

Лесоведение, 2021, № 3, стр. 318-324
Восстановительная динамика населения жужелиц в лесных культурах южной тайги
Е. Д. Коробов *
ФГБУ “Центрально-Лесной государственный заповедник”
172521 Нелидовский район, Тверская область, пос. Заповедный, Россия
* E-mail: edkorobov@yandex.ru
Поступила в редакцию 24.12.2018
После доработки 04.02.2019
Принята к публикации 08.12.2020
Аннотация
Приводятся многолетние данные по сукцессионной динамике населения жужелиц в еловых культурах южной подзоны тайги (Тверская область). Восстановительный потенциал искусственных насаждений оценивается для лесов неморального типа с затрудненным естественным возобновлением ели. В качестве тестового показателя используется ранговая структура комплексов жужелиц. Установлено, что активное восстановление видовых популяций жужелиц идет на уровне 20-летнего возраста еловых посадок и, как правило, совпадает с волнами общего для территории роста численности этих видов. В дальнейшем снижается скорость изменений ранговой структуры и стабилизируется состав доминантной группы видов. Между 15 и 25 годами возраста культур наблюдается быстрое приближение порядков доминирования видов к характеристикам ненарушенных южно-таежных ельников. Сравнение с естественным возобновлением леса на сплошном ветровале показывает, что после ветровала население жужелиц сохраняется намного лучше, чем на вырубке. Однако к 20-летнему возрасту скорость восстановления в еловых культурах компенсирует это различие.
Возобновление еловых лесов на сплошных вырубках с помощью искусственных насаждений – распространенная практика ведения лесного хозяйства на территории южной тайги. По очевидным причинам лесозаготовкам прежде всего подвергается зональный южно-таежный тип – высокопродуктивные ельники сложные. Однако в лесах именно этой категории естественное возобновление коренной хвойной породы ослаблено, проходит длительный цикл, и поэтому создание лесных культур почти не имеет альтернативы (Побединский, 1973; Старостина, 1973).
Исследовательская тематика, которая складывается вокруг искусственного возобновления леса на вырубках, в основном развивается по двум направлениям. Значительно большая ее часть относится к лесохозяйственным вопросам эффективного и ускоренного создания фондов для лесной промышленности. С другой стороны, еловые культуры привлекают внимание исследователей как модельный объект сукцессионной восстановительной динамики, включая развитие во времени различных компонентов биоты лесных экосистем.
К модельным свойствам монокультур ели можно отнести упрощенность состава и ценотической структуры по сравнению с естественными лесами, такими, например, как сложные, неморального типа ельники южной тайги. При этом в экологических исследованиях возникает круг вопросов, связанный с сохранением биоразнообразия и формированием полноценной структуры сообществ в искусственных лесах. Еще одно модельное свойство еловых посадок – то, что они в ускоренном и усиленном режиме воспроизводят конечную стадию восстановительной сукцессии – хвойный лес и, таким образом, возникает возможность проследить в сжатом формате детали восстановительных процессов.
ОБЪЕКТЫ И МЕТОДИКА
В Центрально-Лесном заповеднике ведутся многолетние наблюдения за динамикой лесных южно-таежных экосистем по программе экологического мониторинга. В рамках этой программы сформированы длительные ряды непрерывных ежегодных данных по лесным площадям, подвергшимся естественным и антропогенным катастрофическим нарушениям. Основной тестовый объект – структура сообществ и динамика популяций жужелиц, группы жесткокрылых с признанными свойствами универсального биоиндикатора. Неоднократно изучалось влияние на жужелиц рубок и возобновления леса на вырубках (Арнольди, Матвеев, 1973; Huhta, 1979; Koivula, 2002; Sklodowski, 2006). Как правило, эти работы основаны на подборе пространственного ряда площадей, имитирующего возрастную динамику. Соответственно, этот метод дает фиксацию состояний объекта на возрастных этапах, но остаются неизвестными детали развития во времени между этапами и перехода из одного состояния в другое. Существует также зависимость локальных сукцессий от общих для территории динамических процессов, которая выявляется только в многолетних рядах наблюдений. Многолетние данные обнаруживают затяжные периоды общего для территории роста или спада популяций отдельных видов, часто не имеющие очевидных объяснений.
В качестве тестового показателя для оценки восстановительной динамики в настоящем сообщении использовалась ранговая структура населения жужелиц. Порядок доминирования видов – надежный показатель восстановления при интенсивном замещении видов в ходе сукцессии. Общность подхода при замене значений обилия на ранги здесь компенсируется малой зависимостью от резких колебаний обилия массовых видов и сравнительной простотой расчетов (Песенко, 1982). Изменения ранговой структуры во времени анализировались на основе расчета полной матрицы коэффициентов ранговой корреляции Спирмена и ее обработкой методом неметрического многомерного шкалирования в программе Statistica. Устойчивость состава доминантной группы видов жужелиц оценивалась с помощью индекса общности Жаккара.
Площадь исследований – ситниково-кипрейная вырубка 1979 г. ельника неморально-кисличного. По технологии проведения вырубка сплошная, летняя, с использованием тяжелой лесозаготовительной техники. Поверхность сильно нарушена, фрагменты исходной травяной растительности сохраняются только возле пней. Доминировали на ранней стадии возобновления иван-чай (Chamaenerion angustifolium L.), ситник развесистый (Juncus effusus L.), вейник наземный (Calamagrostis arundinacea L.). В 1980 г. проведена посадка еловых сеянцев в отвалы плужных борозд. На уровне 10-летнего возраста дважды проведена очистка рядов и междурядий от лиственных пород. По мере увеличения сомкнутости происходило замещение пионерных видов нижних ярусов на типичный для лесных южно-таежных экосистем флористический комплекс. К 30-летнему возрасту сформировался разреженный травяной покров с преобладанием кислицы (Oxalis acetosella L.) и неморальных видов: зеленчук (Galeobdolon luteum Huds.), медуница (Pulmonaria obscura Dumort), подмаренник (Galium odoratum L.), ожика волосистая (Lusula pilosa L.). Хорошо развит ярус зеленых мхов с доминированием Pleurozium schreberi Mitt., Hylocomium splendens Hedw., Cirriphyllum piliferum Hedw. и видов рода Plagiomnium.
Для сравнительного анализа дополнительно использовались характеристики ранговой структуры населения жужелиц в ненарушенных неморальных ельниках заповедника и на участках сплошного ветровала 1996 г.
Жужелицы учитывались стандартным методом почвенных ловушек. Использовалась линия из 15 постоянных ловушек с расстоянием между ними 10 м и добавлением в качестве фиксатора 2%-го формалина. Учеты проводились с периодичностью 10 дней, ежегодно, с мая по октябрь.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Многолетний ряд ежегодных наблюдений показывает последовательные смены состояний населения жужелиц по мере развития лесных культур (рис. 1). Восстановительная динамика видовых популяций оценивалась по отношению к структуре населения жужелиц в неморальных южно-таежных ельниках климаксового уровня. Анализ начинается со времени завершения травяной стадии возобновления растительного покрова на вырубке, где в доминантную группу входили лугово-полевые виды жужелиц (Agonum sexpunctatum L., Poecilus versicolor Sturm, Pterostichus vernalis Pz.). С пятого года после рубки быстро формируется мелколиственный растительный покров и одновременно развивается характерный для ранних стадий посткатастрофических сукцессий комплекс гигрофильных видов жужелиц (Agonum fuliginosum Pz., Oxypselaphus obscurus Hbst., Epaphius rivularis Gyll., Pterostichus minor Gyll., Pterostichus diligens Sturm, Dicheirotrichus placidus Gyll.). Отчасти он поддерживается рубками ухода через 10 лет после посадки ели, но на уровне 20-летних культур эти виды становятся малочисленны или регистрируются единично в дальнейших наблюдениях. Наряду с ними уже на ранней стадии возобновления отмечаются типичные виды лесных экосистем южной тайги Epaphius secalis Pk., Leistus terminatus Hellw., Pterostichus niger Schall., которые в дальнейшем сохраняют позиции на всем протяжении восстановительного цикла и входят в состав доминантов зрелых еловых лесов заповедника.
Рис. 1.
Многолетняя динамика видовых популяций жужелиц в еловых культурах. По оси ординат – учетная численность, средняя за период с мая по октябрь. а – Epaphius rivularis, б – Pterostichus minor, в – Oxypselaphus obscurus, г – Pterostichus niger, д – Epaphius secalis, е – Leistus terminatus, ж – Pterostichus melanarius, з – Pterostichus oblongopunctatus, и – Calathus micropterus, к – Carabus hortensis, л – Cychrus caraboides, м – Loricera pilicornis.
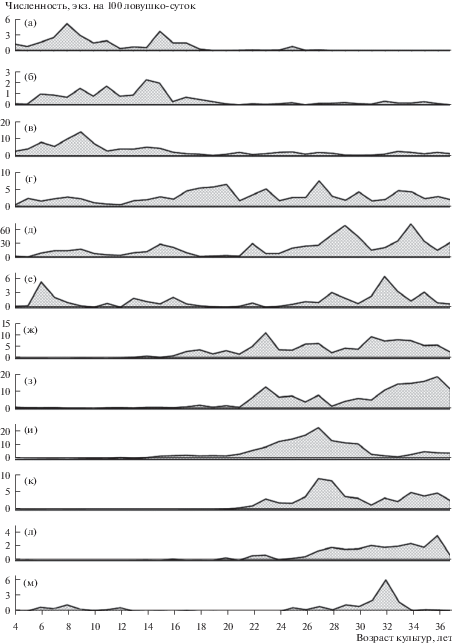
По данным рис. 1 на этом же 20-летнем уровне возраста культур начинается активное восстановление основной части фоновых для ненарушенных южно-таежных ельников видов жужелиц (Carabus hortensis L., Cychrus caraboides L., Pterostichus oblongopunctatus F., Pterostichus melanarius Ill., Calathus micropterus Duft.). Форма динамических кривых в зоне восстановления видоспецифична; наблюдаются относительные смещения во времени и вариации в интенсивности и последовательности нарастания численности популяций. Основной фактор развития этой группы видов – очевидно преобразование среды еловыми культурами. Дополнительные факторы открывает анализ многолетних учетных материалов. Сопоставление данных показывает, что даже для контрастных по типологии и сукцессионному положению экосистем периоды роста и спада видовых популяций жужелиц в значительной степени синхронизированы и должны быть отнесены к территории в целом.
Примеры такой синхронизации показаны на рис. 2, где для сравнения приводятся многолетние материалы по постоянной пробной площади ельника неморально-кисличного. Чтобы отчетливей выделить хронологическую составляющую, годовые показатели обилия видов представлены в долях от суммарной уловистости за все годы наблюдений.
Рис. 2.
Примеры синхронизации многолетней динамики видов жужелиц. 1 – еловые культуры, 2 – ельник неморально-кисличный; а – Calathus micropterus, б – Pterostichus oblongopunctatus, в – Carabus glabratus, г – Carabus coriaceus.
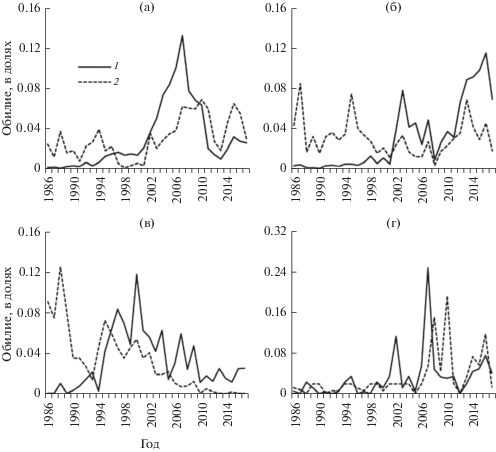
В типичном случае по мере восстановления популяций их динамика синхронизируется с общетерриториальной, характерной для данного вида (рис. 2а, 2б). Сведения по двум видам рода Carabus вынесены за рамки рис. 1 и показывают, что синхронизирующим динамику фактором могут быть многолетние климатические изменения (рис. 2в, 2г). Эти виды контрастны по зональным оптимумам: Carabus glabratus Pk. отчетливо тяготеет к лесам бореальной структуры и доходит в распространении до северной тайги, в отличие от Carabus coriaceus L. – западноевропейского вида, характерного для хвойно-широколиственных и широколиственных лесов (Орлов, 1983; Грюнталь, 2008). Восстановление популяции C. glabratus на площади культур прерывается в конце 1990-х гг. Одновременно его численность последовательно снижается на всех постоянных пробных площадях независимо от лесорастительных условий и сукцессионного положения. Ранее по данным многолетних учетов вид всегда отмечался в доминирующей группе для неморальных и зеленомошных ельников заповедника. Причиной незавершенности восстановительной динамики C. glabratus в культурах может быть рост весенне-летних температур с начала 2000-х гг. (данные метеостанции заповедника). Это подтверждается динамикой C. coriaceus, который с конца 1990-х гг. становится обычным в производных лесах на старых вырубках, а затем и в зрелых ельниках заповедника.
Практически для всех видов жужелиц можно показать, что фаза активного восстановления совпадает с волнами общего для территории роста численности. Вероятно, в эти же периоды увеличиваются миграционные потоки из прилегающих к вырубке лесных экосистем. Стимулирующим или сдерживающим фактором в процессах восстановления, по-видимому, являются климатические изменения.
Непрерывный многолетний ряд данных позволяет оценить формы восстановительного роста популяций жужелиц на площади культур. Почти для всех видов наблюдается фаза медленного нарастания численности, после которой происходит быстрый рост, приближающийся к экспоненциальной зависимости. В отдельных случаях (P. niger, C. micropterus) фаза нарастания выглядит как линейные тренды. Для некоторых видов (L. terminatus, Loricera pilicornis F.) наблюдаются два периода роста популяций – в ранних лиственных молодняках и в возрасте смыкания еловых культур. Как общее явление сразу после фазы роста следует спад, и колебания численности устанавливаются на различных для видов уровнях.
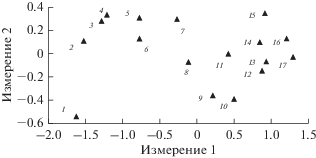
На этом этапе восстановления начинает формироваться видовая и ранговая структура населения жужелиц, характерная для терминальных стадий сукцессии в зрелых ельниках. Двухмерная конфигурация неметрического многомерного шкалирования (НМШ) показывает непрерывные и направленные изменения порядков доминирования видов от начального периода роста культур до сомкнутого насаждения (рис. 3). Признаки стабилизации ранговой структуры населения жужелиц заметны на стадии мелколиственной поросли. После рубок ухода в 10-летних посадках идут интенсивные изменения, связанные с распадом комплекса промежуточных гигрофильных видов и началом восстановления лесной мезофильной группировки. По данным конфигурации НМШ с 25-летнего возраста культур начинаются автономные преобразования внутри комплекса восстанавливающихся видов жужелиц. При этом наблюдается снижение интенсивности структурных изменений в населении жужелиц по сравнению с более ранними возрастами еловых посадок.
Рис. 3.
Ординация изменений ранговой структуры населения жужелиц в еловых культурах методом неметрического многомерного шкалирования. Использованы усредненные значения обилия видов по двухлетиям: 1 – 1984–1985 гг.; 2 – 1986–1987 гг.; 3 – 1988–1989 гг.; … 17 – 2016–2017 гг.
Стабилизацию структурных изменений подтверждает расчет функции автокорреляции с лагом от 1 до 6, отдельно для первого и последнего десятилетия развития еловых посадок (рис. 4). Для всех значений лага устойчивость во времени порядка доминирования видов выше в пределах последнего десятилетия. С увеличением временного промежутка между годами наблюдений структурные различия заметно быстрее увеличиваются для данных первого десятилетия. Отметим, что динамика значений автокорреляции и конфигурация НМШ показывают продолжение направленных изменений структуры населения жужелиц на уровне 30-летнего возраста культур, но по форме эти изменения уже ближе к циклическим.
Рис. 4.
Автокорреляция ранговой структуры населения жужелиц еловых культур в различные периоды временного ряда. 1 – 1984–1994 гг., 2 – 2007–2017 гг.
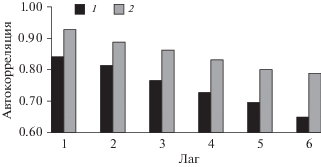
Одновременно со стабилизацией ранговой структуры возрастает устойчивость во времени состава доминантных видов жужелиц. Формируется типичная, в том числе и для зрелых лесных экосистем южной тайги, группа преобладающих по обилию видов (C. hortensis, C. caraboides, P. oblongopunctatus, P. niger, P. melanarius, C. micropterus). Межгодовая общность состава доминантов в последнее десятилетие становится почти независимой от временного лага и составляет в среднем 77% (от 68 до 100% по индексу Жаккара). Сохраняет динамичность состава субдоминантная группа, которая включает экологически разнородные элементы, в том числе виды, адаптированные к нарушениям лесных экосистем. Средняя общность состава этой группы по индексу Жаккара 42% (от 17 до 68%). По данным рис. 4 стабилизация доминантной группы в целом повышает устойчивость ранговой структуры населения жужелиц. Порядок доминирования видов уже меньше зависит от локальных сукцессионных замещений и больше от общих для территории колебаний внешних факторов, таких как погодные условия. Межгодовые изменения численности, связанные с резкими погодными аномалиями, как правило, синхронизированы для массовых видов жужелиц, что поддерживает стабильность ранговой структуры.
Возникает вопрос об оценке восстановительных процессов по отношению к ненарушенным экосистемам ельников заповедника. На уровне 30–35 лет возраста культур общность ранговой структуры населения жужелиц с населением зрелых ельников составляет по коэффициенту Спирмена от 0.75 до 0.83. Рассматривая эти значения как ориентир, у нас есть возможность сравнить данные по культурам с многолетними наблюдениями за естественным возобновлением леса на сплошном ветровале 1996 г. в ельнике неморально-кисличном. На уровне 20 лет после начала сукцессии общность порядков доминирования в населении жужелиц по отношению к населению ненарушенных ельников составляет на ветровале 0.52 и 0.58 в культурах. Однако условия предшествующего развития возобновления резко различаются для этих динамических рядов. Механизированные летние лесозаготовки связаны с обширными повреждениями значимых для биоты верхних почвенных горизонтов. Уничтожение подстилки, минерализация и уплотнение поверхности достигают 90% от площади вырубок (Исаев, 1979). Сходные явления возникают при посадке ели в отвалы плужных борозд. Естественные процессы восстановления прерываются рубками ухода за культурами.
В отличие от вырубок на сплошных ветровалах в значительной мере остаются участки ненарушенной почвы; условия освещенности и гидротермического режима не имеют таких радикальных изменений, как на лесосеках (Скворцова и др., 1983; Карпачевский и др., 1999). В результате намного лучше сохраняется исходное население жужелиц. Это подтверждают сравнительные расчеты коэффициентов Спирмена на разных возрастных уровнях после начала сукцессии. На уровнях от 5 до 15 лет после начала возобновления общность ранговой структуры населения жужелиц с населением зрелых, ненарушенных ельников составляет для культур от –0.06 до 0.09; для ветровала – от 0.77 до 0.58. Характерно последовательное снижение этой корреляции по мере развития на ветровале комплекса видов – индикаторов нарушенности экосистем ельников южной тайги. Сближение к 20 годам значений общности ранговой структуры жужелиц с населением ненарушенных ельников на вырубке и ветровале показывает, что активность восстановительных процессов на ранних этапах возобновления значительно выше в еловых посадках.
ЗАКЛЮЧЕНИЕ
В заключение отметим основные моменты анализа восстановительной динамики населения жужелиц в еловых культурах.
При существующей технологии ухода за посадками на вырубках восстановление популяций фоновых для южно-таежных лесов видов жужелиц происходит на уровне 20-летнего возраста культур. Развитие популяций этих видов на площади культур, как правило, совпадает с общими для территории волнами подъема их численности. Вероятно, в эти периоды также увеличиваются миграционные потоки жужелиц из прилегающих лесов при условии технологии лесозаготовок, которая сохраняет такую возможность.
Восстановительный период в большинстве случаев включает фазу медленного нарастания, затем быстрый рост, приближающийся к экспоненциальной зависимости. Далее, после фазы иногда значительного спада устанавливаются типичные для лесных южно-таежных экосистем уровни относительного обилия видов жужелиц, и начинает формироваться порядок их доминирования в населении. На уровне 30-летнего возраста культур заметно снижается скорость сукцессионных изменений ранговой структуры населения жужелиц и стабилизируется состав доминантной группы видов.
По данным анализа ранговой структуры наблюдается быстрое ее приближение к эталонным характеристикам ненарушенных сложных ельников между 15 и 25 годами возраста культур. Сравнение с естественным возобновлением леса на сплошном ветровале показывает, что на ветровале намного лучше сохраняется исходное население жужелиц ненарушенного ельника. Однако к 20-летнему возрасту скорость восстановления в еловых культурах компенсирует это различие.
Список литературы
Арнольди К.В., Матвеев В.А. Население жужелиц (Carabidae) еловых лесов у южного предела тайги (Марийская АССР) и изменение его на вырубках // Экология почвенных беспозвоночных. М.: Наука, 1973. С. 131–143.
Грюнталь С.Ю. Организация сообществ жужелиц (Coleoptera, Carabidae) лесов Восточно-Европейской (Русской) равнины. М.: Галлея-принт, 2008. 484 с.
Исаев В. И. Изменение экологической среды в результате сплошных рубок леса // Проблемы экологического мониторинга и моделирования экосистем. Л., 1979. Т. 2. С. 249–257.
Карпачевский Л.О., Кураева Е.Н., Минаева Т.Ю., Шапошников Е.С. Демутационные процессы в нарушенных сплошными ветровалами еловых лесах // Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия. СПб.: РБО, 1999. С. 380–387.
Орлов В.А. Жужелицы рода Carabus L. в Московской области // Фауна и экология почвенных беспозвоночных Московской области. М.: Наука, 1983. С. 113–120.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
Побединский А.В. Рубки и восстановление в таежных лесах СССР. М.: Лесная пром-ность, 1973. 199 с.
Скворцова Е.Б., Уланова Н.Г., Басевич В.Ф. Экологическая роль ветровалов. М.: Лесная пром-ность, 1983. 187 с.
Старостина К.Ф. Роль синузий нижних ярусов в регуляции возобновительного процесса ели // Структура и продуктивность еловых лесов южной тайги. Л.: Наука, 1973. С. 246–255.
Huhta V. Evaluation of different similarity indices as measures of succession in arthropod communities of the forest floor after clear-cutting // Oecologia. 1979. V. 41. № 1. P. 11–23.
Koivula M. Boreal carabid beetle (Coleoptera, Carabidae) assemblages in thinned uneven-aged and clear-cut spruce stands // Annales Zoologici Fennici. 2002. V. 39. P. 131–149.
Sklodowski J.W. Anthropogenic transformation of ground beetle assemblages (Coleoptera: Carabidae) in Bialowiesa Forest, Poland: from primeval forest to managed woodlands of varios ages // Entomologica Fennica. 2006. V. 17. P. 296–314.
Дополнительные материалы отсутствуют.