

Лесоведение, 2021, № 4, стр. 426-436
Изменение свойств дерново-карбонатных почв после сплошной рубки в хвойно-широколиственных лесах Северо-Западного Кавказа
К. Ш. Казеев a, *, В. П. Солдатов a, А. К. Шхапацев b, Н. Е. Шевченко c, Е. А. Грабенко d, О. Ю. Ермолаева a, С. И. Колесников a
a Южный федеральный университет
344090 Ростов-на-Дону, Стачки, 194/1, Россия
b Майкопский государственный технологический университет
385000 Майкоп, Первомайская, д. 191, Республика Адыгея, Россия
c Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, Профсоюзная, 84/32, стр. 14, Россия
d Институт географии РАН
119017 Москва, Старомонетный пер., 29, стр. 4, Россия
* E-mail: kamil_kazeev@mail.ru
Поступила в редакцию 23.07.2020
После доработки 12.01.2021
Принята к публикации 02.04.2021
Аннотация
В статье рассматриваются результаты изменения за 9-летний период свойств горно-карбонатных почв после сплошной вырубки в хвойно-широколиственных лесах Северо-Западного Кавказа. В первые 2–3 года после сплошной рубки на участках со слабым и средним уровнями нарушения почвенного покрова, которые занимают большую часть территории вырубки, формируется высокотравная растительность с повышенным видовым разнообразием по сравнению с контрольными участками леса. Существенно изменяются свойства дерново-карбонатных почв исследуемой территории, особенно физические (температура, влажность, плотность сложения, сопротивление пенетрации) и биологические (разнообразие флоры, численность микроорганизмов, содержание гумуса, активность ферментов). Значения показателей биологических свойств почвы при максимальном нарушении почвенно-растительного покрова непосредственно после рубки леса снижаются более, чем в 10 раз по сравнению с контрольными участками леса. При слабом уровне нарушений вследствие проявления экотонного эффекта отмечены случаи повышения биологической активности почв на 20–50% и более. Биологические свойства почв, особенно ферментативная активность, проявили себя как очень чувствительный индикатор изменений, произошедших вследствие рубки. В ходе восстановительной сукцессии за десятилетний период на участках со средним и сильным нарушением почвенного покрова содержание органического углерода и ферментативная активность почв не достигли контрольных значений.
Леса Западного Кавказа являются одними из самых разнообразных и продуктивных природных экосистем России. Пихтовые и пихтово-буковые леса требуют особого внимания при ведении лесного хозяйства в Краснодарском крае и Адыгее, так как они в наименьшей степени нарушены антропогенным воздействием и могут считаться эталонными. Однако эти леса в настоящее время подвергаются значительному антропогенному прессу, связанному с повышающейся рекреационной нагрузкой, строительством баз отдыха, дорог, линий электропередач и т.д. В бассейне р. Белой естественная растительность верхнего горного пояса образована в основном двумя формациями с участием темнохвойных пород: пихтовой и буково-пихтовой, причем последняя преобладает. Рубки лесов приводят к изменению растительности и животного мира, деградации почвенного покрова, развитию эрозионных процессов (Дымов, 2017). В горах в условиях гумидного климата и рассеченного рельефа рубка леса может приводить к полной деградации почв в результате уничтожения лесной подстилки, обесструктуривания, уплотнения, смыва почв и т.д. Общая площадь территории вырубок в Краснодарском крае по расчетам М.В. Придня с соавт. (2009) составила 5539 км2 и достигает 38% всей площади лесов в регионе, что в 3 раза превышает площадь лесов Кавказского биосферного заповедника. Время восстановления биоразнообразия в буковых лесах Кавказа после рубки составляет 50–55 лет (Придня, Ромашин, 2001; Щербина, 2006). В дубравах украинского Полесья, сходных по условиям произрастания с дубравами Северного Кавказа, при большей обеспеченности теплом и светом этот процесс занимает около 40–45 лет (Ткачук, 2007).
Лесные экосистемы, формирующиеся на известняковых породах, имеют широкое распространение на территории Западного Кавказа и значительно отличаются от таковых на бескарбонатных породах. Сложенные юрскими известняками Фишт-Оштенский массив и Лагонакское нагорье представляют большой интерес из-за разнообразия условий местообитания, растительных сообществ и богатства флоры. Почвы известняковых массивов Кавказа – это дерново-карбонатные почвы (рендзины), которые как азональные встречаются во многих районах земного шара (Вальков и др., 2007). В лесных зонах Северного Кавказа эти почвы распространены на площади более 1.2 млн. га в горных территориях под разными растительными ассоциациями на карбонатном элювии известняков, доломитов и мергелей (Вальков и др., 2008). В Классификации и диагностике почв России (2004) года они обозначены как темногумусовые карболитоземы, согласно Мировой реферативной базе почвенных ресурсов WRB – Rendzik leptosol. В условиях Западного Кавказа рендзины встречаются среди зональных бурых и серых лесных почв. Генезис дерново-карбонатных почв в значительной мере отличается от условий почвообразования зональных бурых и серых лесных почв (Вальков и др., 2007; Казеев и др., 2012).
Оценка качества почвы является важной составляющей мониторинга окружающей среды и, наряду с оценками техногенного загрязнения, включает весь комплекс экологических функций почв (Bünemann et al., 2018). Биологические свойства почв имеют большой потенциал для оценки экологического состояния почв (Колесников и др., 2014; Karlen et al., 2019; Kolesnikov et al., 2019; Казеев и др., 2020). В почвах вырубок может быть как активизация биологических процессов (Горшенин, Щербакова, 1972; Краснощеков, Сорокин, 1988; Adamczyk et al., 2015; Молчанов и др., 2017), так и снижение их интенсивности (Казеев и др., 2012). Вследствие рубок изменяется ферментативная активность почв (Казеев и др., 2012; Trasar-Cepeda et al., 2008; Paz-Ferreiro et al., 2010; Brackin et al., 2013). Рубки способствуют оживлению процессов минерализации органических веществ, о чем свидетельствует повышение активности уреазы (Горшенин, Щербакова, 1972). При этом усиливаются окислительно-восстановительные процессы, диагностируемые по активности дегидрогеназ и каталазы. Активность инвертазы в почве вырубок снижается, что, возможно, связано с уменьшением количества корней, служащих основными ее поставщиками в почве. Однако есть данные, свидетельствующие о слабой связи ферментативной активности с изменениями в содержании углерода и азота в почве, что можно объяснить значительной ролью абиотической регуляции активности ферментов на вырубках (McDaniel et al., 2013). На содержание органического вещества и биологические параметры в лесных почвах значительное влияние оказывает сукцессионный статус (Дымов, 2017; Лукина и др., 2018; Шевченко и др., 2019) и пожары (Дымов и др., 2018; Богородская и др., 2019).
Цель работы – исследовать динамику изменения физических и биологических свойств дерново-карбонатных почв известняковых массивов Западного Кавказа после рубки леса.
ОБЪЕКТЫ И МЕТОДИКА
Характеристика лесных насаждений и лесорастительных условий
Исследуемая территория расположена в 10 км от поселка Гузерипль (Республика Адыгея) на высоте 1635 м. над ур. моря. Климатические условия характеризуются повышенным увлажнением, количество осадков за год составляет в среднем 1795 мм. Среднегодовая температура равна 9.8°С, температура июля 19°С, января –3°С (Козунь и др., 2013). Основные лесообразующие породы – бук восточный (Fagus orientalis Lipsky) и пихта кавказская (Abies nordmanniana (Steven) Spac). Формула преобладающего состава леса – 7.8П2.1Бк (Грабенко, Татаренко, 2010). Средний диаметр древостоя у пихты от 24 до 80 см (в среднем 44.4 см), бука от 8 до 44 см (в среднем 28.2 см). Среднее значение густоты – 665 деревьев на 1 га.
Вырубка 2010 г. представляет собой выровненный участок верхней части склона, полностью лишенный растительности. Поверхность почвы на вырубке очень сильно нарушена тяжелой техникой. Для таких сильнонарушенных почв предложено новое таксономическое определение турбоземов детритных на механически нарушенных участках лесосек (Дымов, 2017). Исследования проводили в 2010–2020 гг. на участках вырубки с разной степенью нарушения почвенного покрова. Для этого были выделены участки со слабым (I), средним (II), сильным (III) и очень сильным (IV) уровнем антропогенного нарушения почвы. Критерием выделения были площадь и степень повреждения, определяемые по мощности скальпирования почвы, ее перемешивания и погребения в результате работы тяжелой техники. В большинстве случаев степень нарушения уменьшалась от дороги (очень сильная) до границы вырубки (слабая). В качестве контроля (0) взят участок буково-пихтового леса, граничащий с вырубкой. Почва дерново-карбонатная выщелоченная слабокаменистая суглинистая на элювии известняков (карболитозем темногумусовый).
Методы исследования
Геоботанические описания проводились по общепринятым методикам в соответствии со стандартными подходами (Миркин, Наумова, 2012). Видовая принадлежность растений определялась по региональным определителям (Косенко, 1970; Зернов, 2006). Латинские названия сосудистых растений даны по С.К. Черепанову (1995). Обилие видов оценивалось по шкале Ж. Браун-Бланке (Миркин и др., 2001): r – вид на площадке встречен в единичном экземпляре; + – вид имеет проективное покрытие до 1%; 1 – вид имеет покрытие от 1 до 5%; 2 – от 5 до 25%; 3 – от 25 до 50%; 4 – от 50 до 75%; 5 – выше 75%.
Экспедиционные и лабораторно-аналитические исследования выполнены с использованием общепринятых в экологии, биологии и почвоведении методов. Были исследованы температура, влажность, плотность почв, заложены разрезы и прикопки, определена численность микроорганизмов, ферментативная активность. Влажность почвы в лаборатории определяли весовым методом и в полевых условиях влагомером Thetaprobe (EIJKELKAMP, Нидерланды) в 10-кратной повторности на каждом участке. Температуру почв определяли послойно при отборе образцов (0, 5, 10, 20, 30 см) электронным термометром HANNA CHECTEMP (HANNA, Германия). Плотность почвы определяли объемно-весовым методом в 3-кратной повторности. Твердость почв – сопротивление пенетрации – определяли в полевых условиях пенетрометром EIJKELKAMP (Нидерланды) до глубины 50 см каждые 5 см в 10-кратной повторности.
Активность каталазы, инвертазы и дегидрогеназы определяли по разложению соответствующих субстратов: перекиси водорода, сахарозы и трифенилтетразолия хлористого. Содержание гумуса определяли бихроматным методом по И.В. Тюрину в модификации Никитина (Казеев и др., 2016). Численность аммонифицирующих бактерий определяли посевом на мясо-пептонном агаре. Численность микроскопических грибов учитывали на подкисленной среде Чапека. Обилие азотфиксирующих бактерий рода Azotobacter определяли на среде Эшби методом комочков обрастания. Общая численность микроорганизмов и водорослей определена люминесцентно-микроскопическим методом в 3–6-кратной повторности. Для определения различий в уровне биогенности и биологической активности разных почв определяли интегральный показатель биологического состояния (ИПБС) почвы. Этот показатель оценивает совокупность биологических показателей, выраженных в разных единицах, и позволяет нивелировать случайные колебания, характерные для большинства биологических параметров. Для расчета ИПБС за 100% принимается максимальное значение каждого из показателей и по отношению к нему в процентах выражается значение этого же показателя в остальных образцах:
Б1 = (Бх/Бmax) × 100%,
где Б1 – относительный балл показателя, Бх – фактическое значение показателя, Бmax – максимальное значение показателя.
После этого рассчитывают средний оценочный балл изученных показателей ИПБС почвы – аналогично расчету относительного балла показателя.
Статистическая обработка результатов исследования проведена с использованием статистического пакета Statistica 10.0 и Excel.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Ботаническая характеристика в зависимости от степени нарушенности территории
Уже через год на вырубке появился травяной покров, отмечено снижение повреждений (колеи, борозд и др.) от работы тяжелой техники. Через три года после рубки леса в июне 2013 г. на контрольной площадке в лесу насчитали 20 видов растений с доминированием Abies nordmanniana и Fagus orientalis . В 2015 г. видовой состав насчитывал 37 видов (таблица 1). Первый ярус (древесный) образует пихта Нордманна (Abies nordmanniana (Stev.) Spach) со значительным участием бука восточного (Fagus orientalis Lipsky) и клена Траутфеттера (Acer trautvetteri Medw.). Кроме этих деревьев в состав древесного яруса в незначительном количестве примешиваются рябины (Sorbus aucuparia L., S. caucasica Zinserl.). Высота древесного яруса в среднем составляет 30 м, сомкнутость 0.5. Подъярус (высота 5–15 м, сомкнутость 0.2–0.4) слагают те же виды деревьев: Abies nordmanniana (Stev.) Spach., Fagus orientalis Lipsky, Acer trautvetteri Medw., Sorbus aucuparia L., S. caucasica Zinserl. Кустарниковый ярус не выражен, но из кустарников изредка встречается жимолость восточная (Lonicera orientalis Lam.). Разреженный древесный полог, а также хорошее увлажнение создают благоприятные условия для развития травяного яруса. Второй ярус (травяной) имеет проективное покрытие более 80% и среднюю высоту травостоя – 70 см. Обычными видами с проективным покрытием до 50% (3) являются бор раскидистый (Milium effusum L.), купена мутовчатая (Polygonatum verticillatum (L.) All.), подмаренник душистый (Galium odoratum (L.) Scop.), щитовник мужской (Dryopteris filix-mas (L.) Schott), мятлик длиннолистный (Poa longifolia Trin.).
Таблица 1.
Сравнительная характеристика фитоценозов вырубки и контрольного участка
| Исследуемые участки | Контроль, лес (0) | Слабое нарушение (I) | Среднее нарушение (II) | Сильное нарушение (III) | Очень сильное нарушение (IV) |
|---|---|---|---|---|---|
| Тип фитоценоза | Лесной | Опушечно-лесной | Луговой | Луговой | Пионерное |
| Ярусность | 2 | 3 | – | – | – |
| Древесный ярус | Abies nordmanniana Fagus orientalis Acer trautvetteri |
Sorbus aucuparia, S. caucasica, Acer trautvetteri |
– | – | – |
| Кустарниковый ярус | не выражен | Rubus caucasicus, Ribes Biebersteini, Lonicera orientalis |
– | – | – |
| Травяной ярус | Milium effusum, Polygonatum verticillatum, Galium odoratum и др. |
Petasites hybridus, Polygonatum verticillatum и др. | Poa longifolia, Symphytum asperum, Petasites hybridus и др. |
Galega orientalis, Poa longifoliaSymphytum asperum и др. | Rumex confertus, Plantago major Trifolium ambiguum, Carex contigua |
| Высота травостоя, см | 70 | 120 | 155 | 95 | 36 |
| Проективное покрытие травяного яруса, % | 80 | 100 | 100 | 95 | 10 |
| Число видов | 37 | 51 | 36 | 29 | 14 |
На участках вырубки, имеющих слабую степень антропогенной нагрузки, видовой состав более разнообразен и насчитывает 51 вид растений. Вертикальная структура сообщества трехъярусная. 1-й ярус (древесный) состоит из рябин (Sorbus aucuparia L., S. caucasica Zinserl) и клена Траутфеттера (Acer trautvetteri Medw.). Единично представлены пихты (Abies nordmanniana (Steven) Spach) и буки (Fagus orientalis Lipsky.). На этих площадках отмечены в незначительном числе яблони (Malus orientalis Uglizk.). Древесный ярус имеет неравномерную плотность (сомкнутость 0.5–0.3), его высота варьирует от 35 до 15 м. 2-й ярус (кустарниковый) выражен хорошо, его проективное покрытие до 60%. Его образует ежевика кавказская (Rubus caucasicus Focke) и смородина кавказская (Ribes biebersteinii Berl.), менее обильна жимолость восточная (Lonicera orientalis Lam.). Наибольшую степень проективного покрытия имеет ежевика кавказская (Rubus caucasicus Focke), покрывающая на отдельных участках до 50% площади. 3-й ярус (травяной) выражен хорошо, его проективное покрытие составляет 100%. Высота травостоя варьирует от 100 до 166 см и в среднем составляет 120 см. Видами, имеющими наибольшее обилие среди травянистых растений, являются белокопытник гибридный (Petasites hybridus (L.) Gaertn. B. Mey. & Scherb.) и мятлик узколистный (Poa longifolia Trin.). Среди других видов наиболее представлены виды субальпийского высокотравья (2): ясколка даурская (Cerastium davuricum Fisch. Ex Spreng.), щитовник мужской (Dryopteris filix – mas (L.) Schott.), гравилат городской (Geum latilobum Somm. et Levier.), ясменник кавказский (Asperula caucasica Pobed), бутень золотистый (Chaerophyllum aureum L.) и др.
Сообщества, имеющие среднюю степень антропогенной нагрузки, обеднены по составу (36 видов) и, по сути, представляют собой субальпийские высокотравные поляны. Из деревьев встречаются только клены Траутфеттера (Acer trautvetteri Medw.), единично отмечены пихта кавказская (Abies nordmanniana (Steven) Spach), бук восточный (Fagus orientalis Lipsky.) и яблоня (Malus orientalis Uglizk.). Из кустарников отмечена только ежевика кавказская (Rubus caucasicus Focke). Проективное покрытие травяного яруса около 100%, высота травостоя в среднем составляет 155 см в его основе окопник жесткий (Symphytum asperum Lepech.), белокопытник гибридный (Petasites hybridus (L.) Gaertn. B. Mey. & Scherb.) и мятлик длиннолистный (Poa longifolia Trin.). Для сообщества константными являются виды субальпийского высокотравья, характерные для обедненных и деградированных вариантов субальпийских высокотравных лугов, такие как козлятник восточный (Galega orientalis Lam.), ясколка даурская (Cerastium davuricum Fisch. Ex Spreng.) и др. Обычными являются такие высокотравные виды, как молочай длиннорогий (Euphorbia macroceras Fisch. et C.A. Mey.), девясил высокий (Inula helenium L.), лютик кавказский (Ranunculus caucasicus Bieb) и др. Снижение роли таких представителей пышного субальпийского высокотравья, как живокость опушенноплодная (Delphinium dasycarpum Stev. ex DC.), живокость пирамидальная (D. pyramidatum Albov), мытник черно-пурпурный (Pedicularis atrоpurpurea Nord.), герань лесная (Geranium sylvaticum L.) и др. свидетельствует о нарушенности этого сообщества. В тоже время снижение роли видов, характерных для нарушенных местообитаний при достаточной влажности почвы (например, Rumex alpinus L., Urtica dioica L., Cerastium davuricum Fisch. ex Spreng.), свидетельствуют о некотором восстановлении этих фитоценозов. На участках с максимальной степенью нарушения (в основном грунтовая дорога) растительность практически отсуствует.
Изменение физических свойств
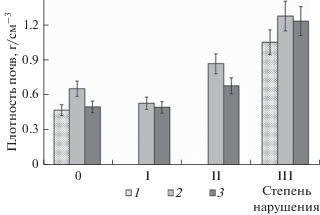
На исследуемой территории были выявлены значительные изменения физических свойств почв. Непосредственно после рубки леса плотность сложения почв вырубки значительно повысилась, по сравнению с значениями под лесом (рис. 1). Переуплотнение вызвано работой тяжелой техники, которая нарушила сложение горизонтов и привела к общему уплотнению почв. Изменение плотности почвы в результате вырубки леса сохраняется в течение всего времени наблюдения. При этом степень увеличения плотности почв зависела от уровня нагрузки на исследуемую территорию. Повышенной плотностью обладают почвы с максимальным уровнем антропогенного воздействия. Через три года плотность сложения почвы на участках вырубки со средней и высокой степенью деградации также была выше (0.8–1.3 г см–3), чем в почве контрольного участка (0.8 г см–3). В то же время на участке со слабым нарушением плотность почвы была меньше, чем на контрольном. Это связано с увеличением поступления растительных остатков в почву вследствие формирования высокотравной флоры при лучшей освещенности на периферии вырубки. Поэтому здесь формируется более мощный органогенный горизонт и, в связи с обильным ростом травяной растительности, больший объем почвы пронизан корнями.
Рис. 1.
Влияние рубки леса на плотность сложения почв (0–10 см) с разной степенью нарушения почвенного покрова, 2010–2019 г. 1 – 2010 г., 2 – 2012 г., 3 – 2019 г.
Уплотнение почвы на вырубке приводит и к изменению ее твердости, измеренной по сопротивлению проникновению (пенетрации). Этот показатель отражает противодействие, которое испытывают при росте корни растений. Его повышение негативно влияет на продуктивность растений. В результате исследований было выявлено повышение значений показателя на нарушенных участках вырубки (рис. 2). При этом в первый год исследований на вырубке были отмечены инверсии профильного распределения твердости почв, связанные с нарушениями естественного сложения почвы.
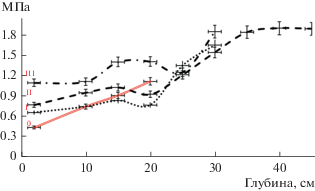
Изменение биологической активности
Рубка леса привела к снижению значений различных биологических показателей: интенсивности дыхания почвы, активности почвенных ферментов и содержания гумуса. Изменения биологических свойств дерново-карбонатных почв касаются не только верхних горизонтов, но, прежде всего, характера их распределения по почвенному профилю. Биологическая активность снижается вниз по профилю рендзин в меньшей степени, чем в зональных горно-лесных почвах. Это объясняется карбонатностью почвообразующих пород рендзин, которая повышает рН, содержание элементов питания и гумуса (Казеев и др., 2012). Кроме того, изменения профильного распределения биологических показателей связано с нарушением поверхности почв, кое-где с частичным скальпированием, а также с гидротермическим режимом почв исследуемых участков. Различия сохраняются в течение всего периода наблюдения и оказывают прямое воздействие на химические и биологические процессы. В связи с повышением уровня солнечной инсоляции на вырубке температура почв значительно выше, а влажность ниже, по сравнению с контрольными значениями в лесу. При этом повышение температуры почвы на вырубке не приводит к ее иссушению, что особенно важно для почвенной биоты и биологических процессов.
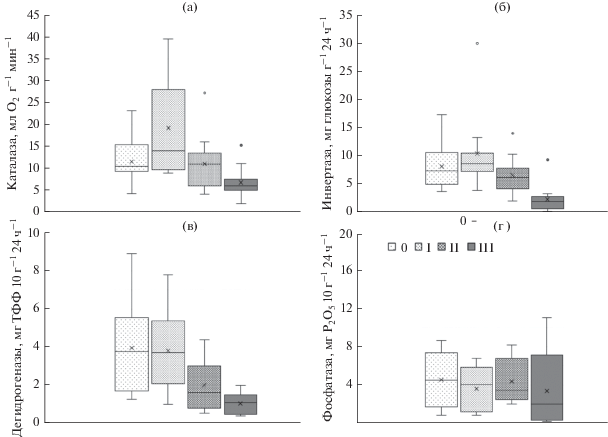
Ферментативная активность в течение всего срока наблюдения различается на участках вырубки в зависимости от степени нарушения и вида фермента (рис. 3). Значительный материал, усредненный за 10 лет, позволил показать значительное ингибирование активности всех исследуемых ферментов в почвах участков вырубки со средним и, особенно, сильным нарушением. Однако на участках со слабым нарушением, расположенных на периферии вырубки, активность каталазы и инвертазы достоверно выше контрольных значений. Активность фосфатазы и дегидрогеназ здесь достоверно не отличается от почвы под лесом. Особенно значительное изменение активности ферментов отмечено в первый год после вырубки леса. Активность гидролаз по сравнению с активностью оксидаз уменьшилась сразу после рубки леса и увеличилась через год. Это связано со сложным сочетанием гидротермических условий с началом сукцессионных изменений растительности, разным уровнем проявления эрозии и др. В дальнейшем активность ферментов разных групп приближается к контрольным значениям, за исключением дегидрогеназы, активность которой на участке со средним и высоким уровнем нарушения почвенного покрова через год снижается вдвое, через 2 года – в 11 раз, а активность инвертазы полностью ингибируется Высокое варьирование и разнонаправленность биологической активности зависит от множества факторов, включая степень нарушения, тип почвы, запас органических веществ, тип растительности (Молчанов и др., 2017).
Рис. 3.
Ферментативная активность почв вырубки с разной степенью нарушения почвенного покрова, 2010–2020 гг. (N = 30). а – каталаза, б – инвертаза, в – дегидрогеназы, г – фосфатаза. 0 – III – степень нарушения.
Применение интегральной оценки при диагностике влияния вырубки выявило существенное снижение значений ИПБС на нарушенном участке. Непосредственно после рубки леса уменьшение составило 28% для поверхностных горизонтов и 24% для всего профиля. Согласно классификации С.И. Колесникова с соавт. (2014) при вырубке леса и связанной с этим работы техники произошло значительное нарушение экологических функций почвы. Вследствие нарушения почвенного покрова, уничтожения лесной подстилки, увеличения плотности почвы произошло снижение общей мощности гумусового горизонта почвы на некоторых участках. Профильное изменение значений ИПБС отражает особенности внутрипрофильного распределения отдельных биологических показателей. В контрольной дерново-карбонатной почве следует типичное для лесных почв почти двукратное снижение ИПБС вниз по профилю на глубине 10–20 см с некоторым относительным увеличением на глубине полуметра, связанным, как уже отмечали ранее, с близостью карбонатных пород. Нарушение сложения почвы и утрата самого биогенного горизонта подстилки привела к тому, что на вырубке практически не было отмечено снижения значений ИПБС по почвенному профилю. Вслед за небольшим снижением значений ИПБС на глубине 10 см следует его относительный рост почти до значений поверхностного горизонта. Это явление в значительной мере отличает почву вырубки от почвы контрольного участка леса.
Мониторинг содержания гумуса в дерново-карбонатных почвах вырубки показал значительные различия на разных участках. Степень изменения зависит от уровня исходного нарушения почвенно-растительного покрова. Выявлены разные закономерности изменения содержания гумуса на участках вырубки со слабым, средним и сильным уровнем нарушений при вырубке и тралевке леса после 2010 г. (рис. 4). Возможно значительное сокращение содержания гумуса на участке с сильным нарушением поверхности почвы. При этом в условиях обильного увлажнения и рассеченного рельефа резко усиливаются процессы водной эрозии, которые приводят к смыву верхнего гумусированного горизонта, укорачиванию гумусового профиля, повышению каменистости почв, а, местами, выходу на поверхность карбонатного элювия и массивных плит известняка. Содержание гумуса при этом неуклонно снижается.
Рис. 4.
Динамика содержания гумуса на участках вырубки с разной степенью нарушения почвенно-растительного покрова (0–10 см), 2010–2019 гг. 0–III – степень нарушения.
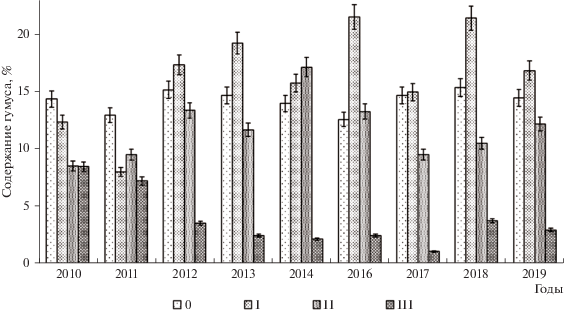
Второй вариант изменений – это постепенное восстановление содержания гумуса в почвах на участках со средним антропогенным нарушением. При этом снижение гумуса в первые годы после рубки леса сменяется его постепенным восстановлением в последующие годы при усилении роли травянистой флоры. Содержание гумуса при этом приближается к контрольным значениям, но степень его варьирования весьма высока, что связано с локальными особенностями рельефа и растительности.
На участках со слабым нарушением почвенно-растительного покрова происходит прогрессирующее увеличение содержание гумуса сверх контрольных значений уже через два года. Это связано с сукцессионным изменением растительности, сопровождаемым повышением разнообразия и продуктивности высокотравной растительности. Повышенная инсоляция на открытых после рубки леса пространствах приводит к бурному росту травяной растительности и, как следствие, усилению дернового и гумусо-аккумулятивного процессов. А замедленное разложение ее остатков вследствие короткого периода биологической активности приводит к быстрому образованию мощной оторфованной подстилки. Со временем дерново-карбонатные почвы могут переходить в перегнойно-карбонатные. В результате слабонарушенные участки за счет экотонного (опушечного) эффекта превосходят контрольные значения леса по биоразнообразию флоры и биологической активности почвы. Здесь отмечены случаи значительного повышения активности разных ферментов на вырубке относительно контрольных значений. Об активизации ферментов и дыхания почв на некоторых участках вырубок благодаря активным процессам разложения и минерализации мертвых корней и накопившегося древесного опада и отходов лесозаготовки сообщали и другие исследователи (Adamczyk et al., 2015; Молчанов и др., 2017). Кроме того на участках вырубки формируются микроклиматические условия, способствующие повышению биологической активности (Дымов, 2017; Молчанов и др., 2017). Некоторые показатели биологической активности, например почвенное дыхание, в значительной мере определяются влажностью почвы (Махныкина и др., 2020).
Динамика активности почвенных ферментов также различна в течение первых 10 лет после вырубки леса. Активность каталазы изменяется с такими же закономерностями, как и для содержания гумуса. Активность инвертазы и дегидрогеназ варьируют в значительно больших пределах, чем активность каталазы. Однако для обоих ферментов выявлен одинаковый характер динамики. Высокая степень нарушения приводит к значительному понижению значений практически без тренда к восстановлению. При слабом нарушении почв после первых лет значительного возрастания ферментативной активности на 70–200% значения дегидрогеназ и инвертазы начинают снижаться. Активность дегидрогеназ уже через 5 лет снижается ниже контроля, активность инвертазы, снижаясь, все также и через 9 лет выше контрольных значений примерно на 30%.
ВЫВОДЫ
1. Уровень деградации почвенных свойств зависит от степени нарушения почвенно-растительного покрова при рубке и тралевке леса. В почвах вырубки значительно увеличились плотность сложения почв и сопротивление пенетрации.
2. В первые годы после рубки леса на свободных участках формируется более разнообразная высокотравная горно-луговая растительность Разнообразие повышается, в основном, за счет высокотравных и синантропных видов.
3. В почвах участков с разной степенью нарушения может происходить разнонаправленное изменение содержания гумуса. При сильном нарушении снижение может превысить 80% от первоначального содержания. На участках слабого нарушения почв при развитии травянистой растительности содержание гумуса может достоверно увеличиться.
4. Активность ферментов в почвах вырубки в значительной степени различается в зависимости от степени нарушения, времени с момента рубки и вида фермента. В целом ферментативная активность после сведения леса ингибируется, однако возможно и повышение активности ферментов, особенно на начальном этапе зарастания вырубки и на участках слабого нарушения.
5. Значения ИПБС через 2 года после рубки леса снижаются в ряду лес (100) → слабое нарушение (92) → среднее нарушение (71) → сильное нарушение (59) → очень сильное нарушение (20). Через 10 лет на участках со значительным нарушением полного восстановления биологических свойств почв не произошло.
Список литературы
Богородская А.В., Кукавская Е.А., Каленская О.П., Буряк Л.В. Микробиологическая оценка состояния почв хвойных лесов Средней Сибири после пожаров разной интенсивности // Лесоведение. 2019. № 2. С. 138–156.
Вальков В.Ф., Казеев К.Ш., Колесников С.И. Почвы юга России: генезис, география, классификация, использование и охрана. Ростов-на-Дону: Изд-во Эверест, 2008. 276 с.
Вальков В.Ф., Казеев К.Ш., Колесников С.И., Кутровский М.А. Почвообразование на известняках и мергелях. Ростов-на-Дону: Ростиздат, 2007. 198 с.
Горшенин Н.М., Щербакова О.Н. Влияние рубок на биологическую активность дерново-подзолистой супесчаной почвы // Почвоведение. 1972. № 5. С. 115–123.
Грабенко Е.А., Татаренко Н.П. Зависимость лесорастительных и почвенных условий от абсолютной высоты в буково-пихтовых лесах Северо-Западного Кавказа // Вестник Майкопского государственного технологического университета. 2010. № 1. С. 134–138.
Дымов А.А. Влияние сплошных рубок в бореальных лесах России на почвы (обзор) // Почвоведение. 2017. № 7. С. 787–798.
Дымов А.А., Старцев В.В., Зуева О.М. Углерод водорастворимых соединений в лесных почвах и его постпирогенная динамика (на примере республики Коми) // Лесоведение. 2018. № 5. С.359–371.
Зернов А.А. Флора Северо-Западного Кавказа. М.: Товарищество научных изданий Изд-во: КМК, 2006. 664 с.
Казеев К.Ш., Колесников С.И., Акименко Ю.В., Даденко Е.В. Методы биодиагностики наземных экосистем. Ростов-на-Дону: Изд-во Южного фед. университета, 2016. 356 с.
Казеев К.Ш., Кутровский М.А., Даденко Е.В., Везденеева Л.С., Колесников С.И., Вальков В.Ф. Влияние карбонатности пород на биологические свойства горных почв Северо-Западного Кавказа // Почвоведение. 2012. № 3. С. 327–335.
Казеев К.Ш., Трушков А.В., Одабашян М.Ю., Колесников С.И. Постагрогенное изменение ферментативной активности и содержания органического углерода чернозема в первые 3 года залежного режима // Почвоведение. 2020. № 7. С. 901–910.
Классификация и диагностика почв России. Смоленск: Ойкумена. 2004. 342 с.
Козунь Ю.С., Казеев К.Ш., Колесников С.И. Влияние климата на биологические свойства почв юга России. Ростов-на-Дону: Изд-во Южного фед. университета, 2013. 112 с.
Колесников С.И., Жаркова М.Г., Везденеева Л.С., Кутузова И.В., Молчанова Е.В., Зубков Д.А., Казеев К.Ш. Оценка экотоксичности тяжелых металлов и нефти по биологическим показателям чернозема // Экология. 2014. № 3. С. 158–163.
Косенко И.С. Определитель высших растений Северо-Западного Кавказа и Предкавказья. М.: Колос. 1970. 614 с.
Краснощеков Ю.Н., Сорокин Н.Д. Почвенно-экологические изменения на вырубках и гарях Восточного Хэнтэя (МНР) // Почвоведение. 1988. № 1. С. 117–127.
Лукина Н.В., Тихонова Е.В., Шевченко Н.Е., Горнов А.В., Кузнецова А.И., Гераськина А.П., Смирнов В.Э., Горнова М.В., Ручинская Е.В., Анищенко Л.Н., Тебенькова Д.Н., Данилова М.А., Бахмет О.Н., Крышень А.М., Князева С.В., Шашков М.П., Быховец С.С., Чертов О.Г., Шанин В.Н. Аккумуляция углерода в лесных почвах и сукцессионный статус лесов. М.: Товарищество научных изданий КМК, 2018. 232 с.
Махныкина А.В., Прокушкин А.С., Меняйло О.В., Верховец С.В., Тычков И.И., Урбан А.В., Рубцов А.В., Кошурникова Н.Н., Ваганов Е.А. Влияние климатических факторов на эмиссию СО2 из почв в среднетаежных лесах Центральной Сибири: Эмиссия как функция температуры и влажности почвы // Экология. 2020. № 1. С. 51–61.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: Гилем, 2012. 488 с.
Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М: Логос., 2001. 264 с
Молчанов А.Г., Курбатова Ю.А., Ольчев А.В. Влияние сплошной вырубки леса на эмиссию СО2 с поверхности почвы // Известия РАН. Серия биологическая, 2017. № 2. С. 190–196.
Придня М.В., Ромашин А.В. Биологическое разнообразие лесов курортных комплексов Кубани и их оздоравливающее значение // Наука Кубани. 2001. №1. С. 3–10.
Придня М.В., Ромашин А.В., Пиньковский М.Д. Экосистемные услуги лесов Западного Кавказа // Успехи современного естествознания. 2009. №11. С. 9–20.
Рабочая группа IUSS WRВ. 2015. Мировая реферативная база почвенных ресурсов 2014, исправленная и дополненная версия 2015. Международная система почвенной классификации для диагностики почв и создания легенд почвенных карт. Доклады о мировых почвенных ресурсах № 106. ФАО, Рим. М.: Изд-во МГУ, 2018. 204 с.
Ткачук В.И. Динамика биоразнообразия в дубовых лесах влажных сугрудков центрального Полесья после сплошных вырубок // Лесоводство и агролесомелиорация., Харьков. 2007. Вып. 111. С. 73–79.
Французов А.А. Флористическая классификация лесов с Fagus orientalis Lipsky и Abies nordmanniana (Stev.) Spach в бассейне реки Белой (Западный Кавказ) // Растительность России. 2006. № 9. С. 76–85.
Черепанов С.К. Сосудистые растения России и сопредельных государств. Спб.: Мир и семья, 1995. 992 с.
Шевченко Н.Е., Кузнецова А.И., Тебенькова Д.Н., Смирнов В.Э., Гераськина А.П., Горнов А.В., Грабенко Е.А., Тихонова Е.В., Лукина Н.В. Сукцессионная динамика растительности и запасы почвенного углерода в хвойно-широколиственных лесах Северо-Западного Кавказa // Лесоведение. 2019. № 3. С. 163–176.
Щербина В.Г. Динамика деструкции листового опада в рекреационных буковых биогеоценозах. // Экологический вестник Северного Кавказа. 2006. Т. 2. № 2. С. 5–9.
Adamczyk B., Adamczyk S., Kukkola M., Tamminen P., Smolander A. Logging residue harvest may decrease enzymatic activity of boreal forest soils // Soil Biology & Biochemistry. 2015. V. 82. P. 74–80.
Brackin R., Robinson N., Lakshmanan P., Schmidt S. Microbial function in adjacent subtropical forest and agricultural soil // Soil Biology & Biochemistry 2013. V. 57. P. 68–77.
Bünemann E.K., Bongiorno G., Bai Z., Creamer R.E., De Deyn G., Goede R., Fleskens L., Geissen V., Kuyper T.W., Mäder P., Pulleman M., Sukke W.l, Groenigen J.W., Brussaard L. Soil Quality – A Critical Review // Soil Biology and Biochemistry. 2018. V. 120. P. 105–125.
Karlen D.L., Veum, K.S., Sudduth K.A., Obrycki J.F., Nunes M.R. Soil Health assessment: Past Accomplishments, Current Activities, and Future Opportunities // Soil & Tillage Research. 2019. V. 195. 104365.
Kolesnikov S.I., Timoshenko A.N., Kazeev K.S., Akimenko Y.V., Soldatov A.V. Comparison of ecotoxicity of nickel and iron oxides and their nanoforms // Rasayan J. Chemistry, 2019. V. 12(2). P. 549–553.
McDaniel M.D., Kaye J.P., Kaye M.W. Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest forest // Soil Biology & Biochemistry. 2013. V. 56. P. 90–98.
Paz-Ferreiro J., Trasar-Cepeda C., Leirós M.C., Seoane S., Gil-Sotres F. Effect of management and climate on biochemical properties of grassland soils from Galicia (NW Spain) // European J. Soil Biology. 2010. V. 46. I. 2. P. 136–143.
Trasar-Cepeda C., Leirós M.C., Gil-Sotres F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality // Soil Biology & Biochemistry, 2008. V. 40. I. 9. P. 2146–2155.
Дополнительные материалы отсутствуют.