

Лесоведение, 2021, № 3, стр. 278-289
Сопряженная динамика восстановления растительности и сообществ раковинных амеб на гарях в лесах Центральной Эвенкии
О. М. Шабалина a, *, А. В. Гренадерова a, И. Н. Безкоровайная a, А. А. Красильникова a, А. В. Кошкарова a
a Сибирский федеральный университет
660041 Красноярск, просп. Свободный, 79, Россия
* E-mail: oshabalina@sfu-kras.ru
Поступила в редакцию 16.12.2019
После доработки 13.02.2020
Принята к публикации 03.02.2021
Аннотация
В статье рассматриваются основные этапы восстановления растительного покрова и сообществ раковинных амеб на лиственничных гарях разного возраста в условиях северной тайги Центральной Эвенкии. Показано, что после сильных устойчивых низовых пожаров, полностью уничтоживших древостой и напочвенный покров, наблюдается сопряженная динамика растительности и сообществ раковинных амеб, сопровождающаяся сменой видов-эксплерентов видами с преимущественно патиентной и виолентной стратегиями. Развитие растительного покрова и подстилки на гарях, сопровождающееся улучшением гидротермического режима, в ценозах раковинных амеб вызывает расширение спектра морфо-экологических групп и появление более влаголюбивых форм. Естественное возобновление лиственницы на “черных гарях” начинается уже на следующий год после пожара, однако, максимума достигает на второй-третий год. В дальнейшем процесс существенно замедляется, но не прекращается вплоть до седьмого-восьмого года, а в отдельных случаях может длиться до 20 и более лет.
Лесные пожары являются одним из основных периодических повторяющихся экзогенных факторов, изменяющих экологические функции экосистем. Совместно с климатом пирогенный фактор контролирует возрастную структуру и мозаичность растительного покрова, потоки вещества и энергии (Abaimov et al., 2000; Auclair, Carter, 1993; Schultze et al., 2012; Fultz et al., 2016).
В лесах Центральной Эвенкии основной лесообразующей породой является лиственница Larix gmelinii (Rupr.) Rupr., формирующая в этих условиях монодоминантные древостои. Ее ареал почти полностью совпадает с зоной сплошного распространения многолетней мерзлоты в Средней и Восточной Сибири, где она занимает 81.3% лесопокрытой площади (Абаимов и др., 1997; Прокушкин, Абаимов, 2008).
Тренд повышения активности пожаров и горимости лесов Сибири подтверждается данными разных лет (Kharuk et al., 2005; Loupian et al., 2006; Пономарев, Харук, 2016). Северотаежные лиственничные леса отличаются повышенной горимостью, которая обусловлена слабой пожарной расчлененностью территории, своеобразным температурным и гидрологическим режимом мерзлотных почв, разреженностью древесного полога и замедленными процессами разложения органического вещества (Софронов, Волокитина, 1996; Zyryanova et al., 2010). Повторяемость пожаров в лиственничниках Центральной Эвенкии составляет 60–100 лет, и большая часть современных лесных ландшафтов исследуемой территории представляет собой разные стадии послепожарных сукцессий (Абаимов и др., 1997).
Устойчивые низовые пожары, характерные для данной территории (Харук, Пономарев, 2017), способны полностью уничтожить растительный покров и верхние горизонты почвы, после чего образуются так называемые “черные гари”, для которых характерен весьма неблагоприятный термический режим (Абаимов и др., 2001; Прокушкин и др., 2000). И это существенно затрудняет восстановление растительного покрова на первых этапах постпирогенной сукцессии.
Изучению процессов восстановительной динамики растительности на гарях в Центральной Эвенкии посвящена обширная литература (Цветков, 1990; Цветков и др., 2001; Цветков, Цветкова, 1995; Зырянова и др., 2008; Zyryanova et al., 2010; Прокушкин и др., 2010), однако подробное описание первых этапов развития всех компонентов растительного покрова до настоящего времени не проводилось.
Обязательными участниками детритных пищевых цепей наземных экосистем являются раковинные амебы – представители нанофауны, имеющие раковинки размером от 10 до 300 мкм. Виды раковинных амеб имеют различные экологические преферендумы по условиям увлажнения и характеризуются чуткой реакцией на их изменение, а благодаря наличию устойчивой к разложению раковинки, используются в качестве биоиндикаторов (Количественные …, 1987; Гельцер и др., 1995).
В данной работе сделана попытка проследить динамику восстановления растительности и сообществ раковинных амеб на лиственничных гарях разного возраста в условиях северной тайги Центральной Эвенкии.
ОБЪЕКТЫ И МЕТОДИКА
Район исследований относится к Нижне-Тунгусскому округу Ангаро-Тунгусской лесорастительной провинции Средне-Сибирской плоскогорной лесорастительной области (Коротков, 1994). Исследования проводили на базе Эвенкийского стационара Института леса им. В.Н. Сукачева СО РАН в окрестностях пос. Тура Эвенкийского муниципального района Красноярского края в период 2016–2018 гг.
Объектами исследования служили лиственничные гари различного возраста, образовавшиеся в результате воздействия сильных устойчивых низовых пожаров, уничтоживших древостой и живой напочвенный покров полностью (Г15, Г13, Г09) или почти полностью (Г93). Характеристика объектов исследования приведена в таблице 1, в названии гарей использованы даты последнего пожара. К 2018-у году гарь 2015 года (Г15) была трехлетней, Г13 – пятилетней, Г09 – девятилетней, Г93 – 25-летней. К моменту начала наблюдений (2016 г.). Гари Г15 и Г13 представляли собой “черные гари”, микрорельеф которых сформировался в результате делювиального смыва и представляет собой чередование бугров-микроповышений и эрозионных борозд-микропонижений глубиной 15–20 см. 9-летняя гарь (Г09) отличается своеобразным микрорельефом, поскольку сформировалась на крупнообломочных продуктах выветривания туфогенных пород с размерами отдельных глыб до 2 м. Поверхность глыб обнажена в местах временных водотоков, а в остальных местах перекрыта слоем формирующейся почвы. 25-летняя гарь (Г93) в отличие от рассмотренных выше, пройдена, по-видимому, более слабым низовым пожаром (Абаимов и др., 1996) и характеризуется наличием допожарной генерации древостоя, представленной единичными довольно крупными плодоносящими лиственницами диаметром до 15–18 см. Микрорельеф выражен слабо, почвы более легкого гранулометрического состава.
Таблица 1.
Общая характеристика объектов исследования
| Гарь | Координаты | Экспозиция склона | Крутизна, ° | Абсолютная высота, м | Микрорельеф | Ассоциация |
|---|---|---|---|---|---|---|
| Г15 | 64°17′14.7″ с.ш., 100°18′51.2″ в.д. |
Юго-западная | 4–5 | 397 | Эрозионные борозды глубиной 15–20 см | Chamerion angustifolium (Calamagrostis lapponica) – Ceratodon purpureus |
| Г13 | 64°19′31.1″ с.ш., 100°15′26.2″в.д. |
Западная | 4–5 | 199 | Эрозионные борозды глубиной 5–15 см | Chamerion angustifolium (Calamagrostis lapponica ) – Ceratodon purpureus – Peltigera aphthosa |
| Г09 | 64°14′41.2″ с.ш., 100°09′47.7″ в.д. |
Северо-западная | 4–5 | 205 | Курумник | Duschekia fruticosa (Larix gmelinii) – Ledum palustre (Vaccinium vitis-idaea) – Ceratodon purpureus |
| Г93 | 64°19′45.6″ с.ш., 100°13′53.4″ в.д. |
Юго-восточная | 3–4 | 218 | Слабовыра-женный | Larix gmelinii (Betula pendula) – Vaccinium vitis-idaea (Ledum palustre, V. uliginosum) – Polytrichum piliferum – Cladonia cornuta |
Основной почвенный фон исследуемой территории составляют подбуры, они занимают не только склоновые позиции, но и междуречья, перекрытые трапповой формацией. Характеризуются хорошим дренажом и оттаиванием мерзлоты в конце вегетационного периода на всю глубину почвенного профиля (Безкоровайная и др., 2017).
На каждой гари закладывались пробные площади размером 400 м2, на которых проводилось полное геоботаническое описание растительного покрова (Методы …, 2002). Характеристика подроста, подлеска и живого напочвенного покрова проводилась на двух учетных профилях 2 × 20 м, заложенных в пределах пробной площади. Учетные профили разбивались на площадки 2 × 2 м, на каждой проводился сплошной перечет подроста, к которому относили растения высотой менее 2 м. Возраст подроста лиственницы определялся по мутовкам. Латинские названия сосудистых растений приведены согласно работе (Флора Сибири, 1988–1997), мхов – по (Игнатов, Игнатова, 2003).
В составе естественного возобновления выделяли всходы (проростки) – растения первого года жизни, характеризующиеся наличием зародышевых структур, имеющие главный корень, гипокотиль, семядоли и верхушечную почку, и собственно подрост – молодые растения, находящиеся в ювенильном и прематурном состоянии (простота организации, несформированность признаков, присущих взрослому растению, или наличие переходных признаков) (Восточноевропейские …, 2004).
Непосредственные наблюдения за послепожарным восстановлением растительности проводились на гарях Г15 и Г13 в период с 2016 по 2018 гг. Для реконструкции динамики естественного возобновления лиственницы на гарях Г09 и Г93 использовали показатели возрастной структуры подроста лиственницы.
Определение типов жизненных стратегий растений живого напочвенного покрова проводилось по классификации Л.Г. Раменского (1938; цит. по Миркин, Наумова, 2012) с уточнениями для мхов по (Сакович, Рыковский, 2012).
Для изучения процесса восстановления сообществ раковинных амеб после пожара на гарях отбиралось по 4–6 проб подстилки с площади 20 × 20 см каждая, по 2–3 на каждом элементе микрорельефа. Для каждой точки отбора проведено измерение температуры подстилки во время отбора проб. Естественная влажность субстрата определена путем взвешивания проб в сыром виде и после высушивания до воздушно-сухого состояния в лаборатории. Подготовка проб к ризоподному анализу проводилась по стандартной методике: 10 г влажного образца помещали в колбу на 100–150 мл, заливали произвольным количеством воды и оставляли на сутки, далее хорошо встряхивали и пропускали через сито с размером ячейки 0.25 мм для отделения крупных растительных остатков. Фильтрат отстаивали сутки, затем надосадочную жидкость сливали, доводили объем осадка до 10 мл (Гельцер и др., 1985). Пробы водной суспензии просматривали под микроскопом “Микромед 2” при увеличении ×200 и ×400. Определение видов раковинных амеб осуществлялось при помощи практических руководств (Гельцер и др., 1985; Мазей, Цыганов, 2006). В каждой пробе было идентифицировано не менее 300 экземпляров раковинок (за исключением свежей гари – Г15, год отбора – 2016, отличающейся наименьшей относительной плотностью раковинных амеб, где было идентифицировано 100 амеб). Полученные величины пересчитаны на 1 г воздушно-сухого вещества (экз г–1 в.с.в.).
Статистическая обработка данных проводилась с помощью стандартного статистического пакета Excel. Для оценки достоверности различий между средними значениями использовался однофакторный дисперсионный анализ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На лиственничных гарях в Центральной Эвенкии режим увлажнения и температурный режим в микропонижениях более благоприятен для растений, чем на микроповышениях (Прокушкин и др., 2000; Абаимов и др., 2001; Зырянова и др., 2008), поэтому на гари Г15 именно эрозионные борозды послужили инициальными местообитаниями, в которых происходило наиболее активное развитие пионерных видов растений.
На следующий год после пожара растительный покров гари был чрезвычайно разрежен (общее проективное покрытие (ОПП) <1%). В бороздах отмечалось развитие пирофитных пионерных видов-эксплерентов – печеночника маршанции многообразной (Marchancia polymorpha L.), слоевища которого местами сплошь покрывали их дно, хохлатки сибирской (Corydalis sibirica (L. f.) Pers.), малины сахалинской (Rubus sachalinensis Levl.), шиповника (Rosa acicularis Lindley). Всходы иван-чая (Chamerion angustifolium (L.) Holub.), а также вейник (Calamagrostis lapponica (Wahlb.) Hartm.), отрастающий от сохранившихся после пожара корневищ, не имели приуроченности к конкретным элементам нанорельефа, встречаясь как по бороздам, так и вне их. Малина сахалинская, по-видимому, возобновлялась из почвенного банка семян. Действительно, близкий вид малина обыкновенная (Rubus idaeus L.), известна своей способностью формировать долговременные почвенные банки семян, что обусловливает активное заселение ею гарей в зоне хвойных бореальных лесов (Марков, 2012). Густота всходов малины была в 4 раза больше, чем шиповника, и составляла 4.2 тыс. шт. га–1, однако из-за небольших размеров растений общее проективное покрытие составляло менее 1%.
На второй год после пожара общее проективное покрытие растений достигло 50–55%. Началось зарастание бугров-микроповышений за счет развития плотных куртин пирофитного листостебельного мха Ceratodon purpureus (Hedw.) Brid. (проективное покрытие (ПП) = 5–10%), а также иван-чая (ПП = 15–20%) и вейника (ПП = 10–15%). К концу третьего года эти виды сохранили доминирующие позиции, общее проективное покрытие растений увеличилось до 60–70%, сформировалась вейниково-кипрейная гарь. К этому моменту сформировалась рыхлая подстилка мощностью до 2 см, состоящая преимущественно из опавших сухих коробочек иван-чая, выполняющая роль мульчи.
Ольховник (Duschekia fruticosa (Rupr.) Pouzar), который является важнейшим компонентом зрелых лиственничников, в первый год единично отрастал от сохранивших жизнеспособность корневищ, однако, побеги были слабыми и в дальнейшем погибли. Черная смородина (Ribes nigrum L.) зафиксирована единично на второй послепожарный год. К третьему году видовой состав подлесочных пород на гари не изменился, но проективное покрытие шиповника и малины увеличилось до 5 и 10% соответственно. Оба вида плодоносили.
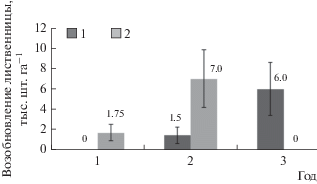
Размещение всходов и подроста древесных пород на гари крайне неравномерно, значения коэффициентов вариации достигают 130–150%, поэтому статистически достоверные различия густоты подроста по годам отсутствуют. Всходы Larix gmelinii как в первый послепожарный год, так и в дальнейшем обнаруживались исключительно в бороздах-микропонижениях. В первый год были зафиксированы только всходы густотой 1.75 тыс. шт. га–1 (рис. 1). На второй год появилась категория подроста (1.5 тыс. шт. га–1), а количество всходов составило 7 тыс. шт. га–1. На третий год было обнаружено около 6 тыс. шт. га–1 подроста, однако, всходы лиственницы полностью отсутствовали. Таким образом, в третьем послепожарном году на гари 2015 года абсолютно доминировало (92%) поколение лиственничного подроста, появившееся через два года после пожара.
Рис. 1.
Динамика естественного возобновления лиственницы на гари Г15 (1–3-й годы после пожара). На рис. 1 – подрост 2 – всходы.
Одновременно с лиственницей наблюдалось и семенное возобновление березы, всходы которой обнаруживались тоже только в бороздах. В первый послепожарный год густота всходов березы была такой же, как и у лиственницы, и составляла 1.75 тыс. шт. га–1. Однако на второй год практически все однолетние растения погибли, и вновь появились всходы – около 750 шт. га–1. На третий год процесс естественного возобновления березы протекал слабо – количество всходов и подроста было менее 1 тыс. шт. га–1.
Параллельно с восстановлением растительности шел процесс заселения гари раковинными амебами. В первый послепожарный год комплекс раковинных амеб в субстрате из озоленной подстилки включал всего 5 видов, относительная плотность тоже была очень низкой – 50–52 экз г–1 в.с.в. (рис. 2). Численно доминировали эврибионты с мелкими (менее 45 мкм) филозными раковинками (рис. 3): Trinema lineare (64%) и Corythion dubium (21%), Corythion orbicularis (7.5%), единично отмечен мелкий педобионт Schoenbornia humicola (1.6%) и один гидрофильный сфагно-бриобионт с крупной раковинкой Placocista glabra (5.9%). Присутствие мелких форм в целом типично для горевших биотопов (Turner, Swindles, 2012; Малышева и др., 2014; Курьина, Климова, 2016; Смолянинова, Гренадерова, 2018), так как на “черных гарях” в результате резкого уменьшения альбедо происходит сильное прогревание и иссушение субстрата. Это способствует развитию преимущественно мелких, быстро размножающихся ксерофилов, довольствующихся мельчайшими запасами влаги, раковинки которых легко разносятся ветром и не требуют морфологических адаптаций к условиям среды (Lousier, 1974; Корганова, 1977), что в целом характерно для эксплерентов. Кроме того, известно, что Trinema lineare и Schoenbornia humicola широко распространены в ландшафтах высоких широт северного полушария (Bobrov, Wetterich, 2012).
Рис. 2.
Относительная плотность и число видов раковинных амеб на гарях разного возраста. На рис. 2–3 в скобках указано число лет после пожара.
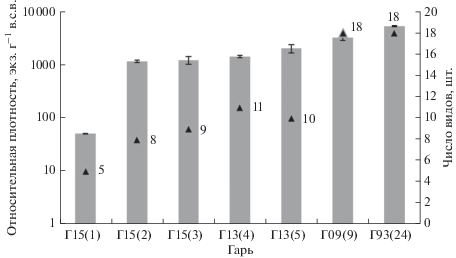
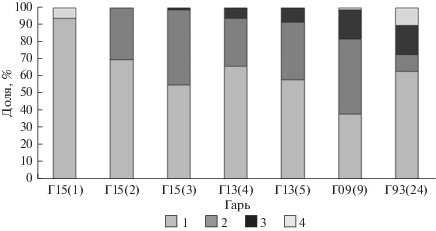
Рис. 3.
Соотношение экологических групп раковинных амеб на гарях разного возраста. На рис. 1 – филозные, эврибионты, 2 – лобозные, эврибионты, 3 – мезофильные, 4 – гигрофильные.
Присутствие вида Placocista glabra – крупной гидрофильной амебы – может быть связано с пятнами маршанции, слоевища которой удерживают влагу и местами сплошь покрывают дно эрозионных борозд.
На второй год после пожара число видов раковинных амеб возросло до 8, а относительная плотность увеличилась более чем в 20 раз и составила в среднем 1160 экз г–1 в.с.в. В этот год также доминировали мелкие виды, их доля составила 70%. Однако резко увеличилось (до 30%) относительное обилие более крупных (лобозных) форм (Cyclopyxis eurystoma, Centropyxis aerophila), относящихся к эврибионтам, способным переживать периоды иссушения в почве (Гельцер и др., 1985; Мазей, Цыганов, 2006).
На третий год после пожара видовое богатство сообществ раковинных амеб увеличилось незначительно и составило 9 видов, однако, существенно изменилась видовая структура. Мелкие эврибионтные формы сохранили доминирующие позиции, но их доля сократилась до 55%, зато участие лобозных форм увеличилось до 45%, в том числе единично встречены ксеро-мезофильные Padaungiella lageniformis (бриобионт) и Cyclopyxis kahli (брио-педобионт).
Известно, что среди абиотических факторов, влияющих на распределение раковинных амеб, ведущее значение принадлежит влажности (Lousier, 1974). Предполагалось, что более высокое видовое разнообразие и плотность будут наблюдаться именно в микропонижениях - эрозионных бороздах, где фиксировалось повышенное значение естественной влажности субстрата (66% против 45% на буграх). Однако анализ распределения амеб по элементам микрорельефа показал, что максимальная плотность (1340 экз г–1 в.с.в.) была отмечена на поверхности бугра, где наблюдалось и максимальное видовое богатство (8 видов). В эрозионных бороздах плотность составила в среднем 1140 экз г–1 в.с.в. и было обнаружено всего 5–6 видов. Статистически достоверные различия отсутствуют. Ранее (Абаимов и др., 2001) указывалось, что на буграх в подстилке температура на 3–5°С выше, чем в понижениях. Действительно, различия температур в день отбора проб на поверхности бугра и борозды составили 4°С (27°С и 23°С соответственно), а на границе с органо-минеральным слоем – 2.1°С (16.3°С и 14.2°С). Таким образом, несмотря на повышенную влажность, периодический вынос субстрата и более низкие температуры в борозде, возможно, не способствуют созданию благоприятных трофических условий для амеб.
Гарь Г13 в 2016 году была трехлетней и имела типичный для этой стадии вейниково-кипрейный живой напочвенный покров (ОПП = 85–90%) со значительным участием Ceratodon purpureus (ПП = 40–45%). В последующие 2 года происходило внедрение новых видов травянистых растений с более патиентной стратегией (Carex media R. Br., Equisetum scirpoides Michx.), появился первый представитель листоватых лишайников – Peltigera aphthosa (L.) Willd.), проективное покрытие которого было весьма значительным – до 5–10%. Представители рода Peltigera наряду с трубчатыми и бокальчатыми лишайниками отмечаются в качестве индикаторов начальных стадий постпирогенных сукцессий в лиственничниках лесотундры Западной Сибири (Морозова и др., 2007; Замараева, 2012). При этом Peltigera преимущественно осваивает куртинки цератодона, где задерживается влага. Кустарнички (Ledum palustre L., Vaccinium uliginosum L., V. vitis-idaea L.) – типичные доминанты напочвенного покрова развитых лиственничников – последовательно увеличивали обилие, и к 2018 году их суммарное проективное покрытие составило 10–15%, доминировала брусника.
Видовой состав подлесочных пород на гари Г13 включает единичные особи ольховника вегетативного происхождения из допожарной генерации, малину, шиповник, а также иву (Salix phylicifolia Pall.), проективное покрытие которой к пятому послепожарному году составило 5–7%.
Естественное возобновление лиственницы на гари Г13 протекало весьма успешно. Густота ее подроста к моменту начала наблюдений составляла около 15 тыс. шт. га–1 (рис. 4). Однако сложные гидротермические условия, характерные для первых стадий зарастания “черных гарей” (Зырянова и др., 2008), приводят к гибели части молодых растений. Так, густота подроста лиственницы в течение последующих двух лет снижалась на 5–6 тыс. шт. га–1 ежегодно и к пятому послепожарному году составляла всего около 4.5 тыс. шт. га–1 (различия статистически достоверны, Fф > Fкрит). При этом процесс естественного возобновления не прекращался – ежегодно на пробной площади фиксировалось 1–2 тыс. шт. га–1 всходов лиственницы.
Рис. 4.
Динамика естественного возобновления лиственницы на гари Г13 (3–5-й год после пожара). См. пояснения к рис. 1.
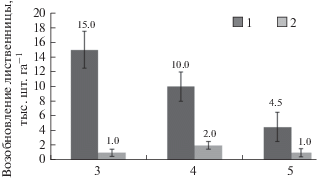
Возрастная структура лиственничного подроста отражает динамику его появления на гари. В составе подроста на пятилетней гари (Г13) преобладают особи 3-летнего (31%) и 4-летнего (56%) возраста. Это поколения, появившиеся в течение первых двух послепожарных лет. В дальнейшем условия для сохранения подроста на гари, по-видимому, ухудшились - доля 1–2-летних растений в совокупности составляет лишь 12%. Береза возобновляется слабо, ее подрост единичен.
Процесс послепожарного восстановления растительного покрова сопровождается увеличением мощности (до 2 см) и степени разложения подстилки, которая на этой стадии еще не дифференцирована на подгоризонты, однако улучшает условия влагообеспеченности для нанофауны. Спустя 4–5 лет после пожара на гари Г13 видовое богатство раковинных амеб увеличилось до 10–11 видов, а относительная плотность превысила 2000 экз. г–1 в.с.в. (рис. 2). На этой стадии отмечается стабильное (6–8%) участие мезофильных видов (например, Nebela tincta, Padaungiella lageniformis) в составе сообществ (рис. 3).
В живом напочвенном покрове 9-летней гари (Г09) еще сохранились виды, свидетельствующие о пирогенном происхождении сообщества: довольно обильны Rubus sachalinensis, Ceratodon purpureus, Chamerion angustifolium. Однако доминирующую роль играют кустарнички (особенно Ledum palustre), что свидетельствует о начале восстановления характерных для данного региона кустарничково-зеленомошных лиственничников. В составе моховой синузии доминируют пирофитные Ceratodon purpureus и Polytrichum juniperinum Hedw., однако в небольшом обилии (ПП = 1–2%) появляются виды, характерные для зрелых сообществ – Aulacomnium palustre (Hedw.) Schwägr., A. turgidum (Wahlenb.) Schwägr., Dicranum elongatum Schleich. ex Schwägr. При этом типичные для зрелых лиственничников доминанты, например, бриовиолент Pleurozium schreberi (Brid.) Mitt., еще отсутствуют. Лишайники немногочисленны и представлены преимущественно листоватыми и бокальчатыми формами.
Видовой состав подлеска включает те же виды, что и на пятилетней гари, но доминирующая роль переходит к ольховнику (проективное покрытие около 5%).
Послепожарное возобновление лиственницы приурочено к участкам между камнями, микропонижениям. Густота подроста составила 28 тыс. шт. га–1, при этом всходы зафиксированы не были. Анализ возрастной структуры подроста показал, что особи, появившиеся на гари в первый-второй послепожарные годы, единичны и на данный момент уже вышли из категории подроста, сформировав молодой древостой высотой около 3 м. В структуре естественного возобновления на данной гари преобладают 3–4-летние особи, появившиеся на пятый и шестой послепожарные годы, после чего естественное возобновление резко пошло на спад – доля поколений седьмого-восьмого послепожарных лет составляет менее 10%. Береза в составе естественного возобновления занимает незначительное место – около 1 тыс. шт. га–1 при средней высоте 0.7 м.
В ходе пирогенной сукцессии на Г09 в микропонижениях увеличилась мощность подстилки (до 3 см) и стал заметным подгоризонт F. В сообществе раковинных амеб общее число видов достигло 18, среди них Euglypha rotunda, E. strigosa glabra, E. tuberculata, Alabasta militaris, Nebela wailesi, Trigonopyxis arcula, средняя относительная плотность тоже существенно увеличилась и достигла 3228 ± 380 экз. г–1. в.с.в. Значительная доля мезо- и гигрофильных видов (рис. 3) может быть обусловлена не только накоплением органики, но и большей влажностью субстрата, характерной для склонов северных экспозиций.
Растительный покров 25-летней гари (Г93) отличается мозаичностью. В живом напочвенном покрове доминируют типичные для зрелых северотаежных лиственничников кустарнички, однако восстановления типичного мохово-лишайникового яруса еще не произошло, сообщество находится на стадии пирофитных мхов и шиловидных и бокальчатых лишайников. На сильно прогоревших участках развиты микроассоциации из пирофитных мхов (Ceratodon purpureus, Polytrichum juniperinum Hedw.) и лишайников (Peltigera aphthosa, Cladonia cornuta (L.) Hoffm., C. fimbriata (L.) Fr.), на сохранившихся лучше – зеленые мхи (Pleurozium schreberi, Dicranum polysetum Sw.) с Empetrum nigrum L. и кустистыми лишайниками (Cladonia rangiferina (L.) F. H. Wigg., C. stellaris (Opiz) Pouzar et Vězda, Flavocetraria cucullata (Bellardi) Kärnefelt et A. Thell). Характерно полное отсутствие типичного для лиственничников подлеска из ольховника.
Большинство авторов (Цветков, Цветкова, 1995; Прокушкин и др., 2000; Зырянова и др., 2008) отмечают, что возобновительный процесс в послепожарных лиственничниках продолжается в первые 7–10 лет и затем затухает, что обусловливается преимущественно восстановлением на гари зеленомошно-кустарничкового растительного покрова. Однако на гари Г93 процесс естественного возобновления продолжался значительно дольше. Густота подроста лиственницы в момент учета составила около 2 тыс. шт. га–1. В составе подроста обнаружены особи от 4-летнего до 11-летнего возраста, представленные приблизительно в равных долях (около 15%). Таким образом, в настоящее время на этой гари в составе естественного возобновления присутствуют поколения лиственницы, появившиеся в период с 14 по 21-й послепожарные годы, причем по площади он распределен крайне неравномерно.
В сообществе раковинных амеб на гари Г93 спустя 24 года после пожара общее число видов составило 18, а средняя относительная плотность достигла 5320 ± 110 экз. г–1. в.с.в. (рис. 2).
Мозаичность растительного покрова обусловливает своеобразие комплекса доминантов сообществ раковинных амеб, включающего наряду с преимущественно ксерофильными Corythion dubium, Schoenbornia humicola, Trinema complanatum, и ксеро-мезофильные Nebela tincta, Assulina seminulum. В микробиотопах с преобладанием плевроциума, брусники и багульника отмечается участие влаголюбивых видов Arcella vulgaris, A. jurassica, Euglypha strigosa glabra. Тем не менее доминантами (63%) по-прежнему являются ксерофильные виды, что можно объяснить летним иссушением мохово-лишайникового яруса и подстилки северных редколесий (Софронов, Волокитина, 1996).
ЗАКЛЮЧЕНИЕ
На начальных этапах заселения “черных гарей” в сообществах растений и раковинных амеб абсолютно доминируют виды-эксплеренты. Это бриоэксплеренты Marchancia polymorpha, Ceratodon purpureus, травы Corydalis sibirica, Chamerion angustifolium, Calamagrostis lapponica, кустарник Rubus sachalinensis. В сообществах раковинных амеб отмечены мелкие филозные эврибионты: Trinema lineare, Corythion dubium.
При заселении свежих гарей с выраженным микрорельефом инициальными местообитаниями для большинства видов растений служат эрозионные борозды, тогда как пионерные сообщества раковинных амеб могут интенсивнее заселять бугры-микроповышения.
Интенсивность и характер естественного возобновления лиственницы Гмелина зависят не только от возраста гари, но и от комплекса складывающихся на ней эдафо-ценотических условий. Наиболее интенсивно возобновительный процесс протекает в первые 2–3 года после пожара. В дальнейшем интенсивность естественного возобновления резко снижается, но оно может продолжаться до 7–8 и даже до 20 лет.
В ходе восстановительной постпирогенной сукцессии в живом напочвенном покрове пионерные растения-эксплеренты постепенно вытесняются видами с патиентной и виолентной стратегиями, способными длительно удерживать свои позиции в сообществах. Через промежуточную стадию брусники, пирогенных долгомошников (Polytrichum juniperinum, P. piliferum), листоватых (Peltigera aphthosa), шиловидных и бокальчатых (Cladonia cornuta, C. fimbriata) лишайников восстанавливается господство типичных для зрелых северотаежных лиственничников зеленых мхов-бриовиолентов (Pleurozium schreberi, Aulacomnium palustre и др.) и кустарничков (Ledum palustre, Vaccinium vitis-idaea, V. uliginosum).
Развитие растительного покрова и подстилки на гарях, сопровождающееся улучшением гидротермического режима, вызывает изменения в структуре сообществ раковинных амеб, выражающееся в появлении более влаголюбивых форм. На старых гарях мозаичность растительного покрова обусловливает существование широкого спектра морфо-экологических групп амеб – от мелких ксерофильных до крупных влаголюбивых форм.
Авторы благодарят доцента кафедры водных и наземных экосистем Сибирского федерального университета (г. Красноярск), канд. биол. наук И.П. Филиппову за помощь в определении мхов и лишайников, научного сотрудника Лаборатории мониторинга лесных экосистем Института мониторинга климатических и экологических систем СО РАН (г. Томск), канд.биол.наук И.В. Курьину за консультации при выполнении ризоподного анализа.
Список литературы
Абаимов А.П., Прокушкин С.Г., Зырянова О.А. Эколого-фитоценотическая оценка воздействия пожаров на леса криолитозоны Средней Сибири // Сибирский экологический журнал. 1996. № 1. С. 51–60.
Абаимов А.П., Прокушкин С.Г., Зырянова О.А., Каверзина Л.Н. Особенности формирования и функционирования лиственничных лесов на мерзлотных почвах // Лесоведение. 1997. № 5. С. 13–23.
Абаимов А.П., Прокушкин С.Г., Зырянова О.А., Каназава Ю., Такахаши К. Экологическая и лесообразующая роль пожаров в криолитозоне Сибири // Там же. 2001. № 5. С. 50–59.
Безкоровайная И.Н., Борисова И.В., Климченко А.В., Шабалина О.М., Захарченко Л.П., Ильин А.А., Бескровный А.К. Влияние пирогенного фактора на биологическую активность почв в условиях многолетней мерзлоты (Центральная Эвенкия) // Вестник Красноярского государственного аграрного университета. 2017. № 9. С. 181–189.
Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004. Кн. 1. 479 с.
Гельцер Ю.Г., Корганова Г.А., Алексеев Д.А. Практическое руководство по идентификации почвенных тестаций. М.: Изд-во МГУ, 1985. 84 с.
Гельцер Ю.Г., Корганова Г.А., Алексеев Д.А. Определитель почвообитающих раковинных амеб (практическое руководство). М.: Изд-во МГУ, 1995. 88 с.
Замараева Т.А. Возобновление шиловидных и бокальчатых лишайников после пожаров в лиственничных лесах лесотундры Западной Сибири // Вестник Тюменского государственного университета. 2012. № 6. С. 104–108.
Зырянова О.А., Абаимов А.П., Чихачёва Т.Л. Влияние пожаров на лесообразовательный процесс в лиственничных лесах севера Сибири // Лесоведение. 2008. № 1. С. 3–10.
Игнатов М.С., Игнатова Е.А. Флора мхов средней части европейской России. М.: Товарищество научных изданий КМК, 2003. Т. 1. С. 1–608; Т. 2. С. 611–960.
Количественные методы в почвенной зоологии / Отв. ред. Гиляров М.С., Стриганова Б.Р. М.: Наука, 1987. 290 с.
Корганова Г.А. Адаптационные особенности почвообитающих раковинных амеб (Protozoa, Testacida) // Адаптация почвенных животных к условиям среды. М.: Наука, 1977. С. 82–102.
Коротков И.А. Лесорастительное районирование России и республик бывшего СССР // Углерод в экосистемах лесов и болот России / Под ред. Алексеева В.А., Бердси Р.А. – Красноярск: Изд-во Институт леса СО РАН, 1994. С. 29–47.
Курьина И.В. Климова Н.В. Сообщества раковинных амеб (Rhizopoda, Testaceafilosea, Testacealobosea) в болотных местообитаниях после воздействия пожаров (юг Западной Сибири) // Вестник Томского государственного университета. 2016. Серия Биология. С. 161–181.
Мазей Ю.А., Цыганов А.Н. Пресноводные раковинные амебы. М.: Товарищество научных изданий КМК, 2006. 300 с.
Малышева Е.А., Цыганов А.Н. Бабешко К.В., Новенко Е.Ю., Мазей Ю.А. Видовой состав и структура сообществ сфагнобионтных раковинных амеб в болотных экосистемах Мордовского государственного природного заповедника // Труды Мордовского государственного природного заповедника им. П.Г. Смидовича. Саранск: Пушта, 2014. Вып. 12. С. 330–336.
Марков М.В. Популяционная биология растений. М.: Товарищество научных изданий КМК, 2012. 387 с.
Методы изучения лесных сообществ / Отв. ред. Ярмишко В.Т., Лянгузова И.В. СПб.: Изд-во Санкт-Петербургский государственный университет, 2002. 240 с.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: Гилем, 2012. 488 с.
Морозова Л.М., Эктова С.Н., Рябицева Н.Ю. Восстановление растительного покрова на гарях в зоне лесотундры // Экосистемы Субарктики: структура, динамика, проблемы охраны. Салехард. 2007. Вып. 6(50). Ч. 1. С. 17–32.
Пономарев Е. И., Харук В.И. Горимость лесов Алтае-Саянского региона в условиях наблюдаемых изменений климата // Сибирский экологический журнал. 2016. № 1. С. 38–46.
Прокушкин С.Г., Абаимов А.П. Толерантность лиственницы Гмелина к гипотермии // Хвойные бореальной зоны. 2008. Т. 25. № 3–4. С. 196–202.
Прокушкин С.Г., Сорокин Н.Д., Цветков П.А. Экологические последствия пожаров в лиственничниках северной тайги Красноярского края // Лесоведение. 2000. № 4. С. 9–15.
Прокушкин С.Г., Бугаенко Т.Н., Прокушкин А.С., Шкикунов В.Г. Сукцессионная трансформация растительного и почвенного покрова на солифлюкционных площадях в криолитозоне Центральной Эвенкии // Известия РАН. Серия биологическая. 2010. № 1. С. 95–104.
Сакович А.А., Рыковский Г.Ф. Анализ жизненных стратегий мохообразных в составе бриофлоры фортификаций линии Молотова (Гродненский район) // Актуальные проблемы экологии: Матер. VIII междунар. научн.-практич. конф. (Гродно, 24–26 октября 2012 г.). Гродно: Гродненский гос. университет, 2012. Ч. 1. С. 63–65.
Смолянинова С.Э., Гренадерова А.В. Сообщества раковинных амеб в подстилке сосняков Юксеевского лесничества после воздействия пожара // Вестник Красноярского государственного аграрного университета. 2018. № 5. С. 307–313.
Софронов М.А., Волокитина А.В. Пожары растительности в зоне северных редколесий // Сибирский экологический журнал. 1996. № 1. С. 43–49.
Флора Сибири. Новосибирск: Наука. 1988. Т. 1. С. 200; 1990. Т. 2. С. 361; 1990. Т. 3. С. 280; 1987. Т. 4. С. 250; 1992. Т. 5. С. 312; 1993. Т. 6. С. 310; 1994. Т. 7. С. 312; 1988. Т. 8. С. 200; 1994. Т. 9. С. 280; 1996. Т. 10. С. 254; 1997. Т. 11. С. 296; 1996. Т. 12. С. 208; 1997. Т. 13. С. 472; 2003. Т. 14. С. 188.
Харук В.И., Пономарев Е.И. Пространственно-временная горимость лиственничников Центральной Сибири // Экология. 2017. № 6. С. 413–419.
Цветков П.А. Возобновление на гарях в лиственничниках Центральной Эвенкии // Лесоведение. 1990. № 1. С. 62–67.
Цветков П.А., Сорокин Н.Д., Прокушкин С.Г., Каверзина Л.Н., Сорокина О.А., Цветкова Г.М. Эдафические условия и лесовосстановление после пожаров в лиственничниках Эвенкии // Там же. 2001. № 2. С. 16–21.
Цветков П.А., Цветкова Г.М. Послепожарное возобновление в северотаежных лиственничниках // Лесн. хоз-во. 1995. № 6. С. 44–47.
Abaimov A.P., Zyryanova O.A., Prokushkin S.G., Koike T., Matsuura Y. Forest Ecosystems of the cryolithic zone of Siberia; regional features: mechanisms of stability and pyrogenic changes // Eurasian Journal of Biosciences Foundation for Enviromental Protection and Research. 2000. № 1. P. 1-10.
Auclair A.N. D., Carter T.B. Forest wildfires as a recent source of CO2 at northern latitudes // Canadian Journal of Forest Research. 1993. V. 23. № 8. P. 1528–1536.
Bobrov A.A., Wetterich S. Testate amoebae of arctic tundra landscapes // Protistology. 2012. V. 7. № 1. P. 51–58.
Fultz L.M., Moore-Kucera J., Dathe J., Davinic M., Perry G., Wester D. & Rideout-Hanzak S. Forest wildfire and grassland prescribed fire effects on soil biogeochemical processes and microbial communities: Two case studies in the semi-arid Southwest // Applied Soil Ecology. 2016. V. 99. P. 118–128.
Kharuk V.I., Dvinskaya M.L., Ranson K.J. The spatiotemporal pattern of fires in northern taiga larch forests of Central Siberia // Russian J. Ecology. 2005. V. 36. № 5. P. 302–311.
Loupian E.A., Mazurov A.A., Flitman E.V., Ershov D.V., Korovin G.N., Novik V.P., Abushenko N.A., Altyntsev D.A., Koshelev V.V., Tashchilin S.A., Tatarnikov A.V., Csiszar I., Sukhinin A.I., Ponomarev E.I., Afonin S.V., Belov V.V., Matvienko G.G., Loboda T. Satellite Monitoring of Forest Fires in Russia at Federal and Regional Levels // Mitigation & Adaptation Strategies for Global Change. 2006. V. 11. P. 113–145.
Lousier J.D. Responce of soil testacea to soil moisture fluctuations // Soil Biology and Biochemistry. 1974. V. 6. № 4. P. 235–239.
Schulze E.-D., Wirth C., Mollicone D., N. von Lüpke, Ziegler W., Achard F., Mund M., Prokushkin A., Scherbina S. Factors promoting larch dominance in central Siberia: fire versus growth performance and implications for carbon dynamics at the boundary of evergreen and deciduous conifers // Biogeosciences, 2012. V. 9. P. 1–17.
Turner T.E., Swindles G.T. Ecology of testate amoebae in moorland with a complex fire history: implications for ecosystem monitoring and sustainable land management // Protist. 2012. V. 163. P. 844–855.
Zyryanova O.A., Abaimov A.P., Bugaenko T.N., Bugaenko N.N. Recovery of Forest Vegetation After Fire Disturbance // Permafrost Ecosystems. Siberian Larch Forests. Springer Dordrecht Heidelberg London New York. Springer Science+Business Media B.V., 2010. P. 83–94.
Дополнительные материалы отсутствуют.