

Лесоведение, 2021, № 4, стр. 437-445
Ферментативная активность почвы под насаждениями клена ясенелистного в условиях пойменных лесных биогеоценозов
О. Л. Цандекова a, *, В. И. Уфимцев a
a ФГБНУ Федеральный исследовательский центр угля и углехимии СО РАН
650065 Кемерово, Ленинградский просп., 10, Россия
* E-mail: zandekova@bk.ru
Поступила в редакцию 15.04.2020
После доработки 23.09.2020
Принята к публикации 06.04.2021
Аннотация
Изучена активность почвенных ферментов под насаждениями клена ясенелистного, произрастающего в пойме р. Томь в пределах г. Кемерово. Отбор почвенных проб проводили по трем категориям сомкнутости крон деревьев. В качестве контроля выбрана внешняя зона одиночных деревьев. В смешанных образцах почвы, отобранных из горизонта 0–10 см в начале, середине и конце вегетационного сезона (2017–2019 гг.), определяли активность гидролитических ферментов (протеазы, фосфатазы и инвертазы). Выявлено, что наибольший уровень активности гидролитических почвенных ферментов отмечен возле одиночных деревьев A. negundo в несомкнутых древостоях. Наименьшая сезонная вариабельность характерна для активности протеазы и фосфатазы. У одиночных деревьев в несомкнутых древостоях коэффициент вариации ферментативной активности ниже в 1.5 раза, чем у других исследуемых образцов. Наиболее тесная отрицательная связь обнаружена между активностью почвенных ферментов под насаждениями A. negundo и содержанием гумуса, общего и нитратного азота. Полученные данные по ферментативной активности могут служить одним из показателей биологической активности почвы под древесными насаждениями, быть использованы при биодиагностике, биомониторинге и биоиндикации состояния почв в природных экосистемах.
Ферментативная активность почв является одним из важнейших показателей, характеризующих их биологическое состояние. Почвенные ферменты играют ведущую роль в биохимических процессах, происходящих в почве, и являются достаточно устойчивыми и чувствительными показателями при почвенной диагностике (Звягинцев, 1991; Хазиев, 2015). Являясь катализаторами химических реакций, они регулируют биохимические процессы, принимают участие в синтезе и распаде гумуса, гидролизе органических соединений, а также способствуют переводу элементов питания в формы, легкодоступные для растений. Ферменты, относящиеся к классу гидролаз (протеаза, фосфатаза, инвертаза), участвуют в основных процессах минерализации органических веществ и катализируют реакции гидролитического расщепления высокомолекулярных органических соединений. Протеазы участвуют в начальных этапах минерализации белковых соединений и обусловливают динамику усвояемых форм азота. Фосфатазы катализируют гидролиз фосфорорганических веществ и регулируют интенсивность процессов мобилизации органического фосфора почвы. Активность фосфатазы тем выше, чем меньше в почве подвижных форм фосфора (Хазиев, 2005). Инвертаза участвует в биохимических превращениях углеводов, которые содержатся в почвенном органическом веществе, микроорганизмах и растениях в значительном количестве. Исследования некоторых авторов показали, что активность инвертазы лучше других ферментов отражает уровень плодородия и биологической активности почв (Казеев и др., 2003; Даденко и др., 2013а). Более высокая ферментативная активность характерна для верхних горизонтов почвы (0–10 см) по сравнению с более глубокими (20–30 см) (Сакбаева, 2014; Li et al., 2018; Yu et al., 2019). В течение вегетации активность почвенных ферментов повышается в период активного роста растений, а также при разложении корневых и растительных остатков (Хежева и др., 2010; Hu et al., 2016; Moghimian et al., 2017).
Конкурентные взаимоотношения растительных сообществ в условиях природных ландшафтов ведут к усилению деятельности почвенных ферментных систем микроорганизмов и корневой системы растений (Гродзинский, 1965; Кавеленова и др., 2008; Saccone et al., 2010; Porté et al., 2011). Под влиянием насаждений клена ясенелистного в его подкроновом пространстве ферментативная активность почвы может претерпевать некоторые изменения, что приводит к сокращению площади напочвенного покрова, подавлению роста самосева и подроста растений, и препятствует их естественному возобновлению в природных сообществах (Ерёменко, 2014; Коношина и др., 2014; Веселкин и др., 2019; Лозбякова, Степанов, 2019). В настоящее время недостаточное внимание уделено особенностям изменения ферментативной активности почв под влиянием растительности, в частности в насаждениях A. negundo. На территории Кемеровской области данные исследования не проводились. В связи с этим, актуален поиск наиболее информативных диагностических показателей состояния почвы и ее ферментативной активности, что позволит расширить сведения об активности почвенных ферментов в конкретных почвенно-климатических условиях.
Цель работы – оценить ферментативную активность почвы под насаждениями A. negundo в условиях пойменных лесных биогеоценозов.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в насаждениях Acer negundo L. (клен ясенелистный) в пойме р. Томь в пределах г. Кемерово. Исследуемые насаждения оцениваются первой категорией жизненного состояния по шкале В.А. Алексеева, I классом бонитета, возраст деревьев – 25–30 лет, высота – 12–14 м. У одиночных деревьев диаметр кроны составлял 8–10 м, у деревьев средней и полной сомкнутости крон – 4–6 м. Живой напочвенный покров образован разнотравно-злаковым сообществом с преобладанием Urtica dioica L., Poa pratensis L., Phleum pratense L., Elytrigia repens (L.) Nevski., Humulus lupulus L., с общим проективным покрытием 20–90%. Почвы района исследований характерны для пойменных ландшафтов Кузнецкой котловины (Трофимов, 1975; Самаров, 2017), классифицируются как лугово-черноземные по эколого-генетической классификации почв СССР (Классификация …, 1977), или как черноземы гидрометаморфизованные по профильно-генетической классификации почв России (Полевой определитель …, 2008), или как Gleyic Chernozems по классификации WRB (IUSS …, 2014).
Отбор почвенных проб проводили в 2017–2019 гг. в начале (III декада мая), середине (III декада июля) и конце (III декада сентября) вегетационного периода. Почвенные пробы отбирали по категориям сомкнутости крон клена ясенелистного на учетных площадках: 1 – одиночные деревья в несомкнутых древостоях; 2 – деревья со средней (50–60%) сомкнутостью крон, 3 – деревья с высокой (100%) сомкнутостью крон. В качестве контроля (4) выбрана внешняя зона одиночных деревьев. Образцы почвы отбирали с каждого исследуемого варианта с глубины 0–10 см, поскольку основная биологическая активность и наибольшая биогенность присущи верхним горизонтам почвенного профиля (Хазиев, 2005).
Определение ферментативной активности почв проводили на свежесобранном смешанном образце в трехкратной повторности. Активность инвертазы определяли по методу В.Ф. Купревича и Т.А. Щербаковой, активность протеазы – по методу А.Ш. Галстяна и Э.А. Арутюнян (Практикум …, 2001), активность фосфатазы – по методу А.Ш. Галстяна (Титова, Козлов, 2012). Агрохимический анализ почвы на различных участках фитогенного поля в насаждениях клена ясенелистного проводили в аккредитованном испытательном центре агрохимической службы ФГУ ЦАС “Кемеровский”. Определяли реакцию почвенного раствора (рHсол) (ГОСТ 26483-85), содержание гумуса (ГОСТ 26213-91), подвижные формы фосфора (P2O5) и калия (K2O) (ГОСТ 26204-91), сумму поглощенных оснований (ГОСТ 27821-88), массовую долю нитратного (ГОСТ 26951-86) и общего азота (ГОСТ 26107-84). Статистическая обработка полученных данных и построение графиков выполнены с помощью стандартного пакета программ StatSoft STATISTICA 8.0. for Windows и Microsoft Office Excel 2007.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты агрохимических анализов показали, что исследуемые почвы характеризовались слабокислой и нейтральной реакцией солевой вытяжки (рН 5.9–6.5). Обеспеченность подвижными формами фосфора характеризуется как очень высокая и варьирует в пределах 610–1650 мг кг–1, что в 2.4–6.6 раз превышает нижний предел данной градации обеспеченности (табл. 1). Некоторые исследователи отмечают, что повышенные концентрации фосфора в почве могут блокировать поступление в растения важнейших элементов питания и, как следствие этого, вызвать приостановку их роста (Адрианов, Сушеница, 2004; Balemil, Negisho, 2012; Ratliff, Fisk, 2016), что является опосредованной характеристикой аллелопатического воздействия A. negundo.
Таблица 1.
Агрохимические показатели почвы на исследуемых площадках
| Показатель | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| рH(сол.) | 6.5 ± 0.1* | 6.1 ± 0.1 | 6.2 ± 0.1 | 5.9 ± 0.1 |
| Гумус, % | 7.6 ± 0.8 | 7.9 ± 0.8 | 10.6 ± 1.01 | 5.6 ± 0.6 |
| P2O5, мг кг–1 | 1170 ± 140 | 610 ± 73 | 1650 ± 198 | 620 ± 74 |
| K2O, мг кг–1 | 290 ± 29 | 191 ± 19 | 400 ± 40 | 140 ± 14 |
| Nнитратный, мг кг–1 | 26.3 ± 5.3 | 27.5 ± 5.5 | 58.9 ± 11.8 | 30.9 ± 6.2 |
| Nобщий, % | 0.39 ± 0.03 | 0.35 ± 0.02 | 0.53 ± 0.03 | 0.30 ± 0.02 |
| Сумма поглощенных оснований, ммоль × 100 г–1 | 42.4 ± 6.4 | 34.0 ± 5.1 | 43.2 ± 6.5 | 23.2 ± 3.5 |
Содержание нитратного азота по шкале Г.П. Гамзикова (1981) на пробных площадках колеблется на уровне высоких значений – от 26.3 до 58.9 мг кг–1. Массовая доля общего азота в исследуемых почвах составила 0.30–0.53%, сумма поглощенных оснований – 23.2–43.2 ммоль × 100–1 г. Содержание калия в почвах варьирует в пределах 140…400 мг·кг–1 и соответствует среднему и высокому уровню (ГОСТ Р 54650-2011), причем, ниже всего в контроле. Содержание гумуса в почвах составляет 5.6–10.6%, что соответствует уровню высокогумусных луговых почв.
Сравнительный анализ агрохимических показателей почвы под насаждениями A. negundo показал, что более высокие показатели изучаемых параметров (подвижный фосфор и калий, азот нитратный и общий, гумус) характерны для почв пробных площадок под деревьями с высокой сомкнутостью крон, в сравнении с контролем и другими участками. Содержание азота в почве подтверждает зависимость между его количеством и органическим веществом. Со снижением содержания гумуса в почве следует снижение содержания азота, и, наоборот, с увеличением содержания гумуса увеличивается содержание азота. Так, для почв контрольного участка характерен минимальный уровень содержания органического вещества – 5.6%, а для почв под насаждениями клена на третьей учетной площадке – максимальный уровень (10.6%), при этом массовая доля общего азота составила 0.30 и 0.53% соответственно.
Анализ экспериментальных данных по ферментативной активности почвы показал, что более высокие показатели на исследуемых участках характерны для вегетационных периодов 2018–2019 гг., в сравнении с 2017 г. Очевидно, сезонные изменения активности ферментов связаны с различными гидротермическими условиями данных периодов. Климатические условия вегетационного периода 2017 г. характеризовались достаточно теплой и влажной погодой. В мае среднемесячная температура воздуха составляла 11.7°С, что превышало среднемноголетнюю норму на 0.5°С. Осадков выпало 24 мм (60% от нормы). В июле и августе отмечались понижение температуры (на 0.5–0.8°С ниже нормы) и избыток увлажнения (144–146% от нормы). Особенностью вегетации 2018 и 2019 гг. являлось понижение температуры воздуха (ниже нормы на 1–1.1°С) и увеличение влажности (153–198% от нормы) в начале и в середине вегетации, к концу вегетации среднемесячная температура превысила норму на 1.1–1.7°С, осадков выпало 51–70 мм (103–124% от нормы). Некоторые исследователи отмечают, что в активные фазы роста растений при высокой температуре почвы и достаточной влажности в летние месяцы активность почвенных ферментов максимальна (Даденко и др., 2013б; Фаизова, 2014; Хазиев, 2014). Наши исследования подтвердили данную закономерность. Выявлено, что наибольшая активность ферментов у контрольных и опытных образцов отмечена в период активного роста деревьев (июль), к концу вегетации установлено их понижение. Сравнительный анализ данных по активности гидролитических ферментов почвы под насаждениями A. negundo выявил некоторые различия у исследуемых образцов на площадках наблюдений. Активность инвертазы во всех почвенных образцах выше, чем активность протеазы и фосфатазы.
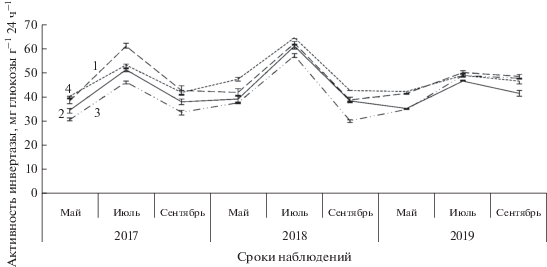
Анализ экспериментальных данных показал, что в течение вегетации во все сроки наблюдений активность инвертазы отличалась значительным варьированием по отношению к контролю (рис. 1). Так, у исследуемых почвенных образцов ферментативная активность варьировала в пределах от 30.20 до 56.93 мг глюкозы г–1 × 24 ч–1, что ниже на 2–23% относительно контроля. Более высокие показатели данного показателя отмечены в 2018 г., особенно в начале и середине вегетации в сравнении с другими годами исследований. Так, активность фермента в мае составила 37.5–47.21 мг глюкозы г–1 × 24 ч–1, в июле – 56.93–64.21 мг глюкозы г–1 × 24 ч–1. Наибольшие отличия от контроля (11–21%) характерны для деревьев с высокой сомкнутостью крон. Согласно шкале сравнительной оценки биологической активности почвы, предложенной Э.И. Гапонюк и С.Г. Малаховым (Казеев и др., 2003), в мае и сентябре степень активности фермента характеризовалась как средняя, в июле – относительно высокая на всех исследуемых площадках.
Рис. 1.
Сезонная динамика активности инвертазы на исследуемых площадках. На рис. 1–3: 1 – одиночные деревья в несомкнутых древостоях; 2 – деревья со средней (50–60%) сомкнутостью крон; 3 – деревья с высокой (100%) сомкнутостью крон; 4 – контроль (внешняя зона одиночных деревьев).
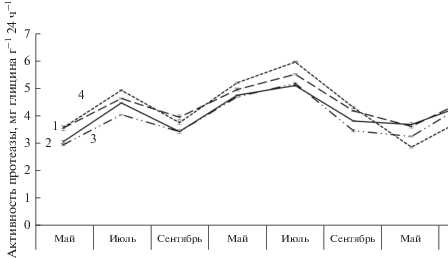
В течение вегетации у контрольных и опытных почвенных образцов на учетных площадках активность протеазы варьировала в пределах от 2.88 до 5.98 мг глицина г–1 × 24 ч–1 (рис. 2). Сравнительная характеристика исследуемых площадок выявила некоторые различия данного показателя в течение вегетации относительно контроля. Опытные образцы, собранные под одиночными деревьями в несомкнутых травостоях, характеризовались более высокими показателями протеолитической активности (3.57–5.53 мг глицина г–1 ×24 ч–1) в сравнении с другими исследуемыми участками. В вегетационный период 2019 г. значения по активности протеазы у этой категории деревьев превысили контроль на 12–26%. По анализу результатов исследований протеолитической активности установлено, что почва относится к средней степени активности.
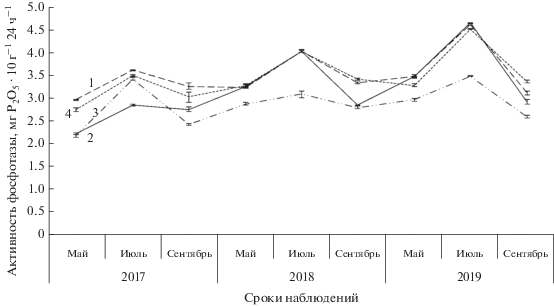
Анализ данных по активности фосфатазы показал, что в течение вегетации она варьировала в пределах от 2.18 до 4.65 мг P2O5 × 10 г–1 ч–1. Это соответствовало средней степени активности почв. Более высокие показатели фосфатазной активности почвы на исследуемых участках отмечены в вегетационный период 2019 г. Так, в мае и июле активность фермента у деревьев в несомкнутых травостоях и со средней сомкнутостью крон выше в среднем на 4–5%, с высокой сомкнутостью крон – ниже на 17% относительно контроля. В сентябре все опытные образцы уступили контрольным значениям на 7–23% (рис. 3).
Рис. 3.
Динамика активности фосфатазы в течение вегетации на исследуемых площадках. Обозначения см. рис. 1.
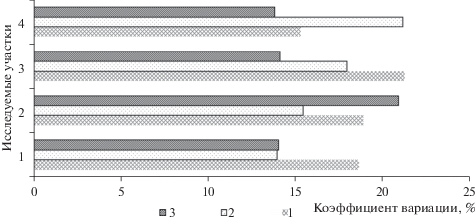
Анализ значений коэффициентов вариации почвенных ферментов выявил большую однородность активности протеазы и фосфатазы по сравнению с активностью инвертазы (рис. 4). Вариабельность активности протеазы и фосфатазы отличалась в почвах различных площадок наблюдений. Так, у одиночных деревьев в несомкнутых древостоях коэффициент вариации составил 13.97 и 14.06%, что меньше в 1–1.5 раза в сравнении с другими исследуемыми площадками. В результате анализа ферментативной активности почв было выявлено, что наиболее пригодными биоиндикационными признаками для почвенной оценки являются активность протеазы и фосфатазы.
Рис. 4.
Коэффициент вариации ферментативной активности почвы на исследуемых участках. Ферменты: 1 – фосфатаза; 2 – протеаза; 3 – инвертаза.
На основе полученных экспериментальных данных проведен расчет корреляционных связей между агрохимическими свойствами почв под насаждениями клена ясенелистного на исследуемых площадках и активностью почвенных ферментов. Наиболее тесная отрицательная связь обнаружена между активностью почвенных ферментов, особенно инвертазы и фосфазаты, и содержанием гумуса (r = –0.42…–0.44), нитратного и общего азота (r = –0.31…–0.39, при p < 0.05, N = 189).
ЗАКЛЮЧЕНИЕ
Под насаждениями клена ясенелистного в условиях пойменных лесных биогеоценозов выявлен средний уровень активности гидролитических почвенных ферментов. Наибольший уровень ферментативной активности выявлен возле одиночных деревьев A. negundo в несомкнутых древостоях по сравнению с другими группами деревьев и контролем. Почвенные образцы характеризовались наименьшей вариабельностью по активности протеазы и фосфатазы в сравнении с активностью инвертазы. У одиночных деревьев в несомкнутых древостоях коэффициент вариации ниже в 1.5 раза в сравнении с другими исследуемыми площадками. Наиболее тесная отрицательная связь обнаружена между активностью почвенных ферментов под насаждениями A. negundo и содержанием гумуса, общего и нитратного азота. Полученные данные можно использовать в качестве диагностических признаков состояния почвы, при биодиагностике, биомониторинге и биоиндикации почв в природных экосистемах.
Список литературы
Адрианов С.Н., Сушеница Б.А. Роль фосфора в современном земледелии // Плодородие. 2004. № 3(18). С. 13–16.
Веселкин Д.В., Рафикова О.С., Екшибаров Е.Д. Почва из зарослей инвазивного Acer negundo неблагоприятна для образования микоризы у аборигенных трав // Журн. общей биологии. 2019. Т. 80. № 3. С. 214–225.
Гамзиков Г.П. Азот в земледелии Западной Сибири. М.: Наука, 1981. 267 с.
ГОСТ 26107-84 ПОЧВЫ. Методы определения общего азота. М.: Издательство стандартов, 1984.
ГОСТ 26483-85 Почвы. Определение рН солевой вытяжки, обменной кислотности, обменных катионов, содержания нитратов, обменного аммония и подвижной серы методами ЦИНАО: Сб. ГОСТов. М.: Издательство стандартов, 1985.
ГОСТ 26951-86 Почвы. Определение нитратов ионометрическим методом. М.: Издательство стандартов, 1986.
ГОСТ 27821-88 Почвы. Определение суммы поглощенных оснований по методу Каппена. М.: Издательство стандартов, 1988.
ГОСТ 26213-91 Почвы. Методы определения органического вещества. М.: Издательство стандартов, 1992.
ГОСТ 26204-91 Почвы. Определение подвижных соединений фосфора и калия по методу Чирикова в модификации ЦИНАО. М.: Издательство стандартов, 1992.
ГОСТ Р 54650-2011 Почвы. Определение подвижных соединений фосфора и калия по методу Кирсанова в модификации ЦИНАО. М.: Стандартинформ, 2013.
Гродзинский А.М. Аллелопатия в жизни растений и их сообществ: основы химического взаимодействия растений. Киев: Наукова думка, 1965. 198 с.
Даденко Е.В., Денисова Т.В., Казеев К.Ш., Колесников С.И. Оценка применимости показателей ферментативной активности в биодиагностике и мониторинге почв // Поволжский экологический журн. 2013а. № 4. С. 385–393.
Даденко Е.В., Мясникова М.А., Чернокалова Е.В., Казеев К.Ш., Колесников С.И. Сезонная динамика ферментативной активности чернозема обыкновенного // Современные проблемы науки и образования. 2013б. № 6. С. 743.
Ерёменко Ю.А. Аллелопатическая активность инвазионных древесных видов // Российский журнал биологических инвазий. 2014. № 2. С. 33–39.
Звягинцев Д.Г. Методы почвенной микробиологии и биохимии. М.: Изд-во МГУ, 1991. 304 с.
Кавеленова Л.М., Вандышева Е.А., Розно С.А. Влияние древесных интродуцентов на некоторые показатели почвы в условиях дендрария ботанического сада // Бюллетень Самарская Лука. 2008. Т. 17. № 2. С. 407–415.
Казеев К.Ш., Колесников С.И., Вальков В.Ф. Биологическая диагностика и индикация почв: методология и методы исследований. Ростов-на-Дону: Изд-во Ростовский гос. университет, 2003. 216 с.
Классификация и диагностика почв СССР / Сост. В.В. Егоров, В.М. Фридланд, Е.Н. Иванова, Н.Н. Розов и др. М.: Колос, 1977. 221 с.
Коношина С.Н., Хилкова Н.Л., Прудникова Е.Г. Аллелопатическая активность листового опада древесных растений Орловской области // Ученые записки Орловского государственного университета. Серия: Естественные, технические и медицинские науки. 2014. № 3. С. 152–155.
Лозбякова А.И., Степанов М.В. Влияние химических веществ из опада листьев клена остролистного // Самарская Лука: проблемы региональной и глобальной экологии. 2019. Т. 28. № 1. С. 100–102.
Полевой определитель почв России / Под ред. Остриковой К.Т. М.: Почвенный институт им. В.В. Докучаева, 2008. 182 с.
Практикум по агрохимии / Под. ред. Минеева В.Г. М.: Изд-во МГУ, 2001. 689 с.
Сакбаева З.И. Влияние ферментативной активности фосфатаз на экологическое состояние сероземных почв предгорий Ферганы // Современные проблемы науки и образования. 2014. № 5. С. 615.
Самаров В.М. Почвы и климат Кузнецкой котловины: Учеб. пособие. Кемерово: ИИО Кемеровского ГСХИ, 2017. 79 с.
Титова В.И., Козлов А.В. Методы оценки функционирования микробоценоза почвы, участвующего в трансформации органического вещества: Научно-методическое пособие. Нижний Новгород: Нижегородская сельскохозяйственная академия, 2012. 64 с.
Трофимов С.С. Экология почв и почвенные ресурсы Кемеровской области. Новосибирск: Наука, 1975. 299 с.
Фаизова В.И. Ферментативная активность черноземов Центрального Предкавказья // Вестник АПК Ставрополья. 2014. № 3(15). С. 154–157.
Хазиев Ф.Х. Методы почвенной энзимологии. М.: Наука, 2005. 252 с.
Хазиев Ф.Х. Ретроспективы и проблемы почвенно-энзимологических исследований в Башкортостане // Вестник Академии наук Республики Башкортостан. 2014. Т. 19. № 3. С. 5–15.
Хазиев Ф.Х. Функциональная роль ферментов в почвенных процессах // Вестник Академии наук Республики Башкортостан. 2015. Т. 20. № 2(78). С. 14–24.
Хежева Ф.В., Улигова Т.С., Темботов Р.Х. Оценка ферментативной активности черноземов естественных биоценозов степной зоны и лесостепного пояса Центрального Кавказа // Известия Самарского НЦ РАН. 2010. Т. 12. № 1(4). С. 1075–1078.
Balemil T., Negisho K. Management of soil phosphorus and plant adaptation mechanisms to phosphorus stress for sustainable crop production: a review // Journal of Soil Science & Plant Nutrition. 2012. V. 12(3). P. 547–561. https://doi.org/10.4067/S0718-95162012005000015
Hu R., Wang X., Zhang Ya., Shi W., Chen N. Insight into the influence of sand-stabilizing shrubs on soil enzyme activity in a temperate desert // Catena. 2016. V. 137. P. 526–535. https://doi.org/10.1016/j.catena.2015.10.022
IUSS Working Group WRB. World Reference Base for SoilResources 2014. International soil classification system for namingsoils and creating legends for soil maps. World Soil Resources Reports. N 106. Rome: FAO. 2014. 181 p.
Li J., Tang X., Awasthi M.K., Wu F., Ha S., Ma J., Sun X., He Ch. Dynamics of soil microbial biomass and enzyme activities along a chronosequence of desertified land revegetation // Ecological Engineering. 2018. V. 111. P. 22–30. https://doi.org/10.1016/j.ecoleng.2017.11.006
Moghimian N., Hosseini S.M., Kooch Ya., Darki B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activities // Catena. 2017. V. 157. P. 407–414. https://doi.org/10.1016/j.catena.2017.06.003
Porté A.J., Lamarque L.J., Lortie C.J., Michalet R., Delzon S. Invasive Acer negundo outperforms native species in non-limiting resource environments due to its higher phenotypic plasticity // BMC Ecology. 2011. P. 11–28. https://doi.org/10.1186/1472-6785-11-28
Ratliff T.J., Fisk M.C. Phosphatase activity is related to N availability but not P availability across hardwood forests in the northeastern United States // Soil Biology & Biochemistry. 2016. V. 94. P. 61–69. https://doi.org/10.1016/j.soilbio.2015.11.009
Saccone P., Pagès J.P., Girel J., Brun J.J., Michalet R. Acer negundo invasion along a successional gradient: early direct facilitation by native pioneers and late indirect facilitation by conspecifics // New Phytologist. 2010. V. 187. P 831–842. https://doi.org/10.1111/j.1469-8137.2010.03289
Yu P., Tang X., Zhang A., Fan G., Liu Sh. Responses of soil specific enzyme activities to short-term land use conversions in a salt-affected region, northeastern China // Science of The Total Environment. 2019. V. 687. P. 939–945. https://doi.org/10.1016/j.scitotenv.2019.06.171
Дополнительные материалы отсутствуют.