

Лесоведение, 2021, № 2, стр. 156-172
Эффекты фитогенного поля сосны в различных климатических условиях
О. В. Трефилова a, *, И. П. Беланов b, В. И. Уфимцев c, Д. Ю. Ефимов d
a Сибирский государственный университет науки и технологий имени академика М.Ф. Решетнева
660037 Красноярск, просп. им. газеты Красноярский рабочий, 31, Россия
b Институт почвоведения и агрохимии СО РАН
630099 Новосибирск, просп. Академика Лаврентьева, 8/2, Россия
c Федеральный исследовательский центр угля и углехимии СО РАН
650065 Кемерово, Ленинградский просп., 10, Россия
d Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 пос. Борок, Некоузский р-н, Ярославская обл., 109, Россия
* E-mail: ovtrefilova_ilsoran@mail.ru
Поступила в редакцию 28.11.2018
После доработки 27.07.2020
Принята к публикации 08.12.2020
Аннотация
В рамках настоящей работы предпринята попытка оценить характер (однотипность или уникальность) влияния дерева на почвы в различных климатических условиях. Для достижений цели исследования подобрана серия опорных полигонов, где одна и та же древесная порода – сосна обыкновенная (Pinus sylvestris L.) – развивается в разных эколого-географических районах (ЭГР): остепненном; северном лесостепном и среднегорном таежном, но в условиях близкого эдафического фона. Формирование насаждений происходит на слаборазвитых почвах отвалов. Это молодые почвы, для которых процессы формирования “педопамяти” (по Таргульяну) только запускаются, что позволило оценивать непосредственное влияние современного древостоя на почвы. Методологической основой работы служит концепция фитогенного поля (ФП). Показано, что влияние отдельно стоящего дерева сосны II класса возраста распространяется на 131–155 м2 и только 10–13% данной площади находится в подкроновом пространстве – зоне максимального воздействия источника ФП. Сопоставление степени проявления прямых и косвенных эффектов ФП (параметры радиально-концентрического рисунка ФП, степень освещенности, запас лесной подстилки и фитомассы, интенсивность почвенного дыхания, температура и влажность почвы) и корреляционный анализ биогенных и абиогенных параметров позволяют сделать вывод о неоднотипности влияния сосны на молодые почвы угольных отвалов при разной степени увлажнения. Радиально-концентрические рисунки проекции изучаемых фитогенных полей различаются по площади прикроновой зоны. Увеличение площади прикроновой зоны в избыточно увлажненном среднегорно-таежном ЭГР позволяет предполагать, что воздействие сосны на почвы отвалов в этих условиях проявляется более интенсивно.
Исследования различных аспектов взаимодействия в системе “лес – почва” имеют более чем вековую историю (Binkley, 1995). Возросший в последнее время интерес к данной проблеме во многом является следствием развития биогеоценотического направления в почвоведении (Головенко, 1977; Смагин, 1989, 1996; Федорец и др., 2006; Карпачевский и др., 2007; Бобровский, 2010; Лукина и др., 2010; и др.). С позиции данного направления лесной фитоценоз рассматривается как пример мощного “давления жизни” (по В.И. Вернадскому) на биокосную материю. Именно в этих условиях можно ожидать наиболее сильного проявления роли биотического фактора в почвообразовательном процессе, эволюции почв и почвенного покрова (Орловский, 1968; Зонн, 1983; Карпачевский и др., 2007; и др.).
Прижизненное направленное воздействие фитоценозов на среду прежде всего определяется материально-энергетическим метаболизмом (по: Дылис и др., 1964) формирующих его видов растений-эдификаторов. Изменение растением среды тем значительнее, чем крупнее это растение, и чем продолжительнее его влияние (Ястребов, Лычная, 1993; Hokkanen et al., 1995; Кожевников, 1998). Поэтому особое внимание исследователей привлекают деревья, средообразующая роль которых хорошо выражена, усиливается в ходе роста растения и длительно существует (Самойлов, Тархова, 1993; Демьянов, 1996; Журавлёва и др., 2012).
Часть пространства, в пределах которого среда приобретает новые свойства, определяемые присутствием в ней особи конкретного вида, А.А. Уранов (1965) назвал “фитогенное поле” (ФП). Характеристики среды, которые изменяются в отклике на рост и развитие вида-эдификатора, и посредством трансформации, которых источник ФП контролирует видовой состав, продуктивность и пространственное распределение растений подчиненных ярусов, состав и численность беспозвоночных животных, грибов и бактерий, режимы и свойства почв – рассматриваются как эффекты ФП (Уранов, Михайлова, 1974; Кожевников, 1998).
Воздействие дерева на среду распространяется неравномерно в пределах ФП. Наиболее значимое влияние отдельно стоящего дерева, как правило, регистрируется в приствольном и подкроновом пространствах, по мере удаления от ствола напряженность поля ослабевает (Уранов, Михайлова, 1974; Демьянов, 1980; Крышень, 2000; Ипатов, 2007; Ипатов и др., 2009). В структуре ФП, организованного градиентом напряженности воздействия эдификатора, как правило, выделяются радиально-концентрические круги, именуемые тессерами (Лукина и др., 2010), микрозонами (Маслов, 1986; и др.) или зонами (Уфимцев и др., 2016). В сомкнутых насаждениях ФП деревьев перекрываются, а их радиально-концентрическая дифференциация оказывается ограниченной (Крышень, 2000).
Для обозначения участков, в пределах которых проявляется средообразующая роль деревьев, используются и другие термины: “экологическое поле” (Wu et al. 1985; Walker et al., 1989); “сфера средообразующего воздействия” (Работнов, 1984); “эдасфера” (Быков, 1978) и т.д. По мнению большинства авторов, термин, предложенный А.А. Урановым, по сути, имеет тот же экологический смысл, синонимичен, но при этом он получил более широкое распространение и считается удобным (Кожевников, 1998; Крышень, 2000; Ипатов и др., 2009). Возможно, именно поэтому концепция ФП довольно активно развивается в настоящее время, обсуждается и формализуется ее теоретическая основа (Демьянов, 1996; Ястребов, 1996; Кожевников, 1998; Крышень, 2000; Жукова, 2012; Горелов, 2013; и др.). Более детальные сведения по истории и о современном состоянии изучения ФП представлены в работе Е.В. Черняевой и В.П. Викторова (2016).
Изначально концепция привлекалась только для изучения межвидовых взаимодействий растений и горизонтальной структуры фитоценозов, т.е. воздействие источника ФП оценивалось по поведению других видов или вида, так называемых растений-фитомеров (Уранов, Михайлова, 1974; Галанин, 1980; Маслов, 1986; Ястребов, Лычная, 1993). Позднее при изучении фитогенных полей предметом исследования стали и параметры среды (или эффекты ФП), в том числе освещенность, количество осадков и опада, мощность подстилки, температура и влажность почвы, почвенные свойства (кислотность, концентрация обменных оснований, биологическая активность, содержание гумуса, азота и др.), трансформируя которые источник ФП влияет на соседние растения (Самойлов, 1983; Hokkanen et al., 1995; Finzi et al., 1998; Priha et al., 1999; Паркина, 2006; Пуртова, Зимина, 2007; Ипатов, 2007; Ипатов и др., 2009; Лебедева и др., 2009; Chodak, Nikli$\mathop {\text{n}}\limits^{\text{`}} $ska, 2010; Журавлева и др., 2012; Прокушкин и др., 2004; и др.).
Применительно к изучению взаимодействий леса и почвы привлечение концепции ФП нельзя считать чем-то новым. Она перекликается с учением о парцеллярной структуре биогеоценоза, сформулированным Н.В. Дылисом с соавт. (1964) и получившим свое развитие в работах по почвоведению (Карпачевский, 1977, 1981; Ведрова, 1980; Федорец и др., 2006; Карпачевский и др., 2007; Лукина и др., 2010; и др.), а также и с теорией экологического поля (Wu et al., 1985; Walker et al., 1989).
Результаты наиболее всестороннего изучения вопросов влияния леса на почвы, в основном, получены при проведении полевых многолетних экспериментов. Таких экспериментов в мире не много. Наиболее известные из них: эксперименты с тремя древесными породами в Финляндии (Priha et al., 1999); шестью – в Дании (Vesterdal et al., 2008, 2012), четырнадцатью – в Польше (Reich et al., 2005); в России – это 100-летний опыт Н.В. Орловского с шестью породами в условиях одного эдафического фона (Shugalei, 2005; Vedrova, 2005; Меняйло, 2006; Ведрова, Решетникова, 2014; и др.) и эксперимент с географическими культурами сосны обыкновенной, дополнительно используемый для оценки влияния климатипов на трансформацию почвенных свойств (Наумова и др., 2009).
Все упомянутые эксперименты проводятся на почвах, имеющих многовековую историю почвообразования. Как справедливо замечает С.В. Зонн (1983), не всегда известно имеют ли почвы лесной генезис на всeм протяжении своего формирования или стали лесными в результате поселения лесной растительности. Фактически, влияние древесных растений на свойства почв в таких условиях оценивается “от исходного” их состояния. При этом изучается воздействие древесных растений на почвы применительно к отдельно взятым условиям местообитания. В то же время остается не ясным, насколько однозначным окажется действие одного и того же вида растения на материнскую породу и почву в различных климатических условиях? Поставленный еще в работах С.В. Зонна (1954) вопрос не нашел исчерпывающего ответа и по мнению D. Binkley (1995) остается ключевым в развитии исследований взаимодействия компонентов системы “лес – почва”.
Приведенные ниже результаты получены в ходе исследований, где одна и та же древесная порода – сосна обыкновенная (Pinus sylvestris L.) – развивается в разных климатических условиях. Спецификой постановки исследований служит формирование насаждений на почвах техногенных ландшафтов. В отличие от природных аналогов, их называют почвами с практически полностью трансформированной матрицей (Солнцева и др., 1990). Это молодые почвы, для которых процессы формирования “педопамяти” (Таргульян, 2008) только запускаются. При этом, начальная “точка отсчета” в их формировании известна, как и тип растительности, при котором развиваются почвообразовательные процессы.
Перед авторами стоял ряд задач: 1) Оценивая и сопоставляя эффекты фитогенного поля сосны, определить, в пределах какой площади поверхности проявляются влияние источника фитогенного поля; 2) Изучая направленность и интенсивность проявления эффектов фитогенного поля сосны, произрастающей в разных климатических условиях, оценить характер (однотипность или уникальность) влияния породы на почвы.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили на территории Кузнецкой котловины, биогеографическая структура которой представлена чередованием степных, лесостепных и таежных районов. Неоднородность климатических условий здесь определяется меридиональной протяженностью горных систем Салаира и Кузнецкого Алатау (рис. 1), представляющих значимое орографическое препятствие для влажных воздушных масс, поступающих с северо- и юго-запада, и вызывающих конденсацию осадков на наветренных макросклонах (Трофимов, 1975). Таким образом, на сравнительно небольшом расстоянии для проведения исследований доступны различные по климатическим условиям районы (Экологическая карта …, 1995).
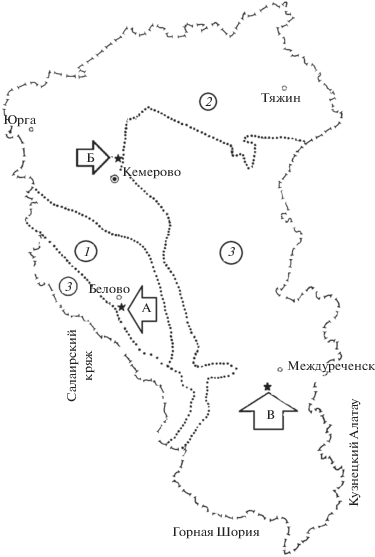
Рис. 1.
Географическое положение опорных полигонов проведения исследований: “Бачатский” (А), “Кедровский” (Б), “Красногорский” (В), расположенных в различных ЭГР: центральном остепненном (1), северном лесостепном (2) и среднегорно-таежном (3).
Кроме того, в пределах Кузнецкой котловины активно ведется добыча угля открытым способом, что предполагает извлечение на дневную поверхность и складирование в отвалы пород, слабо затронутых процессами почвообразования. При восстановлении нарушенных земель в лесохозяйственном направлении отвалы рекультивируются без нанесения потенциально-плодородных и плодородных слоев почвы и, чаще всего, засаживаются культурами сосны обыкновенной (Уфимцев, 2017). Широкое использование древесной породы при искусственном лесовосстановлении на породных отвалах обусловлено еe высокой толерантностью к низкой трофности и слабой влагообеспеченности молодых почв (Уфимцев, 2017).
Таким образом, в пределах Кузнецкой котловины удалось подобрать опорные полигоны, которые характеризуют формирование искусственных сосновых биогеоценозов на слаборазвитых лесных почвах в различных эколого-географических районах (ЭГР): центральный остепненный (54°16′09″ с.ш.; 86°09′00″ в.д.) – “Бачатский”11; северный лесостепной (55°32′33″ с.ш.; 86°04′11″ в.д.) – “Кедровский” и среднегорный таежный (53°42′01″ с.ш.; 88°06′27″ в. д.) – “Красногорский” (рис. 1).
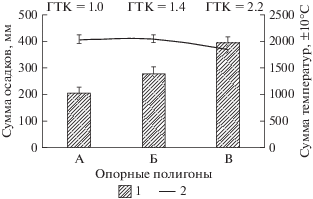
Для характеристики гидротермических условий опорных полигонов (рис. 2) привлекались значения среднесуточной температуры и осадков, фиксируемых на метеостанциях, ближайших к объектам исследования, за период 2012–2017 гг. (архив сайта https://rp5.ru). Анализ полученных параметров (рис. 2) подтверждает выше упомянутую особенность климата Кузнецкой котловины: при близких величинах суммы температур вегетационного периода, наблюдаются существенные различия ЭГР по сумме осадков. Рассчитанный по Г.Т. Селянинову гидротермический коэффициент (ГТК) для засушливых условий центрального остепненного ЭГР составил 1.022, 1.4 – для северного лесостепного – умеренная степень увлажнения и 2.2 – в пределах среднегорно-таежного – избыточное увлажнение.
Рис. 2.
Гидротермические условия опорных полигонов: “Бачатский” (А), “Кедровский” (Б), “Красногорский” (В); 1 и 2 – соответственно, сумма средне многолетних осадков и температур периода вегетации. Приведены средние и их стандартные ошибки.
По результатам маршрутно-рекогносцировочных исследований растительного покрова анализируемых полигонов, подобраны массивы искусственных сосновых насаждений, соответствующие I–II классам бонитета, II классу возраста, I категории жизненного состояния. В пределах каждого из них выбиралось 3 модельных дерева сосны обыкновенной. Таксационные параметры модельных деревьев опубликованы ранее (Уфимцев и др., 2016).
Для исключения мешающего влияния других деревьев исследования проводились на примере фитогенных полей, образованных отдельно стоящими деревьями сосны. Данное допущение считаем оправданным, поскольку исследования направлены (как сказано выше) на изучение эффектов ФП сосны на почвы в различных климатических условиях.
Согласно современной классификации (Классификация …, 2004), с учетом системы таксономических единиц почв, составленной для почв угольных отвалов Кемеровской области (Брагина, 2016), культуры изучаемых опорных полигонов формируются на гумусово-слаборазвитых, натечно-карбонатных, литостратных скелетно-гетерогенных техногенных петрозёмах. По результатам оценки ряда параметров эдафического фона петроземы, формирующиеся в различных ЭГР Кузнецкой котловины, мало различаются между собой. Аналогичные выводы сделаны П.С. Брагиной с соавт. (2014) при изучении слаборазвитых почв в различных районах Кузбасса. Это нейтральные и слабощелочные почвы (рН 7.2–7.9, коэффициент вариации – С.V. = 4%), легко- и среднесуглинистого гранулометрического состава. Содержание валового азота составляет 0.360–0.364%, фосфора и калия – варьирует от 62 ± 11 до 201 ± 40 мг кг–1 (С.V. = 42–48%) и от 6 ± 1 до 15 ± ± 2 мг кг–1 (С.V. = 26–44%) соответственно.
В практике изучения ФП нет единого мнения по процедуре выделения отдельных структурных единиц и их количества (Демьянов, 1980, 1996; Крышень, 2000). В пределах проекции радиально-концентрического рисунка ФП на поверхности почвы мы выделяли три отдельные зоны: подкроновую, прикроновую и внешнюю.
Внешние границы концентрических кругов подкроновой и прикроновой зон сосны визуально хорошо различимы по изменению характеристик напочвенного покрова (видовой состав, обилие и др.), а также градиенту распределения запасов подстилки. Радиусы зон измерялись в северном и южном направлениях. На основании полученных величин по формуле эллипса рассчитывали площади зон. Границу внешней зоны определяли по дальности распространения корней растения-эдификатора (Левичев, 1975; Демьянов, 1980). По данным (Орлов, Кошельков, 1971), горизонтальные корни сосны могут распространятся на расстояние в 3 раза большее радиуса кроны.
Несмотря на то, что по влиянию на почвы древесные растения в значительной степени превосходят травянистые (Журавлева и др., 2012), факт определённого влияния видов напочвенного покрова на свойства поверхностных почвенных горизонтов не отрицается (Бганцова, 1991; Карпачевский и др., 2007; Лебедева и др., 2009). По данным В И. Уфимцева с соавт. (2016) напочвенный покров в пределах ФП изучаемых моделей сосны, несмотря на произрастание в различных ЭГР, характеризуется высокой мерой сходства.
Учитывая, что на начальных этапах эволюции почва находится в определяющей зависимости от внешних факторов, накапливает биогенную энергию и вещество (Смагин, 1996), логично предположить, что влияние сосны на почвы, прежде всего, будет проявляться в специфике радиального перераспределения органического вещества и пространственном градиенте интенсивности биологических процессов.
Чтобы определить (установить) различия в пространственном перераспределении фитомассы напочвенного покрова и детрита по зонам ФП модельных деревьев в различных климатических условиях, на радиальных трансектах в северном и южном направлениях от ствола сосны в трехкратной повторности для каждого направления отбирались укосы с учетной площадки 30 × 60 см. Запасы подстилки сосредоточены только в подкроновой зоне, их также учитывали для северной и южной экспозиции (n = 3) на площадках 30 × 30 см. В лаборатории отобранные образцы высушивали, взвешивали. Запасы фитомассы (в абсолютно сухом веществе) пересчитывали на площадь соответствующих зон ФП.
Интенсивность почвенного дыхания, как интегрального показателя биологических процессов молодых почв проводили методом закрытых (статичных) камер объемом 0.125 м3, площадки-основания для которых устанавливались в каждой зоне ФП в трехкратной повторности. Для предотвращения перегрева камеры ее поверхность покрывали фольгированным теплоизолирующим материалом. Площадка-основание камеры с водяным замком врезалась на глубину не менее 15 см, перед его установкой надземная фитомасса травяно-кустарничкового яруса предварительно скашивалась. На всех опорных полигонах замеры проводились в полдень при одинаковых погодных условиях. В камере устанавливался автоматический логгер-самописец SenseAir (Швеция), фиксирующий температуру воздуха (°C) и концентрацию СО2. Для создания равномерной газовой среды в камере был установлен портативный вентилятор. Время насыщения камеры составляло для каждого опыта 1 ч. В дальнейшем концентрацию СО2 рассчитывали по уравнению Клапейрона–Менделеева с учетом температуры газа непосредственно в момент измерения. Содержание влаги оценивали весовым методом в лаборатории. Для этого отбирались образцы почвы. Температура и влажность почвы фиксировалась для слоя 0–10 см.
Освещенность в пределах проекции ФП измерялась по подзонам Люксметром TESTO 540 (Германия).
Для зависимых количественных переменных (интенсивность почвенного дыхания, запас подстилки и фитомассы) рассчитывались стандартные статистики: среднее, медиана, ошибка среднего значения, стандартное отклонение, коэффициенты вариации.
Статистический анализ проводился с помощью программного пакета PAST (Hammer et al., 2001). Нормальность распределения оценивалась по критерию Шапиро–Уилка. Для этого критерия, а также при проведении однофакторного дисперсионного, рангового корреляционного анализа Спирмена, тестов сравнения дисперсий (Левена) и множественного сравнения средних (Тьюки) использовался уровень достоверности 0.05 (Кобзарь, 2006; McDonald, 2014).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Площадь зон проекции ФП. Согласно классификации Е.Н. Журавлевой с соавт. (2012), изучаемые нами модельные деревья сосны обладают средней силой влияния. Средообразующая роль эдификатора наиболее полно проявляется в пределах подкроновой зоны. Ее площадь в пределах проекции радиально-концентрического рисунка ФП модельных деревьев на поверхности почвы в различных эколого-географических районах оценивается, в целом, близкими величинами (рис. 3). Отмечается тенденция к увеличению площади подкроновой зоны с ростом величины ГТК: 15 м2 – для модельных деревьев в центральном остепненном районе (ГТК = 1.0), 16 м2 – в лесостепном (ГТК = = 1.4) и 17 м2 – в среднегорно-таёжном ЭГР (ГТК = 2.0). Различия между полученными значениями площадей подкроновой зоны не превышают 10%.
Рис. 3.
Схема радиально-концентрической проекции зон фитогенного поля (а – подкроновая, б – прикроновая и в – внешняя) для модельных деревьев изучаемых ЭГР: центральный остепненный (а), северный лесостепной (б) и среднегорный таежный (в). Представлены средние биометрические параметры фитогенных полей в масштабе 1 : 200 с учeтом сторон света (север и юг).
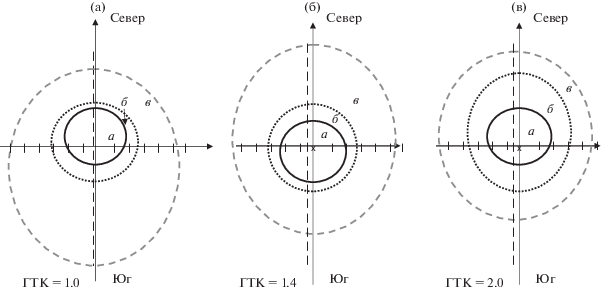
Согласно расчетам, общая площадь проекции радиально-концентрического рисунка ФП изучаемых модельных деревьев составляет: 155 – в остепненном, 141 и 131 м2 в лесостепном и среднегорно-таежном ЭГР, соответственно. Несмотря на то, что величины общей площади ФП являются расчетными, а при их получении привлекалась закономерность, полученная в естественных сосняках, есть основания полагать, что мы не далеки от истины. Так, по данным А.И. Лукьянец (1975), площадь поверхности, занимаемая корневыми системами 12-летних сосен первого бонитета на угольных отвалах севера Свердловской области, составила 108 м2. Раскапывались корневые системы сосен диаметром 10 см, что почти в 2 раза меньше, чем диаметры изучаемых нами модельных деревьев.
В структуре проекции ФП на долю подкроновой зоны приходится от 10 до 13% общей площади. Подкроновую зону отличает наиболее низкий уровень освещенности: от 4 до 12 тыс. лк, что составляет 5–15% от показателей, полученных на открытом пространстве (фоне). Травяной покров здесь сильно разрежен, его общее проективное покрытие варьирует в широких пределах: от 0 до 60%. Доминантами выступают Poa angustifolia L. и Fragaria vesca L.
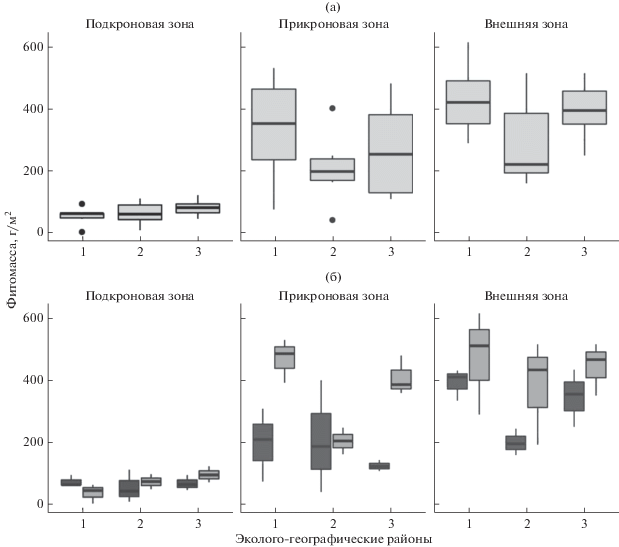
Фитодетрит и фитомасса в пределах проекции ФП. Запас надземной фитомассы напочвенного покрова в пределах подкроновой зоны ФП модельных деревьев одного ЭГР изменяется в широких приделах: от 2 до 92 г м–2 в засушливых условиях центрального остепненного ЭГР; от 7 до 108 и от 61 до 119 г м–2 в умеренно и избыточно увлажненных ЭГР соответственно (рис. 4). По результатам ANOVA средние значения фитомассы подкроновой зоны ФП изучаемых ЭГР статистически не различаются (F = 1.07, p = 0.367). На изменчивость фитомассы травянистого покрова подкроновой зоны экспозиция ФП не влияет (F = = 0.05, p = 0.822).
Рис. 4.
Вариабельность запаса фитомассы напочвенного покрова в зонах фитогенного поля сосен в различных ЭГР: центральный остепненный (1), северный лесостепной (2) и среднегорный таeжный (3). Анализ проведен без учeта (а) и с учетом (б) влияния экспозиции фитогенного поля: темная заливка – север, светлая – юг. Обозначены: медиана-линия внутри коробки; 25 и 75% квартили-границы коробки; минимальные и максимальные значения внутри “ограды” – усы; “⚫” – значения (выбросы), превышающие 1.5 межквартильных расстояния.
Почвы подкроновой зоны, особенно в южной экспозиции ФП, в меньшей степени обеспечены влагой, что, вероятно, обусловлено более низким (на 25–40% по сравнению с открытым пространством, вне ФП) количеством поступающих осадков. Определенный вклад в иссушение поверхностных слоев вносит и корневая система подроста сосны. Несмотря на то, что он угнетен, его высота ограничивается высотой прикрепления кроны (10–50 см), густота подроста может достигать 50 экз. м–2.
Индикаторным признаком подкроновой зоны является сплошной, довольно мощный (5–15 см) слой лесной подстилки, сформированной преимущественно за счет опада сосны. Морфологически подстилка подразделяется на 2 подгоризонта: опада, на долю которого приходится 30–40%, и ферментации. Подгоризонт гумификации не имеет сплошного распространения, его относительное участие не превышает 10%. Фитогенные поля модельных деревьев эколого-географических районов достоверно различаются по запасу подстилки (F = 7.43, p = 0.006) на поверхности почвы, при этом средние оценки запаса снижаются с увеличением величины ГТК (рис. 5).
Рис. 5.
Вариабельность запаса подстилки в подкроновой зоне фитогенных полей модельных деревьев, произрастающих в центральном остепненном (1), северном лесостепном (2) и среднегорном таёжном ЭГР (3). Обозначены: медиана-линия внутри коробки; 25 и 75% квартили-границы коробки; минимальные и максимальные значения внутри “ограды” – усы.
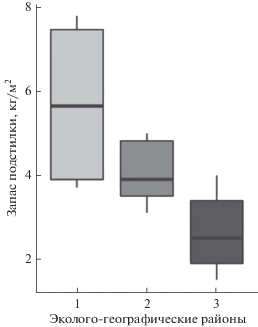
Масса растительных остатков подстилки увеличивается по мере приближения к источнику ФП. Так, в условиях центрального остепненного ЭГР запас подстилки у границы подкроновой зоны составляет 3.7–3.8 кг м–2, а на учетных площадках, расположенных вблизи ствола сосны – 7.6–8.0 кг м–2. В северном лесостепном ЭГР при умеренной степени увлажнения запас подстилки изменяется по направлению к стволу от 3.1–3.5 до 5 кг м–2, а в пределах избыточно увлажненного среднегорно-таежного ЭГР (ОП “Красногорский”) – от 1.5–1.8 до 3.6–4.0 кг м–2.
Изменчивость запаса подстилки значимо и с высокой силой связи коррелирует с температурой (rs = –0.85, p = 0.007) и влажностью (rs = – 0.87, p = = 0.005) почвы подкроновой зоны ФП, что может указывать на значимую роль сосны в регулировании процессов аккумуляции растительных остатков лесной подстилки.
Полученные средние показатели запаса подстилки в подкроновой зоне ФП модельных деревьев согласуются с данными литературных источников, характеризующих накопление массы органогенного горизонта почв в искусственных сосняках II класса возраста. Так, для культур сосны, формирующихся на отвалах Назаровского угольного разреза, запасы подстилки изменяются в пределах от 1.3 до 3.3 кг м–2 (Шугалей, Чупрова, 2015). В опытном Серебряноборском лесничестве (Московская область) запасы органогенного горизонта в мертвопокровной парцелле достигали 3–3.5 кг м–2 (Бганцова, 1983). В лесостепной зоне Приобья на одном квадратном метре поверхности почвы сосредоточено от 2.3 до 3.0 кг растительных остатков (Габеев, 1990). Аналогичные показатели, но для естественных 20–летних сосновых молодняков Енисейской равнины (средняя тайга) составляют 1.6–3.8 кг м–2 (Трефилова, Ведрова, 2018). Наши оценки выше величин, полученных А.В. Почикаловым с соавт. (2015) в 15–33-летних культурах на отвалах фосфоритового рудника Хорловского месторождения (Московская область) – 0.3–0.8 кг м–2, М.Г. Семечкиной (1983) – 0.6 кг м–2 для сосновых молодняков разнотравного типа в лесостепной зоне Средней Сибири и А.С. Курочкиным (1983) для Ленинградской области – от 0.5 до 2 кг м–2. Различия могут быть обусловлены спецификой как условий биотопа, так и методических подходов, используемых при учете запаса подстилки.
Вторая зона ФП – прикроновая, простирается от границы прямой проекции края кроны модельного дерева на 0.8–3 м. В центральном остепненном и северном лесостепном ЭГР на долю прикроновой зоны приходится 11–13% общей площади проекции ФП. В условиях избыточного увлажнения среднегорно-таежного ЭГР прикроновая зона занимает уже 31% ФП, а ее площадь в 2.4 и 2.2 раза больше аналогичных показателей в ОП “Бачатский” и “Кедровский”, соответственно.
Согласно опубликованным данным (Волокитина, 1979) и результатам собственных наблюдений (Уфимцев и др., 2016) на край кроны сосны, как правило, приходятся участки более высокой концентрации атмосферных осадков. Здесь количество осадков достигает 150–180% от фоновых значений, полученных для северной и 100–120% – для южной экспозиции. Условия повышенного увлажнения, вероятно, способствовали развитию мохового покрова – общее проективное покрытие яруса в пределах прикроновой зоны достигает 100%. В числе доминантов выступают три вида: Ceratodon purpureus (Hedw.) Brid., Polytrichum juniperinum Hedw. и Brachythecium salebrosum (F.Weber & D.Mohr) Bruch et al.
На фоне увеличения освещенности прикронового пространства до 35–68 тыс. лк, более активно развивается и травянистый покров. Количество видов, формирующих покров, не изменяется – 7–15, преобладает Melilotus officinalis (L.) Pall. (общее проективное покрытие составляет 70–100%). Средние величины запаса надземной фитомассы в прикроновой зоне по сравнению с подкроновой возрастают в 3–6 раз. Однако по результатам ANOVA средние значения фитомассы прикроновых зон ФП, формирующихся в различных ЭГР, статистически не различаются (F = 1.04, p = 0.379). При анализе изменчивости фитомассы в прикроновой зоне отчетливо проявляется влияние экспозиции ФП (F = 15.69, p = 0.002, см. рис. 4).
Подрост в прикроновой зоне единичный – 1–3 шт. м–2, в большинстве своем (60–80%) сомнительный, разновозрастный. Высота молодых сосен изменяется от 15 до 140 см.
По мере перехода во внешнюю зону ФП модельных сосен, влияние эдификатора ослабевает, освещенность возрастает до фонового уровня 66–81 тыс. лк, проективное покрытие мохового покрова снижается до 10–40%, активно развиваются подрост и травяной покров. Преобладающая часть подроста (80–100%) является жизнеспособной. В зависимости от возраста высота деревьев подроста может изменяться в пределах от 60 до 460 см. Видовой состав травяного покрова представлен 17–25 видами, из них в пределах центрального остепненного ЭГР доминируют Calamagrostis epigeios (L.) Roth и Poa angustifolia, в северном лесостепном и среднегорно-таежном ЭГР – Dactylis glomerata L. и Centaurea scabiosa L., Melilotus officinalis имеет повсеместное распространение и встречается во всех ЭГР, однако его общее проективное покрытие ниже, чем в прикроновой зоне, и не превышает 10–30%.
По сравнению с прикроновой зоной средние запасы надземной фитомассы напочвенного покрова внешней зоны увеличиваются на 28–40%. Внешние зоны ФП модельных деревьев, формирующихся в различных климатических условиях, схожи по среднему запасу надземной фитомассы напочвенного покрова (F = 2.16, p = 0.15). Из числа факторов, определяющих пространственную изменчивость запаса надземной фитомассы, можно выделить экспозицию ФП (F = 5.04, p = 0.044). Масса напочвенного покрова в южной части внешней зоны изучаемых ФП выше значений, полученных для северной экспозиции (рис. 4).
Необходимо отметить, что отсутствие статистически значимых различий в средних значениях надземной фитомассы напочвенного покрова всех зон ФП сопоставляемых опорных полигонов является дополнительным подтверждением близкой трофности эдафического фона отвалов.
Принимая во внимание, что практически вся надпочвенная фитомасса травяного покрова в течение вегетационного сезона постепенно перейдет в состав фитодетрита, а продуктивность травяных сообществ в пределах ФП контролируется модельным деревом-эдификатором, мы попытались сравнить ЭГР по общей массе отмерших растительных остатков, взяв за основу величины запаса фитомассы и площадь соответствующей зоны. Согласно полученным данным, в пределах ФП центрального остепненного ЭГР сосредоточено 161 кг растительных остатков, как поступающих со свежими порциями опада, так и ранее накопленных в составе лесной подстилки. Для северного лесостепного ЭГР эта величина равна 101, а в среднегорном таежном – 85 кг. Примерно половина (55–67%) данного количества сосредоточена в пределах подкроновой зоны, т.е. в наибольшей степени подконтрольно влиянию дерева.
Интенсивность почвенного дыхания. Результаты сопоставления оценок интенсивности почвенного дыхания (ИПД) по ЭГР (рис. 6) указывают на значительный уровень их дифференциации в подкроновой (F = 97.7, p < 0.001) и прикроновой зонах (F = 10.73, p = 0.001) ФП модельных деревьев. Средние значения ИПД внешней зоны ФП в изучаемых ЭГР отличаются меньшим варьированием значений и слабыми общими (на уровне тенденции) различиями (F = 2.71, p = 0.099). Полученные численные результаты сравнений хорошо согласуются с визуальной оценкой различий (рис. 6): наиболее высокие средние показатели ИПД зарегистрированы в подкроновой и прикроновой зонах ФП сосен северного лесостепного ЭГР.
Рис. 6.
Вариабельность интенсивности почвенного дыхания в зонах проекции фитогенного поля сосен в центральном остепненном (1), северном лесостепном (2) и среднегорном таeжном ЭГР (3). Анализ проведен без учёта (а) и с учетом (б) влияния экспозиции фитогенного поля: темная заливка – север, светлая – юг. Обозначены: медиана-линия внутри коробки; 25 и 75% квартили-границы коробки; минимальные и максимальные значения внутри “ограды” – усы; “⚫” – значения (выбросы), превышающие 1.5 межквартильных расстояния.
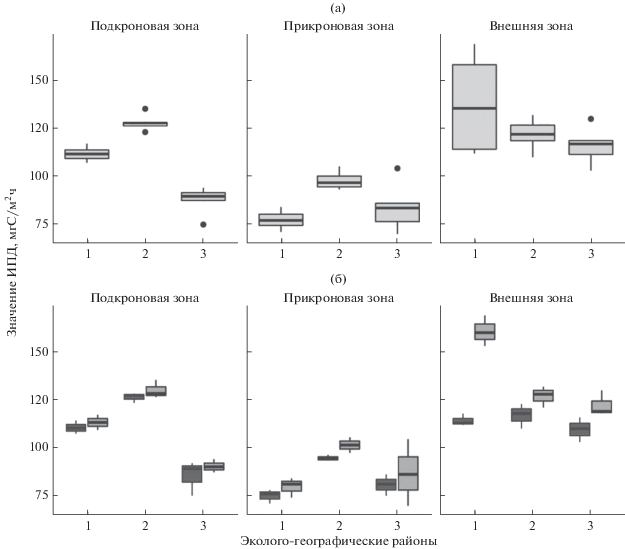
Значимое влияние экспозиции на изменчивость ИПД отсутствует как в подкроновой (F = = 2.53, p = 0.138), так и прикроновой (F = 2.39, p = = 0.148) зонах ФП анализируемых эколого-географических районов. Во внешней зоне, где влияние ФП существенно сглаживается, отмечается сильное воздействие экспозиции на изменчивость значений ИПД (F = 60.69, p < 0.001).
Межзональные изменения ИПД в изучаемых эколого-географических районах имеют схожий, “U-образный” характер: величины почвенного дыхания постепенно снижаются по мере перехода от подкроновой к прикроновой зоне и вновь возрастают во внешней зоне. Механизм пространственной дифференциации ИПД в пределах ФП модельных деревьев пока не ясен. Полагаем, что данное явление может быть обусловлено различиями подкроновой и прикроновой зон ФП по массе и биохимическому составу разлагающегося материала, а также развитием мохового покрова в пределах более обеспеченной атмосферными осадками прикроновой зоны. Более низкие показатели ИПД на участках, покрытых моховым покровом, по сравнению с травяным, получены и в работе (Sedia et al., 2005).
В целом, полученные оценки ИПД согласуются с литературными данными. Интенсивность потока СО2 для молодых почв 25-летних отвалов Назаровского угольного разреза (Красноярский край, лесостепная зона) изменяется в пределах от 70 ± 21 до 107 ± 35 мг С м–2 ч–1 (Наумов, 2009). Аналогичный показатель для отвалов Хорловского месторождения фосфотитов под 22–25-летними культурами сосны в первый год проведения исследования в среднем составил – 202 ± 75, а в следующий – только 19 ± 4 мг С м–2 ч–1 (Почикалов и др., 2015). Немногим выше оказались показатели ИПД в приспевающих сосняках Ангаро-Енисейского региона, формирующихся на дерново-подзолистой и серой лесной оподзоленной почвах: 128 и 145 мг С м–2 ч–1 соответственно (Сорокин и др., 1985). В округе Брадфорд штата Флорида (США) на бедных песчаных почвах, под 9- и 29-летними плантациями Pinus palustris Mill. средний за период вегетации поток углекислоты достигал 99 ± 14 и 135 ± 20 мг С м–2 ч–1 соответственно (Ewel et al., 1986). По данным июля, ИПД в 16-летнем разреженном разнотравно-злаковом сосняке на постагрогенном агроземе иллювиально-железистом в течение двух лет наблюдения изменялась в пределах 120–180 мг С м–2 ч–1 (Люри и др., 2013).
Корреляционный анализ данных ИПД показал наличие средней положительной связи между изменчивостью значений интенсивности почвенного дыхания прикроновой и подкроновой (rs = 0.55, p = 0.014), а также внешней и прикроновой (rs = 0.18, p = 0.003) и внешней и подкроновой (rs = 0.30, p = 0.036) зон ФП. Из этого следует, что регистрируемый уровень ИПД регулируется воздействием одного определенного вида-эдификатора. Таким образом, есть основания полагать, что интенсивность почвенного дыхания, в данном случае может рассматриваться как интегральный эффект ФП, позволяющий на более “тонком” уровне оценить влияние деревьев на техногенные молодые почвы.
Выводы: 1. На основании полученных данных, можно утверждать, что эффекты ФП отдельно стоящих модельных деревьев сосны (II класса возраста), со средней силой влияния (по: Журавлева и др., 2012) распространяются как минимум в пределах 131–155 м–2 и только 10–13% данной площади находится в подкроновом пространстве – зоне максимального воздействия источника ФП. Радиально-концентрические рисунки изучаемых фитогенных полей различаются по площади прикроновой зоны. Увеличение площади прикроновой зоны в избыточно увлажненном среднегорном таежном ЭГР позволяет предполагать, что воздействие сосны на почвы отвалов в этих условиях проявляется более интенсивно.
2. Запас лесной подстилки в подкроновой зоне убывает по мере увеличения степени увлажнения: от 89 кг при ГТК = 1.0 до 45 кг – ГТК = 2.2. Сильная отрицательная корреляционная связь изменчивости запаса подстилки с температурой в подкроновой и влажностью в подкроновой и прикроновой зонах ФП указывает на значимую роль сосны в регулировании процессов аккумуляции растительных остатков.
3. Зоны ФП модельных деревьев, формирующихся в различных ЭГР, схожи по среднему запасу надземной массы напочвенного покрова. В определении пространственной изменчивости запаса ведущая роль отводится экспозиции ФП, которая четко проявляется только в прикроновой и внешней зонах. Фитомасса напочвенного покрова в южной части зон выше значений, полученных для северной экспозиции.
4. Общей закономерностью является снижение разброса показателей интенсивности почвенного дыхания по мере удаления от источника фитогенного поля. Наиболее высокие показатели интенсивности почвенного дыхания зарегистрированы в северном лесостепном эколого-географическом районе (условия умеренного увлажнения).
5. Приведенный анализ эффектов ФП показал, что характер влияния сосны на молодые почвы отвалов, формирующиеся в условиях увлажнения различной степени, при определенном сходстве параметров нельзя считать полностью однотипным.
Список литературы
Бганцова В.А. Связь травяного покрова с запасом подстилки в сосновых культурах. // Роль подстилки в лесных биогеоценозах. М.: Наука, 1983. С. 19.
Бганцова В.А. Влияние травянистых растений на свойства почвы в лесном БГЦ // Почвоведение. 1991. № 10. С. 60–78.
Бобровский М.В. Лесные почвы Европейской России: биотические и антропогенные факторы формирования М.: Товарищество научных изданий КМК, 2010. 359 с.
Брагина А.С. Почвообразование на отходах горнодобывающих предприятий Кемеровской области: Автореф. дис. … канд. биол. наук: 25.00.23. М. 2016. 23 с.
Брагина П.С., Цибарт А.С., Завадская М.П., Шарапова А.В. Почвы на отвалах вскрышных пород в лесостепной и горно-таёжной зонах Кузбасса // Почвоведение. 2014. № 7. С. 878–889.
Быков Б.А. Геоботаника. Алма-Ата: Наука, 1978. 288 с.
Ведрова Э.Ф. Влияние сосновых насаждений на свойства почв. Новосибирск: Наука, 1980. 104 с.
Ведрова Э.Ф., Решетникова Т.В. Масса подстилки и интенсивность ее разложения в 40-летних культурах основных лесообразующих пород Сибири // Лесоведение. 2014. № 1. С. 42–50.
Волобуев В.Р. Экология почв (очерки). Баку: Изд-во Акад. наук АзССР, 1963. 260 с.
Волокитина А.В. Особенности распределения осадков под пологом хвойного леса // Лесоведение. 1979. № 2. С. 40–48.
Габеев В.Н. Экология и продуктивность сосновых лесов. Новосибирск: Наука, 1990. 229 с.
Галанин А.В. Влияние фитогенного поля на распределение растений // Экология. 1980. № 6. С. 76–78.
Головенко С.В. Экологическое почвоведение-предмет и метод // Методология и методика почвенных и ландшафтно-геохимических исследований. М.: Изд-во МГУ, 1977. С. 25–37.
Горелов А.М. Фитогенное поле и его структура // Электронный журн. “Вестник Московского государственного областного университета”. 2013. № 1. С. 1–9.
Демьянов В.А. Анализ ценотической роли лиственницы Гмелина на крайнем северном пределе распространения древесной растительности (Таймыр) // Ботанический журнал. 1980. Т. 65. № 7. С. 926–937.
Демьянов В.А. Представление о фитогенном поле растений и проблема сущности фитоценоза // Известия РАН. Серия биологическая. 1996. № 3. С. 359–363.
Дылис Н.В., Уткин А.И., Успенская И.М. О горизонтальной структуре лесных биогеоценозов // Бюллетень Московского общества испытателей природы. Отдел биологический. 1964. Т. 30. Вып. 4. С. 65–72.
Жукова Л.А. Концепция фитогенных полей и современные аспекты их изучения // Известия Самарского НЦ РАН. 2012. Т. 14. № 1(6). С. 1462–1465.
Журавлева Е.Н., Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю. Изучение растительности на лугах под влиянием сосны обыкновенной (Pinus sylvestris L.) // Вестник Санкт-Петербургского университета. 2012. Вып. 2. Сер. 3. С. 3–12.
Зонн С.В. К вопросу о взаимодействии лесной растительности с почвами // Почвоведение. 1954. № 4. С. 51–60.
Зонн С.В. Современные проблемы генезиса и географии почв. М.: Наука, 1983. 168 с.
Ипатов В.С. Фитогенные поля одиночных деревьев некоторых пород в одном экотопе // Ботанический журн. 2007. Т. 92. № 8. С. 1186–1192.
Ипатов В.С., Журавлёва Е.Н., Лебедева В.Х., Тиходеева М.Ю. Фитогенное поле Picea abies, P. obovate (Pinaceae) // Ботанический журн. 2009. Т. 94. № 4. С. 558–568.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во МГУ, 1977. 312 с.
Карпачевский Л.О. Лес и лесные почвы. М.: Лесная промыш-сть, 1981. 264 с.
Карпачевский Л.О., Зубкова Т.А., Ташнинова Л.Н., Руденко Р.Н. Почвенный покров и парцеллярная структура лесного биогеоценоза // Лесоведение. 2007. № 6. С. 107–113.
Классификация и диагностика почв России / Сост. Л.Л. Шишов, В.Д. Тонконогов, И.И. Лебедева, М.И. Герасимова. Смоленск: Ойкумена, 2004. 342 с.
Кобзарь А.И. Прикладная математическая статистика. М.: Физматлит, 2006. 816 с.
Кожевников Ю.П. О концепции фитогенного поля // Известия АН. Серия биологическая. 1998. № 3. С. 356–362.
Крышень А.М. Фитогенное поле: теория и проявление в природе // Известия АН. Серия биологическая. 2000. № 4. С. 437–443.
Курочкин А.С. Динамика запасов и состав лесной подстилки в плантационных культурах сосны обыкновенной. Роль подстилки в лесных биогеоценозах. М.: Наука, 1983. С. 108.
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. Фитогенное поле герани лесной Geranium sylvaticum (Geraniaceae) // Ботанический журн. 2009. Т. 94. № 6. С. 833–847.
Левичев И.Г. О динамике травянистых микрогруппировок в фисташниках Бадхыза (Туркменская ССР) // Ботанический журн. 1975. Т. 60. № 7. С. 969–976
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва–растительность //Лесоведение. 2010. № 5. С. 45–56.
Лукьянец А.И. Архитектоника и фитомасса корневой системы естественных сосняков на железнодорожных отвалах Богославского и Веселовского месторождений бурого угля // Рекультивация земель, нарушенных при добыче полезных ископаемых: Тез. докл. координационного совещания. Тарту, 1975. С. 151–157.
Люри Д.И., Карелин Д.В., Кудиков А.В., Горячкин С.В. Изменение почвенного дыхания в ходе постагрогенной сукцессии на песчаных почвах в южной тайге // Почвоведение. 2013. № 9. С. 1060–1072.
Маслов А.А. О взаимодействии фитогенных полей деревьев в сосняке чернично-брусничном // Ботанический журн. 1986. Т. 71. № 12. С. 1646–1652.
Меняйло О.В. Влияние древесных пород Сибири на образование и потребление N2O // Известия РАН. Серия биологическая. 2006. № 5. С. 606–612.
Наумов А.В. Дыхание почв: составляющие, экологические функции, географические закономерности. Новосибирск: Изд-во СО РАН, 2009. 208 с.
Наумова Н.Б., Макарикова Р.П., Тараканов В.В., Кузьмина Н.А., Новикова Т.Н., Милютин Л.И. Влияние климатипов сосны обыкновенной на некоторые химические и микробиологические свойства почв // Сибирский экологический журн. 2009. № 2. С. 287–292.
Орлов А.Я., Кошельков С.П. Почвенная экология сосны. М.: Наука, 1971. 323 с.
Орловский Н.В. Послесловие. Лес и почва: Тр. Всесоюз. научн. конф. по лесному почвоведению. Красноярск: Красноярское книж. издательство, 1968. С. 593.
Паркина И.Н. Особенности биологической активности почвы в фитогенном поле березы повислой // Вестник Самарского государственного университета. Естественнонаучная серия. 2006. № 7(47). С. 148–152.
Почикалов А.В., Ларин Я.А., Арешин А.В., Карелин Д.В. Компоненты бюджета углерода в лесных посадках при рекультивации открытых горных выработок // Лесоведение. 2015. № 6. С. 447–457.
Прокушкин С.Г., Бугаенко Т.Н., Сорокин Н.Д., Каверзина Л.Н., Зырянова О.А. Роль фитогенного поля лиственницы допожарной генерации в формировании эдафических условий на гари // Известия АН. Серия биологическая. 2004. № 1. С. 49–58.
уртова Л.Н., Зимина М.П. Изменчивость физико-химических показателей бурых лесных почв в пределах фитогенных полей деревьев (юг Дальнего Востока) // Почвоведение. 2007. № 1. С. 31–37
Работнов Т.А. О конкуренции между растениями в растительных сообществах // Бюллетень Московского общества испытателей природы Отдел биологический. 1984. Т. 89. Вып. 5. С. 82–93.
Самойлов Ю.И. Структура фитогенного поля на примере одиночных дубов Quercus robur (Fagaceae) // Ботанический журн. 1983. Т. 68. № 8. С. 1022–1034.
Самойлов Ю.И., Тархова Т.Н. Анализ фитогенных полей одиночных деревьев с использованием метода главных компонент // Ботанический журн. 1993. Т. 78. № 5. С. 61–77.
Семечкина М.Г. Запасы подстилки в сосняках разнотравных островных лесостепей Средней Сибири. Роль подстилки в лесных биогеоценозах. М.: Наука, 1983. С. 182.
Смагин А.В. Почва как результат самоорганизации биогеоценоза // Доклады АН СССР. 1989. Т. 308. № 3. С. 729–731.
Смагин А.В. Биогеоценотическое направление в почвоведении // Почвоведение. 1996. № 3. С. 298–309.
Солнцева Н.П., Герасимова М.И., Рубилина Н.Е. Морфогенетический анализ техногенно-преобразованных почв // Почвоведение. 1990. № 8. С. 124–129.
Сорокин Н.Д., Горбачев В.Н., Гиголян Д.К. Микробоценозы и биологическая активность лесных почв Ангаро-Енисейского региона // Биологическая активность лесных почв. Красноярск: Институт леса и древесины, 1985. С. 35.
Таргульян О.В. Память почв: общие подходы к проблеме // Память почв: почвы как память биосферно-геосферно-антропосферных взаимодействий. Отв. ред. Таргульян О.В., Горячкин С.В. М.: Изд-во ЛКИ. 2008. С. 24–57.
Трефилова О.В., Ведрова Э.Ф. Минерализационный поток углерода в постпирогенных сосняках Среднего Енисея // Лесоведение. 2018. № 3. С. 210–224.
Трофимов С.С. Экология почв и почвенные ресурсы Кемеровской области. Новосибирск. 1975. 300 с.
Уранов А.А. Фитогенное поле. Проблемы современной ботаники. М. Л: Наука. 1965. Т. 1. С. 251–254.
Уранов А.А., Михайлова Н.Ф. Из опыта изучения фитогенного поля Stipa pennata L. // Бюллетень Московского общества испытателей природы. Отдел биологический. 1974. T. LXXIX(5). С. 151–159.
Уфимцев В.И. Опыт и современное состояние лесной рекультивации в Кузбассе // Сибирский лесной журн. 2017. № 4. С. 12–27.
Уфимцев В.И., Беланов И.П., Куприянов О.А. Эколого-ценотическая роль фитогенных полей сосны обыкновенной на отвалах угольной промышленности // Сибирский экологический журн. 2016. № 1. С. 164–175.
Федорец Н.Г., Морозова Р.М., Солодовников А.Н. Пространственная изменчивость почвенного покрова в сосняке лишайниковом // Лесоведение. 2006. № 3. С. 64–79.
Черняева Е.В., Викторов В.П. История и современное состояние изучения фитогенных полей // Социально-экологические технологии. 2016. № 1. С. 89–105.
Шугалей Л.С., Чупрова В.В. Почвообразование в техногенных ландшафтах. Красноярск: Красноярский гос. аграрный ун-т, 2015. 239 с.
Экологическая карта Кемеровской области. Масштаб 1 : 500 000. М.: Федеральная служба геодезии и картографии России, 1995. Л. 1.
Ястребов А.Б. Напряженность фитогенных полей деревьев в лишайниково-зеленомошных сосняках // Экология. 1996. № 1. С. 3–9.
Ястребов А.Б., Лычная Н.В. Исследование фитогенных полей деревьев в лишайниково-зеленомошных сосняках // Ботанический журнал. 1993. Т. 78. № 5. С. 78–92.
Binkley D. The influence of tree species on forest soils: Processes and patterns // Proceeding of the Trees and Soil Workshop. Eds.: Mead D.J., Cornforth I.S. Canterbury: Lincoln University Press, 1995. P. 1–33.
Chodak M., Nikli$\mathop {\text{n}}\limits^` $ska M. The effect of different tree species on the chemical and microbial properties of reclaimed mine soils // Biology & Fertility of Soils 2010. V. 46. P. 555–566.
Ewel K.C., Cropper W.P., Jr., Gholz H.L. Soil CO2 evolution in Florida slash pine plantations. I. Changes through time // Canadian J. Forest Research. 1986. V. 17. P. 325–329.
Finzi A.C., Canham C.D., Breemen N.V. Canopy tree-soil interactions within temperate forests: species effects on pH and cations // Ecology Applications. 1998. № 8(2). P. 447–454.
Hammer Ø., Harper D. A. T., Ryan P. D. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. V. 4. № 1. P. 9.
Hokkanen T.J., Järvinen E., Kuuluvainen T. Properties of top soil and relationship between soil and trees in a boreal Scots pine stand// Silva Fennica. 1995. V. 29(3). P. 189–203.
McDonald J.H. Handbook of Biological Statistics, 3rd ed. Sparky House Publishing, Baltimore, Maryland: 2014. 299 p.
Priha O., Lehto T., Smolander A. Mycorrhizas and C and N transformations in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings // Plant and Soil. 1999. V. 206. P. 191–204.
Reich P.B., Oleksyn J., Mrozinski P., Hobbie S.E., Eissenstat D.M., Chorover J., Chadwick O.A., Hale C.M., Tioelker M.G. Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species // Ecology Lettets. 2005. V. 8. № 8. P. 811–818.
Sedia E.G., Ehrenfeld J.G. Differential effects of lichens, mosses and grasses on respiration and nitrogen mineralization in soils of the New Jersey Pinelands // Oecologia. 2005. V. 144. № 1. P. 137–147.
Shugalei L.S. The Siberian afforestation experiment: history, methodology, and problems // Tree species effects on soil: implications for global change: Proceedings of the N-ATO Advanced Research Workshop on Trees and Soil Interactions, Implications to Global Climate Change August 2004 Krasnoyarsk, Russia. Series IV: Earth environmental sciences / Ed. by D. Binkley, O. Menyail. Amsterdam: IOS Press and Springer 2005. 2005. P. 257–268.
Vedrova E.F. Biochemistry of carbon and nitrogen in the Siberian afforestation experiment // Tree species effects on soil: implications for global change: Proceedings of the NAT-O Advanced Research Workshop on Trees and Soil Interactions, Implications to Global Climate Change August 2004 Krasnoyarsk, Russia. Series IV: Earth environmental sciences / Ed. by D. Binkley, O. Menyail. Amsterdam: IOS Press and Springer 2005. P. 257–268.
Vesterdal L., Elberling B., Christiansen J.R., Callesen I., Schmidt I. K. Soil respiration and rates of soil carbon turnover differ among six common European tree species // Forest Ecology and Menegement. 2012. V. 264. P. 185–196.
Vesterdal L., Schmidt I. K., Callesen I., Nilsson L.O., Gundersen P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species // Forest Ecology & Menegement. 2008. V. 255. P. 35–48.
Walker J., Sharpe P.J.H., Penridge L.K., Wu H. Ecological field theory: the concept and field tests // Vegetation. 1989. V. 83. P. 81–95.
Wu H., Sharpe P.J.H., Walker J., Penridge L.K. Ecological field theory (EFT): A spatial analysis of resource interference among plants // Ecological Modelling. 1985. № 29. P. 215–243.
Дополнительные материалы отсутствуют.