

Лесоведение, 2021, № 4, стр. 379-392
Влияние рубок разной интенсивности на генетическое разнообразие подроста сосны в северной лесостепи Средней Сибири
И. В. Тихонова a, *, А. К. Экарт b, А. Н. Кравченко b, Н. А. Тихонова b, Д. А. Семенякин b
a Западно-Сибирское отделение Института леса им. В.Н. Сукачева СО РАН – филиал ФИЦ КНЦ СО РАН
663082 Новосибирск, ул. Жуковского, 100/1, Россия
b Институт леса им. В.Н. Сукачева СО РАН – обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: selection@ksc.krasn.ru
Поступила в редакцию 19.04.2019
После доработки 19.05.2020
Принята к публикации 06.04.2021
Аннотация
Представлены результаты анализа аллозимной изменчивости в популяциях сосны обыкновенной, произрастающих в Красноярской, Ачинской и Канской лесостепи Средней Сибири. Сравниваются основные показатели генетического разнообразия у подроста сосны обыкновенной, появившегося на вырубках разной интенсивности, и в контроле. Установлено сокращение числа редких и низкочастотных аллелей в полиморфных локусах изоферментных маркеров, среднего числа аллелей на локус у подроста, появившегося после сплошной рубки и выборочных рубок высокой интенсивности. Не выявлено существенных отклонений в генетическом разнообразии у подроста предварительной и последующих генераций после выборочной рубки интенсивностью 35%, проведенной через 10 лет после низового пожара. Из остальных вариантов рубок наименьшее сокращение числа редких аллелей наблюдалось у подроста после узколесосечной рубки. В наиболее нарушенных рубками популяциях отмечен самый низкий уровень гетерозиготности, высокие значения индекса фиксации Райта (F), наименьшее число аллелей изоферментных локусов, что в дальнейшем может привести к снижению продуктивности и устойчивости новых поколений деревьев в небольших популяциях сосны обыкновенной, произрастающих в лесостепной зоне, где ареал вида характеризуется высокой фрагментированностью. Рассматриваются методические аспекты организации генетического мониторинга хвойных лесов как необходимого дополнения в комплексе с лесохозяйственным мониторингом их нарушенности. Отмечена необходимость организации генетических резерватов во всех крупных лесостепных борах, а также в других местах ограниченного распространения вида по всему его ареалу.
Как известно, Красноярская, Ачинская и Канская лесостепь Средней Сибири является “островной” и расположена в окружении тайги (Берг, 1947). Ведущая роль в составе ее растительного покрова также отводится лесным фитоценозам (Антипова, 2012), лесистость варьирует от 20 до 50% – наименьшая в Канской лесостепи. Вместе с тем лесные массивы тяготеют к берегам крупных рек, занимаемая ими площадь обратно пропорциональна обилию населенных пунктов, и по мере удаления от городов и основных транспортных магистралей заметно сокращается доля распаханных земель, повышается лесистость территории, а лесостепь постепенно переходит в подтайгу и далее – в тайгу. Все это указывает на антропогенное происхождение северной (как, вероятно, и южной, с более длительной историей антропогенеза) лесостепи Средней Сибири, появившейся в результате вырубки южной тайги (Леса СССР, 1969; Побединский, 1973; Сочава, 1980), в отдельные эпохи голоцена территория была покрыта темнохвойными лесами, периодически сменяемыми светлохвойной подтайгой (Савина, 1986). Необходимо отметить, что в составе лесов здесь преобладают березняки и сосново-березовые леса, доля чистых сосняков невелика, они приурочены ближе к окраинам лесостепи. Поэтому их можно рассматривать в качестве резервного генофонда для восстановления светлохвойных лесов при смене экономической и лесохозяйственной политики России на более рациональную (национальную) ресурсосберегающую.
От интенсивных рубок пострадали как пригородные леса лесостепной зоны, так и значительная часть таежных лесов Сибири в целом (Сочава, 1980; Калашников, 1998; Бузыкин, Пшеничникова, 2008; Степанов, 2014). Это вызывает тревогу не только у экологов, но и лесопромышленников, учитывая, что большая часть вырубаемой древесины (до 70–80%) отправляется на экспорт (Синицких, 2015), вырубаются лучшие леса, они часто сменяются вторичными мелколиственными сообществами. При этом многие фундаментальные научные проблемы лесопользования мало изучены. Например, недостаточно исследовано влияние рубок на показатели генетического разнообразия популяций видов. Не разработаны общие методические подходы к мониторингу состояния генофондов лесных древесных видов (Ирошников и др., 1989; Fussi et al., 2016). Не исследованы пределы допустимого сокращения генетического разнообразия популяций для их полноценного воспроизводства и устойчивости. Эта проблема активно изучается за рубежом, начиная с 90-х годов ХХ века (Ratnam et al., 2014). В России и в странах бывшего СССР такие работы пока единичны (Динамика …, 2004; Падутов и др., 2008; Ильинов и др., 2010).
Цель данного исследования заключалась в изучении генетического разнообразия аллозимов (изоферментов) в сосняках северной лесостепи Средней Сибири и оценке влияния вариантов рубок на показатели генетического разнообразия у подроста.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в популяциях сосны обыкновенной, произрастающих вблизи населенных пунктов Красноярск, Сухая, Ачинск, Погорелка, Большая Мурта. Всего проанализировано 445 деревьев в 15 выборках, по 25–30 деревьев в каждой. В их числе 6 выборок подроста и 2 выборки взрослых деревьев из оставшихся на лесосеке на месте сплошных и выборочных рубок разной интенсивности: сплошной широколесосечной рубки 28-летней давности площадью 352 га – “Малый Кемчуг” и 8 га – “Ачинск” (проведена 5 лет назад), 35-летние культуры на месте сплошной широколесосечной рубки площадью 37 га “Погорелка”; сплошной узколесосечной вырубки 22-летней давности площадью 1 га – “Сухая”; выборочной рубки интенсивности 70% площадью 32 га (проведена 8 лет назад) – “Большая Мурта”, 35% и 50%, проведенной 9 лет назад методом узких лент шириной 25 м (пасеки) и технологическим коридором (волоком) 5 м, общей площадью 50 га – “Погорелка” (табл. 1). В определении типа вырубки мы учитывали его кратковременность, как одного из этапов процесса восстановления исходного типа леса после его нарушения (Солодухин, 1961). Следует отметить, что на большей части сплошной вырубки “Малый Кемчуг” были полностью удалены верхние горизонты почвы вместе с живым напочвенным покровом (и весь подрост), а подстилающие почву глинистые отложения спустя 28 лет после рубки сильно эродированы и травяной покров на них отсутствует. На участке выборочной рубки интенсивностью 35% за 10 лет до рубки прошел (или был проведен) низовой пожар, вызвавший активное возобновление сосны (28 тыс. шт. га–1). Так как все популяции расположены недалеко от населенных пунктов и подвержены длительному антропогенному влиянию (от рекреации и пожаров до рубок), для сравнительного анализа использовали 2 контрольных участка: ненарушенные недавней рубкой части популяций, и один общий региональный контроль – наименее нарушенная популяция (часть популяции) с наиболее высокими показателями генетического разнообразия. Предварительно оценивали генетическую изменчивость популяций вида в этой части ареала на контрольных, нетронутых рубками участках (9 популяций), и для ранее полученных нами данных (4 популяции) (Экарт и др., 2014). С этой целью всего проанализировано 676 деревьев, по 30–60 в каждой популяции. В дополнение к выше перечисленным, это пункты Тарутино, Назарово, Юксеево, Березовка, Есаулово, Кускун, Рыбное, Гмирянка, Канск.
Таблица 1.
Краткая характеристика популяций
| Популяция, давность рубки | Воз-растная группа | Выборка | Состав древостоя/ Тип вырубки |
Возраст, лет |
Полнота (густота у подроста, тыс. шт.га–1) |
Географические координаты | |
|---|---|---|---|---|---|---|---|
| с. ш. | в. д. | ||||||
| Красноярск, контроль |
В | 35 | 7С2Б1Л С. ос.разн.зелм. |
70–120 | 1.0 | 56°00′ | 92°45′ |
| Сухая, СУР, 20 лет | П | 30 | 6Б4Сед.Е С-Б. мох.ос.разн. |
10–20 | 48.0 | 56°13′ | 92°58′ |
| М. Кемчуг, СШР, 10 лет | П | 25 | 6Ос3Б1СедЕП Б-Ос. разн.вейн. |
5–8 | 0 0.1 |
56°12′ | 92°27′ |
| Ачинск, контроль | В | 29 | 10С | 80–150 | 0.7 | 56°18′ | 90°30′ |
| П | 30 | 7С2Б1Е С.разн.зелм. |
7–14 | 2.0 | 56°14′ | 90°33′ | |
| Ачинск, СШР, 5 лет | В | 30 | 6С4Б | 50–60 | 0.1 | 56°16′ | 90°33′ |
| П | 30 | 6Б3С1Ос С-Б. ос.вейн.разн. |
5–10 | 0.5 | |||
| Погорелка, | В | 28 | 10С | 50–120 | 1.0 | 56°22′ | 92°58′ |
| контроль | П | 30 | 10С С. бр.разн.зелм. |
5–18 | 5.6 | ||
| Погорелка СШР, 40 лет, культуры | В | 29 | 10С С. зелм.разн. |
35 | 1.3 | 56°23′ | 92°59′ |
| Погорелка ВР35, 5 лет | П | 30 | 10С С. кипр.бр.разн.зелм. |
5–10 | 28.0 | 56°22′ | 92°59′ |
| ВР50, 5 лет | П | 29 | 10С С. бр.зелм.вейн.разн. |
5–8 | 7.2 | ||
| Б. Мурта ВР70, 5 лет | П | 30 | 8С2Е С. разн.вейн.кипр. |
5–7 | 1.0 | 56°53′ | 93°05′ |
| Б. Мурта, контроль | В | 30 | 8С2Б | 40–100 | 0.8 | 56°53′ | 93°12′ |
| П | 30 | 10С С. мох.ос.разн. | 6–15 | 3.0 | |||
Примечание: СШР – сплошная широколесосечная вырубка, СУР – сплошная узколесосечная вырубка, ВР35 и ВР70 – выборочная рубка интенсивностью 35 и 70%. Возрастная группа: В – взрослые деревья, П – подрост. Состав древостоя и тип вырубки (тип леса в контроле): С – сосна, Л – лиственница, Е – ель, П – пихта, Б – береза, Ос – осина, бр. – брусничный, разн. – разноравный, зелм. – зеленомошный, ос. – осочковый, вейн. – вейниковый, кипр. – кипрейный.
С каждого дерева для электрофоретического анализа изоферментов осенью отбирали вегетативные почки. Проанализировано 20 локусов, 17 из них полиморфны при 99%, а 16 – при 95%-ном уровне значимости – это локусы ферментных систем малатдегидрогеназы (Mdh–2, Mdh–3, Mdh–4), глутаматоксалоацетаттрансаминазы (Got–1, Got–2, Got–3), шикиматдегидрогеназы (Skdh–1, Skdh–2), 6-фосфоглюконатдегидрогеназы (6-Pgd–2), формиатдегидрогеназы (Fdh), фосфоглюкомутазы (Pgm–1), глутаматдегидрогеназы (Gdh), флуоресцентной эстеразы (Fe–2), алкогольдегидрогеназы (Adh–1, Adh–2), лейцинаминопептидазы (Lap–1, Lap–2). Используемые в работе методы электрофоретического анализа изоферментов и гистохимического окрашивания были описаны нами ранее (Экарт и др., 2014).
Для каждой популяции с помощью программы GenAlex 6 (Peakall, Smouse, 2006) вычисляли следующие показатели генетической изменчивости: процент полиморфных локусов (Р), среднее (NА) и эффективное число аллелей на локус (NЕ), наблюдаемая (HО) и ожидаемая гетерозиготность (HЕ), несмещенная оценка гетерозиготности (UHe), индекс разнообразия Шенона (I), индекс фиксации Райта (F), число иммигрантов на поколение (Nm). Соответствие распределения генотипов уравнению Харди–Вайнберга также оценивали с помощью программы GenAlex 6, используя критерий χ2, по каждому локусу в каждой выборке, по всем локусам в каждой выборке и в целом по всем локусам, объединив все выборки. Степень генетической подразделенности между популяциями и внутри популяций определяли с помощью F-статистик Райта FIS (уровень инбридинга особи относительно популяции), FIT (уровень инбридинга особи относительно вида), FST (уровень инбридинга популяции относительно вида – подразделенность популяций) (Wright, 1978; Хедрик, 2003), их значимость оценивали также с помощью χ2. Степень различий между выборками определяли по генетическому расстоянию Неи (Nei, 1972), достоверность различий по частотам аллелей вычисляли c помощью точного критерия Фишера с использованием метода Марковских цепей для всего набора локусов и полокусно в программе Genepop (Raymond, Rousset, 1995). Также с помощью точного критерия Фишера вычисляли достоверность различий с контролем по частотам редких аллелей.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Основные показатели генетической изменчивости 13 популяций, без учета подроста с вырубок варьировали в пределах: Р = 70–85%, NА = 2.46 (2.25–2.65), NЕ = 1.392 (1.357–1.432), HО = 0.225 (0.202–0.248); HЕ = 0.224 (0.214–0.243), F = –0.002 (–0.069…0.072). Межпопуляционные различия, оцененные с помощью FST, составили 1.7%. На различия между группами популяций трех лесостепей приходится только 0.2%, внутри популяций остается 98.3% генетической изменчивости. Такая низкая дифференциация, в общем, характерна для изучаемого вида (Гончаренко и др., 1996; Санников и др., 2002; Семериков и др., 2014). В более широких географических пределах всей сибирской части ареала и Монголии различия между популяциями по аналогичному набору локусов достигают 4.5% (Экарт и др., 2014), наибольшие – по локусам Skdh–2 (15.8%), Adh–2 (7.1%), Pgm–1 (7.1%), Mdh–4 (6.2%).
В суммарной выборке деревьев всех популяций северной лесостепи Средней Сибири установлены достоверные отклонения в распределении генотипов 6 локусов относительно равновесия Харди–Вайнберга, в 5 из них (Mdh–3, Got–2, Skdh–1, Adh–2, Pgm–1) наблюдается превышение доли гомозиготных генотипов над гетерозиготными. По-видимому, это объясняется в большей степени эффектом Валунда – влиянием различий между популяциями по частоте встречаемости аллелей (Хедрик, 2003), а также инбридингом, связанным с их изолированностью друг от друга, значительно большей в лесостепной зоне, чем в таежной (Санников и др., 2002). Генетические дистанции Неи (Nei, 1972) между популяциями в границах исследуемой части ареала вида, рассчитываемые по частотам аллелей, составили 0.002–0.010, наибольшие – между популяциями, удаленными на расстояние ≥200 км. Более низкие значения Nm и более выcокие FIS и FIT (0.040–0.153) получены по 5 перечисленным выше локусам и локусам Gdh и Lap–2.
В связи с тем, что данное исследование не было запланированным экспериментом с предваряющим рубку детальным обследованием насаждений и анализом генетического разнообразия древостоев до рубки, не все особенности возобновления удалось учесть: исходную полноту, точный состав и строение древостоев, численное соотношение предварительного и последующего возобновления, длительность периода после рубки. На данном этапе перед нами стояла задача выяснить, могут ли сплошные и выборочные рубки оказать влияние на генетическую структуру одного из ветроопыляемых хвойных видов, приуроченных к первому этапу восстановительно-возрастных сукцессий. В дальнейшем, при условии полноценного бюджетного финансирования тематики исследования (размера регионального гранта, на который оно было проведено, для решения такой задачи недостаточно) можно было бы собрать необходимые данные, координируя работы с лесопользователями и детальнее проработав методическую сторону вопроса.
В 4 исследуемых популяциях сосны обыкновенной, где проводились рубки, показатели генетического разнообразия выборок подроста и взрослых деревьев в местном контроле и на вырубках варьировали в следующих пределах: Р = = 70–80%, NА = 2.303 (2.100–2.500), NЕ = 1.376 (1.288–1.433), HО = 0.217 (0.195–0.235); HЕ = 0.219 (0.191–0.243), F = 0.002 (–0.075–0.064) (табл. 2). В целом данные популяции сосны обыкновенной находятся в состоянии относительного генетического равновесия (по числу гомо- и гетерозиготных генотипов). Достоверные отклонения от равновесия Харди–Вайнберга при полокусном анализе отмечены в 8 выборках из 15 по 1–2 локусам. В обобщенном многолокусном анализе достоверное превышение гомозиготных генотипов над ожидаемым их числом обнаружено только в локусах Adh–2 (χ2 = 17.4, d. f. = 6, р < 0.008) и Lap–2 (χ2 = 29.2, d. f. = 10, р < 0.0012). Согласно результатам анализа AMOVA, 0.8% генотипической изменчивости приходится на межпопуляционные различия, 1.1% – на различия между выборками внутри популяций, подразделенность всех выборок по FST составляет 2.3%. Больший вклад в межпопуляционные различия вносит локус Adh–1 (FST= 3.0%, Nm = 8.0). Внутри популяций дифференциация между выборками наблюдалась по локусам: Mdh–3 (3.5%) и Skdh–1 (4.1%) в популяции “Красноярск”; Lap–1 (2.7%) в популяции “Погорелка”; Got–2 (4.2%), 6-Pgd–2 (6.5%), Pgm–1 (6.3%), Fe–2 (3.5%) в популяции “Ачинск”.
Таблица 2.
Основные характеристики генетической изменчивости внутри 4 популяций сосны обыкновенной
| Популяция: | Выборка | Возрастная группа | Р | NA | NE | I | HO | HE | UHE | F |
|---|---|---|---|---|---|---|---|---|---|---|
| Красноярск | ||||||||||
| Общая | 90 | 80 | 2.650 | 1.365 | 0.389 | 0.214 | 0.220 | 0.222 | 0.015 | |
| выборка | ±0.284 | ±0.081 | ±0.070 | ±0.041 | ±0.042 | ±0.042 | ±0.017 | |||
| МК | 35 | В | 80 | 2.400 | 1.364 | 0.384 | 0.229 | 0.22 | 0.226 | –0.035 |
| СУР | 30 | П | 75 | 2.350 | 1.376 | 0.384 | 0.204 | 0.220 | 0.224 | 0.027 |
| СШР | 25 | П | 75 | 2.250 | 1.352 | 0.364 | 0.203 | 0.208 | 0.211 | 0.041 |
| Погорелка | ||||||||||
| Общая | 146 | 85 | 3.000 | 1.388 | 0.406 | 0.219 | 0.228 | 0.228 | 0.021 | |
| выборка | ±0.363 | ±0.086 | ±0.075 | ±0.042 | ±0.044 | ±0.045 | ±0.013 | |||
| МК | 28 | В | 80 | 2.500 | 1.418 | 0.416 | 0.231 | 0.239 | 0.243 | 0.028 |
| МК | 30 | П | 75 | 2.300 | 1.359 | 0.381 | 0.205 | 0.212 | 0.215 | 0.011 |
| СШР Ку | 29 | В | 80 | 2.350 | 1.361 | 0.383 | 0.216 | 0.217 | 0.221 | –0.020 |
| ВР50 | 29 | П | 70 | 2.200 | 1.406 | 0.385 | 0.231 | 0.228 | 0.232 | –0.024 |
| ВР35 | 30 | П | 80 | 2.450 | 1.378 | 0.396 | 0.215 | 0.226 | 0.230 | 0.031 |
| Б. Мурта | ||||||||||
| Общая | 90 | 70 | 2.400 | 1.398 | 0.391 | 0.233 | 0.225 | 0.226 | –0.029 | |
| выборка | ±0.320 | ±0.095 | ±0.080 | ±0.050 | ±0.047 | ±0.047 | ±0.018 | |||
| МК | 30 | В | 70 | 2.300 | 1.382 | 0.408 | 0.233 | 0.214 | 0.217 | –0.075 |
| 30 | П | 70 | 2.350 | 1.434 | 0.419 | 0.235 | 0.243 | 0.247 | 0.030 | |
| ВР70 | 30 | П | 70 | 2.100 | 1.373 | 0.365 | 0.229 | 0.213 | 0.217 | –0.064 |
| Ачинск | ||||||||||
| Общая | 119 | 80 | 2.750 | 1.363 | 0.393 | 0.205 | 0.219 | 0.220 | 0.035 | |
| выборка | ±0.354 | ±0.081 | ±0.070 | ±0.038 | ±0.042 | ±0.042 | ±0.020 | |||
| МК | 29 | В | 75 | 2.450 | 1.390 | 0.391 | 0.215 | 0.223 | 0.226 | 0.012 |
| 30 | П | 70 | 2.300 | 1.396 | 0.390 | 0.217 | 0.223 | 0.227 | 0.017 | |
| СШР | 30 | В | 75 | 2.200 | 1.288 | 0.334 | 0.197 | 0.191 | 0.194 | –0.020 |
| 30 | П | 75 | 2.250 | 1.357 | 0.370 | 0.195 | 0.216 | 0.220 | 0.064 | |
Примечание: Возрастная группа: В – взрослые деревья, П – подрост. Показатели генетического разнообразия: P – процент полиморфных локусов, NA и NE – среднее и эффективное число аллелей на локус, I – индекс разнообразия Шенона, HO и HE – наблюдаемая и ожидаемая гетерозиготность, UHE – несмещенная оценка гетерозиготности, F – индекс фиксации Райта. Обозначения выборок: МК – местный контроль, РК – региональный контроль, СШР – сплошная широколесосечная вырубка, СУР – сплошная узколесосечная вырубка, СШР Ку – культуры на сплошной широколесосечной вырубке, ВР35, ВР50 и ВР70 – выборочная рубка интенсивностью 35, 50 и 70%.
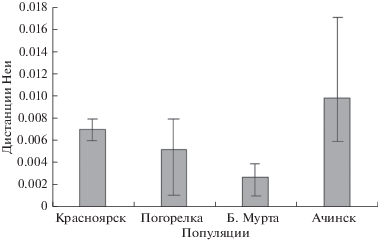
Как видно из табл. 2, наибольшие различия между участками, пройденными рубками, и контролем наблюдаются по среднему числу аллелей на локус (NA) и индексу разнообразия Шеннона (I). Оба этих показателя уменьшаются на вырубках интенсивностью выше 35%. Почти не отличался от контроля подрост после выборочной рубки умеренно-высокой интенсивности (35%) “Погорелка” и, в меньшей степени, после сплошной узколесосечной рубки “Сухая”. Необходимо отметить, что между взрослыми деревьями и подростом на всех сплошных широколесосечных вырубках по сравнению с контролем в 1.5–3 раза увеличились генетические дистанции Неи (Nei, 1972): между 4 популяциями они варьировали в пределах 0.002–0.007, внутри популяций между контролем и вырубкой, между взрослыми деревьями и подростом на вырубке – от 0.004 до 0.017 (рис. 1). При этом различия между вырубками и контролем в популяциях “Красноярск”, “Большая Мурта”, “Погорелка” были сопоставимы с различиями между популяциями внутри региона, а в популяции “Ачинск” соответствовали межрегиональному уровню: близкие по значениям генетические дистанции Неи были получены нами при сравнении выборок сосны обыкновенной из европейской части ареала с сибирскими популяциями (Тихонова и др., 2011), а также популяций из Средней Сибири с популяциями из Восточной Сибири и Монголии (Экарт и др., 2014). Отметим, что по мнению Ю.П. Алтухова (2004) видоспецифичное соотношение внутри- и межпопуляционной компонент генетической изменчивости должно быть одним из основных показателей в мониторинге популяционных систем, испытывающих антропогенные воздействия. По другим генетическим характеристикам заметное снижение уровня наблюдаемой гетерозиготности (НО) и повышение индекса фиксации Райта (F) отмечено только после сплошной широколесосечной вырубки в популяции “Ачинск” у оставшихся на вырубке взрослых деревьев и подроста.
Рис. 1.
Дистанции Неи (Nei, 1972) между выборками деревьев на вырубках и в контроле внутри популяций (столбцы – средние значения, линии – максимальные и минимальные значения).
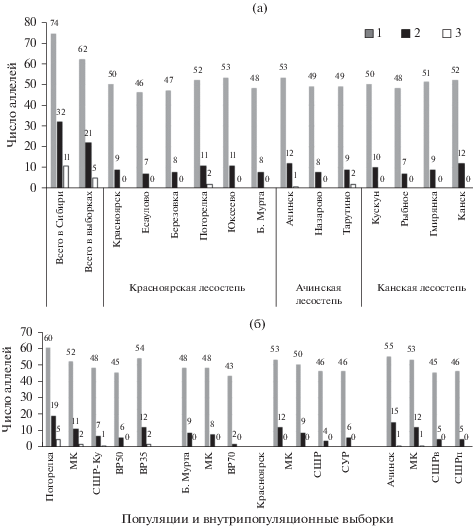
Число аллелей в 20 локусах в популяционных выборках варьировало от 48 до 60, число редких – от 8 до 19. У каждого дерева суммарное число аллелей было 21–28, редких – 0–2. Напомним, что исследуемые ферменты задействованы в адаптивных биохимических реакциях деревьев, а их аллели внутри локусов различаются по физико-химическим условиям функционирования ферментов, и следовательно, приспособительными особенностями (Животовский, 1984). Поэтому сокращение их разнообразия вполне определенно может привести к снижению устойчивости и продуктивности будущих поколений деревьев в меняющихся условиях внешней и внутренней среды лесных биогеоценозов.
Всего в исследуемых выборках было определено 62 аллеля, из них 21 редкий (рис. 2). За все время исследований, включая данную работу, в Сибири для аналогичного набора локусов было обнаружено 74 аллеля, из них 31 общий, 32 редких (в их числе 11 встречались только в отдельных популяциях). При выборе регионального контроля учитывали как степень нарушенности популяции, так и аллельное разнообразие всех выборок, включая выборки, отобранные на вырубках. Наибольшее разнообразие аллозимов и меньшая нарушенность части популяции были отмечены в Погорельском бору (доля полиморфных локусов Р = 0.85, суммарное число выявленных аллелей 60, включая 19 редких – это один из самых высоких показателей с учетом всех, исследованных нами ранее популяций вида в Сибири (Тихонова и др., 2011, 2014; Экарт и др., 2014). В других популяциях общее число аллелей было меньше (“Б. Мурта” – 48, “Красноярск” – 53, “Ачинск” – 55). Поэтому данная популяция была выбрана основным (региональным) контролем и ее можно предварительно рассматривать в качестве кандидата на генетический резерват. Отметим, что несмотря на ограниченную площадь, значительное генетическое разнообразие сосны сохраняется в городских парках гг. Ачинск (парк “Железнодорожников” площадью около 8 га) и Канск (с 1985 г. памятник природы “Сосновый бор в г. Канск” площадью 130 га), в припоселковых борах (сс. Юксеево, Гмирянка).
Рис. 2.
Число аллелей, обнаруженных на ненарушенных рубками участках (а) популяций сосны обыкновенной; всего в популяции (первый столбец) и у подроста с вырубок (б) на исследуемой территории, 1 – всего, 2 – редких, 3 – очень редких. Обозначения выборок см. в табл. 2.
Так как наибольший негативный эффект от рубок проявился в сокращении аллельного разнообразия аллозимов, основное внимание было уделено сравнительному анализу частот распределения редких аллелей исследуемых полиморфных локусов, присутствующих с частотой менее 5%: Mdh–283, Mdh–490, Got–1107, Got–332, 310, Skd-h–185, 90, 103, 107, Skdh–286, Lap–1103, Lap–298, 102, 105, Fdh45, 206, Pgm–193, 95, Adh–259, Fe–285, 113 (табл. 3). В данных популяциях не обнаружено следующих 10 аллелей, редко встречаемых в сибирской части ареала: Got–188, Got–2148, Got–3505, Skdh–193, 98, 6-Pgd–267, 91, Adh–1112, 116, Idh74.
Таблица 3.
Частота встречаемости редких аллелей (%) у подроста на вырубках и в контроле исследуемых популяций сосны обыкновенной
| Локус Аллель |
Погорелка | Б. Мурта | Красноярск | Ачинск | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| СШРКу | ВР50 | ВР35 | МК | ВР70 | МК | СУР | СШР | МК | СШРп | СШРв | МК | |
| Mdh–283 | – | – | 3.3 | – | – | – | – | – | – | – | – | – |
| Mdh–490 | – | 1.7 | 3.3 | – | – | 0.8 | – | – | 1.4 | 1.7 | – | 1.7 |
| Got–1107 | 1.7 | – | 1.7 | 1.8 | – | – | – | – | 1.4 | – | – | 1.7 |
| Got–332 | – | – | – | – | – | – | – | – | – | – | – | 3.4 |
| Got–3310 | 1.7 | 1.7 | 3.3 | 1.8 | 3,3 | 3.3 | 1.7 | 2.0 | 1.4 | 5.0 | 1.7 | 2.5 |
| Skdh–185 | 1.7 | 3.4 | 1.7 | 1.8 | – | 1.7 | – | – | – | – | – | 1.7 |
| Skdh–190 | – | – | 1.7 | – | – | – | – | – | 1.4 | – | 1.7 | – |
| Skdh–1103 | – | 1.7 | – | 2.6 | – | 3.3 | 1.7 | – | 2.9 | – | – | 1.7 |
| Skdh–1107 | 5.2 | 1.7 | 2.6 | – | – | 0.9 | – | – | – | – | – | 5.1 |
| Skdh–286 | – | – | – | – | – | – | 1.7 | – | 1.4 | – | – | – |
| Lap–1103 | – | – | – | 1.7 | – | – | – | – | – | – | – | – |
| Lap–298 | – | 1.7 | 1.7 | 1.8 | – | – | 3.3 | 2.0 | – | 4.7 | 3.3 | 1.7 |
| Lap–2102 | – | – | – | 1.7 | – | – | – | – | – | – | – | 0.9 |
| Lap–2105 | 1.7 | – | 1.7 | – | – | – | – | – | – | – | – | – |
| Fdh45 | 1.7 | – | 1.7 | – | 3.3 | 3.4 | – | – | 1.4 | 3.3 | 1.7 | – |
| Fdh206 | – | – | – | 1.8 | – | 1.7 | 1.7 | – | 1.4 | 1.7 | 3.3 | 1.7 |
| Pgm–193 | – | – | – | 1.7 | – | – | – | – | – | – | – | – |
| Pgm–195 | – | – | – | – | – | – | – | 4.0 | – | – | – | – |
| Adh–259 | 3.4 | – | 4.2 | 2.6 | – | 2.0 | 3.3 | – | 1.4 | – | – | 1.7 |
| Fe–285 | – | – | 1.7 | – | – | – | – | – | – | – | – | – |
| Fe–2113 | – | – | – | 2.7 | – | – | – | 2.0 | – | – | – | 0.9 |
| р | 0.121 | 0.033 | 0.202 | – | 0.034 | – | 0.160 | 0.047 | – | 0.032 | 0.032 | – |
| Всего | 7 | 6 | 12 | 11 | 2 | 8 | 6 | 4 | 9 | 5 | 5 | 12 |
Примечание. р – уровень достоверности различий с местным контролем. Обозначения выборок см. в табл. 2.
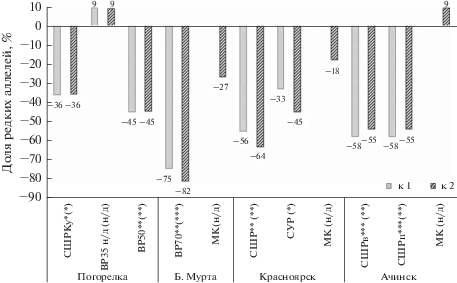
Предварительный сравнительный анализ всех 15 выборок по частотам аллелей всех локусов показал высоко достоверные различия между ними по точному критерию Фишера (р < 0.0000), а при полокусном анализе – по 8 локусам: Mdh–3, Got–2, Skdh–1, Lap–2, 6Pgd–2, Pgm–1, Adh–1, Adh–2. В ограниченном наборе 12 локусов, которые содержали редкие аллели, наиболее дифференцированы по частотам аллелей выборки с вырубок и контрольных участков в популяциях “Ачинск” (χ2 = 57.26, d. f. 22, р < 0.00006) и “Красноярск” (χ2 = 98.45, d. f. 22, р < 0.00000), в меньшей степени популяциях “Погорелка” (χ2 = 34.91, d. f. 24, р < 0.069) и “Б. Мурта” (χ2 = 17.25, d. f. 22, р < 0.750). Достоверные различия по частоте встречаемости редких аллелей у подроста на вырубках по сравнению с местным контролем выявлены на вырубках интенсивностью выше, чем “умеренно-высокая” 30–40% в популяциях “Погорелка”, “Б. Мурта”, “Красноярск” и “Ачинск”. Достоверно различались между собой также 2 варианта выборочной рубки интенсивностью 35 и 50% (“Погорелка”). Дополнительно сравнивали выборки по общему числу редких аллелей. В результате этого анализа подтвердилось достоверное сокращение генетического разнообразия в тех же вариантах рубок, а также в культурах после сплошной широколесосечной рубки в Погорельском бору по сравнению с контролем (рис. 3). Наряду с этим наиболее обедненная по разнообразию аллелей ферментов популяция “Б. Мурта” существенно отличалась от всех популяций при попарном сравнении по 3–8 (в общем наборе локусов) и по ограниченному набору 12 локусов (χ2 = 58.71–112.34, d. f. 32–34, р < 0.0027–0.00000). Несмотря на достоверные наибольшие различия между вырубкой и контролем по числу и частотам редких аллелей в этой популяции (табл. 3, рис. 3), между выборками внутри популяции получены самые низкие генетические дистанции Неи (рис. 1). Это свидетельствует о том, что отмеченное нами выше общее увеличение дистанции Неи на вырубках по сравнению с контролем не может служить самостоятельным критерием в оценке влияния лесопользования на генетическую изменчивость популяций и должно рассматриваться только вместе с данными о динамике и пространственной структуре аллельного и генотипического разнообразия популяций. Добавим также, что такие показатели генетической изменчивости, как снижение уровня гетерозиготности (НО) и повышение индекса фиксации Райта (F), по-видимому, недостаточно чувствительны к данному фактору, и поэтому тоже могут служить только в качестве дополнительных критериев наиболее сильных повреждающих воздействий на сосновые леса. В результате исследования насаждений сосны веймутовой также сделан вывод о непригодности величины НО для целей генетического мониторинга (Buchert et al., 1997; Rajora, 2000). В большей части работ, посвященных изучению данной проблемы, установлено сокращение числа аллелей, изменение генетической структуры популяций у разных древесных видов, в основном, после сплошных рубок (Savolainen, Kärkkäinen, 1992; Ledig, 1992; Buchert et al., 1997; Macdonald et al., 2001; Marquard et al., 2007; Ratnam et al., 2014), реже – в результате выборочных и постепенных (с большим воздействием) рубок (Buchert et al., 1997; Adams et al., 1998; Marquard et al., 2007). В отдельных работах аналогичные изменения зафиксированы при использовании искусственного лесовосстановления сосняков (Raja et al., 1998; Rajora, 1999). Вместе с тем, в ряде работ каких-либо существенных изменений в генофонде популяций после рубок и в лесных культурах выявлено не было (Nale, 1985; Ильинов и др., 2010; Ratnam et al., 2014) либо получены более высокие показатели в культурах по числу аллелей у потомства представительной выборки деревьев, так как уровень генетического разнообразия является функцией от числа деревьев, с которых были собраны семена (Adams et al., 1998).
Рис. 3.
Сокращение числа редких аллелей (в %) на вырубках по сравнению с местным (1) и региональным (2) контролем, *…*** уровни достоверности различий по критерию Фишера р < 0.05, 0.01, 0.001 относительно 1 (в скобках – относительно 2), н/д – недостоверно.
Таким образом проведенное нами исследование на вырубках в лесостепных борах Средней Сибири в целом подтверждает существенное влияние лесохозяйственной деятельности человека на генетическое разнообразие популяций сосны обыкновенной, даже после однократного приема рубок, не рассматривая проблему лесопользования и обеднения генофонда древесных видов в историческом аспекте. Оно согласуется с выводами других исследователей, установивших неполное воспроизводство генетического разнообразия в лесах после рубок вследствие фрагментации ареалов, сокращения численности популяций, искусственного восстановления и селекции (Ledig, 1992; Buchert et al., 1997; Adams et al., 1998; Тараканов и др., 2004; Видякин, 2007; Lise et al., 2007; Marquard et al., 2007; Падутов и др., 2008; Ortego et al., 2010 и др.). Разногласия в оценке влияния лесохозяйственной практики на генетическое разнообразие древесных растений, на наш взгляд, объясняется различиями между объектами исследования (биоэкологическими особенностями видов, уровнем обеднения/богатства и истории генофондов разных видов и их популяций); различиями в размерах популяций, площади вырубок, геометрической форме этой площади (квадрат – узкий прямоугольник), размеров окружающей рубку малонарушенной территории; возможное воздействие пожаров до и после рубки, наличие факторов, препятствующих восстановлению лесов либо способствующих ему – проведение рубки в годы с высоким урожаем семян, удаление напочвенного покрова и верхнего горизонта почвы. Большое значение имеют также различия между используемыми генетическими маркерами.
Следует отметить, что на вырубках могут сохраняться носители очень редких для популяции аллелей, поэтому после включения в анализ подроста с вырубок их число в популяционных выборках увеличилось в популяциях “Погорелка” с 11 до 19, “Красноярск” – с 9 до 12, “Ачинск” – с 12 до 15, в том числе за счет увеличения выборки (Тихонова и др., 2014). При этом на вырубках увеличиваются различия между поколениями взрослых деревьев и подроста, а также между ценопопуляциями (или субпопуляциями) в пространстве по частоте встречаемости одноименных редких аллелей, об этом свидетельствуют данные табл. 3, рис. 1 и результаты корреляционного анализа. Коэффициенты ранговой корреляции Спирмена (R) по частоте встречаемости одних и тех же редких аллелей в выборках взрослых деревьев и подроста существенно снижаются после сплошной широколесосечной рубки. Так, в Погорельском бору между парами выборок подроста с выборочной вырубки интенсивностью 35%, выборочной рубки интенсивностью 50% и в культурах после сплошной широколесосечной рубки с контрольным участком R, соответственно, составил 0.73, 0.62, и 0.56; в популяции “Красноярск” в парах выборок со сплошной узколесосечной и широколесосечной рубки с контролем, соответственно, R = 0.70 и 0.59; в популяции “Ачинск”, соответственно, R = 0.66 и 0.48 в парах подроста и взрослых деревьев со сплошной широколесосечной рубки и контроля (все значения R достоверны при р < 0.05–0.01).
Лучшая сохранность и даже более высокие показатели генетического разнообразия по сравнению с контролем и низкие дистанции Неи с местным контролем наблюдались на участке с выборочной рубкой интенсивностью 35% в Погорельском бору (рис. 2б, рис. 3, табл. 3). Поскольку эта рубка была проведена через 10 лет после низового пожара, который вызвал вспышку возобновления на участке, на наш взгляд, целесообразно рассмотреть возможность внедрения в практику лесного хозяйства предварительного контролируемого выжигания участков сосняков, планируемых к выборочной или узколесосечной рубке, в пожаробезопасный период в некоторых типах леса и лесорастительных условий лесостепи. Их стимулирующее влияние на возобновление сосны неоднократно отмечалось исследователями (Колесников и др., 1973; Фуряев, 1996), в том числе с использованием контролируемых неинтенсивных выжиганий (Ковалева и др., 2018). В одной из работ, посвященных исследованию старовозрастных смешанных хвойных древостоев с участием сосен Жеффрея и Ламберта в Калифорнии также отмечается отсутствие признаков негативного влияния рубок в древостоях послепожарного происхождения (Macdonald et al., 2001). Однако в целом лесостепные боры относятся к группе лесов с высокой пожарной опасностью: после рубки и пожаров леса здесь не всегда хорошо восстанавливаются, особенно в засушливые годы (Полувековой …, 1966; Фуряев, 1996; Ильичев и др., 2003; Буряк и др., 2011; Куулар и др., 2015; Иванова и др., 2015). Высока опасность перехода низового пожара в верховой, после которого значительно сокращаются все показатели генетической изменчивости аллозимов, что наблюдалось нами в послепожарном сосняке на Куртушибинском хр. Западного Саяна (Экарт и др., 2014). Поэтому для лесостепных боров, учитывая их ценность и связанные с пожарами риски, возможно, больше подойдет не выжигание (или даже не сплошное выжигание, – мозаичное), а предварительная небольшая минерализация поверхности почвы. Вторым вариантом, характеризующимся меньшим обедняющим эффектом на генофонд популяций сосны обыкновенной в условиях лесостепи, является чересполосная узколесосечная рубка, которую рекомендуют для лесостепных боров Западной Сибири (Ильичев и др., 2003).
В данной работе ввиду ограниченного набора популяций и небольшой территории мы не затронули такое важное условие рационального лесопользования, как необходимость сохранения пространственной популяционной структуры древесных видов, на что обращают внимание многие отечественные исследователи (Мамаев и др., 1988; Ирошников и др., 1989; Гончаренко и др., 1996; Милютин, 2003; Динамика …, 2004; Видякин, 2007; Тараканов, 2009).
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования было установлено, что генетическая структура и разнообразие аллозимов в лесостепных популяциях сосны обыкновенной в Средней Сибири сохраняются на участке после первого приема выборочной рубки невысокой интенсивности, проведенной методом узких лент после низового пожара, что в совокупности стимулировало процесс возобновления и появления большого количества подроста. На втором месте, с меньшим сокращением числа редких аллелей по сравнению с остальными вариантами, была узколесосечная рубка. В сравнении с ними при рубках более высокой интенсивности (50%, 70% и сплошной массивной рубке) число редких аллелей у подроста сокращалось на 45–82% относительно контроля, что равносильно сокращению общего числа аллелей у подроста на 8–15% относительно местного и на 13–28% – относительно регионального контроля за один прием рубки. Для популяций с большой площадью при небольшой площади рубок это разнообразие может быстро восстановиться, однако в условиях лесостепи возможен постепенно накапливаемый эффект обеднения генофонда популяций в связи с их изолированным положением и сокращением притока генов извне, незаметный для одного поколения исследователей. Наблюдаемое нами в наиболее нарушенных рубками небольших популяциях сокращение аллельного разнообразия, низкий уровень гетерозиготности особей и высокие значения коэффициента инбридинга (F) могут привести к снижению продуктивности и устойчивости новых поколений деревьев.
Ввиду того, что в Погорельском бору было выявлено наибольшее среди всех исследованных нами ранее популяций сосны обыкновенной число редких аллелей, на части его территории необходимо выделить генетический резерват (надежнее 2–3 в разных участках бора). Подобные генетические резерваты желательно выделять во всех лесостепных борах, оставленных в окрестностях крупных населенных пунктов лесостепей на расстоянии не более 100–150 км друг от друга на основе существующих естественных изолированных популяций сосны обыкновенной и, возможно, включая искусственно созданные микро-культуры (или одиночные деревья) для восстановления утраченной генетической изменчивости наиболее нарушенных из них.
Исследования показали, что наряду с такими составляющими лесохозяйственного мониторинга, как лесопожарный и лесопатологический, необходимо организовать дополняющий генетический мониторинг популяций основных лесообразующих древесных видов. В качестве некоторых методических рекомендаций можно отметить, что для этих целей желательно увеличение числа особей в популяционных выборках (до ≥120–150). Для целей мониторинга наиболее информативными являются показатели NA, I, число и частота встречаемости редких и низкочастотных аллелей, в качестве дополнительных характеристик могут быть использованы дистанции Неи (Nei, 1972), НО, F и Nm. Необходимо учесть, что на результаты оценок генетической изменчивости у подроста после рубок определенное влияние оказывают различия между исходными популяциями, поэтому желательно использовать не менее 2-х контролей и разработать поправочные коэффициенты к показателям степени обеднения генофонда популяций, учитывающие степень их изолированности, эффективную численность, уровень генетического разнообразия и нарушенность. Отметим также, что преобладание в лесостепях Средней Сибири вторичных березняков является объективным критерием нерационального лесопользования и необходимости не только развития плантационного лесоводства на землях сельскохозяйственного назначения (в настоящее время крестьян повсеместно заставляют вырубать подрост на заросших лесом участках), но и увеличения доли хвойных пород в естественных лесах этой зоны.
Список литературы
Алтухов Ю.П. Динамика генофондов при антропогенных воздействиях // Вестник ВОГиС. 2004. Т. 8. № 2. С. 40–59.
Антипова Е.М. Флора внутриконтинентальных островных лесостепей Средней Сибири. Красноярск: Красноярский гос. пед. университет им. В.П. Астафьева, 2012. 662 с.
Берг Л.С. Географические зоны Советского Союза. М.: Изд-во ОГИЗ, 1947. 397 с.
Бузыкин А.И., Пшеничникова Л.С. Ресурсно-экологический потенциал лесов Красноярского края // Хвойные бореальной зоны. 2008. Т. 25. № 3–4. С. 327–332.
Буряк Л.В., Сухинин А.И., Каленская О.П., Пономарев Е.И. Последствия пожаров в ленточных борах юга Сибири // Сибирский экологический журн. 2011. № 3. С. 135–140.
Видякин А.И. Фенетика, популяционная структура и сохранение генетического фонда сосны обыкновенной (Pinus sylvestris L.) // Хвойные бореальной зоны. 2007. № 2–3. С. 159–166.
Гончаренко Г.Г., Дробышевская В.В., Силин А.Е., Падутов В.Е. Генетические ресурсы сосен России и сопредельных государств // Доклады РАН. 1996. Т. 346. № 3. С. 419–423.
Динамика популяционных генофондов при антропогенных воздействиях / Под ред. Алтухова Ю.П. М.: Наука, 2004. 618 с.
Животовский Л.А. Интеграция полигенных систем в популяциях. М.: Наука, 1984. 183 с.
Иванова Г.А., Иванов В.А., Кукавская Е.А. Периодичность пожаров в лесах республики Тыва // Хвойные бореальной зоны. 2015. Т. 33. № 5–6. С. 204–209.
Ильинов А.А., Политов Д.В., Раевский Б.В. Влияние способов лесовозобновления на генетическую структуру популяций ели финской Picea x fennica (Regel) Kom // Ученые записки Петрозаводского государственного университета. 2010. Т. 109. № 4. С. 50–55.
Ильичев Ю.Н., Бушков Н.Т., Тараканов В.В. Естественное лесовосстановление на гарях Среднеобских боров. Новосибирск: Изд-во СО РАН, 2003. 195 с.
Ирошников А.И., Мамаев С.А., Некрасов В.И. Генетический фонд лесных древесных пород в СССР (дифференциация, использование, охрана, мониторинг). В кн.: Материалы межд. Симпозиума “Лесная генетика, селекция и физиология древесных растений” (25–29 сентября 1989 г., Воронеж). М.: ЦНИИЛГиС РАН, 1989. С. 9–16.
Калашников Е.Н. Мониторинг нарушенности лесов Сибири // Сибирский экологический журн. 1998. № 1. С. 49–57.
Ковалева Н.М., Собачкин Р.С., Екимова Е.Ю. Динамика нижних ярусов растительности после экспериментальных пожаров в сосновых древостоях // Сибирский лесной журн. 2018. № 2. С. 61–70.
Колесников Б.П., Санникова Н.С., Санников С.Н. Влияние низового пожара на структуру древостоя и возобновление древесных пород в сосняках черничном и бруснично–черничном // Горение и пожары в лесу. Красноярск: Изд-во Института леса и древесины им. В.Н. Сукачева СО АН ССР, 1973. С. 301–321.
Куулар Х.Б., Намзын Ш.А., Хертек С.Б. Мониторинг Балгазынского бора по данным дистанционного зондирования // Известия высших учебных заведений. Геодезия и аэрофотосъемка. 2015. № 3. С. 67–72.
Леса СССР: Леса Урала, Сибири и Дальнего Востока / Под ред. Жуков А.Б. М.: Наука, 1969. Т. 4. С. 318.
Мамаев С.А., Семериков Л.Ф., Махнёв А.К. О популяционном подходе в лесоводстве // Лесоведение. 1988. № 1. С. 3–9.
Милютин Л.И. Генетико-эволюционные основы устойчивости лесных экосистем // Лесоведение. 2003. № 1. С. 16–20.
Падутов В.Е., Хотылева Л.В., Баранов О.Ю., Ивановская С.И. Генетические эффекты трансформации лесных экосистем // Экологическая генетика. 2008. Т. VI. № 1. С. 3–11.
Побединский А.В. Рубки и возобновление в таёжных лесах СССР. М.: Лесная пром-ть, 1973. 200 с.
Полувековой опыт лесовосстановления в ленточных борах Казахстана и Алтая / Под общ. ред. Грибанова Л.Н. Алма-Ата: Кайнар, 1966. 131 с.
Савина Л.Н. Таежные леса Северной Азии в голоцене. Новосибирск: Наука, 1986. 190 с.
Санников С.Н., Петрова И.В., Семериков В.Л. Генофеногеографический анализ популяций Pinus sylvestris L. на трансекте от северной до южной границы ареала // Экология. 2002. № 2. С. 97–102.
Семериков В.Л., Семерикова С.А., Дымшакова О.С., Зацепина К.Г., Тараканов В.В., Тихонова И.В., Экарт А.К., Видякин А.И., Жамьянсурен С., Роговцев Р.В., Кальченко Л.И. Полиморфизм микросателлитных локусов хлоропластной ДНК сосны обыкновенной (Pinus sylvestris L.) в Азии и Восточной Европе // Генетика. 2014. Т. 50. № 6. С. 660–669.
Синицких П. Краткий обзор состояния рынков продаж хвойных пиломатериалов // Леспроминформ. 2015. № 2. С 90–91.
Солодухин Е.Д. К типологии дальневосточных вырубок // Лесной журн. (Известия высших учебных заведений). 1961. № 4. С. 22–23.
Сочава В.Б. Географические аспекты сибирской тайги. Новосибирск: Наука, 1980. 256 с.
Степанов Н.В. Сосудистые растения Приенисейских Саян: флористический и биоресурсный анализ: Автореф. дис. … д-ра биол. наук (спец. 03.02.14). Красноярск: Сибирский федеральный университет, 2014. 40 с.
Тараканов В.В. Достижения и ошибки в области сохранения и рационального использования лесных генетических ресурсов Сибири // Лесное хоз-во. 2009. № 5. С. 10–12.
Тараканов В.В., Самсонова А.Е., Ильичев Ю.Н. Влияние естественных и антропогенных факторов на генетическую изменчивость сосны в Приобье: состав терпентинных масел хвои // Лесоведение. 2004. № 5. С. 50–57.
Тихонова И.В., Семериков В.Л., Семерикова С.А., Дымшакова О.В., Полежаева М.А., Экарт А.К. О выборках в исследованиях внутривидового генетического разнообразия хвойных // Сибирский лесной журн. 2014. № 4. С. 99–109.
Тихонова И.В., Семериков В.Л., Шишикин А.С., Тараканов В.В. О необходимости особого режима хозяйствования и охраны в рефугиумных (реликтовых) популяциях видов хвойных в Сибири // Лесное хозяйство. 2011. № 3. С. 41–42.
Фуряев В.В. Роль пожаров в процессе лесообразования. Новосибирск: Наука, 1996. 253 с.
Хедрик Ф. Генетика популяций. М.: Техносфера, 2003. 592 с.
Экарт А.К., Ларионова А.Я., Зацепина К.Г., Кравченко А.Н., Жамьянсурэн С., Тихонова И.В., Тараканов В.В. Генетическое разнообразие и дифференциация сосны обыкновенной в Южной Сибири и Монголии // Сибирский экологический журн. 2014. № 1. С. 69–78.
Adams W.T., Zuo J., Shimizu J.Y., Tappeiner J.C. Impact of alternative regeneration methods on genetic diversity in coastal Douglas-fir // Forest Science. 1998. V. 44. P. 390–396.
Buchert G.P., Rajora O.P., Hood J.V., Dancik B.P. Effects of harvesting on genetic diversity in old-growth eastern white pine (Pinus strobes L.) in Ontario, Canada // Conservation Biology. 1997. № 11. P. 747–758.
Fussi B., Westergren M., Aravanopoulos F., Baier R., Kavaliauskas D., Finzgar D., Alizoti P., Bozic G., Avramidou E., Konnert M., Kraigher H. Forest genetic monitoring: an overview of concepts and definitions // Environmental Monitoring & Assessment. 2016. V. 188. № 493. 12 pp. https://doi.org/10.1007/s10661-016-5489-7
Ledig F. Human impacts on genetic diversity in forest ecosystems // Oikos. 1992. V. 63. P. 87–108.
Lise Y., Kaya Z., Isik F., Sabuncu R., Kandemir I., Onde S. The impact of overexploitation on the genetic structure of Turkish red pine (Pinus brutia Ten.) populations determined by RAPD markers // Silva Fennica. 2007. V. 41. P. 211–220.
Macdonald S.E., Thomas B.R., Cherniawsky D.M., Purdy B.G. Managing genetic resources of lodgepole pine in west-central Alberta: patterns of isozyme variation in natural populations and effects of forest management // Forest Ecology & Management. 2001. V. 152. P. 45–58.
Marquard P.E., Echt C.S., Epperson B.K., Pubanz D.M. Genetic structure, diversity, and inbreeding of eastern white pine under different management conditions // Canadian J. Forest Research. 2007. V. 37. P. 2652–2662.
Nei M. Genetic distance between populations // American Naturalist. 1972. V. 106. P. 283–292.
Nale D.B. Genetic implications of shelterwood regeneration of Douglas-fir in southwest Oregon // Forest Science. 1985. V. 31. № 4. P. 995–1005.
Ortego J., Bonal R.L., Munoz A. Genetic consequences of habitat fragmentation in long-lived tree species: the case of the Mediterranean Holm Oak (Quercus ilex L.) // Heredity. 2010. V. 101. P. 717–726.
Peakall R., Smouse P.E. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research // Molecular Ecology Notes. 2006. № 6. P. 288–295.
Raja R.G., Tauer C.G., Wittwer R.F., Huang Y. Regeneration methods affect genetic variation and structure in shortleaf pine (Pinus echinata Mill.) // Forest Genetics. 1998. № 5. P. 171–178.
Rajora O.P. Genetic biodiversity impacts of silvicultural practices and phenotypic selection in white spruce // Theoretical and Applied Genetics. Genet. 1999. V. 99. P. 954–961.
Ratnam W., Rajora Om P., Finkeldey R., Aravanopoulos F., Bouvet J-M., Vaillancourt R. E., Kanashiro M., Fady B., Tomita M., Vinson Ch. Genetic effects of forest management practices: Global synthesis and perspectives // Forest Ecology & Management. 2014. 14 p. https://doi.org/10.1016/j.foreco.2014.06.008
Raymond M., Rousset F. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism // Heredity. 1995. V. 86. P. 248–249.
Savolainen O., Kärkkäinen K. Effect of forest management on gene pools // New Forests. 1992. V. 6. P. 372–383.
Wright S. Evolution and the genetics of populations. V. 4. Variability within and among natural populations. Chicago: University of Chicago, 1978. 590 p.
Дополнительные материалы отсутствуют.