

Лесоведение, 2021, № 3, стр. 227-239
Информативность спектральных и морфометрических признаков оконной структуры полога древостоя на основе спутниковых данных
А. В. Комаров a, *, Д. В. Ершов a, Е. В. Тихонова a
a Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32 стр. 14, Россия
* E-mail: vins90nuar@gmail.com
Поступила в редакцию 24.05.2020
После доработки 04.09.2020
Принята к публикации 03.02.2021
Аннотация
Исследованы различия морфометрических признаков оконной структуры полога древостоя в трех доминирующих типах лесных сообществ Валуевского лесопарка на основе данных дистанционного зондирования высокого и детального пространственного разрешения. Классификация групп лесных сообществ (лиственный лес с преобладанием липы, лиственный лес с преобладанием березы или осины и хвойный лес с преобладанием ели или сосны) методом “случайный лес” выполнена по разносезонным мультиспектральным спутниковым изображениям Sentinel-2/MSI с точностью 0.96 (k = 0.88). По этим же изображениям создан слой сегментов – спектрально-однородных участков леса. Маска окон древесного полога создавалась по панхроматическому изображению Ресурс-П1/Геотон методом кластерного анализа с визуальным дешифрированием кластеров. Рассчитывались восемь морфометрических признаков для каждого окна. Анализ различий проводился на уровне сегментов (U-тест Манна–Уитни) и всей совокупности окон (H-тест Краскела–Уоллиса) каждой группы лесных сообществ. Наибольшие показатели U-теста для средних значений морфометрических признаков на уровне сегментов лесных сообществ имеют площадь окна S (U = 24), его периметр P (U = 19.3), комплексный индекс формы окна GSCI (U = 19.0) и отношение периметра к площади окна P : S (U = 18.7). Наибольшие значения H-теста на уровне отдельных окон лесных сообществ были получены для фрактальной размерности окна PFD (H = 2229.2), отношения периметра к площади окна P : S (H = 2064.9) и площади окна S (H = 1718.4). Анализ результатов и литературы позволил определить возможные причины различий в оконной структуре и параметрах окон хвойных, мелколиственных и липовых сообществ модельной территории.
Лесные экосистемы, развивающиеся в спонтанном режиме при отсутствии катастрофических внешних воздействий, характеризуются мозаичной структурой, представленной “окнами возобновления” на разных стадиях развития (Schliemann, Bockheim, 2011; Muscolo et al., 2014). “Окна” – просветы в пологе взрослого древостоя (treefall gaps) – возникают в результате смерти одного или нескольких крупных деревьев. Оконная мозаика взрослого древостоя определяет пространственную гетерогенность лесного фитоценоза, которую связывают со многими важными экологическими структурами и процессами на разных пространственных масштабах. Окна играют ключевую роль в лесовозобновлении, поддержании таксономического и структурного биоразнообразия лесных сообществ (Широков, 2005). Появление окон отмечается уже в 40–60-летних древостоях, когда начинают отмирать деревья раннесукцессионных видов (Смирнова и др., 2001). Образовавшись, окна не остаются статичными, они существуют в различных стадиях “заполнения”. При рассмотрении всего цикла развития окон, от момента их возникновения до распада нового поколения деревьев, уместно использовать термин “возрастная парцелла”, предложенный Н.В. Дылисом (1978). В хвойно-широколиственных лесах европейской части России в первые 15 лет в окнах доминируют крупные травы и виды кустарников, которые после смыкания полога подроста замещаются на лесные неморальные и бореальные виды растений. Через 35–40 лет с момента образования окна восстанавливается исходная структура нижних ярусов (Широков, 2005).
В работе мы рассматриваем окна первых этапов развития – от момента формирования до закрытия (gap closure). L.E. Tyrrell, T.R. Crow (1994) приводят три критерия закрытия окон: деревья в окне имеют высоту от половины до двух третей окружающего полога; деревья в окне имеют диаметры (на уровне груди) больше 25 см; древесный полог настолько густой, что исходное окно трудно выявить.
Еще один важный параметр, который необходимо учитывать при исследовании оконной структуры верхнего полога древостоя — это верхний и нижний пределы размера окон. По определению, под окнами понимаются небольшие прорывы в пологе, образующиеся при отмирании одного или нескольких деревьев. J. McCarthy (2001) считает необходимым различать понятия “окно” (gap) и “участок” (patch). Это необходимо, потому что участки, сформированные крупномасштабными катастрофическими нарушениями (пожары, вспышки насекомых, массовые ветровалы), сильно отличаются от небольших окон по микроклимату, времени возобновления, освещенности, доступности для светолюбивой флоры и другим параметрам. Нижний предел размера окон определяется площадью от вывала единичного дерева, которая обычно варьирует от 50 до 200 м2 (Drobyshev, 1999; McCarthy, 2001).
Для пространственного анализа оконной структуры традиционно используются различные наземные методы: 1) трансекты (Широков, 2005); 2) дендроэкологический анализ (Hobi et al., 2015); 3) фотосъемка с использованием полусферической камеры (Nyamgeroh et al., 2018). Однако наземные методы требуют значительных затрат времени и труда на проведение измерений и подготовку схем пространственного расположения окон в пологе древостоя (Hobi et al., 2015; Nyamgeroh et al., 2018), поэтому их применение ограничено небольшими площадями (до нескольких гектар).
Возможности применения данных аэрокосмической съемки для картографирования и изучения оконной структуры лесов исследованы в ряде работ (Runkle, 1992; Fox et al., 2000; Hobi et al., 2015; Barton et al., 2017; Bagaram et al., 2018; Nyamgeroh et al., 2018; White et al., 2018). J.R. Runkle (1992) исследовал возможности данных аэрофотосъемки для картографирования окон древесного полога. В более поздней работе T.J. Fox (2000) провел сравнение точности карт окон, созданных на основе наземной съемки полога леса, с результатами визуального дешифрирования аэрофотоснимков высокого пространственного разрешения (1 : 15 000). Было показано, что созданные на основе аэрофотоснимков карты оказались более точными, так как они имели намного меньший процент пропусков (4.7%) картографирования окон по сравнению с наземными обследованиями (25.6%). Однако, хотя аэрофотосъемка позволяет с более высокой точностью выявлять окна в пологе древостоя, с ее помощью невозможно детально описать характеристики растительности в окнах, что обеспечивают наземные методы.
Отдельный вопрос − влияние собственных теней деревьев на точность определения границ и площадей окон верхнего полога. Из-за различий высоты деревьев часть крон оказывается затененной и включается в окна при картографировании (Zielewska-Büttner et al., 2016; Barton et al., 2017; Nyamgeroh et al., 2018). Исследование B.B. Nyamgeroh с соавт. (2018) показало, что при использовании методов выделения окон по изображениям (объектно-ориентированный анализ изображений OBIA и текстурный анализ изображений ITBA) происходит переоценка их площади (особенно для окон малых размеров) по сравнению с наземными измерениями. Хуже всего выделяются окна в пологе широколиственных пород.
Преимущество спутниковых измерений заключается в том, что они позволяют исследовать оконную структуру лесов в труднодоступных регионах, а также за счет повторной съемки одних и тех же территорий обеспечивают возможность анализа динамики площади окон и, соответственно, изменения породно-возрастной структуры лесов (Hobi et al., 2015; Nyamgeroh et al., 2018). Оконная структура древесного полога различается в лесных сообществах разного типа и сукцессионного статуса. Это может использоваться как косвенный дешифровочный признак при их картографировании. M.L. Hobi с соавт. (2015) в своей работе использовали стереопары спутниковых изображений WorldView-2 для изучения параметров окон древесного полога старовозрастных буковых лесов украинских Карпат. С помощью спутниковых изображений было выявлено, что размер окон в пологе букового леса не превышает 200 м2, а доля нарушений, вызванных ураганными ветрами и снеголомами, меньше 1% площади исследуемой территории. Проверка спутниковых карт окон древостоя на основе наземных данных оказалась сложной задачей, так как не все пробные площади можно было точно пространственно привязать в ходе наземного обследования. Но в целом оценки размеров окон, полученные наземными и дистанционными методами, хорошо согласуются. Таким образом, литературный анализ показывает сложность реализации задачи, а с другой стороны − перспективу разработки и применения дистанционных методов для картографирования оконной структуры лесов и их мониторинга на больших лесных территориях.
Цель настоящего исследования – оценка различий параметров оконной структуры разных групп лесных сообществ на основе спутниковых изображений детального разрешения. Для этого решались следующие задачи: (1) оценить информативность спектральных каналов и провести классификацию основных групп лесных сообществ по спутниковым данным Sentinel-2b/MSI (10–20 м); (2) сформировать маску окон полога древостоя с использованием панхроматического изображения Ресурс-П1/Геотон (0.8 м); (3) рассчитать морфометрические признаки оконной структуры древостоя и статистическими методами оценить различия значений морфометрических признаков для групп лесных сообществ.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводились на модельной территории Валуевского лесопарка (Новомосковский административный округ г. Москва) (рис. 1). Общая площадь исследуемой территории составляет 1156.33 га (Публичная …, 2020). Валуевский лесопарк лежит в пределах Апрелевско-Кунцевского ландшафта с доминированием моренных равнин с абсолютными высотами 190–210 м. В почвенном покрове преобладают дерново-подзолистые почвы на покровном суглинке. Почвообразующая порода характеризуется средне-тяжелосуглинистым гранулометрическим составом (Аккумуляция …, 2018). По материалам таксации последнего лесоустройства леса представлены широколиственными (с доминированием липы) и широколиственно-еловыми сообществами возрастом от 60 до 110 лет (Материалы лесоустройства …, 2005). Значительное распространение имеют вторичные мелколиственные леса, находящиеся на разных стадиях послерубочной сукцессии. Лесные культуры ели и сосны занимают незначительные площади. В статье рассматриваются три доминирующие группы лесных сообществ: (1) лиственный лес с преобладанием липы; (2) лиственный лес с преобладанием березы или осины; (3) хвойный лес с преобладанием ели или сосны.
Рис. 1.
План лесонасаждений Валуевского лесопарка по преобладающим породам
1 – места расположения наземных площадок 2016–2019 гг.; лесные формации: 2 – Picea abies; 3 – Betula pendula и Populus tremula; 4 – Tilia cordata; 5 – Pinus sylvestris; 6 – другие породы.
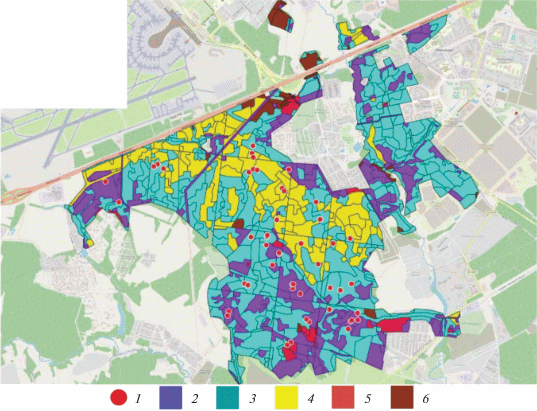
Выбор групп лесных сообществ проводился на основе анализа материалов последнего лесоустройства и данных выборочных наземных геоботанических и таксационных обследований.
Для оценки пространственного распределения лесных сообществ Валуевского лесопарка использовали разносезонные мультиспектральные изображения спутников Sentinel-2, запущенных в 2015 и 2017 г. в рамках научной космической программы Европейской комиссии Copernicus. Мультиспектральный прибор MSI каждого спутника позволяет вести съемку земной поверхности с шириной захвата 290 км, с периодом повторной съемки 5 суток в 13 каналах видимого, ближнего и среднего инфракрасных (ИК) диапазонов различного пространственного разрешения (10, 20 и 60 м) (Sentinel-2 …, 2020).
Для исследований собран временной ряд безоблачных изображений Sentinel-2 с уровнем обработки Level-2A (Level-2A Algorithm Overview, 2020) за 2017, 2018 и 2019 г. В таблице 1 приводятся параметры пяти каналов и характеристики диапазонов значений (минимум, максимум, среднее значение) спектральных яркостей растительности лесопарка в указанные периоды съемки.
Таблица 1.
Характеристика набора данных Sentinel-2
| Спектральный канал* | Статисти-ческий параметр | Спектральный коэффициент яркости лесов модельной территории для разных дат съемки** | ||||||
|---|---|---|---|---|---|---|---|---|
| янв. 2017 г. | апр. 2017 г. | май 2018 г. | авг. 2017 г. | сент. 2017 г. | дек. 2017 г. | окт. 2018 г. | ||
| B3, зеленый (559 нм, 10 м) |
Мин. | 917 | 576 | 490 | 560 | 632 | 978 | 619 |
| Макс. | 8231 | 1732 | 2596 | 1764 | 1186 | 6452 | 1661 | |
| Среднее | 2128 | 754 | 656 | 684 | 765 | 2099 | 814 | |
| B4, красный (665 нм, 10 м) |
Мин. | 917 | 402 | 490 | 343 | 399 | 890 | 390 |
| Макс. | 8002 | 2065 | 2596 | 2150 | 1203 | 6808 | 1506 | |
| Среднее | 1417 | 632 | 656 | 432 | 506 | 2105 | 726 | |
| B8, БИК (833 нм, 10 м) |
Мин. | 893 | 399 | 344 | 482 | 643 | 1128 | 355 |
| Макс. | 10512 | 1203 | 5586 | 4066 | 3490 | 6836 | 14 332 | |
| Среднее | 2480 | 506 | 2744 | 2245 | 1944 | 2522 | 2998 | |
| B11, срИК 1 (1610.4 нм, 20 м) |
Мин. | 62 | 343 | 228 | 251 | 484 | 302 | 389 |
| Макс. | 1439 | 2150 | 3692 | 3166 | 1984 | 2884 | 2702 | |
| Среднее | 687 | 432 | 1201 | 1065 | 1050 | 807 | 1310 | |
| B12, срИК 2 (2185.7 нм, 20 м) |
Мин. | 42 | 0 | 138 | 2740 | 220 | 2501 | 86 |
| Макс. | 1310 | 2168 | 3360 | 2260 | 1038 | 2501 | 2183 | |
| Среднее | 428 | 210 | 138 | 112 | 456 | 769 | 507 | |
Для спектральных каналов с пространственным разрешением 20 м было проведено геометрическое преобразование стандартным алгоритмом обработки изображений “resampling” с использованием функции бикубической интерполяции (ERDAS Imagine …, 2014) для получения единого пространственного разрешения всех спектральных каналов.
Для пространственной идентификации окон в древесном пологе использовалось панхроматическое изображение спутника Ресурс-П1, с пространственным разрешением около 1 м. Ресурс-П1 – спутник с оптическими приборами на борту, предназначенный для мониторинга природных и антропогенных объектов Земли (Космические …, 2020).
Помимо спутниковой информации подготовлены цифровые данные таксации лесов по материалам лесоустройства 2005 г. В период с 2016 по 2019 г. проводились наземные обследования территории, геоботанические описания были выполнены на 53 площадках размером 20 × 20 м, подеревная таксация насаждений – на 15 площадках размером 50 × 50 м (рис. 1).
Схема последовательности этапов исследований представлена на рис. 2. Набор эталонов для классификации групп лесных сообществ формировался на основе материалов повыдельной таксации лесов. Кандидатом в эталоны являлся выдел с участием семи единиц и более древесных пород в составе насаждения, входящих в группу лесных сообществ.
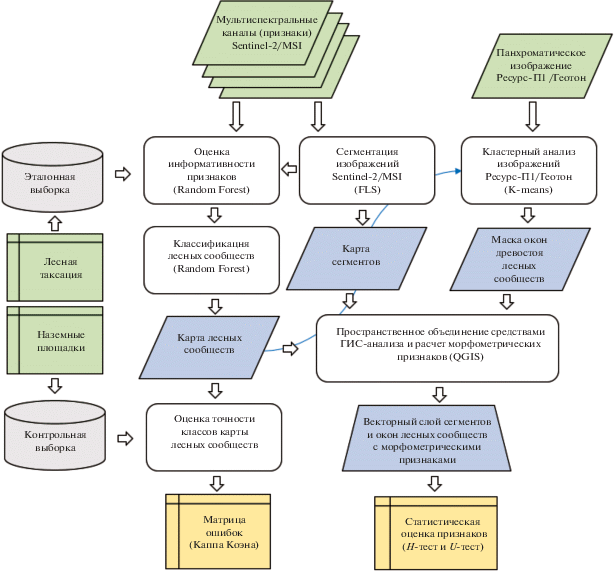
В случаях, когда доля породы в составе насаждения была меньше определенного в исследовании порога (семь единиц), породы одной группы (сосна и ель, береза и осина) суммировались и повторно сравнивались с пороговым значением. Контроль соответствия выделов их таксационным характеристикам осуществлялся методом визуального анализа синтезированных спутниковых изображений и данных наземных площадок. Для получения спектрально-однородных эталонов внутри выделов выполнялась сегментация разносезонных мультиспектральных изображений Sentinel-2 методом FLS (Full Lambda Schedule) (Redding et al., 1999) в программе обработки спутниковых изображений ERDAS Imagine. Сегменты использовались в дальнейшем для статистического анализа морфометрических признаков в их границах. С одной стороны это позволило минимизировать влияние ошибок классификации групп лесных сообществ по изображениям Sentinel-2, а с другой стороны − оценить уровень различимости усредненных на сегмент значений морфометрических признаков по отношению к их исходным значениям на уровне отдельного окна.
Эталонная выборка использовалась для оценки информативности мультиспектральных каналов (признаков) и настройки модели классификации с обучением “случайный лес” (Breiman, 2001). Выбор алгоритма “случайный лес” обусловлен его более высокой точностью классификации лесов по сравнению с другими методами тематической обработки изображений (Maxwell et al., 2018).
Проверку результатов классификации групп лесных сообществ выполняли на основе данных наземных обследований. Каждая площадка была отнесена к одной из групп сообществ. Затем пространственно площадки совмещали с тематической картой лесных сообществ, из которой извлекались пикселы с классами, попавшие на площадки. Оценку точности тематической карты лесных сообществ выполняли на основе анализа матрицы ошибок с оценкой показателя каппы Коэна (k) (Крамер, 1975).
В рамках работы мы принимаем минимальный размер окна, равный 20 м2, что согласуется со средним размером крон ели и липы в условиях Подмосковья – 15–27 м2 (Абатуров, Меланхолин, 2004). По причине отсутствия на территории исследования участков крупных нарушений (более 1 га, McCarthy, 2001) верхний предел размера окон нами не устанавливался.
На следующем этапе исследований выполнялся кластерный анализ спутниковых изображений детального пространственного разрешения Ресурс-П1/Геотон для выделения оконной структуры древесного полога модельной территории. Кластеры окон включают пикселы с низкими значениями яркостей, которым соответствуют не только окна древостоя, но и затененные участки крон деревьев верхнего и подчиненных ярусов. Так как применение пороговых методов невозможно из-за высокой вариабельности значений яркости окон для лесных сообществ, мы использовали неуправляемую избыточную классификацию на основе метода K-means (Mirkes, 2011), с последующим визуальным дешифрированием классов для каждой группы лесных сообществ отдельно и их объединением в два класса “окно” и “лес”.
Далее средствами ГИС растровые слои сегментов и маски окон древесного полога преобразовывали в векторный формат и пространственно пересекались, используя функции расширения PostGIS − st_intersects (https://postgis.net/docs/ ST_Intersects.html), st_within (https://postgis.net/ docs/ST_Within.html). Для векторного слоя окон были рассчитаны следующие морфометрические характеристики: площадь (S), периметр окна (P), их отношение (P : S), округлость форм (1) и ряд фрактальных признаков (2)–(5), а именно (Getzin et al., 2012):
где CIRC – округлость окна, S – площадь окна, P – периметр окна где GSCI – комплексный индекс формы окна, S – площадь окна, P – периметр окна где PFD – фрактальная размерность окна, S – площадь окна, P – периметр окна где FDI – индекс фрактальной размерности окна, S – площадь окна, P – периметр окна где FD – фрактальная размерность, S – площадь окна, P – периметр окнаДля определения различий в значениях морфометрических признаков трех групп лесных сообществ использовались непараметрический H‑тест Краскела−Уоллиса (Kruskal, Wallis, 1952) и его параметрический аналог – U-тест Манна−Уитни (Mann, Whitney, 1947). Анализ морфометрических признаков проводился с использованием двух наборов данных: (1) на уровне отдельных окон, принадлежащих группам лесных сообществ, с применением H-теста; в этом случае каждый полигон в векторном слое рассматривался как отдельное окно, принадлежащее к одной из групп лесных сообществ; (2) на уровне сегментов с применением U-теста; второй тест проводился для выборки из усредненных и взвешенных значений морфометрических признаков в границах сегмента, принадлежащего одной из трех групп лесных сообществ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ информативности спектральных каналов Sentinel-2/MSI показал, что увеличение их числа влияет на результаты классификации лесных сообществ. Однако вклад различен и зависит от фенологического периода съемки лесов и спектрального диапазона (длины волны). Наибольший вклад в распознавание трех групп лесных сообществ из 35 спектральных каналов (признаков), полученных в разные вегетационные сезоны, вносят средние ИК диапазоны B11 (1565–1655 нм) за май, сентябрь и декабрь 2017 г. и B12 (2100–2280 нм) – за август 2017 и май 2018 г. Следующими по значимости являются ближний ИК диапазон B8, полученный при съемке в мае и декабре 2018 г., а зеленый B3 и красный B4 диапазоны − для апреля 2017 г. и октября 2018 г., соответственно. Классификация групп лесных сообществ алгоритмом “случайный лес” выполнялась по трем наборам признаков, а именно 15, 21 и 35 спектральных диапазонов.
Интегральная точность классификации лесных сообществ с 15 признаками составила 0.80 (каппа k = 0.64), 21 признаками – 0.89 (k = 0.84) и для 35 признаков – 0.96 (k = 0.88). Наилучший результат классификации групп лесных сообществ по спутниковым изображениям Sentinel-2 представлен на рис. 3а.
Рис. 3.
Результаты обработки спутниковых изображений: (a) карта групп лесных сообществ с границами сегментов; (б) маска окон лесов с границами выделов. 1 – с преобладанием липы, 2 – мелколиственные, 3 – с преобладанием хвойных.
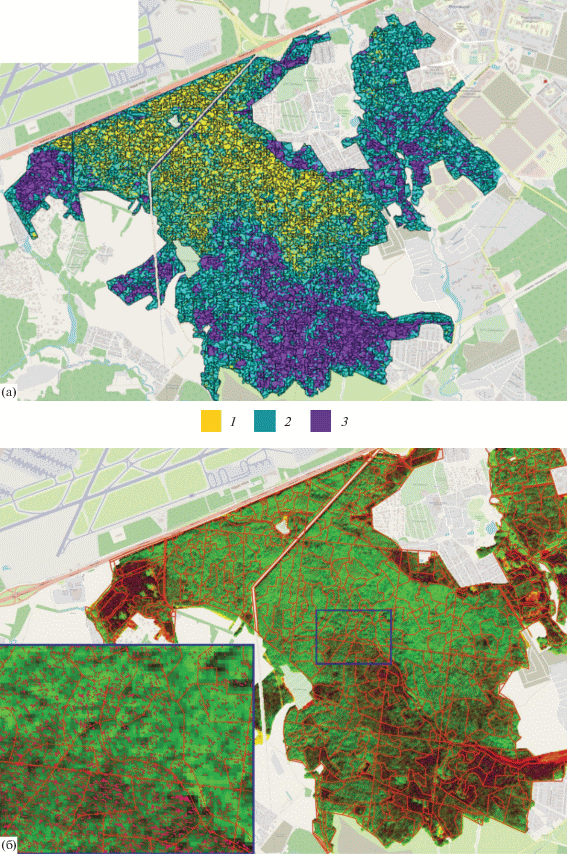
Результат классификации панхроматического изображения Ресурс-П1/Геотон содержал 200 классов, из которых классы с низкими значениями пикселов изображения были визуально объединены в группу классов, характеризующих “окно”. Все остальные классы были сгруппированы в группу “лес”. Полученная бинарная маска окон использовалась для исследования различий морфометрических параметров оконной структуры трех групп лесных сообществ (рис. 3б).
Результаты тестирования различий морфометрических признаков двумя тестами позволили выявить наиболее информативные из них для обоих уровней (табл. 2). Наибольшие значения H-теста на уровне отдельных окон лесных сообществ имеют фрактальная размерность окна (PFD) (H = 2229.2), отношение периметра к площади окна (P : S) (H = 2064.9) и площадь окна (S) (H = 1718.4).
Таблица 2.
Различия морфометрических признаков оконной структуры лесных сообществ
| Тест | CIRC | S | P | GSCI | PFD | FDI | FD | P : S |
|---|---|---|---|---|---|---|---|---|
| На уровне отдельных окон лесных сообществ | ||||||||
| H-тест | 517.97 | 1718.4 | 996.61 | 517.95 | 2229.2 | 29.079 | 37.219 | 2064.9 |
| p-значение | <2.2e-16 | <2.2e-16 | <2.2e-16 | <2.2e-16 | <2.2e-16 | 4.848e-07 | 8.278e-09 | <2.2e-16 |
| На уровне средних характеристик окон в сегментах лесных сообществ | ||||||||
| U-тест | – | 24.0 | 19.3 | 19.0 | – | – | – | 18.7 |
| p-значение | >0.05 | 0.000116 | 0.00321 | 0.00311 | >0.05 | >0.05 | >0.05 | 0.01139 |
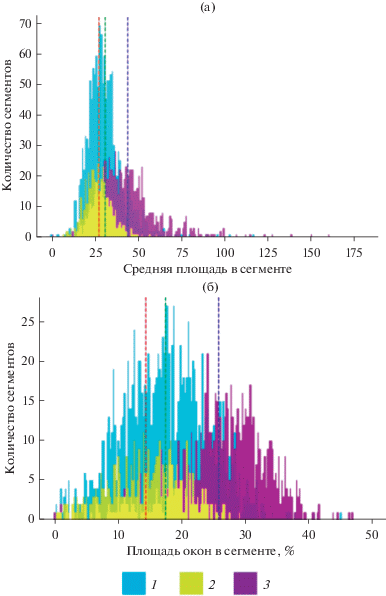
Наибольшие значения U-теста для средних значений морфометрических признаков на уровне сегментов лесных сообществ имеют S (U = = 24.0), периметр окна (P) (U = 19.3), комплексный индекс формы окна (GSCI) (U = 19.0) и P : S (U = 18.7). Таким образом, P : S и S сохраняют свои различия в группах лесных сообществ даже при их осреднении на уровне сегментов. Статистический анализ также показал, что средняя площадь окна в хвойных древостоях и доля площади окон в сегменте превышает аналогичные значения для мелколиственных и широколиственных пород (табл. 3, рис. 4).
Таблица 3.
Средняя площадь и доля площади окон в сегменте для хвойных, мелколиственных и широколиственных (липовых) сообществ
| Группа лесных сообществ | Площадь окна в сегменте, га | Доля площади окон в сегменте, % | ||
|---|---|---|---|---|
| µ | σ | µ | σ | |
| Хвойные | 70.3 | 75.4 | 25.9 | 7.6 |
| Мелколиственные | 48.1 | 49.2 | 17.5 | 6.7 |
| Широколиственные | 42.4 | 32.2 | 14.1 | 5.9 |
Рис. 4.
Распределение числа сегментов хвойных, мелколиственных и широколиственных (липовых) сообществ по среднему значению площади окна (а) и доли площади окон в сегменте (б). 1 – мелколиственные, 2 – липа, 3 – ель.
Анализ литературных источников показывает, что оконная структура полога лесных сообществ определяется одновременно идущими процессами образования и закрытия окон (Kenderes et al., 2009). Параметры оконной структуры специфичны для разных типов лесных сообществ и зависят от многих факторов, которые можно объединить в две группы, соответствующие названным выше динамическим процессам. Первая группа включает факторы, связанные с интенсивностью отпада (смертностью) деревьев: возраст и размер деревьев, ветровальность, различные патогены, положение в ландшафте. Вторая группа объединяет факторы, определяющие скорость закрытия окон, такие как боковой рост, предварительный подрост и др.
В работе S.E. Dahir, C.G. Lorimer (1996) исследовалась оконная структура в широколиственных лесах на разных стадиях развития (молодых, спелых и старовозрастных) на севере США, установлена зависимость размера окон от возраста и размера деревьев. Средняя площадь окна в молодых насаждениях была в 4 раза меньше, а доля окон в общей площади насаждений – в 2 раза меньше, чем в старовозрастных насаждениях. Сходные результаты получены для лесов из дугласовой пихты (псевдотсуги) (Spies, Franklin, 1989). Показано, что на оконную динамику могут оказывать влияние различные патогены (Kneeshaw, Bergeron, 1998; Pham et al., 2004; Messier et al., 2005). Одним из важных факторов, определяющих интенсивность образования окон, является ветровальность деревьев, которая зависит от вида (Kenderes et al., 2009) и возраста деревьев, а также положения в ландшафте (Мирин, 2010).
Скорость закрытия окон, в первую очередь, определяется их размером. Мелкие окна (диаметр которых не превышает высоту окружающих деревьев) закрываются намного быстрее крупных, обычно это происходит в результате бокового разрастания ветвей деревьев (Ulanova, 2000; McCarthy, 2001; Kucbel et al., 2010). Скорость бокового роста широколиственных деревьев оценивается в 4–26 см в год (Runkle, 1985, цит. по: McCarthy, 2001). Однако хвойные деревья с моноподиальным ростом имеют ограниченную способность для заполнения освобождающегося пространства посредством гибкого изменения ветвления (Kuuluvainen, 1994; Pham et al., 2004), поэтому окна в хвойных лесах могут быть более долговечными. На скорость закрытия окон может оказывать влияние хорошо развитый подчиненный (второй) подъярус древостоя, деревья которого способны быстро заполнять образующиеся в пологе открытые участки (Kenderes et al., 2009).
В нашей работе наибольшие значения среднего размера окна и доли площади окон в сегменте получены для группы хвойных сообществ. Этот результат можно объяснить несколькими причинами: хвойные насаждения на исследуемой территории имеют самый большой возраст, деревья ели и сосны достигают наибольшей высоты (более 30 м), еловые сообщества довольно сильно пострадали от вспышки короеда-типографа в 2010–2014 гг., ель, имеющая поверхностную корневую систему, в большей степени подвержена ветровалам, чем лиственные деревья. Возраст мелколиственных насаждений (80–100 лет) приближается к пределу долговечности березы и осины. Многие деревья имеют стволовые гнили, что определяет их ветровальность и ветроломность. Отпад деревьев верхнего яруса происходит достаточно активно, но из-за того, что в мелколиственных сообществах обычно уже сформировался высокий второй подъярус из ели и, в меньшей степени, липы и клена, процесс окнообразования в этих сообществах выражен не так сильно. Для липовых сообществ характерны наименьшие значения среднего размера и доли окон. Это определяется способностью липы к боковому разрастанию кроны, большей долговечностью по сравнению с мелколиственными видами и меньшей ветровальностью вследствие относительно небольшой высоты деревьев (до 25 м) и глубокой корневой системы.
ЗАКЛЮЧЕНИЕ
Проведены исследования по оценке различий параметров оконной структуры трех групп лесных сообществ Валуевского лесопарка. Для этого создана карта лесных сообществ с использованием 35 спектральных признаков разносезонных изображений Sentinel-2/MSI (10–20 м). Наибольшую информативность при классификации групп лесных сообществ показали средние ИК каналы Sentinel-2 за май, август, сентябрь и декабрь 2017 и май 2018 г. Точность карты составила 0.96 (k = 0.88). С помощью кластерного анализа из 200 классов и визуального дешифрирования панхроматического изображения Ресурс-П1/Геотон (0.8 м) была сформирована маска окон полога древостоя и рассчитаны восемь морфометрических признаков на основе литературных данных. На уровне отдельных окон лесных сообществ значения площади, периметра и их отношения, а также фрактальная размерность окна имеют статистически значимые различия для каждой из трех групп. Для средних значений морфометрических признаков на уровне сегментов статистически значимыми признаками являются площадь и периметр окна, их отношение и комплексный индекс формы окна. Выявлены различия в значениях средней площади окна и его доли в сегменте для хвойных и лиственных сообществ. Связано это с различным световым режимом, влияющим на восстановление подроста в сообществах, а также строением крон хвойных и лиственных пород, последние из которых способны занимать освободившееся пространство за счет активного роста боковых побегов.
Список литературы
Абатуров А.В., Меланхолин П.Н. Естественная динамика леса на постоянных пробных площадях в Подмосковье. Тула: Гриф и К°. 2004. 333 с.
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов / Под ред. Лукиной Н.В. М: Товарищество научных изданий КМК, 2018. 232 с.
Дылис Н.В. Основы биогеоценологии. М.: Наука, 1978. 172 с.
Космические аппараты типа “Ресурс-П” (находятся в эксплуатации). URL: http://www.ntsomz.ru/ks_dzz/satellites/resurs_p (дата обращения: 23.04.2020).
Крамер Г. Математические методы статистики. М.: Мир, 1975. 648 с.
Материалы лесоустройства лесхоза Экспериментальный Москворецкий, Таксационное описание (по состоянию на 01.01.05 года).
Мирин Д.М. Причины и следствия высокой ветровальности приручьевых еловых лесов // Ученые записки Российского государственного гидрометеорологического университета. 2010. № 13. С. 111–120.
Публичная кадастровая карта Московской области. Интернет ресурс. URL: https://публичная-кадастровая-карта.рф/московская-область/ (дата обращения 21.01.2020).
Смирнова О.В., Бобровский М.В., Ханина Л.Г. Оценка и прогноз сукцессионных процессов в лесных ценозах на основе демографических методов // Бюллетень МОИП. Отд. биол. 2001. Т. 106. Вып. 5. С. 25–33.
Широков А.И. Использование метода парцеллярного анализа для оценки структурного биоразнообразия лесных сообществ // Лесоведение. 2005. № 1. С. 19–27.
Bagaram M.B., Giuliarelli D., Chirici G., Giannetti F., Barbati A. UAV Remote Sensing for Biodiversity Monitoring: Are Forest Canopy Gaps Good Covariates? // Remote Sensing. 2018. V. 10. P. 1397.
Barton I., Király G., Czimber K., Hollaus M., Pfeifer N. Treefall Gap Mapping Using Sentinel-2 Images // Forests. 2017. V. 8. № 11. P. 426.
Breiman L. Random forests // Machine Learning. 2001. V. 45. № 1. P. 5–32.
Dahir S.E., Lorimer C.G. Variation in canopy gap formation among developmental stages of northern hardwood stands // Canadian J. Forest Research. 1996. V. 26. № 10. P. 1875–1892.
Drobyshev I.V. Regeneration of Norway spruce in canopy gaps in Sphagnum–Myrtillus old-growth forests // Forest Ecology and Management. 1999. V. 115. Iss. 1. P. 71–83.
ERDAS IMAGINE 2014 Product Description. URL: https://ru.scribd.com/document/254309118/ERDAS-IMAGINE-2014-Product-Description-sflb (дата обращения: 23.04.2020).
Fox T.J., Knutson M.G., Hines R.K. Mapping Forest Canopy Gaps Using Air-Photo Interpretation and Ground Surveys // Wildlife Society Bulletin. 2000. V. 28. № 4. P. 882–889.
Getzin S., Wiegand K., Schöning I. Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles // Methods in Ecology and Evolution. 2012. V. 3. Iss. 2. P. 397–404.
Hobi M.L., Ginzler C., Commarmot B., Bugmann H. Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing // Ecosphere. 2015. V. 6. Iss. 5. P. 1–15.
Kenderes K., Král K., Vrška T., Standovár T. Natural Gap Dynamics in a Central European Mixed Beech-Spruce-Fir Old-Growth Forest // Ecoscience. 2009. V. 16. P. 39–47.
Kneeshaw D., Bergeron Y. Canopy gap characteristics and tree replacement in the southeastern boreal forest // Ecology. 1998. V. 79. № 3. P. 783–794.
Kruskal W.H., Wallis W.A. Use of ranks in one-criterion variance analysis // Journal of the American Statistical Association. 1952. V. 47. № 260. P. 583–621.
Kucbel S., Jaloviar P., Saniga M., Vencurik J., Klimaš V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians // European Journal of Forest Research. 2010. V. 129. P. 249–259.
Kuuluvainen T. Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: a review // Annales Zoologici Fennici. 1994. V. 31. № 1. P. 35–51.
Level-2A Algorithm Overview. URL: https://earth.esa.int/ web/sentinel/technical-guides/sentinel-2-msi/level-2a/ algorithm (дата обращения: 23.04.2020).
Mann H.B., Whitney D.R. On a test of whether one of two random variables is stochastically larger than the other // Annals of Mathematical Statistics. 1947. № 18. P. 50–60.
Maxwell A.E., Warner T.A., Fang F. Implementation of machine-learning classification in remote sensing: an applied review // International J. Remote Sensing. 2018. V. 39. Iss. 9. P. 2784–2817.
McCarthy J. Gap dynamics of forest trees: A review with particular attention to boreal forests // Environmental Reviews. 2001. V. 9(1). P. 1–59.
Messier J., Kneeshaw D., Bouchard M., de Römer A. A comparison of gap characteristics in mixedwood old-growth forests in eastern and western Quebec // Canadian J. Forest Research. 2005. V. 35. P. 2510–2514.
Mirkes E.M. K-means and K-methods applet. University of Leicester, 2011. URL: http://www.math.le.ac.uk/people/ ag153/homepage/KmeansKmedoids/Kmeans_Kmedoids. html (дата обращения 28.01.2021 г.).
Muscolo A., Bagnato S., Sidari M., Mercurio R. A review of the roles of forest canopy gaps // Journal of Forestry Research. 2014. V. 25. № 4. P. 725−736.
Nyamgeroh B.B., Groen T.A., Weir M.J.C., Dimov P., Zlatanov T. Detection of forest canopy gaps from very high resolution aerial images // Ecological Indicators. 2018. V. 95. P. 629–636.
Pham A.T., De Grandpré L., Gauthier S., Bergeron Y. Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec // Canadian J. Forest Research. 2004. V. 34. P. 353–364.
Redding N.J., Crisp D.J., Tang D.H., Newsam G.N. An efficient algorithm for Mumford-Shah segmentation and its application to SAR imagery // Proceedings of the 1999 Conference on Digital Image Computing: Techniques and Applications (DICTA-99). 1999. P. 35–41.
Runkle J.R. Guidelines and sample protocol for sampling forest gaps. General technical report PNW-GTR-283. Portland, Or., U.S.: Department of Agriculture, Forest Service, Pacific Northwest Research Station. 1992. 44 p.
Schliemann S.A., Bockheim J.G. Methods for studying treefall gaps: a review // Forest Ecology and Management. 2011. V. 261. P. 1143–1151.
Sentinel-2 MSI instrument: URL https://sentinels.copernicus.eu/web/sentinel/technical-guides/sentinel-2-msi/msi-instrument (дата обращения 29.01.2020 г.).
Spies T.A., Franklin J.F. Gap characteristics and vegetation response in coniferous forests of the Pacific Northwest // Ecology. 1989. V. 70. P. 543–545.
Tyrrell L.E., Crow T.R. Structural characteristics of old-growth hemlock–hardwood forests in relation to age // Ecology. 1994. V. 75. P. 370–386.
Ulanova N.G. The effects of windthrow on forests at different spatial scales: a review // Forest Ecology and Management. 2000. V. 135. P. 155–167.
White J.C., Tompalski P., Coops N.C., Wulder M.A. Comparison of airborne laser scanning and digital stereo imagery for characterizing forest canopy gaps in coastal temperate rainforests // Remote Sensing of Environment. 2018. V. 208. P. 1–14.
Zielewska-Büttner K., Adler P., Ehmann M., Braunisch V. Automated Detection of Forest Gaps in Spruce Dominated Stands Using Canopy Height Models Derived from Stereo Aerial Imagery // Remote sensing. 2016. V. 8. № 3. P. 175.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение