теоретическая биофизика леса. М.: Издво. АН ССР, 1957. 206 с.
Цибульский В.Р., Коновалов А.А., Арефьев С.П. Кривые хода роста и прироста хвойных деревьев на севере Западной Сибири // Вестник ТюмГУ. Экология и природопольование. Т. 2. № 3. С. 59–69.
Evstigneev O.I., Korotkov V.N. Ontogenetic stages of trees: an overview // Russian Journal of Ecosystem Ecology. 2016. V. 1. № 2. P. 1–31.
Soukhovolsky V., Ivanova Y. Modeling Production Processes in Forest Stands: An Adaptation of the Solow Growth Model // Forests. 2018. № 9. P. 391–403.
Thomasius H. Untersuchungen über die Brauchbarkeit einiger Wachstumgrössen von Bäumen und Beständen für die quantitative Standortsbeurtei-lung // Archiv Forst. 1963. Bd. 12. H 12. S. 1267–1323.
Дополнительные материалы отсутствуют.